Окаменелость. Кто это был?

Интересно, кто это был? Найдено в районе Кохила, Эстония.
Показать полностью
1
Интересно, кто это был? Найдено в районе Кохила, Эстония.

Бирманский янтарь - один из самых интересных источников окаменелостей. В нём удалось найти череп редкой амфибии-хамелеона, раковину аммонита, древнейших спаривающихся остракод, а также лапу птицы, похожей на руконожку.
Взял информацию из https://extinct-animals.fandom.com/ru/wiki/Вымершие_животные...

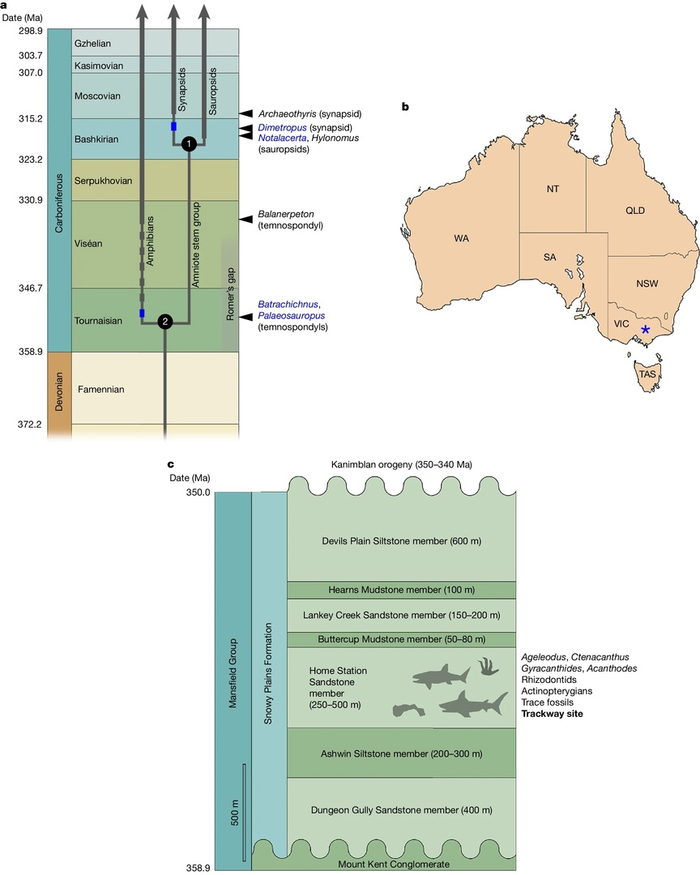
Австралия преподнесла палеонтологам сенсацию: окаменелые следы, найденные в формации Snowy Plains на севере штата Виктория. Эти отпечатки лап рептилеподобного существа, датированные ранним турнейским веком каменноугольного периода (~355 млн лет), отодвигают появление настоящих рептилий на целых 35–40 миллионов лет вглубь веков по сравнению с самыми древними из ранее известных остатков. Открытие указывает, что центром происхождения этих животных следует искать в южном суперконтиненте Гондвана, сердцем которого была древняя Австралия.
До этой находки общепринятая картина эволюции четвероногих позвоночных (тетрапод), вышедших на сушу из воды около 390 миллионов лет назад в девоне, выглядела так: древнейшие ископаемые скелеты или следы настоящих рептилий (амниот кроновой группы) относились к позднему карбону (около 320 млн лет). Новые следы из Австралии, сохранившие характерные царапины от когтей – ключевой признак ранних амниот, практически не встречающийся у других групп четвероногих, – решительно ломают эту схему. Плита со следами, обнаруженная палеонтологами-любителями в районе Мэнсфилда, запечатлела движение маленького, приземистого, похожего на варана существа.

"Уже через несколько секунд я заметил отчетливо сохранившиеся следы когтей. Сочетание этих царапин и формы ступней не оставляет сомнений – следы оставила примитивная рептилия", – комментирует один из исследователей. Это делает австралийскую плиту древнейшим в мире свидетельством существования рептилеподобных животных на суше.
Значение открытия выходит далеко за рамки простого удревнения происхождения рептилий. «Это переписывает целый раздел эволюционной истории», – отмечают ученые. Оно означает, что ключевые ветви эволюционного древа четвероногих – и сами настоящие четвероногие (кроновая группа), и настоящие рептилии (кроновая группа амниот) – должны были сформироваться гораздо раньше. Узел настоящих амниот не может быть значительно моложе границы девонского и каменноугольного периодов (около 359 млн лет назад), а истоки настоящих четвероногих уходят глубоко в девон, возможно, в ранний франский век. Это говорит о том, что эволюция четвероногих в девоне происходила гораздо быстрее, а палеонтологическая летопись этого критически важного периода – гораздо менее полная, чем считалось.
«Скелет может рассказать лишь ограниченное количество информации о том, что могло делать животное, но следовая цепочка фактически фиксирует его поведение и показывает, как оно двигалось», – объясняет ценность следов один из соавторов.


Череп был найден Майком Харрисоном и родом из местных юрских скал. Ему около 195 миллионов лет. Над этим черепом - копия первого черепа, найденного Марией и Джозефом Эннингами.
Это гигантский череп ихтиозавра темнодонтозавра (Temnodontosaurus), представленный на Lyme Regis Museum
Передние/верхние конечности, сверху вниз
Человек
Тирекс
Зухомимус

1. Эволюция и историческая биогеография
Аулакостефаниды – одни из самых интересных и в то же время мало изученных верхнеюрских аммонитов Русской платформы. Само это семейство и своей историей, и обликом некоторых своих представителей очень напоминает уже обсуждавшихся здесь космоцератид.
Аулакостефаниды – тоже настоящее суббореальное семейство и, как и остальные группы аммонитов, обитавших в экотонной зоне между двумя биогеографическими надобластями (Панбореальной и Тетис-Панталасса), существовали они недолго. Появившись в начале позднего оксфорда, аулакостефаниды полностью исчезли в конце кимериджского века. Но за полтора «века» существования в этом семействе успело появиться чуть меньше десятка родов (если считать макроконхов) и что-то около 50 видов, заметная часть которых служит видами-индексами зон, подзон и биогоризонтов суббореального и бореального оксфорда и кимериджа. Именно к этому семейству принадлежат все виды-индексы «стандартной» (английской) шкалы кимериджского яруса. Как и у большинства других позднеюрских аммонитов, у аулакостефанид был хорошо развит диморфизм. Микроконхи отличались по своей более грубой и сохраняющейся до конечной жилой камеры скульптуре, а также по более мелким размерам и присутствию ушек, а макроконхи, соответственно, в основном обладали более слабо скульптурованной раковиной с простым устьем. Диаметр раковины макроконхов мог превышать таковой микроконхов в 10 и более раз. Таксономический статус микро- и макроконхов Aulacostephanidae (опять же, как и у многих других перисфинктид) не совсем устоялся: их относили к разным родам, подродам или видам. Аулакостефаниды – первые из подотряда перисфинктин, у которых появилась лопастная линия «краспедитового» типа – с ростом раковины её усложнение шло в первую очередь за счет увеличения числа элементов, а не их сложности (как это происходит у обычных пери
сфинктид).

Фото Аммонит Microbiplites или Prorasenia, автор Alex

Фото Ringsteadia cuneata, автор Dimka
Микро- и макроконх аулакостефанид
Древнейшие аулакостефаниды (Decipia), как предполагал Дж. Кэлломон, а вслед за ним и другие аммонитчики, произошли от перисфинктид рода Liosphinctes. От своих предков Decipia отличались несколько меньшими размерами раковины, ранним появлением многоветвистых пучков ребер, меньшим углом, под которым лопастная линия подходила ко шву, а также большим количеством боковых лопастей (которые у них образовывались за счет деления внутренней боковой лопасти I, как и у остальных представителей подотряда Perisphinctina). Эти аммониты сразу же расселились по всей Бореально-Атлантической области (хотя Северной Атлантики ещё не существовало, название это широко используется) – от Восточной Гренландии до Русской платформы. Севернее и восточнее находки деципий не известны. Японские палеонтологи, правда, отнесли к Decipia несколько обломков аммонитов из Японии, но, скорее всего, эти экземпляры принадлежат к роду Dichotomosphinctes. К роду Decipia, судя по всему, принадлежит один «забытый» вид – Perisphinctes tenuifurcatus Nikitin, описанный в сборнике посмертно изданных трудов С.Н. Никитина и с тех пор никем никогда не упоминавшийся (Никитин, 1916, табл. II, фиг. 15-16). Никитин, правда, указывал этот вид из «кордатовых слоев», но эта привязка, видимо, ошибочна. Другие деципии стали известны у нас совсем недавно – статья с их описанием вышла только в прошлом году (Glowniak et al., 2010).
Потомки деципий Rinsteadia более далеко распространились на юг, их находки встречаются вместе с субтетическими аммонитами в Польше, Южной Германии и Франции; присутствуют они также в разрезах Англии, Шотландии и, конечно, Русской платформы. А вот в Гренландии рингстедии пока не найдены. Рингстедии были весьма разнообразными, их слабо скульптированные раковины были от полуэволютных до почти почти совершенно инволютных, а лопастная линия приобрела типичный «краспедитовый» облик. Если микроконхи деципий известны плохо, то с микроконхами рингстедий дело обстоит совершенно иначе. Это – небольшие хорошо скульптированные аммониты, относящиеся к роду Microbiplites. На Русской платформе рингстедии не слишком хорошо известны. Хотя их первые описания относятся к XIX веку – именно тогда Г. Траутшольд описал Ammonites cuneatus - в дальнейшем их описывали и/или изображали всего несколько раз. Микроконхи не изображались до самого последнего времени (первые изображены здесь – Glowniak et al., 2010, вместе с макроконхами), макроконхов изобразили Давиташвили (1926), Иловайский и Флоренский (1941 – под названием Ammonia kurmani). Не совсем понятно, были ли рингстедии на Приполярном Урале, откуда они изображались М.С. Месежниковым – это достаточно эволютные формы, которые могут относиться и к Pictonia. В любом случае, в Западной Сибири и восточнее этих аммонитов нет.

Фото Обломок крупной Ringsteadia?, автор Dimka
С границей оксфорда и кимериджа у аулакостефанид связаны важные изменения. Сама эта граница проводится по смене Ringsteadia на Pictonia, Vineta и Pomerania у макроконхов и Microbiplites на Prorasenia (=Desmosphinctes) у микроконхов. Столько родовых названий для аулакостефанид из основания кимериджа предложено не случайно – уж больно они разнообразные. Pictonia – почти эволютные, с хорошо скульптированными внутренними и гладкими внешними оборотами. Vineta, чьи раковины достигали в диаметре более полуметра - полуинволютные, с гладкой раковиной (только на ранних оборотах у этих аммонитов видны ребра). Pomerania были близки к Pictonia, но отличались появлением грубых одиночных ребер-складок на внешних оборотах. Все эти аммониты, как и оксфордские представители семейства, до последнего времени оставались у нас почти неизвестными. Пока известно только два района на Русской платформе, где пограничный интервал оксфордского и кимериджского ярусов представлен достаточно полно – это бассейн р. Унжи (Макарьев, Михаленино) и Соль-Илецкий район Оренбургской области (Ханская гора). Судя по присутствию в Москве и окрестностях Desmosphinctes, этот интервал должен присутствовать и здесь, но, насколько я могу понять, в разрезах он сейчас не обнажается. К началу кимериджа приурочено расселение аулакостефанид как на север и восток, так и на юг. В Арктике, вплоть до Таймыра и Хатангской впадины распространяются своеобразные Pictonia из группы P. involuta Mesezhnikov. В субсредиземноморских разрезах в это же время появляются «средиземноморские рингстедии», недавно выделенные А. Вежбовским в отдельный род Vielunia.

Фото Vineta cf. jaekeli Dohm, автор Dimka
В начале фазы Cymodoce всё это разнообразие вновь свелось к одному единственному роду Rasenia. Макроконхи разений, у которых появились боковые бугорки, удивительно напоминают макроконхов нижнекелловейских космоцератид Kepplerites (Gowericeras), а вот микроконхи у них заметно различаются. Как и пиктонии, разении были широко распространены в Северном полушарии. Достигли они и Арктических островов – есть разении на Шпицбергене и на Земле Франца-Иосифа. Кстати, у микроконхов аулакостефанид бугорки в месте ветвления ребер появились ещё в самом начале кимериджа. Многие исследователи по этой причине считали Prorasenia предками Rasenia.
В конце раннего-начале позднего кимериджа разнообразие аулакостефанид вновь увеличилось. В это время и у микро- и у макроконхов появляется перерыв ребер на вентральной стороне. Сначала, видимо, такой перерыв появляется у Zonovia, которые в остальном очень похожи на разений. Потом – у эволютных, грубо скульптированных Zenostephanus и у первых, в основном слабо скульптированных Aulacostephanus (Aulacostephanoides [M], Aulacostephanites [m]). Эти аммониты несколько отличались по своему географическому распространению. Зоновии в основном характерны для довольно узкой полосы, охватывающей северную часть Суббореальной области и южную – Арктической. Аулакостефанусы были распространены более широко. Они встречались по всей Суббореальной области, заходили на север Субтетической области, а местами достигали и Арктики (Хатангской впадины). Zenostephanus [M] – Xenostephanoides [m] встречались в Суббореальной области, но наиболее широко расселились в Арктике. Это – единственные верхнекимериджские аулакостефаниды, известные на Шпицбергене и на Земле Франца-Иосифа. И это – единственные аулакостефаниды, которые встречаются в Северной Америке (на севере Британской Колумбии, в Канаде), хотя они там исключительно редки: за последние 50 лет их было найдено всего несколько экземпляров. В самом начале позднего кимериджа существовали также «южные» разении (Eurasenia), распространенные в как в Суббореальной (Бореально-Атлантической), так и в Субсредиземноморской области. У этих аммонитов (порой достигавших весьма немалых размеров – до 60 см) перерыва ребер на вентральной стороне не было. Интересно, что у нижнекимериджских микроконхов (Prorasenia) ослабление ребристости на вентральной стороне есть, но оно имеет иную природу, чем у верхнекимериджских аулакостефанид, образуясь за счет утолщение ребер при переходе на вентральную сторону (Месежников, Алексеев, 1974)

Фото Rasenia с Унжи, автор Alex

Фото Zenostephanus sachsi (Mesezhnikov), автор MHorn

Фото Zenostephanus, автор Dimka
В начале фазы Eudoxus верхнего кимериджа появились достаточно грубо скульптированные аулакостефанусы, обычно рассматриваемые в составе подрода Pararasenia, но в действительности являющиеся гомеоморфами Центральноамериканских параразений (см. ниже). Их нужно относить к подроду Aulacostephanus (Sarygulia) [M] (Khudyaev in Sasonov, 1960).
Скорее всего, эти аммониты являлись потомками подрода Aulacostephanoides. Чуть позже появились настоящие Aulacostephanus (Aulacostephanus) [M] – полуинволютные, часто очень крупные и обычно не слишком сильно скульптированные аммониты, просуществовавшие до конца кимериджского века. Микроконхи аулакостефанусов из зон Eudoxus и Autissiodorensis объединятся в подрод Aulacostephanus (Aulacostephanoceras).
В самом конце кимериджа аулакостефаниды вымерли. Скорее всего, это событие произошло одновременно в Суббореальной и Субтетической областях, немного раньше аулакостефаниды исчезли в Арктике (в Хатангской впадине их уже нет в верхней зоне кимериджа, а в Восточной Гренландии – в верхней подзоне).

Фото Aulacostephanus yo (d9;Orbygny), автор MHorn

Фото Устье Aulacostephanus, автор Kach
2. Почти аулакостефаниды
Кроме настоящих аулакостефанид в кимериджском веке существовали также другие аммониты, очень на них похожие. В первую очередь это Pararasenia – аммониты, описанные из слоев с Idoceras Мексики К. Бурхардтом как Aulacostephanus zakatekanus. Потом Л. Спэт (1925) сделал этот вид типовым для рода Pararasenia. Он указал, что эти аммониты очень похожи на Aulacostephanus, но у них отсутствует перерыв скульптуры на вентральной стороне, чем они напоминают Rasenia. Спэт предполагал, что этот род связывает Aulacostephanus (для которого он в этой работе предложил типовой вид A. eudoxus, d'Orbigny) и Rasenia. Кроме мексиканского вида, к параразениям Спэт отнес также европейские виды A. pseudomutabilis (de Loriol) и A. circumplicatum (Quenstedt). Однако мексиканские аммониты обладают несколько отличающейся лопастной линией и меньшими размерами по сравнению с европейскими аммонитами, а также четко изолированным ареалом. Скорее всего, они не относятся к аулакостефанидам. В кимеридже Мексике встречаются и другие очень похожие на аулакостефанид аммониты. Это известный по единственной находке аммонит, описанный А. Канту-Чапой как Pictonia (Colladites) с гладкой жилой камерой и покрытыми одиночными ребрами внутренними оборотами. Скорее всего, он близок к идоцератинам – подсемейству, широко распространенному в кимеридже Мексики. Ещё в верхнем кимеридже Мексики и Восточной Африки встречаются аммониты, очень похожие на Craspedites или небольших Ringsteadia – Procraspedites. Как было установлено Ф. Олоризом с соавторами (Oloriz et al., 1998) по присутствию характерных пережимов на внутренних оборотах эти аммониты должны относиться к семейству атаксиоцератид.
3. Номенклатурные сложности
Многим кимериджским аулакостефанидам не повезло с типовыми видами. Так, вид Ammonites cymodoce d’Orbigny стал типовым видом и Rasenia и Pictonia. А. д'Орбиньи под названием Ammonites cymodoce изобразил различных аммонитов (хотя, видимо, и принадлежащих одному виду), и в качестве типового для обоих родов был выбран экземпляр, изображенный на табл.202, фиг.3-4. В. Аркелл сначала написал статью, где среди прочего обрисовал всю непростую ситуацию (Arkell, 1935), а затем направил предложения в Международную комиссию по зоологической номенклатуре (МКЗН) – высший орган, принимающий решения в спорных случаях. В результате в 1951 году МКЗН приняла решение, зафиксировавшее типовые виды рассматриваемых родов (соответственно, Pictonia baylei и Rasenia involuta), а название cymodoce d'Orbigny, 1850 поместившее в список отклоненных названий, которые не должны в дальнейшем использоваться (Hemming, 1951). Позднее, после обсуждения проблемы с В. Дж. Аркеллом, было решено ещё раз поставить вопрос на голосование в МКЗН: по мнению Аркелла название Cymodoce, которое к этому времени уже давно стало широко используемым названием для зоны, лучше всё же было сохранить в качестве используемого, с типовым экземпляром, выбранным из коллекции д’Орбиньи в 1904 г. В результате члены комиссии большинством голосов восстановили в правах вид Cymodoce (Hemming, 1956). Ну а вместо R. involuta Spath 1935 в дальнейшем было предложено название Rasenia anglica Geyer, 1961 (т.к. вид Ammonites involutus Quenstedt угодил в род Rasenia, и название, предложенное Спэтом стало его младшим омонимом). В. Аркелл с Дж. Кэлломоном (Arkell, Callomon, 1961) считали, что это микроконх, но впоследствие Т. Биркелунд (Birkelund et al., 1978) показала, что это может быть и молодой макроконх.
Столь же непростая ситуация была и с типовым видом рода Aulacostephanus. Оно было первоначально опубликовано Торнквистом (Tornquist, 1896) без диагноза, со ссылкой на то, что название предложено Сутнером и Помпецким. Единственным видом, отнесенным к данному роду, стал Ammonites mutabulis Sowerby – однако, судя по всему, Торнквист имел в виду аммонита, изображенного под этим названием д’Орбиньи (и который в свою очередь был отнесен де Лориолем к новому виду pseudomutabilis); по правилам МКЗН этот вид и должен являться типовым для Aulacostephanus, хотя Торнквист про это ничего и не писал. Тем не менее, в 1925м году типовой вид для рода Aulacostephanus был предложен сразу двумя выдающимися аммонитчиками – Леонардом Спэтом (Spath, 1925) и Отто Шиндевольфом (Schindewolf, 1925). Спэт предложи в качестве такового A. eudoxus, а Шинденвольф – A. phorcus. Однако оба предложенных названия не были в числе видов, изначально отнесенных к роду Aulacostephanus Торнквистом, и такое обозначение типового вида не является валидным. Позднее В.Дж. Аркелл (Arkell, 1935) предложил считать типовым видом рода Aulacostephanus A. pseudomutabilis de Loriol, и это было закреплено специальным решением МКЗН в 1951 году.
Название Sarygylia было опубликовано И. Худяевым (1932) только в виде подписи к одному из рисунков, тогда как в тексте тот же самый вид был отнесен к Aulacostephanus. Такое выделение нового рода без диагноза и ссылки на типовой вид недействительно. Позднее единственным, кто использовал предложенное Худяевым название, был Н.Т. Сазонов (1960), который привел диагноз данного рода и перечислил другие относящиеся к нему виды. Сазоновым в той же работе был установлен новый род Tobolia, который оказался младшим омонимом установленного ранее рода фораминифер и заодно синонимом Aulacostephanoides.
Род Xenostephanus, установленный В. Аркеллом и Дж. Кэлломоном в 1963 году, оказался омонимом рода млекопитающих, установленных годом раньше, и для аммонитов в 2010м году было предложено новое название Zenostephanus.
Не очень понятной получилась ситуация с Ammonites uralensis d’Orbigny in Murchison et al.,1845 – типовым видом рода Zonovia. Орбиньи изобразил под этим названием двух аммонитов. Позднее Р. Зальфельд (Salfeld, 1913) посчитал, что к данному виду относится только более крупный экземпляр (Тове Биркелунд указывала, что Зальфельд таким образом выбрал лектотип – но в действительности в статье Зальфельда лишь было указано, какого из изображенных Орбиньи аммонитов он относит к данному виду). Вскоре Р. Дювие (Douville, 1911) изобразил двух аммонитов из коллекции Орбиньи, один из которых, происходящий из окрестности Кинешмы, ранее не изображался. Сам Дювийе решил, что это – нижнекелловейские Kepplerites. Более крупный из изображенных Дювийе аммонитов был впоследствии выбран в качестве лектотипа Б. Циглером (Ziegler, 1962). Поскольку более крупный аммонит с Урала, изображенный Орбиньи, был утерян, М.С. Месежников (1984) после консультаций с Дж. Кэлломон предложил одного из аммонитов из своей коллекции в качестве неотипа вида Z. uralensis – это, однако, не является валидным обозначением, поскольку лектотип был выбран Циглером корректно.

Моя небольшая коллекция юрских аммонитов Подмосковья.
Этой небольшой публикацией я хочу открыть серию статей, посвященных различным семействам аммонитов, обитавших на территории Центральной России.
Как известно, аммониты, жители палеозойских и мезозойских морей, были очень разнообразны. Каких только раковин у них не было – были и дисковидные, и шаровидные, и похожие на колесо (иногда не только по форме, но и по размеру). Причина такого разнообразия в очень быстрой эволюции аммонитов. Они сменяли друг друга, приспосабливаясь к изменчивым условиям окружающей среды. Это только нам, «со стороны» кажется, что тот же юрский период был временем равномерно теплого климата и полного спокойствия в биосфере – на самом деле и в юре, и в мелу формировались горы, наступали или наоборот отступали и становились мелкими моря, температура воды в них падала или повышалась. Конечно, не было таких перепадов как сейчас, не было оледенений, но все равно было множество факторов, которые не давали аммонитам жить спокойно и заставляли постоянно приспосабливаться и изменяться.

Фото Михайловцемент. Стратиграфия в картинках., автор Dimka
На этой схеме, которую нарисовал уважаемый Dimka, хорошо видно разнообразие аммонитов в одном единственном карьере.
Иногда, в разговоре об аммонитах от начинающих любителей палеонтологии можно услышать сожаление – «эх, не вымерли бы аммониты, ловили бы мы сейчас в морях виргатитов и космоцерасов!». Увы, даже если бы аммониты не вымерли, ни аммонитов рода Kosmoceras, ни рода Virgatites мы бы не встретили. Они и друг с другом-то не встречались никогда, их разделяли несколько миллионов лет, оба эти рода исчезли навсегда, когда до окончания мелового периода и заката «эры аммонитов» оставалось больше 80-ти миллионов лет. Аммониты появлялись и вымирали постоянно. То же самое можно сказать и о других животных, хотя некоторые группы – например мезозойские наутилусы двустворчатые моллюски эволюционировали значительно медленнее.
Если бы аммониты не вымерли, то даже сложно себе представить, какими бы были их потомки через 65 миллионов лет эволюции. Но как они (вернее, их раковины) выглядели в прошедшие эпохи, мы знаем и о них, об их эволюции я и хочу рассказать в этом цикле статей.
Это будет серия небольших статей, посвященных разным семействам аммонитов. Я буду писать только о тех аммонитах, с которыми знаком лично, поэтому начну рассказ с юрских семейств, а потом, может быть, дойду и до мела.
Героями моих статей будут в основном аммониты, обитавшие в морях Русской платформы, точнее, в Центральной России и Поволжье, аммониты бореальные и суббореальные. Здесь необходимо пояснить, что море, занимавшее в юрском и меловом периодах большую часть современной России, было относительно мелководным и относительно холодным. На севере оно соединялось с Бореальным (полярным) океаном, на юге и востоке – с заливами океана Тетис.
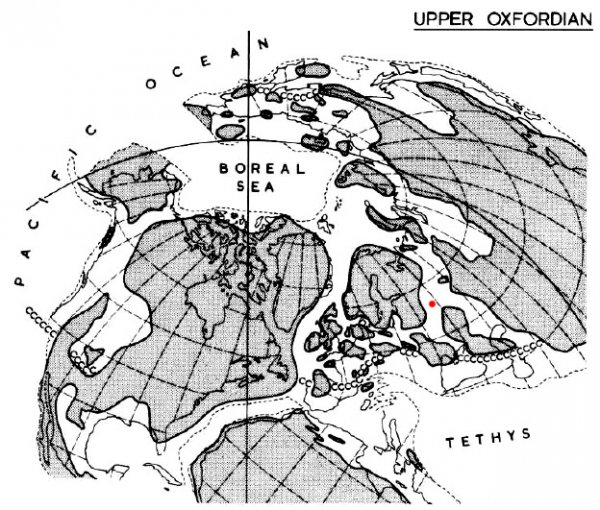
Северное полушарие в оксфордском веке юрского периода. Иллюстрация из книги Callomon J.H. (1985) "The evolution of the Jurassic ammonite family Cardioceratidae". Красной точкой отмечена Москва. Рядом букв "с" - южная граница распространения аммонитов семейства Cardioceratidae в то время.
Конечно, это описание – довольно приблизительная реконструкция, так как ситуация постоянно менялась, например в волжском веке практически пересохли проливы, соединяющие наше море с Тетисом. Но главное тут вот в чем – в океане Тетис были значительно большие глубины и широкие просторы. Там жили аммониты, которые не встречались в мелководных морях Центральной России. В первую очередь это представители подотрядов Phylloceratina и Lytoceratina. Именно поэтому их много в Крыму и на Кавказе (они располагались у края океана Тетис), но совсем нет в Подмосковье или на Приполярном Урале. Бореальные аммониты могли довольно далеко заходить на юг, практически до самого современного Средиземноморья, за которым уже начинался открытый океан. В обратном направлении – на север, аммониты тоже расселялись, например представители тетического семейства Oppeliidae. Но все эти мигранты с юга относились к подотряду Ammonitida, в то время как литоцератины и филлоцератины не хотели покидать материковые склоны океана, мелководные просторы Центральной России их не привлекали.
Поэтому мои истории пока будут посвящены только аммонитидам. Выбор в качестве главных объектов именно семейств аммонитов, а не подсемейств или родов, связан с тем, что чем меньшего уровня таксон мы рассматриваем, тем меньше получается однозначности и ясности. Если на уровне семейств аммонитов у специалистов не так уж много разногласий, то в вопросах, касающихся родов и видов, этих самых разногласий не просто много, а очень много. Кроме того, именно семейства аммонитов можно считать достоверными эволюционными рядами – в их выделении гораздо меньше субъективности, чем в выделении родов и видов, аммониты, отнесенные к тому или иному семейству действительно обладают определенными наборами хорошо заметных признаков и по изменениям этих признаков можно прослеживать ход эволюции аммонитов.

Моё фото. Аммонит Ammonitoceras. Ранний мел: Северный Кавказ. Палеонтологический музей. им Ю.А Орлова

Моё фото. Аммонит Ammonitoceras. Ранний мел: Северный Кавказ. Палеонтологический музей. им Ю.А Орлова

Моё фото. Аммонит Speetoniceras Versicolor. Ранний мел: Ульяновская область. Палеонтологический музей. им Ю.А Орлова

Ярославская область, волжский век. 149 млн. лет назад. https://www.ammonit.ru/foto/54662.htm

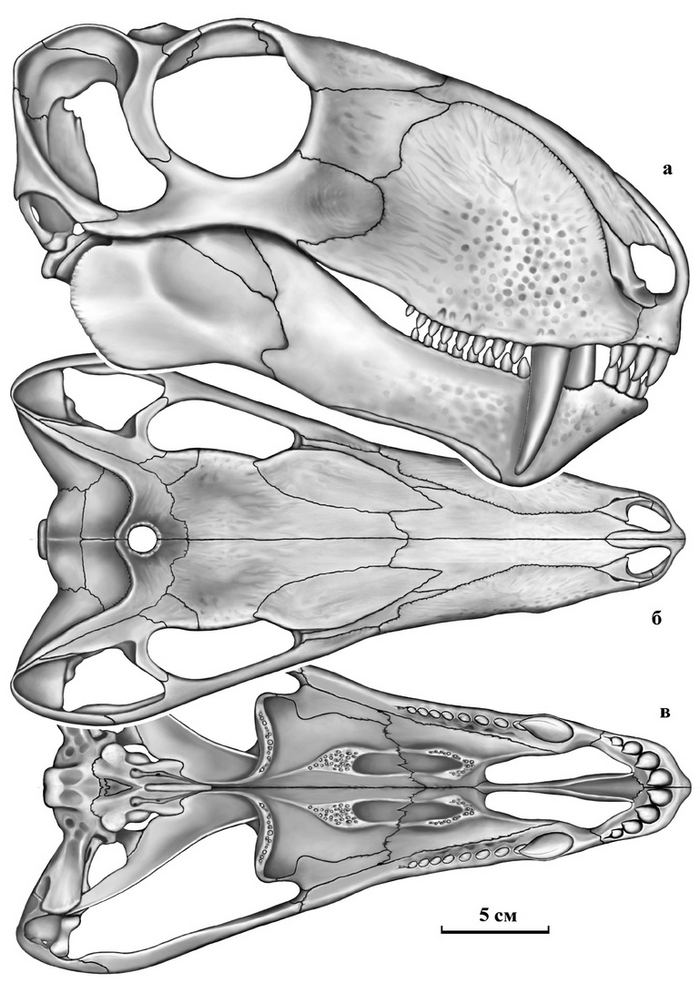
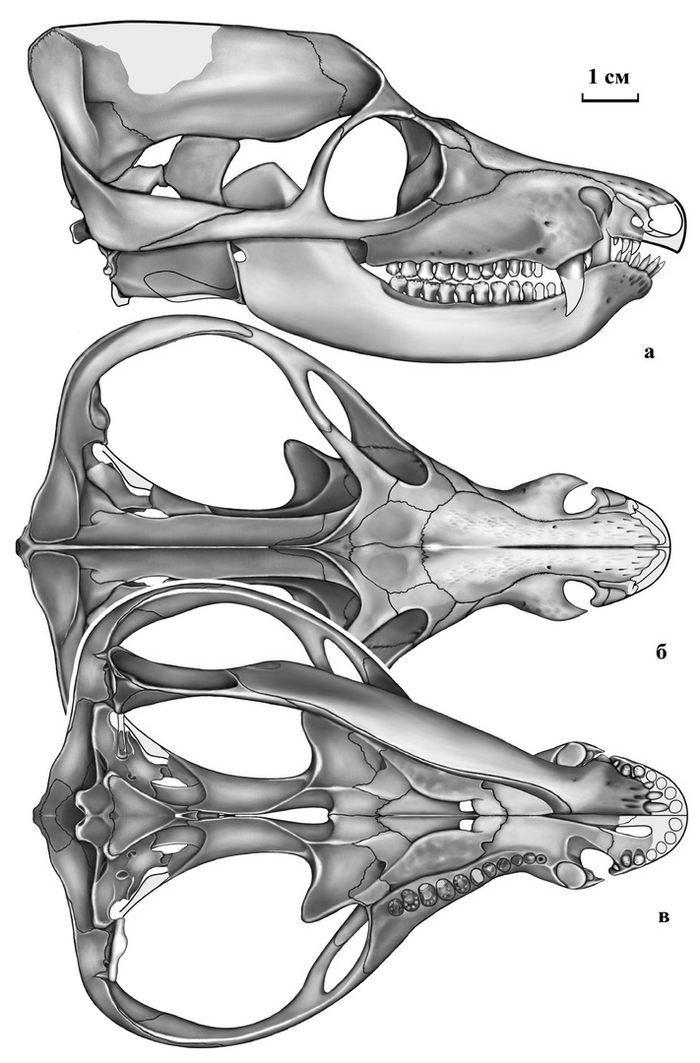
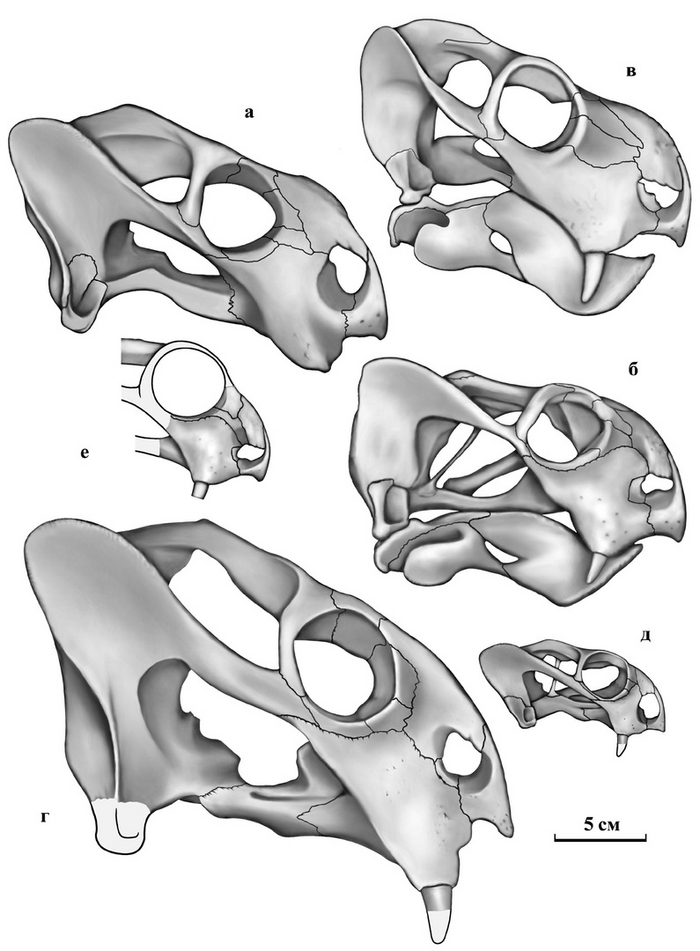
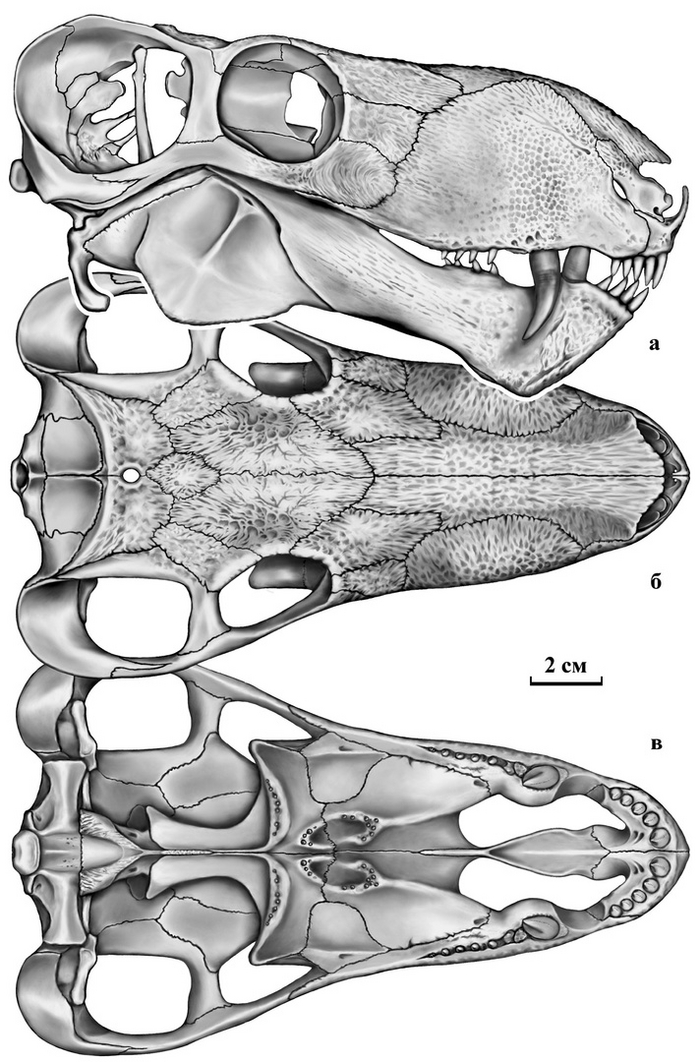
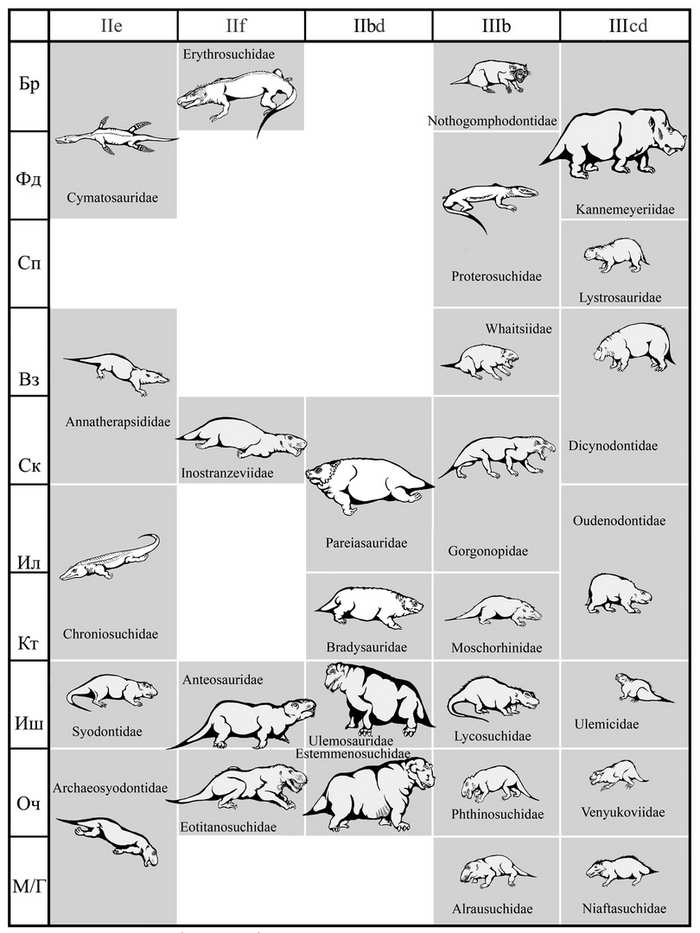
Тероморфы Восточной Европы / М.Ф. Ивахненко (Тр. ПИН РАН. Т. 301). – М.: РАН. – 2024. – 466 с. ISBN 978-5-907645-35-6
От научного редактора монографии, палеонтолога В.К. Голубева:
"10 июля 2015 г. ушел из жизни выдающийся палеогерпетолог, крупнейший специалист по позднепалеозойским и раннемезозойским тетраподам Михаил Феодосьевич Ивахненко. Более 45 лет он был сотрудником Палеонтологического института, где занимался изучением пермских и триасовых амфибий и рептилий Восточной Европы и северной Азии.
С 1990-х гг. главным объектом его исследований были зверообразные,
или тероморфы. По этой теме он опубликовал 7 монографий и множество статей. К сожалению, почти все важные работы вышли только на английском языке, и поэтому, как считал Михаил Феодосьевич, труднодоступны для большинства русскоязычных ученых и палеонтологов-любителей.
К тому же они посвящены морфологии, систематике и палеоэкологии лишь отдельных групп Theromorpha. По мнению М.Ф. Ивахненко, все эти разрозненные данные необходимо было сопоставить, обобщить, свести в единую схему и опубликовать в виде монографии на русском языке. К данной работе, являющейся, по сути, итогом его многолетних исследований уникальных коллекций восточноевропейских тероморфов, он приступил в последние годы жизни. Монография была почти готова, оставалось только в «Заключение» подвести итоги изучения восточноевропейских тероморфов, когда из-за осложнений от неудачно проведенной операции Михаил Феодосьевич попал в больницу. Там он продолжил работу: начал писать «Заключение», но скоропостижная смерть не позволила довести дело до завершения.
В настоящем томе Трудов Палеонтологического института РАН посмертно публикуется монография М.Ф. Ивахненко по восточноевропейским тероморфам. При подготовке рукописи к публикации были исправлены очевидные опечатки, ошибки в коллекционных номерах и в стратиграфической терминологии. Научное редактирование текста было сведено к минимуму, чтобы непреднамеренно не исказить смысл, который автор хотел донести до читателя. По этой же причине «Заключение» было оставлено практически в незавершенном виде: точно не известно, на чем еще хотел акцентировать в нем внимание М.Ф. Ивахненко. В процессе редактирования выяснилось, что отсутствует рис. 82 (вместо него был продублирован рис. 81). Этот рисунок был частично реконструирован в файлах на домашних и рабочем компьютерах М.Ф. Ивахненко (все электронные научные материалы М.Ф. Ивахненко, хранившиеся на домашних компьютерах, были любезно предоставлены его вдовой, Любовью Фоминичной Ивахненко). К сожалению, сам рисунок обнаружить не удалось.
Возможно, он так и не был сделан, или вместе с исходными графическими материалами был по ошибке удален самим автором.
В конце монографии добавлено приложение со списком научных трудов М.Ф. Ивахненко, списком новых таксонов, описанных М.Ф. Ивахненко лично и с соавторами, и списком родов и видов, названных в честь М.Ф. Ивахненко. Приложение составлено А.Е. Нелиховым и В.К. Голубевым."