R1b — гаплогруппа Y-хромосомы, наиболее распространённая в Западной Европе и на Южном Урале. Встречается также в Центральной Азии, Восточной Европе, Северной Африке, Западной Азии. После миграций европейцев в Америку и Австралию она составляет значительную долю и там. Определяется однонуклеотидным полиморфизмом M343, открытым в 2004 году
С 2001 по 2005 год R1b определяли наличием ОНП P25. В старых системах классификации — Hg1 и Eu18
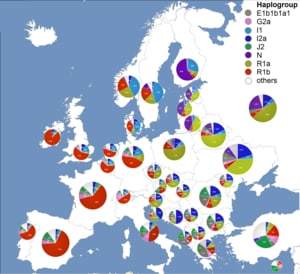
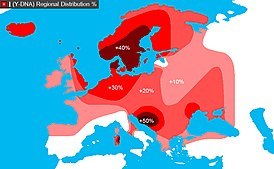
Наиболее часто встречается в отцовской линии в Западной Европе, а также в некоторых частях России — у нескольких башкирских родов и в Дагестане. В центральной Африке — Чад и Камерун. В более низком содержании гаплогруппа есть в Восточной Европе, Западной Азии, в некоторых частях Северной Африки и Центральной Азии.
Происходит из гаплогруппы R1, также как и гаплогруппа R1a.
Гаплогруппа R1b возникла от мутации гаплогруппы R1, произошедшей у мужчины, жившего ок. 22 800 лет назад (дата определена по снипам компанией YFull). Последний общий предок носителей R1b жил 20,4 тыс. лет назад
Согласно ряду исследований в области ДНК народов, гаплогруппы R1b и R1a распространены столь обширно в том числе и благодаря миграциям носителей индоевропейских языков.
Британские генетики Брайан Сайкс и Стивен Оппенгеймер утверждают, что гаплогруппа R1b не имеет отношения к индоевропейскому заселению Западной Европы и происходит от (по их мнению) палеолитического (доиндоевропейского) населения Иберии (баски). Большинство исследователей не считает басков палеолитическими. Взгляды Сайкса и Оппенгеймера получили широкое распространение в Европе благодаря написанным ими популярным бестселлерам о генетической истории Европы.
С другой стороны, «европейская палеолитичность» R1b сталкивается с непреодолимыми противоречиями. Последующие исследования Барбары Арреди с коллегами установили, что разнообразие субклад данной гаплогруппы увеличивается по мере движения на восток, что скорее говорит о восточном происхождении данной гаплогруппы и связано с распространением сельского хозяйства из Западной Азии в Европу
Ряд современных генетиков полагают, что R1b зародилась в Центральной
или Западной Азии
Сначала была выдвинута гипотеза, что R1b является коренной для Западной Европы, поскольку именно там она преобладает. Впоследствии было доказано, что гаплотипы R1b демонстрируют большее разнообразие малых побочных ответвлений в Анатолии и на Кавказе, чем в Европе. Однако известно что древние греки практически отождествляли население Кавказа (Иберия) и современной Испании (Пиренейский полуостров). Также европейские субклады более молоды по сравнению со средневосточными или центральноазиатскими. Основная западноевропейская ветвь R-P312/S116 восходит всего лишь к 3500 или 3000 до н. э., следовательно, старейший общий предок этой линии жил по крайней мере 5000 — 5500 лет назад в долине нижнего Дуная или в Причерноморье. В любом случае, эти временные рамки слишком малы для палеолитического происхождения или неолитического пришествия R1b. Открытие субкладов гаплогруппы R1b в Средней Азии, Пакистане и Индии окончательно опровергло её палеолитическое происхождение в Западной Европе и подтвердило её связь с индоевропейцами
Этот вывод подтверждается дальнейшими исследованиями подгруппы R1b1a2, носителем которой, возможно, являлись фараоны Эхнатон и Тутанхамон. Она, предположительно, зародилась на Кавказе около 9500 лет назад — известно, что до расселения семитов кавказские народы (урарты; хатты; хурриты) были широко расселены по территории Ближнего Востока — и начала миграцию в Европу около 7000 лет назад (см. Балканский неолит). Часть её представителей мигрировала в Северную Африку[16] (примером массовой миграции из Ближнего Востока в Египет является нашествие гиксосов, которое имело место за полтысячелетия до рождения Тутанхамона). Ок. 5700-7300 лет назад (неолитический субплювиал) из Евразии в район озера Чад пришли носители субклады R1b1a2-V88
R1b возникла от гаплогруппы R. Место — Северная Азия. Промежуток — между 26 500 — 19 000 лет назад, в этот период был последний ледниковый максимум. Самая древняя находка гаплогруппы R* — останки мальчика MA1 со стоянки Мальта́ (Алтайский край) возрастом 24 000 лет. Популяция из Мальты произошла от популяции северо-восточных сибиряков (Ancient North Siberians, ANS), представленных обитателями Янской стоянки
Носители гаплогруппы R внесли большой вклад в происхождение современных европейцев и современных жителей Южной Азии. В этих регионах гаплогруппа R наиболее распространена — R1b в Западной Европе, R1a в Восточной Европе, Центральной и Южной Азии, и R2 в Южной Азии. Данные выводы подтверждаются тем, что самые старые мутации R1b — M343, P25, L389. обнаруживаются на огромной площади от Западной Европы до Индии. Объяснению этому — именно охотники позднего палеолита, которые перемещались на этих пространствах во время последнего ледникового периода
Три основные субклады R1b1 — R1b1a, R1b1b, R1b1c происходят с Ближнего Востока. Южная ветвь R1b1c — R1b1a2-PF6279/V88 обнаруживается в Леванте и Африке. Северная ветвь R1b1a — R1b1a1a-P297/PF6398 возникла в районе Кавказа, восточной части Анатолии и возможно в Северной Месопотамии. Затем носители R1b1a1a-P297 двинулись в Европу и Центральную Азию, разделившись на две ветви. R1b1b1-M335 обнаруживается только в Анатолии.
Взрывообразный рост числа потомков основателя субклады R1b-P310/L11/L151/PF6542/P311/L52 (R1b1a1a2a1a) произошёл примерно 5,9—4,8 тыс. л. н., что совпадает с распространением культуры колоколовидных кубков в Западной Европе и культуры боевых топоров в Восточной Европе
Скотоводы периода неолитаПравить
Учёными выдвинута теория, что носители R1b (соседствовавшие в период неолита с носителями J2) стали первыми племенами, одомашнившими крупный рогатый скот. Место этого события — Месопотамия около 10 500 лет назад. Племена носители R1b произошли от племён, охотившихся на мамонтов в период палеолита, до окончания (Последняя ледниковая эпоха) ледникового периода. Когда наступило потепление примерно 12 000 лет назад, стала быстро исчезать огромная биологическая зона — Тундростепь и стали резко вымирать представители мегафауны того периода, в том числе мамонты, шерстистые носороги, гигантские олени и ряд других животных, ныне встречающиеся только как ископаемые виды. Бывшие охотники на мамонтов перепрофилировались на охоту на более мелких представителей мегафауны — бизонов и зубров.
В регионе Плодородного полумесяца, где проживали эти племена R1b, уже 12 000 лет назад стала быстро расти плотность населения, поголовье диких животных сокращалось и племена постепенно перешли к выборочной, контролируемой охоте и затем стали создавать первые стада, чтобы прокормиться. Быстрее всего удалось приручить и затем одомашнить коз, затем кабанов. Носители R1b стали скотоводами (см. Пастушеские племена) кочевого и полу-кочевого типа.
Другие племена проживавшие в Плодородном полумесяце, представленные генетически E1b1b, G и T, выбрали путь оседлого земледелия, делая упор на растениеводство и содержа меньшее количество домашнего скота.
Генетический анализ ДНК современных коров и его сравнение с ископаемыми образцами животных, показали, что все современные коровы, которые происходят от дикого быка, прирученного в период неолита и зарождения земледелия, ведут свой род от популяции, насчитывающей 80 животных, первых одомашненных коров. Самые ранние свидетельства одомашнивания коров датируются 10 500 лет назад, находки сделаны в культурах, владевших гончарным мастерством в период неолита, место нахождения — Тавр (хребет) южные прибрежные горы на территории современной Турции. Самые древние поселения со следами одомашнивания коров — Чайоню на Юго Востоке Турции и в месте Джауд эль Мугара,
на Севере Ирака.
Эти поселения периода неолита расположены в 250 км друг от друга. Именно из этой области, как предполагают генетики и археологи, племена носители R1b начали своё расширение и таким образом эта область считается «родиной» гаплогруппы R1b.
Уже на раннем этапе, как только началась миграция, племена R1b разделились на три группы.
1 — Ветвь M335 осталась на территории Анатолии, при этом сегодня она почти не встречается среди современных людей, что говорит о не успешном развитии её носителей, возможно вынужденных конкурировать с другими культурами периода неолита, жившими на территории Анатолии — Малой Азии, данный регион был уже тогда густо населён, при этом он гористый и пастбищ было недостаточно. Это доиндоевропейская ветвь
2 — Ветвь V88 мигрировала на Юг и обосновалась в Леванте. Из Леванта ряд носителей V88 дошли до Египта, затем колонизировали почти всю Северную Африку от побережья Средиземного моря до зоны Сахеля. Это доиндоевропейская ветвь.
3 — Ветвь P297 перешла Кавказ и попала в степи, расположенные между Чёрным и Каспийским морями — Понто Каспийские степи. Данный регион в тот период времени обеспечил идеальные площади для выпаса большего количества скота. В регионе Понто Каспийских степей произошло разделение 3-ей ветви P297 на две — R1b1a1 M73 и R1b1a2 M269.
Типично индоевропейскими МТ-ДНК являются — H2a1, H6, H8, H15, I1a1, J1b1a, W3, W4 и W5, эти гаплогруппы женской линии будут обнаруживаться в Ямной культуре и Унетицкой культуре и затем уже в других индоевропейских культурах.
Лошади были одомашнены около 6 600 лет назад в Каспийской степи или в районе Дона или в районе Нижней Волги. Этот прорыв стал определяющим для степных культур. Однако R1b не одомашнивали лошадей, они пришли в Понтийско Каспийские степи, когда эти животные уже были поставлены на службу людям. Наиболее вероятно, что лошадей одомашнили носители R1a, вкупе со старой ветвью R1b-P297, немногочисленной по сравнению с R1a. R1b-P297 ещё ни в коем случае не была индоевропейской, это были люди охотники ещё жившие в ледниковом периоде и не знавшие скотоводчества. Эта ветвь поселилась в Понтийско Каспийских степях в позднем палеолите или мезолите, что подтверждается находками в Самаре и Латвии. R1b-P297 и R1a по составу геному, были почти чисто коренными людьми, периода мезолита, жившими в Восточной Европе. В них имелась небольшая примесь от Сибирских охотников палеолита и мезолита, но отсутствовала примесь от жителей Кавказа. При этом примесь жителей Кавказа найдена у культур халколита — Афанасьево, Ямной и Культуры шнуровой керамики.
Точный маршрут миграции R1b-M269 от Южного Кавказа до Понтийско Каспийской степи пока не ясен. Возможно это было в период Днепро Донецкой культуры, 7 100 — 6 300 лет назад. Данная культура стала первым обществом уровня неолита в Прикаспийской степи, у днепро донецких носителей R1a и / или I2a1b, коровы и козы уже паслись в степи и появились похоронные ритуалы, они так же импортировали медь и медные изделия с территории Балкан и Карпат. Часть своих культурных традиций они переняли у охотников периода мезолита. Знания о домашних животных пришли к данной культуре от земледельцев с Ближнего Востока, это были носители I, I2a2, R1a, R1b1a (L754) и один R1b1a2 (L388). Что важно — среди этих земледельцев ещё не было носителей ветвей R1b-M269 или R1b-L23 — которые затем доминировали в Ямной культуре. МтДНК днепро донецкой культуры представлена исключительно женщинами европейского происхождения, периода мезолита — U4a, U4b, U4d, U5a1, U5a2, U5b2, а также одна J2b1 и одна U2e1, все эти мтДНК гаплогруппы не индоевропейские, а до индоевропейские. Вплоть до периода бронзового века, индоевропейские мтДНК не обнаруживаются в образцах периода неолита, найденных на территории Украины. Таким образом, Днепро Донецкая культура, которая уже имела и скотоводческие знания и земледельческие и являлась самой ранней подобной культурой в Понтийской степи, не относится ни к индоевропейской культуре ни к прото индоевропейской культуре
Первыми прото индоевропейскими культурами были Хвалынская (7 200 — 6 500 лет назад) и Среднестоговская (6 600 — 5 900 лет назад). В этот период появляются небольшие захоронения курганного типа, с характерным положением мёртвых на спине, с поднятыми коленями, ориентированными на Северо Восток. В этих культурах население активно вступало в связи с другими племенами, это заметно по разнообразным формам черепов. 7000 лет назад элитная прослойка этих культур начала активное развитие с использованием стад КРС, лошадей и меди. Медь была именно статусным металлом, дорогим и относительно трудно добываемым и обрабатываемым. На рубеже периодов Хвалынской и Среднестоговской культур, появилась ветвь R1b-M269 — L23, это произошло примерно 6 500 лет назад. В итоге в современный период 99 % индоевропейской R1b ведёт свой корень от этой ветви — L23.
От R1b-M269 так же произошла другая клада PF7562, но она немногочисленна и представлена на Балканах, в Турции и Армении.
Вторая миграция через Кавказ произошла около 5 700 лет назад и совпадает с возникновением Майкопской культуры, первой в мире культуры Бронзового века. Её возникновение на Северо Западе Кавказа до сих пор вызывает вопросы — откуда пришли её носители.
Наиболее вероятно, что носители Майкопской культуры родом из Северной Месопотамии, Сирии и Восточной Анатолии. На Южном Кавказе в Грузии и Азербайджане обнаружена Лейлатепинская культура (6 350 — 6 000 лет назад). керамика майкопской и лейлатепинской культур имеет схожие черты, так же похожа на их керамику посуда из поселений Куро Аракской культуры.
Важно, что археологические находки показывают явное распространение курганных погребений с бронзовыми изделиями из Майкопской культуры в Понтийской степи, именно в этой степи вскоре возникал Ямная культура — около 5 500 лет назад. Курганный тип погребений стал доминирующими является одним из маркеров для определения принадлежности культуры к индоевропейской. Курганы как способ погребения знати затем использовались кельтами, римлянами, германскими племенами, скифами.
Между 5 500 — 4 500 лет назад активно развивалась Ямная культура бронзового века. Она считается бесспорно индоевропейской и наиболее важной в основании индоевропейской культуры и общества. R1b-M269 носители с Ближнего востока в Ямной культуре относительно активно вступали в отношения с земледельцами и пастухами-скотоводами R1a на протяжении более чем 2-х тысяч лет и так же после Ямной культуры, в других сообществах.
Эти контакты между R1a и R1b не носили спорадического характера, а были плотными и многоуровневыми, как на уровне знати, так на уровне простых людей и торговцев. Это привело к постепенному формированию своеобразной общности населения Ямной культуры, у Ямной культуры появился относительно единый язык, который сегодня называют — Праиндоевропейский язык, который стал впоследствии основой для почти всех современных Европейских языков, включая все славянские языки, все англосаксонские и так далее. Лингвисты на современном этапе изучения, сошлись во мнении, что именно регион Понтийско Каспийских степей является родиной праиндоевропейского языка. Это в том числе подтверждается сходством праиндоевропейского языка с кавказскими и хуриитскими языками, ведущими своё начало с Ближнего Востока и с другой стороны с Волго — Уральскими языками. Только в Понтийско каспийской степи исторически могли пересекаться так плотно пути носителей этих языков, что бы в итоге возник сначала праиндоевропейский язык — Доиндоевропейский субстрат, затем индоевропейский язык.
Ямная культура наиболее чётко характеризуется тем, что люди её составлявшие использовали телеги на четырёх колёсах, для того чтобы менять место жительства, перевозили на них еду и домашние принадлежности. Подобный транспорт обеспечивал возможность длительных походов в степях, что делало ямников одним из самых мобильных народов, при этом занимавшимся и земледелием и скотоводством одновременно. Этот набор умений положил начало великим миграциям индоевропейцев. В более поздние времена, в период Древнего Рима, галлы и древние германцы так же вели свои миграции.
Ямная культура, учитывая её большие размеры, не была единой. На Юге, вдоль Северного побережья Чёрного моря до Северного Кавказа, была полноценная степь, почти без леса, эта степь простиралась дальше, до Каспийского моря, до Сибири и до Монголии и в англоязычной литературе называется Евразийской степью.
В настоящее время ряд исследований.убедительно доказали, что древние носители R1a и R1b
способствовали распространению мутации (белый) гена. Эта мутация и ген определяют более светлую кожу европейцев. Частота встречаемости аллелей (белый) гена почти идеально коррелирует с распространением R1a и R1b по Европе, Ближнему Востоку, Южной и Центральной Азии. Эта мутация уже в период раннего неолита была передана земледельцам Ближнего Востока и поэтому они уже несли в себе аллель (белый), хотя с меньшей частотой, нежели современные жители Европы и Центральной Азии. Аллель светлой кожи (белый) встречается в концентрации от 15 до 30 % у различных народов в Северной Африке к Югу от Сахары. В том числе это племена фулани и хауса R1b-V88. Это говорит о том, что ген светлой кожи уже был у носителей R1b до разделения их на два потока V88 и P297, на Ближнем Востоке.
Народы, у которых высока концентрация R1a, имеют ту же высокую частоту аллели светлой кожи (белый) что и R1b. С другой стороны данная аллель (белый) не была обнаружена в образце R, который был найден в Сибири и имеет возраст 24 000 лет.
Так же данная аллель отсутствует в большинстве популяций, носителей R2 в Индии и Юго Восточной Азии. Это позволяет обоснованно предполагать, что аллель светлой кожи (белый) возникла уже в линии R1 в период позднего палеолита, в промежутке между 20 000 — 13 000 лет назад.
Наличие светлых волос является ещё одной физиологической чертой, присущей древним индоевропейцам.
Наличие светлых волос является ещё одной физиологической чертой, присущей древним индоевропейцам.
Но голубые глаза уже были среди охотников мезолита Y гаплогруппы — I и возникли независимо от индоевропейских генов. Распространение светлых волос лучше всего коррелирует с гаплогруппой R1a. Распространение рыжих волос лучше всего коррелирует с гаплогруппой R1b и рыжие волосы не обнаруживаются в Европе до периода бронзового века.
Субклад R1b1a1a2a2-CTS1078/Z2103 обнаружен у представителя энеолитической вучедольской культуры
Гаплогруппа R1b была обнаружена у одного представителя культуры полей погребальных урн из саксонской пещеры Лихтенштейн в Германии (около 3000 лет назад, поздний бронзовый век), у представителей культуры колоколовидных кубков (около 2800—2000 лет до н. э.)
куро-аракской, ямной (Z2103), межовской культур, у представителей средней и поздней бронзы Армении
Поскольку самый ранний пример данной гаплогруппы датирован не позднее 14 000 лет назад и принадлежит R1b1a (R-L754), R1b должен был возникнуть относительно скоро после появления R1. Предполагается, что гаплогруппа R1b возникла в Западной Азии.
R1b является подклассом внутри макро-гаплогруппы K-M9, которая является одной из преобладающих гаплогрупп за пределами Африки. Место возникновения гаплогруппы K — Азия. Татьяна Карафет (2014) утверждает, что:
"быстрый процесс диверсификации K-M526, вероятно, произошел в Юго-Восточной Азии, с последующим распространением на западе предков гаплогрупп R и Q
Три генетических исследования в 2015 году оказали поддержку курганской гипотезе Марии Гимбутас относительно прародины индоевропейцев. Согласно этим исследованиям, гаплогруппы R1b и R1a, теперь наиболее распространенные в Европе (R1a также распространена в Южной Азии), были распространены из Западной Евразийской степи вместе с индоевропейскими языками. Также обнаружили аутосомный компонент, присутствующий в современных европейцах, которого не было у неолитических европейцев, который был привнесен с отцовскими линиями R1b и R1a, а также с индоевропейскими языками
Как заметили Барбара Арреди и её коллеги, распределение R1b в Европе образует клин с Востока на Запад, что соответствует теории проникновения её носителей в Европу из Западной Азии с распространением земледелия
Дальнейшие исследования 2009 года ch..., указывает на то, что гаплогруппа R1b имеет наиболее старые субклады в Западной Азии и наиболее поздние в Западной Европе.
При этом исследователи не отрицают того, что гаплогруппа R1b могла попасть в Европу в период неолита, то есть в более позднее время относительно прошлых оценок, в связи с миграциями земледельцев с Ближнего Востока
В 2016 году, генетики Монголии и Южной Кореи, провели исследования останков тел, найденных на территории Таван Толгой — это месторождение каменного угля на юге Монголии, в пустыне Гоби, в аймаке Умнеговь. Монгольские археологи, участвовавшие в раскопках могил Тавана Толгоя, предполагали, что 5 из 7 могил Тавана Толгой принадлежали Золотой семье, и одна из этих 5 могил считалась принадлежащей монгольской королеве.
Проведены исследования в мтДНК (женская) и Y-ДНК (мужская). Члены семьи являлись носителями гаплогруппы R1b — субклада M343 .
Изначально генетики предполагали, что сам Чингисхан и его семья были носителями гаплогруппы C3c субклада M48 — которая распространена в высокой концентрации среди современных монголов, казахов, эвенов, эвенков,
Но при исследованиях это не подтвердилось. Таким образом Чингисхан нёс в себе генетический вклад, связанный с Западной Европой и Передней Азией.
Люди, которых исследовали, могли произойти от браков между династией Борджигинов (к этой династии принадлежал Чингисхан) и кланами Онгуд или Хонгирад.
Часть южноазиатских генетических предков происходит от населения Западной Евразии, и некоторые исследователи предполагают, что Z93, возможно, попал в Индию через Иран и распространился там во время цивилизации долины Инда.
Маскаренья и др. (2015) предположили, что корни Z93 лежат в Западной Азии, и предположили, что «Z93 и L342.2 расширились в юго-восточном направлении из Закавказья в Южную Азию», отметив, что такое расширение совместимо с «археологическими данными экспансии населения Западной Азии на восток в 4-м тысячелетии до н. э., кульминацией которой стали так называемые миграции Кура-Аракс в период после Урука IV». Тем не менее, Лазаридис отметил, что образец I1635 из Лазаридис и др. (2016), их образец Кура-Аракс, содержал Y-гаплогруппу R1b1-M415 (xM269) (также называемую R1b1a1b-CTS3187).
Согласно Андерхилл и др. (2014) диверсификация Z93 и «ранняя урбанизация в долине Инда… произошла [5600 лет назад], и географическое распределение R1a-M780 может отражать это». Позник и др. (2016) отмечают, что «поразительные расширения» произошли в пределах R1a-Z93 ~ 4500-4000 лет назад, что «на несколько столетий предшествует краху цивилизации долины Инда».
Однако, согласно Нарасимхан и др. (2018), степные скотоводы являются вероятным источником R1a в Индии.
Праиндоевропейцы
О том, как восстанавливают поэтику праиндоевропейцев
Современные данные исторических лингвистов дают повод помещать индоевропейскую прародину в Понтийско-Каспийскую степь. Изначально до праиндоевропейцами были представители гаплогруппы R1a «обитавшие» в Самарской и Хвалынской культурах в 6-5 тысячелетиях до нашей эры. Приблизительно к 4600 году до нашей эры представители R1a уже одомашнили лошадь. К тому же времени представители гаплогруппы R1b уже вышли из-за Закавказья и одомашнили корову, благодаря чему у них появилась способность переваривать молочный сахар — лактозу.
С появлением Ямной культуры появилась и её неоднородность. Северную часть культуры населяли представители R1a, тогда как южную часть культуры населяли представители R1b. На симбиозе гаплогрупп появились так называемые — праиндоевропейцы, Ямной культуры.
Представители R1a, будущие носители славянских, балтийских, индо-иранских, албанского, армянского, фракийского и дакского языков, стали носителями сатемной изоглоссы, т. е. в этой группе велярные и лабиовелярные звуки праиндоевропейского языка развились в велярные, а палатовелярные стали шипящими и свистящими.
Тогда как представители R1b, будущие носители италийских (> романских), кельтских, германских, греческого, и вымерших венетского, древнемакедонского и тохарских языков, стали носителями кентумной изоглоссы, т. е. в этой группе велярные и палатовелярные звуки праиндоевропейского языка были объединены в велярные.
Затем представители гаплогруппы R1b мигрируют на восток, вплоть до западного Китая, став известными под именем — тохары, и на запад, в виде германских, кельтских, италийских, венетских, греческих и фригийских племен. Именно тохары, благодаря миграциям на восток, принесли свою R1b современным уйгурам, башкирам и т. д.
Появление балтийской группы
Считается, что балтийская ветвь возникла из Фатьяновской культуры (3200—2300 гг. до н. э.), северо-восточного продолжения культуры Шнуровой керамики. Кочевники раннего бронзового века R1a из северных степей и лесостепей смешались с уральскоязычными жителями (линии N1c1) региона. Это подтверждается сильным присутствием гаплогрупп R1a и N1c1 от Южной Финляндии до Литвы и на северо-западе России.
По лингвистическим данным славянская и балтийская группы объединяются в одну балто-славянскую ветвь с одним общим реконструированным языком путём сравнительно-исторических методов, и методов исторической Акцентологии, которая занимается реконструированием ударения.
Появление славянской группы
Происхождение славян восходит примерно к 3500 году до нашей эры с северной части Ямной культуры и её экспансией через Центральную и Северо-Восточную Европу с культурой Шнуровой керамики. Линии M458 и Z280 распространились по всей Польше, Беларуси, Украине и Западной России и сформировали ядро прабалто-славянской культуры. Высокая распространенность R1a в прибалтийских и славянских странах в настоящее время обусловлена не только экспансией Шнуровой керамики, но и длительной чередой более поздних миграций из России, последняя из которых имела место с 5 по 10 век нашей эры.
Появление индо-иранской группы, (ариев)
Праиндо-иранские носители языка, люди, которые позже называли себя «ариями» в Ригведе и Авесте, возникли в Синташта-Петровской культуры (2100—1750 гг. до н. э.), в Тобольской и Ишимской долинах, к востоку от Уральских гор. Он был основан кочевниками-скотоводами из Абашевской культуры (2500—1900 гг. до н. э.), простирающейся от Верхнего Дона-Волги до Уральских гор, и Полтавской культуры (2700—2100 гг. до н. э.), простирающейся от нижнего Дона-Волги до Каспийской впадины.
Появление греческой группы
Мало что известно о приходе прагреческих носителей из степей. Микенская культура началась около 1650 года до нашей эры и явно является импортированной степной культурой. Тесная связь между микенским и праиндо-иранским языками позволяет предположить, что они разделились довольно поздно, примерно между 2500 и 2000 годами до нашей эры. Археологически микенские колесницы, наконечники копий, кинжалы и другие бронзовые предметы обнаруживают поразительное сходство с культурой Сейма-Турбино (С. 1900—1600 гг. до н. э.) северных русских лесостепей, известных большой подвижностью своих кочевых воинов (Сейма-Турбинские стоянки были найдены аж в Монголии). Поэтому вполне вероятно, что микенцы спустились из России в Грецию между 1900 и 1650 годами до нашей эры, где они смешались с местными жителями, чтобы создать новую уникальную греческую культуру.
Появление германской группы
Германские языки, вероятно, не появились до нордического бронзового века (1800—500 до н. э.). Прагерманский язык, вероятно, развился как смесь двух ветвей индоевропейских языков, а именно прабалто-славянского языка Шнуровой культуры (R1a-Z283) и более позднего прихода праитало-кельто-германских народов из культуры Унэтиса (R1b-L11). Это подтверждается тем фактом, что германские народы являются гибридом R1a-R1b, что эти две гаплогруппы пришли разными путями в разное время, и что прагерманский язык ближе всего к праитало-кельтскому, но также имеет сходство с прабалто-славянским.
Анатолийская ветвь
Хетты (между 4000—3178 лет назад) были первыми индоевропейцами, которые столкнулись в военном отношении с Древним Египтом и Древней Месопотамией и одержали победы. Хетты пришли либо с Балканского полуострова либо с территории Майкопской культуры. Имеются значительные археологические и лингвистические факты, что Троя была построена индоевропейцами, связанными с культурой из степей и люди в ней имели гаплогруппу R1b. Теоретически, Майкопская культура могла основать Трою как колонию, для обеспечения торговых путей между Чёрным и Эгейским морями. Троя была основана примерно 5,000 лет назад и совпадает с временем создания первых галер.
С Балканского полуострова скорее всего пришли фригийцы и праармяне. Они пришли в Малую Азию — Анатолию около 3200 лет назад в период «великих потрясений» в Восточном Средиземноморье. Фригийцы основали королевство 3,200 лет назад, оно просуществовало 500 лет и находилось в Центральной части Анатолии, в западном её сегменте
Это королевство вобрало в себя большую часть разрушавшейся империи хеттов. Праармяне пересекли Анатолию, дошли до озера Ван и обосновались в Армянском нагорье. В современный период 30 % армян это носители R1b с субкладой L584 в Z2103. Большая часть R1b в современной Греции это балканская разновидность Z2103.