Немного эротики

Показать полностью
1
ГДЕ ПЕРЬЯ, БЛЭТ!!!
Палеонтологи, конеш, еще не нашли останки Ти-Рекса, однозначно позволившие бы идентифицировать наличие у него этой формы "внешнего бета-каротинового слоя кожи рептилий". Однако останки азиатских родственников показывают, что тираннозавры Северной Америки с большей вероятностью тоже были пернатыми.




В очередной раз поднимаем тему оперённости динозавров. На этот раз обсудим три интересных и свежих находки за последние три года. Помимо разбора этих интересных представителей ужасных ящеров мы докинем пару доказательств в тероподовую теорию происхождения птиц.

Начнём мы с самого интересного представителя юрского пернатого - странное крыло или Yi Qi. Этот малыш пытался летать задолго до появления первых птиц, только с немного другой механикой. ~160 млн лет назад на востоке Китая жил динозавр по имени И и его полёт был похож на птерозавровый или на полёт летучей мыши. Весил кроха-манираптор менее 500 гр. На передних конечностях была натянута кожаная перепонка. Мягкие ткани перепонки сохранились в хорошем состояние, поэтому сомневаться в ней не приходится. Только вот, судя по строению передних конечностей, полёт был пассивным.
Yi Qi был найден в далеком 2007 году, но тщательные исследования были проведены в 2015. Тогда же малыш и получил своё имя.

Останки Yi Qi дошли до нас в превосходном состояние

Странное крыло хоть и был похож на летучую мышь, но имел совершенно иное строение крыла
Дракон из Чжэньюань был намного моложе И, но обладал уже птичьем крылом и маховыми перьями. Описанный в 2015 году род Zhenyuanlong из семейства дромеозавридов был найден в провинции Ляонин. Это первый крупный вид динозавров с длинными маховыми крыльями (позже в 2015 года был открыт дакотараптор с такими же маховыми перьями, который превосходил дракона в размерах почти в 10 раз). Наш герой впервые поднял вопрос о практической пользе маховых перьев у столь крупных особей. При двухметровой длине Zhenyuanlong весил более двух десятков кг. Ни планировать, ни летать он явно не мог. Подобное строение передних конечностей косвенно подтверждают тероподовую теорию, которая говорит, что сначала динозавры развивали крылья для быстрого бега, а с помощью длинного оперения поддерживали равновесие. Скорее всего, данный род динозавров имел общего предка с родственными ему микрорапторами.
Дракон из Чжэньюань, как и Странное крыло, прекрасно сохранился и задал очень много вопросов учёным о вымерших более 100 млн лет назад рептилиях и их образе жизни.

У останков Zhenyuanlong suni отчётливо сохранились следы маховых перьев

Дракон из Чжэньюань жил 125 млн лет назад
Кулиндадромеус забайкальский смог удивить русских палеонтологов. Ведь это самый древний представитель ужасных ящеров в России (жил ~150 млн лет назад) и подтверждение оперения у птицетазовых! Удивительная находка была обнаружена ещё в 2010 году, однако описали её только 2014. После многочисленных раскопок, где были найдены 6 черепов и сотни различных фрагментов динозавра, был составлен подробный облик Kulindadromeus zabaikalicus. Это небольшое животное из семейства Neornithischia с трудом превышал длину в 1,5 метра. Перьевой покров имел отличную от манирапторов структуру. Мелкий и густой слой протоперьев служил для сохранения тепла во время перепадов температуры, что свойственно для теплокровных животных. При этом, птицетазовый пушистик имел и чешуйки.

Один из фрагментов окаменелостей забайкальского динозавра со следами протоперьев

Внешний облик животного. Кулиндадромеус совмещал перьевой и чешуйчатый покровы


В нашей лиге было уже множество постов о динозаврах и везде есть арты с оперёнными ящерами. Но стабильно найдётся один-два человека, которые в такие реконструкции не верят. Так вот, господа неверующие, этот пост специально для вас. Сегодня я вам объясню, почему теория оперённости и теплокровности ужасных ящеров справедлива не только для маленьких рапторов, но и для всех тероподов.

Для начала, следы перьев.
Справедливости ради, это самые слабые доказательства. Но все гипотезы строились именно вокруг следов перьев, а позже всё подтвердилось более увесистыми доказательствами. Вокруг скелетов отпечатались чёткие контуры и/или ауры. Мягкие ткани успели разложиться, но перьевой покров жёстче и дольше разлагается, поэтому он успел оставить свой след на камне. В случае с микрораптором, следы были очень отчётливы и наблюдались не просто пушистые перья, а длинные контурные. Сомневаться не приходилось, это очевидно перья. Но это всего лишь маленький дромеозаврид, близкий птице. А вот в 2012 году свершилось новое открытие, которое ни чуть не уступало по значимости первому открытию оперённости. В китайской провинции Ляонин, родине огромному количеству оперённых останков, найден древний родственник тираннозавров в идеальном состояние, ютиран хуали. Останки сохранили отчётливый контур оперения по всему телу. Но такое уже находили десятки раз, в чём особенность, в чём прорыв для палеозоологии? Во-первых, ютиран жил 125 млн лет назад, задолго до появления знаменитых тираннозавридов. Получается, что оперённость хищникам эволюция выдала до становления динозавров на птичий путь. Во-вторых, ютиран никак родственно не связан с микрорапторами и им подобными. Они давно разделились. Даже тираннозавру ютиран так себе родственник. Их объединяет лишь надсемейство тираннозавроидов. В-третьих, ютиран - гигантская машина убийств длинной 9 метров и весом 1,5 тонны. Густой пуховой покров был свойственен не только малышам, но и гигантам, в этом больше не приходится сомневаться.

Отчётливые следы перьев у микрораптора. Сложно такие следы интерпретировать как не перьевой покров.

Такие следы были и у меловой птицы джехолорниса. Разве можно сомневаться, что это следы перьев?

Внешний вид динозавров с перьями. Чёрный - микрораптор, бело-коричневый - джехолорнис.

Один из камней, на котором отчётливо сохранились следы оперения ютирана.

Пернатый арт гиганта.
Но скептики не верят в следы перьев. Извращаясь над логикой, они приводят совсем дикие теории. Для вас, ребята, я приведу другие аргументы. Маховые бугорки.
Маховые бугорки на предплечьях свойственны, к сожалению, лишь дейнонихозаврам и птицам. Ведь они, бугорки, появились для крупных маховых перьев, которые не зачем отращивать крупным тероподам. В 2007 году были найдены очень важные, с точки зрения палеонтологии, останки монгольского велоцирапторы. Дело в том, что на предплечье малыша чётко сохранились маховые бугорки, свойственные всем современным птицам. Маховые бугорки - это места крепления маховых перьев на кости. После тщательного исследование останков и их сравнения со скелетами современных птиц сомнений не осталось. Велоцираптор обладал не просто пухом, а длинными маховыми перьями. Это ещё и небольшое доказательства эволюционного пути птиц от динозавров.
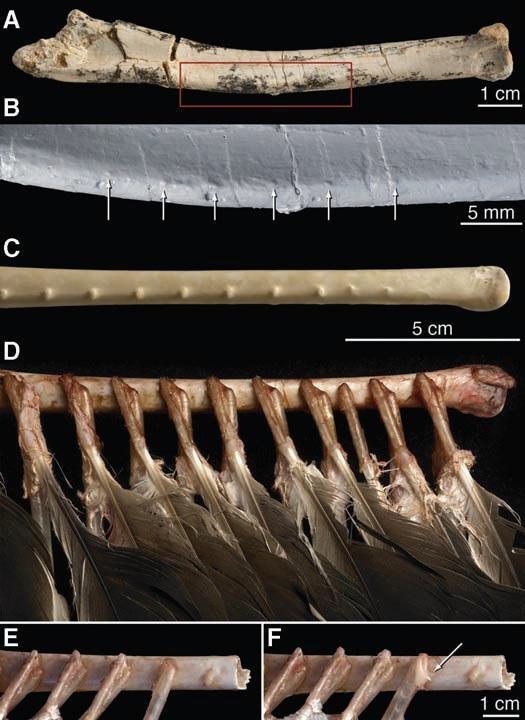
~(A) Дорсальный вид правой локтевой кости Velociraptor IGM 100/981.
~(B) Детальное изображение из красного поля в (A), со стрелками, показывающими шесть равномерно расположенных маховых бугорков перьев. В (B), использовался слепок IGM 100/981.
~(С) Дорсальный вид правой локтевой кости грифа-индейки (Cathartes).
~(D) Тот же вид Cathartes, как и в (С), но с рассечёнными мягкими тканями, для показа размещения вторичных маховых перьев и больших вторичных кроющих, относительно маховых бугорков.
~(E) Детальное изображение Cathartes с одним, полностью удалённым стержнем, для демонстрации махового бугорка.
~(F) Тот же вид, что и в (E), но с отведённым влево стержнем, для демонстрации расположения стержня, бугорка и фолликулярной связки. Фолликулярная связка обозначена стрелкой.

Мой любимый арт велоцирапторов.

Маховые бугорки сохранились и у дакотараптора. Это самый крупный дейнонихозавр, чьё оперение не ставится под сомнение. Вес дакотараптора близился к четырём центнерам.

Внешний вид дромеозаврида-гиганта.
Важнейшее доказательство оперения у тероподов было открыто в минувшем году. Сохранившиеся перья динозавра.
В 2016 году был найден законсервированный в янтаре хвост с перьями. Находка датировалась меловым периодом (около ~100 млн лет назад) и принадлежит очевидно не птице, ведь структура пера была слишком примитивна даже для меловых птиц. После долгих исследований был вынесен вердикт, хвост принадлежит очевидно кому-то из целурозавров (группа динозавров, объединяющая большинство тероподов). Сомнений не оставалось, следы перьев и маховые бугорки, очевидно, не спекуляции и фальсификации. Что ещё нужно неверующим, если уже найден пушистый хвост, подтверждающий на 100% оперённость динозавров?
В наши дни неизвестно ни одного примера, чтобы животные мира сего полностью лишались перьев естественным путём. Это уже не эволюция, это деградация. Но я согласен, что в исключительных случаях такое могло быть. Шерстистый мамонт и современный саванный слон доказывают, что млекопитающие могут лишаться меха. Современные безногие ящерицы доказывают, что порой рептилии могут отказаться и от ног. Но с перьями такого не было ни разу. Они имеют отличную от шерсти природу. Моё личное мнение, что некоторые динозавры избавлялись от перьевого покрова, частично или полностью, но оставались склонны к оперённости. Любые изменения в окружающей среде вынудили бы "лысых" ящеров моментально (по меркам эволюции) отрастить себе перья обратно.


Найденный хвост целурозавра.

Возможно так выглядел динозавр, который умудрился потерять свой хвост.
Оперённость, как доказательство теплокровности.
Немножко о том, что такое теплокровность и хладнокровность.
Пойкилотермия (ранее использовался термин холоднокровность) — эволюционная адаптация вида, при котором температура тела живого существа меняется в широких пределах в зависимости от температуры внешней среды.
Гомойотермиия (теплокровность) — способность живого существа сохранять постоянную температуру тела, независимо от температуры окружающей среды.
Дело в том, что при холоднокровности животное регулирует свою температуру за счёт тепла солнца или разогретых поверхностей, а охлаждается в тени или в воде. При наличии такого густого оперения - это просто невозможно. Пуховой слой динозавров изолировал их тело от внешних температурных факторов. Перья просто напросто не пускали солнечное тепло. С разогретой поверхностью всё то же самое. Тепло будет поглощаться внешним покровом, и не дойдёт до тела. В то же время, теплокровным животным густой пух как раз свойственен. Ведь они поддерживают тепло за счёт быстрого метаболизма, химического процесса внутри организма. Тепло у них вырабатывается внутри тела и его нужно там удержать. Этому будет способствовать внешний покров. В учёном мире уже нет спора: были ли динозавры теплокровны; сейчас у учёных спор: какие именно динозавры были теплокровны. Тероподы вне всяких сомнений были теплокровными и возможно могли жить и охотиться в снегах при минусовых температурах. Сейчас главный вопрос, когда именно динозавры получили столь полезную вещь, как теплокровность?


Оперённый хвост динозавра с примитивными перьями, застывший в
среднемеловом янтаре
Лида Син, Райан МакКеллар, Син Сюй, ... Куовэй Цен, Хао Жань, Филип Карри
КРАТКОЕ СОДЕРЖАНИЕ
За прошедшие с открытия пернатых динозавров два десятилетия спектр известного
оперения нептичьих теропод весьма расширился, и были подтверждены некоторые черты,
предсказанные моделями развития эволюции перьев. Однако, трёхмерная морфология
пера и эволюционные паттерны остаются трудными для интерпретации, ввиду
сплющивания в осадочных породах. Недавние открытия в меловом янтаре из Канады,
Франции, Японии, Ливана, Мьянмы, и США дают нам гораздо лучшую чёткость структурных
деталей, но таксономическое положение находок неясно, так как перья редко
ассоциированы с подходящим для идентификации скелетным материалом. Здесь мы
описываем оперённый хвост нептичьего теропода, сохранившийся в среднемеловом
(99м.л.н.) янтаре из Kachin State, Мьянма, со структурой перьев, прямо рассказывающей об
эволюционном пути их развития. Этот образец даёт возможность задокументировать
древние перья в непосредственной связи с предполагаемым ювенильным целурозавром,
сохранившим мелкие морфологические детали, в том числе пространственное положение
фолликул и перьев на теле, и особенности оперения по шкале микрометра. Многие перья
демонстрируют короткие, нежные стержни с чередующимися бородками и однообразной
серией соприкасающихся бородочек, что поддерживает гипотезу, что эти бородки уже
обладали ими, когда срослись, образовав стержень. Карбонизированные мягкие ткани под
перьями позволяют взглянуть на потенциал сохранности и историю инклюзии; обилие Fe2+
говорит об остатках гемоглобина и феррита, заключённых в хвосте. Новая находка
подчёркивает уникальный потенциал янтаря для сохранения останков и понимания
морфологии и эволюции покровных структур целурозавров.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сохранность
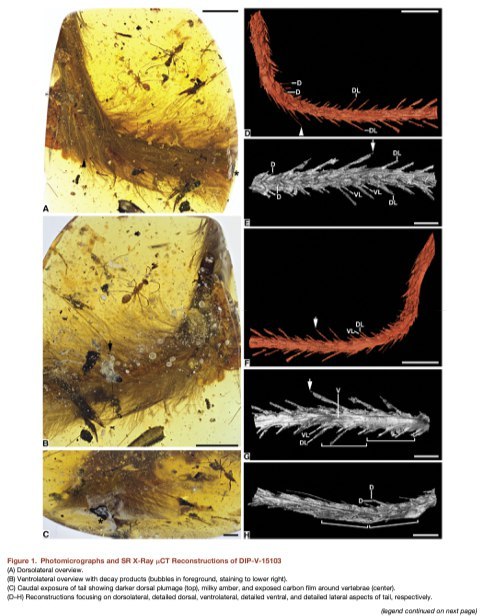
Хвост в образце DIP-V-15103 виден невооружённым глазом, как вытянутое и плавно
изогнутое образование (длина =36.73мм). Из него растёт густой перьевой покров,
закрывающий находящиеся под ним детали, так что для изучения скрытых особенностей
остеологии и мягких тканей было использовано синхротронное фазоконтрастное
рентгеновское микротомографическое сканирование (рис.1). Мягкие ткани — видимо,
мышцы, покровы, и кожа — спорадически видны под оперением, и держатся на костях
образом, напоминающим высыхание, обыкновенное для остатков позвоночных в янтаре.
Эти ткани в основном редуцированы до углеродной плёнки, и содержат только следы
своего исходного химического состава. На основании анализов, описанных в
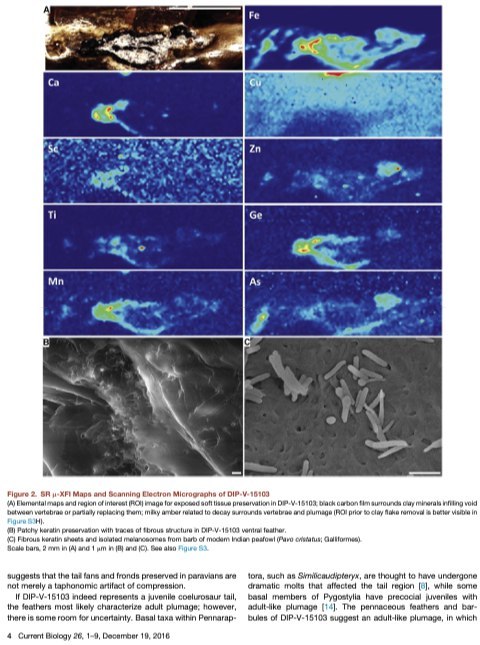
дополнительных материалах, SRm-XFI показывает, что в карбонизированных мягких тканях
есть железо, и оно присутствует как серия чётких линейных образований, соответствующих
открытому оперению (рис.2). Медь несколько более обильна в содержащем перья янтаре,
но этот сигнал криптичен, и не является чётким индикатором сохранившихся пигментов.
Такие элементы, как Ca, Sc, Zn, Ti, Ge, и Mn, по-видимому, связаны с минералами в глине,
заполнившей пустоты в янтаре. Мы вывели валентность железа в образце методом
сравнения со стандартной спектроскопией поглощения железной фольги, Fe2O3, Fe3O4, и
FeO. Наши вычисления указывают, что больше 80% железа в нём двухвалентно (Fe2+).
Аналогичные измерения были выполнены для сохранившихся сосудов в костях
Tyrannosaurus и Brachylophosaurus, и интерпретированы как говорящие о присутствии
гётита и биогенных оксидов железа, получающихся от распада гемоглобина. Наличие
большого количества Fe2+ в DIP-V-15103 предполагает, что гемоглобин или феррит в
останках остался пойман в инклюзии. Анализы с помощью сканирующего электронного
микроскопа показывают, что также там есть пирит (FeS2), но он не имеет существенного
вклада в распределение железа в образце (рис.S3). Тесный контакт между кожей и
окружающим янтарём, вместе с её мумифицированным внешним видом, при котором она
сморщилась вдоль поверхности позвонков, говорит об одном из двух сценариев. Либо
хозяин хвоста был мёртв и частично иссох перед заточением в смоле, либо он очень
быстро высох из-за взаимодействия со смолой. Высыхание на ранней стадии
подтверждает некоторое количество мутного янтаря, окружающее хвост (рис. 1C и S2), что
есть a особенность сохранения, связанная с продуктами разложения или влаги,
взаимодействующей со смолой. Однако, высыхание и последующее попадание в смолу
недостаточно для сохранения деталей клеток в мягких тканях. Судя по глине там, где кость
разрывает поверхность янтаря, скелетный материал, похоже, был обнажён после
полимеризации смолы. Кость частично растворена и заполнена глиной из окружающей
матрицы, как полости тела насекомого в этом отложении (рис. S2A). Наличие Fe2+ в
карбонизированных остатках намекает, что органические компоненты были заключены
там рано, и оставались непотревоженными. Дальнейшие тафономические ограничения
вывести трудно. Неясно, может ли отсутствие меланосом в кератиновых листках перьев
(рис. 2B и S3) дать дополнительную информацию, или оно следует из слабой
пигментированности оперения или малой площади, доступной для анализа сканирующего
электронного микроскопа. Эксперименты по искусственному насыщению показали
разрушение современных меланосом, но эти работы проводились при температурах,
которые разрушили бы и янтарь. Тафономический путь, привёдший к сохранению DIP-V-
15103 не вполне ясен, но он требует более детального изучения органики и пигментации
во включениях позвоночных в янтаре.
Остеология
Синхротронное фазоконтрастное рентгеновское микротомографическое сканирование
образца DIP-V-15103 (рис.1) показало, что плотность мягких тканей отличается от частично
замещённых скелетных элементов незначительно, чтобы позволить получить
рентгеновские изображения и виртуально исследовать только остеологию.
Соответственно, её многие диагностические и сравнительные детали остаются
неизвестными. Но два позвонка чётко разграничены вентрально (рис. 1F–1H).
Экстраполяция их длин даёт, что сохранившийся кусок хвоста содержит как минимум
восемь целых позвонков и часть девятого. Позвонки вытянуты, их антеропостериоральная
длина в два раза больше максимального диаметра хвоста (табл. S1). Их пропорции и
изогнутость хвоста исключают принадлежность к Pygostylia. Даже с кожей, прижатой к
костной поверхности, никаких других особенностей, кроме вентральных желобков на двух
телах позвонков, не наблюдается. Такое отсутствие топографии предполагает, что позвонки
не имели заметных нейральных или гемальных дуг, или продольных отростков. Таким
образом, сохранившийся сегмент - лишь небольшой срединно-дистальный кусок того, что
было довольно длинным хвостом с общим числом позвонков не меньше 15, а вероятно,
больше 25. Судя по величине образца, он также видимо принадлежал ювенильной особи.
DIP-V-15103 интерпретируется как хвост нептичьего целурозавра: строение позвонков и
расчётная длина вычёркивают веерохвостых птиц, овирапторозавров, и
скансориоптеригий — линии, характеризуемые главным образом коротким хвостом с
примерно равными телами позвонков, за исключением эпидендрозавра. Перья с
бородками имеют слабо сцепленное строение, похожее на таковое у нептичьих
целурозавров, в частности, паравиев. Хотя они скорее контурные, ни одна видимая
остеологическая черта не исключает родство их носителя с компсогнатами. Расположение
таких перьев парами по длине хвоста может говорить о принадлежности к Pennaraptora, и
показывает нижнюю границу филогенетического положения образца. Однако, их
распределение и форма сильно указывает на место ближе к крон-группам базальных
целурозавров, таких, как тираннозавриды и компсогнатиды. Касательно верхней границы,
образец надёжно может быть исключён из Pygostylia; кроме того, он, вероятно, может
быть исключён и из ящерохвостых птиц, судя по выраженным вентральным желобам на
телах позвонков. Вспомогательные таксономические детали приведены в Дополнительной
информации.
Оперение
И синхротронное фазоконтрастное рентгеновское микротомографическое сканирование, и
стандартная оптическая микроскопия подтверждают, что перья прикреплены по всей
сохранившейся длине хвоста (рис.1). Двусторонний парный ряд перьев, ориентированных
прстеродорсально, тянется по срединной дорсальной линии (рис. 1D и 1E). Другой ряд
перьев есть на средней части высоты с каждой стороны хвоста, и перья растут
постеролатерально под углом примерно 45' к их длинной оси (рис. 1D–1G). Их пары
фолликул выглядят распределёнными равномерно по длине хвоста. Там, где видны
очертания двух позвонков, фолликулы находятся на середине их длины и на
межпозвоночных суставах. Вентральное оперение редко, и состоит из тонких лёгких
перьев, растущих близко к длинной оси хвоста (рис. 1B, 1G, и 1H). В общем, перья образуют
направленные латерально кили с обеих сторон позвоночного столба, что даёт нам
уникальную возможность видеть их количество и положение в хвостовом, похожем на
рулевое, оперении целурозавра. DIP-V-15103 не показывает расширения крупных рулевых
перьев, наблюдаемого на задней части хвоста ящерохвостых птиц. Его либо не было у этой
особи, либо оно было вне сохранившейся области. Однако, то, как перья DIP-V-15103
выстроены в боковые кили, похоже на веер или кисточку на хвосте у паравиев. Такое
устройство, с разными типами перьев, может быть не только на его кончике, но и по всей
длине. Сохранение в янтаре предполагает, что эти паравиевые веера и кисточки - не
тафономический артефакт раздавливания. Если DIP-V-15103 действительно является
хвостом ювенильного целурозавра, то перья, вероятнее всего, характеризуют взрослых
особей; но остаётся некоторая неопределенность. Базальные таксоны Pennaraptora, такие,
как Similicaudipteryx, как считается, подвергались радикальной линьке, затрагивавшей и
хвост, а у некоторых базальных представителей Pygostylia был выводковый молодняк с
оперением, как у взрослых. Рулевые перья и бородки DIP-V-15103 предполагают оперение,
похожее на взрослое, при котором перья в последующих линьках не будут замещены
другими морфотипами. Как альтернатива, носитель перьев мог иметь такие типы линьки,
которых нет у современных птиц. Все сохранившиеся перья DIP-V-15103 похожи друг на
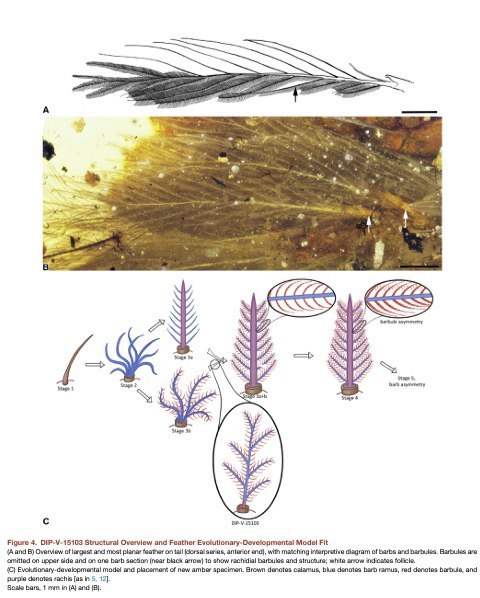
друга морфологией, независимо от положения на хвосте (рис.3 и S4). Все они имеют слабо
выраженный стержень, почти неотличимый от бородок, и слегка утолщённый внизу
(рис.3). И стержни, и бородки в сечении субцилиндричны. Хотя стержень книзу утолщается,
его максимальный диаметр у фолликула примерно в три раза больше ближайшей бородки
(рис. 3 и S4). Перья у переднего конца дорсальной серии имеют самое широкое основание,
где ширина оси достигает 60 мм (рис. 3,4A, и 4B). Оси этих перьев в апикальной позиции не
шире 18 мм, а ширина бородок варьирует от 15 до 23 мм. У отдельных перьев бородки
расположены чередуясь вдоль оси, с большими промежутками между ними и слабой
плоскостной организацией (рис.4). Изгиб внутри янтаря указывает, что бородки были
гибки, как и в известной мере стержень. Такая свободная структура этих перьев более
похожа на современные декоративные, чем на предназначенные для полёта, и имеют
структурные сходства с дистальными компонентами контурных перьев некоторых
Anseriformes (гусеобразных) (рис. 3 и S5). Их парное устройство похоже на таковое у
рулевых перьев современных птиц, что намекает на то, что эта черта хвостового оперения
сложилась до развития пигостиля, и это оперение с течением времени стало более
специализированным. Если на всём хвосте были такие перья, как в DIP-V-15103, их
обладатель, скорее всего, не мог летать. Перья DIP-V-15103 демонстрируют великолепно
сохранившиеся бородочки. Удивительно, но простые бородочки ветвятся не только на
отдельных бородках, но и на стержне (рис. 3,4,S4G, и S4H). В этом отношении, перья
сравнимы с контурными многих современных птиц, на которых также имеются отдельные
бородочки на стержне, хотя обычно они растут из основания ближайшей бородки и в
редуцированном виде. В DIP-V-15103, они ветвятся на равном расстоянии, парно и почти
симметрично. Этот паттерн сохраняется на проксимальных и дистальных бородочках, от
проксимальных к дистальным бородкам, и вдоль стержня. Бородочки имеют форму
лезвия, с пигментацией, окаймляющей пять базовых клеток, за которыми следует
слабодифференцированный пеннулюм (верхняя часть бородочки), лишённый выраженных
узлов или узловых выступов (рис.3E–3H). Малое расстояние между бородочками, в
сочетании с положением их плоских поверхностей (параллельно длинной оси пера),
являет нам скорее контурные перья. Слаборазвитый стержень и ветвление
соприкасающихся бородочек у DIP-V-15103 является новой для теропод комбинацией. В
рамках модели эволюционного развития перьев, DIP-V-15103 выглядит промежуточным
между стадиями IIIa (ось с голыми бородками) и IIIb (бородки с бородочками, без оси), но
не вполне точно соответствует стадии IIIa+b (ось с бородками, несущими бородочки)
(рис.4C). У DIP-V-15103, бородки чередуются вдоль слабо оформленного стержня, ветвясь
апикально почти дихотомически, и бородочки идут вдоль и стержня, и бородок.
Слаборазвитый стержень, по-видимому, сформировался путём слияния отдельных
бородок, на которых уже были бородочки (стадия IIIb), а не слияния голых (стадия IIIa).
Схема их ветвления не меняется до самого фолликула, как и у многочисленных,
недифференцированных бородочек. Если наблюдаемое в DIP-V-15103 положение не
является вторичной редукцией оси, эволюционный путь перьев этого целурозавра мог
проходить через стадию IIIb (бородки с бородочками), а не IIIa (слияние голых бородок).
Цитологические наблюдения за развитием бородочек по краю опахала бородок
поддерживают версию эволюционного слияния бородок с бородочками. Морфология
перьев DIP-V-15103 контрастирует с редуцированной осью и длинными, голыми,
нитевидными бородками на хвостовом оперении дромеозаврида Sinornithosaurus и
теризинозавроида Beipiaosaurus. Это намекает либо на большее разнообразие такового
оперения у целурозавров, чем полагалось прежде, либо у DIP-V-15103 упрощённая форма
более продвинутых контурных перьев. Необычная конфигурация бородочек DIP-V-15103
предполагает, что они примитивно распределялись по всей длине пера, и только после
ограничились бородками и прилегающей осью и ориентированы так, что их стороны
смотрят на поверхность перьев, как у современных птиц. У них клетки бородочек
зарождаются в субперидерме (второй слой эпителия), и соединяются в синцитий
(многоядерная протоплазма, не разделённая на отдельные клетки) на обеих сторонах
краёв опахала бородки. Симметричное устройство бородочек вдоль бородок у DIP-V-15103
говорит о симметрии их клеток вдоль этих краёв. Постоянное ветвление бородочек вдоль
оси, вероятно, шло по краю опахала бородки к самой апикальной из них. В линии, ведущей
к птицам, бородочки стали ограничены бородками и проксимальной частью оси, скорее
всего, как приспособление к росту числа и густоты бородок на жёстких контурных перьях
(стадия IIIa+band/или стадия IV). Как альтернатива, паттерн бородок у DIP-V-15103 может
быть весьма продвинутой и потенциально экспериментальной особенностью, не
связанной с линией птиц. Как бы то ни было, DIP-V-15103 предполагает, что нептичьи
тероподы имели большую вариативность форм перьев, чем предсказывалось по
онтогенетическим фенотипам современных перьев. В захороненном оперении есть следы
пигментации. Отдельные полосы, соответствующие базовым клеткам в каждой бородочке,
видны благодаря заключенным пигментам (рис. 3C–3H). Пигментация более выражена в
апикальных частях бородочек и на бородках и стержнях дорсальных перьев (рис. 1C и S4H).
Окраска у отдельных перьев отличается мало, но те из них, что сверху, заметно темнее, чем
те, что снизу. Сохранившийся окрас говорит об орехово-коричневой верхней поверхности,
контрастирующей с бледным или почти белым нижним оперением (рис. 1A–1C и S4A–S4D);
впрочем, влияние тафономии на видимый цвет неясно. Небольшой участок бледных
нижних перьев доступен для исследования SEM. Меланосом не было обнаружено, что
предполагает, что эти перья либо лишены пигмента, либо были окрашены иным способом,
например, каротеноидами. Листы кератина видны в слое перьев, показывая чёткую,
пористую, пластинчатую структуру, наблюдаемую также на бородочках современных птиц
с помощью SEM (рис. S2A и S2B). Рассмотренный здесь хвост теропода - изумительное
ископаемое, являющее нам уникальный потенциал янтаря для сохранения органики. Что
важно, в контексте происхождения птиц перья и полёт - ключевые элементы успеха этой
клады. Недавние находки в Азии открыли нам неожиданное разнообразие морфологий
перьев и способов полёта среди мелких теропод юрского и мелового периодов,
процветавших около времени появления птиц с машущим полётом. DIP-V-15103 добавляет
к этому разнообразию ещё один морфотип. Интеграция исследований развития и
палеонтологии даёт нам более глубокие модели эволюции морфологических черт,
помогающие нам объяснить важнейшие переходы, такие, как от теропод к птицам. При
сохранении в янтаре, тончайшие детали видны трёхмерно, и показывают нам конкретный
вид морфологии перьев и их расположение на хвосте, также как и подтверждают важную
роль бородок и бородочек в эволюции перьев.
Автор перевода: Aoife Cormach
Источник: https://vk.com/dromeo