


Ответ на пост «Мужчина построил мини-отель для котиков в Подмосковье — в дождь там полная бронь!»
Оригинальная исследовательская статья
Front. Ecol. Evol. , 27 сентября 2018 г.
Раздел «Городская экология»
Том 6 - 2018 |https://doi.org/10.3389/fevo.2018.00146
Изменения во времени и пространстве использования крыс в ответ на хищничество диких кошек в городской экосистеме
Michael H. Parsons 1*
Peter B. Banks 2
Michael A. Deutsch 3
J
Jason Munshi-South 4
1. Department of Biological Sciences, Fordham University, Bronx, NY, United States
2. School of Life and Environmental Sciences, University of Sydney, Sydney, NSW, Australia
Абстрактный
Дикие кошки ( Felis catus ) — хищники, вызывающие массовую гибель местной дикой фауны в городских экосистемах. Несмотря на эти риски, кошек часто выпускают в качестве средства борьбы с городскими крысами ( Rattus spp.). Кошки могут влиять на свою добычу напрямую, убивая её, или косвенно, изменяя её рацион питания или использование пространства. Однако кошки предпочитают беззащитную добычу, и нет данных, свидетельствующих о влиянии кошек на крупных (>300 г) городских крыс. Мы использовали существующий метод радиочастотной идентификации (чипированные крысы и полевые камеры) и этограммы для оценки воздействия кошек, включая временные и пространственные модели использования, на активную колонию крыс. С 27 декабря 2017 года по 28 мая 2018 года мы записали 306 видеороликов с предварительно идентифицированными кошками и/или крысами, находившимися в одном и том же пространстве. Было зафиксировано три случая хищничества и 20 случаев преследования. Логистическая регрессия показала, что вероятность обнаружения крысы в определенный день связана с количеством кошек, замеченных в тот же день (ОР = 0,1, p < 0,001) или в предыдущий день (ОР = 0,15, p < 0,001). Также наблюдалось влияние на использование пространства. При каждом дополнительном обнаружении кошки вероятность перемещения крысы в направлении убежища увеличивается в 1,19 раза. Наши данные о низком уровне хищничества подтверждают мнение экологов о том, что риски для местной дикой природы перевешивают любые преимущества выпуска кошек. Несмотря на то, что крыс стали реже замечать, они просто изменили свои маршруты передвижения и остались в экосистеме. Наши данные о том, что присутствие кошек привело к уменьшению количества обнаружений крыс, могут объяснить распространенное представление об их ценности как хищников, несмотря на связанные с этим риски.
Кошечка, кошечка, где ты была?
Я был в Лондоне, чтобы навестить королеву .
Кошечка, кошечка, что ты там сделала?
Я напугал маленькую мышку под ее стулом .
… Джеймс У. Эллиот (1870)
Введение
Хищничество кошек по отношению к грызунам, особенно к мышам ( Mus, Peromyscus spp.), является одним из наиболее известных примеров взаимодействия хищника и жертвы: вдохновляющие детские стишки, мультфильмы и общественное восприятие. Однако многие неподготовленные наблюдатели не могут отличить мышей весом 20–35 г друг от друга (Zielinski et al.,1992) и городских крыс, которые часто весят в 10 раз больше (Парсонс и др.,2017Комбс и др.2018Это заблуждение, возможно, частично объясняет широкое распространение мнения о кошках как о «естественном» средстве борьбы с крысами. Однако у хищников есть энергетический бюджет, при котором их потребление калорий максимизируется (Jeschke et al.,2002) против потенциальных рисков, связанных с крупной или агрессивной добычей (Эмбар и др.,2014Таким образом, кошки, скорее всего, будут охотиться на птиц и более легкую добычу (например, мелких млекопитающих и их детенышей), прежде чем переходить к крысам. Кроме того, будучи одомашненными животными, кошки хорошо приспособлены к выживанию на подачках от людей (Монтагю и др.,2014Одомашнивание кошек и обилие более доступных альтернативных продуктов питания ставят под сомнение склонность кошек охотиться на городских крыс и сокращать их численность.
Дикие кошки могут влиять на динамику численности своей добычи, но существуют гораздо более убедительные данные, указывающие на то, что кошки влияют на местную дикую природу (Kauhala et al.,2015; Кикиллус и др.,2017чем городские крысы. Например, в Австралии (Дэвис и др.,2017) и США (Лосс и др.,2013Кошки представляют собой основной источник антропогенной смертности для местных птиц и млекопитающих. Это исторически признавалось в островных экосистемах, где кошки являются эволюционно новым видом (Войнарски и др.,2017), но это также верно для некоторых континентальных (Лосс и Марра,2017) и городских систем (Пиллай и др.,2018При нынешних темпах урбанизации последние представляют собой самые быстрорастущие экосистемы в мире, и ожидается, что к 2050 году более 70% населения будет жить в городах (Zhou et al.,).2013В отличие от местных позвоночных, наиболее пострадавших от кошек, городские крысы сосуществуют с кошками на протяжении веков. Эти крысы хорошо адаптированы к городским экосистемам; они представляют собой крупную добычу для всех, кроме самых смелых кошек, и могут защитить себя. Тем не менее, некоторые густонаселенные города, такие как Чикаго, штат Иллинойс, США, широко освещаются в средствах массовой информации (Глантон,2017) как массовые попытки использовать кошек в качестве средства борьбы с крысами. Выпуск кошек не только представляет опасность для дикой природы, но и вызывает опасения за благополучие самих кошек. Например, общеизвестно, что хорошо накормленная и ухоженная кошка может мирно сосуществовать с крысами. Таким образом, те, кто использует кошек в качестве средства борьбы с вредителями, могут намеренно избегать их кормления, чтобы побудить их к охоте (MP, личные наблюдения). Необходимы более убедительные доказательства, прежде чем можно будет оправдать использование кошек в качестве средства борьбы с крысами.
Из-за своих больших размеров городские крысы могут быть менее уязвимы для кошек, чем местные грызуны. Дикие кошки, как правило, предпочитают более мелкую добычу, в том числе мелких грызунов весом менее 250 г (Чайлдс,1986Большая часть доказательств, подтверждающих влияние кошек на виды рода Rattus , получена на островах, где островная карликовость может приводить к тому, что животные оказываются меньше своих континентальных сородичей (например, правило Фостера, Роцци и Ломолино).2017Например, Карл и Бест (1982В ходе исследования были изучены 229 образцов экскрементов диких кошек с острова Стюарт, Новая Зеландия. Крысы ( Rattus spp.) были обнаружены в 93% образцов экскрементов (по сравнению с 44% птиц). Однако взрослые крысы в этих системах обычно весят около 150 г (или вдвое меньше нью-йоркской крысы; Parsons et al.,2017Комбс и др.2018), включая полинезийских крыс ( Rattus exulans ), которые являются самыми маленькими из симбиотических с человеком крыс . Аналогично, Фицджеральд и др. (1991В ходе исследования были изучены экскременты диких кошек, обитавших на острове Рауль в течение 8 лет. Девяносто процентов крыс были полинезийскими, и менее 10% — более крупными (хотя и все же меньшими, чем обитающие на материке) норвежскими крысами ( R. norvegicus ).
Среди немногочисленных континентальных исследований, изучавших влияние кошек на крыс, результаты оказались неоднозначными. В Финляндии исследователи обнаружили, что 72% всей добычи, принесенной кошками домой, составляли грызуны. Однако авторы не проводили различия между мышами и крысами и указали, что почти половина всей добычи приходилась на исключительно крупных кошек (Kauhala et al.,2015Экспериментальный выпуск 20 местных длинношерстных крыс ( Rattus villosissimus ) в Австралии привел к их быстрому истреблению кошками (Франк и др.,2014Однако эти крысы были примерно вдвое меньше (150 г), чем большинство городских крыс. Также в Австралии анализ содержимого желудков 80 диких кошек показал, что 80% их рациона составляли грызуны, но большинство крыс были идентифицированы как длинношерстные (Yip et al.,2015).
Нет доказательств того, что кошки могут напрямую подавлять популяции городских крыс. Однако воздействие хищничества может также возникать косвенно, через нелетальные эффекты для жертв, связанные с риском хищничества. Запаха кошек ( Felus catus ) самого по себе достаточно, чтобы снизить репродуктивную способность некоторых мышей (Келлихер и Версингер,2009Вознессенская,2014) и крыс (Найденко и др.,2003Вознессенская и др.2003Вознессенская,2014Риск хищничества также может ограничивать доступ к пище (Герман и Валоне,2000), эффект, который может быть преувеличен в городских условиях. В городской среде крысы должны перемещаться по открытым пространствам в поисках нового убежища или доступа к пище или воде на некотором расстоянии (в отличие от мышей, которые могут метаболически производить собственную воду; Шмидт-Нильсен и О'Делл,1961Крысы могут выпивать до 30 мл воды в день (Сигел и Стакки).1947Избегание открытых участков характерно для многих видов мелких, «уязвимых» животных (Апфельбах и др.,2015), от мелких сумчатых животных Тасмании (Парсонс и Блюмштейн,2010) к полевым мышам ( Peromyscus polionotus ) в Южной Калифорнии (Orrock et al.,2004Оррок и Дэниелсон,2009Считается, что риск хищничества вызывает страх перед открытыми пространствами или полнолунием, когда животные оказываются на открытом воздухе, реже или на более короткие промежутки времени ищут пищу и остаются ближе к дому (Оррок и др.).2004Оррок и Дэниелсон,2009Однако данные о косвенном влиянии кошек на городских крыс также ограничены. Смешанные запахи хищников, исходящие от собак и кошек, отпугивали грызунов от кормушек, хотя авторы не проводили различий между мышами ( Mastomys natalensis ) и корабельными крысами ( R. rattus ) (Themb'alilahlwa et al.,2017). Совсем недавно Картэй и Бэнкс (2018Исследования показали, что свободноживущие корабельные крысы распознавали запах кошки, но не проявляли никаких антихищнических реакций ни в бдительности, ни в пищевом поведении, что позволяет предположить, что крысы рисковали, чтобы получить доступ к ресурсам.
Действительно, несмотря на существенные пробелы в знаниях о городских крысах, такие как хорошо описанное влияние феромонов на поведение и физиологию грызунов (Херст и Бейнон,2004; Такач и др.,2016), практически не проводились исследования непосредственно в городской экосистеме (Парсонс и др.,2017Это объясняется тем, что если крысы появляются в городе, то они обитают на чьей-то собственности. Владельцы не горят желанием разрешать эксперименты исследователей, если только эти эксперименты не приведут к истреблению крыс. Таким образом, у исследователей не было возможности изучать городских крыс в их естественной среде обитания, как это было сделано в работе Франка и др. (2014эксперимент, что позволяет получить более убедительные доказательства прямого или косвенного влияния кошек в данном месте на городских крыс.
Цели
В данной работе мы исследуем, как дикие кошки прямо или косвенно влияют на городских крыс. В рамках нашей долгосрочной программы исследований крыс в Нью-Йорке, США, мы имеем доступ к активной колонии крыс внутри мусороперерабатывающего предприятия, где, используя уже существующее исследование с применением радиочастотной идентификации (RFID), мы отлавливаем, чипируем и выпускаем животных, изучая при этом их индивидуальные жизненные истории, демографические характеристики популяции и поведение (Parsons et al.,2017Кроме того, в непосредственной близости от этого исследовательского участка уже несколько лет живут около двух десятков диких кошек. В частности, мы оценивали, влияют ли изменения в количестве кошек, присутствующих в данный день или в предыдущий день, на количество наблюдаемых крыс, или же количество крыс или людей может предсказывать количество наблюдаемых кошек. Нашими второстепенными интересами было определить, оказывает ли присутствие кошек какое-либо влияние на распространенность 8 распространенных поведенческих моделей крыс или направление движения по часто используемой тропе основной колонии.
Методы
Область исследования
Нью-Йорк (NYC), расположенный по координатам 40,71° с.ш., 74,01° з.д., является вторым по численности населения городом в Северной Америке, с плотностью населения около 2000 человек на квадратную милю (Гриффит и Вонг,2007Город с населением около 8,5 миллионов человек состоит из 5 районов, расположенных между рекой Гудзон и Атлантическим океаном. Климат характеризуется теплым, влажным летом (средняя месячная максимальная температура летом составляет 27,8°C, минимальная — 19,3°C) и холодной зимой (средняя месячная максимальная температура — 5,0°C, минимальная — −1,5°C) с годовым количеством осадков 50–200 см (NOAA). Бурые крысы ( Rattus norvegicus ) прибыли из Европы на кораблях между 1700 и 1750 годами (Puckett et al.,2016Комбс и др.2018Из-за малого количества естественных хищников они быстро превратились в вредителей. К хищникам относятся краснохвостые ястребы ( Buteo jamaicensis ), койоты ( Canis latrans ), рыжие лисы ( Vulpes vulpes ), домашние собаки (особенно терьеры; Canis familiaris ), куньи, такие как куница ( Pekania pennanti ), сбежавшие домашние хорьки ( Mustela putorius furo ) и одичавшие кошки ( Felis domesticus ). Этот район называют одним из «самых крысиных городов» на Земле (Роберт Корриган, Департамент здравоохранения и психической гигиены, Нью-Йорк, в отставке). Это разговорное обозначение является результатом обилия отходов, оставленных на ночь, количества мусора, оставленного на улице и в парках, и относительного отсутствия хищников.
Место проведения исследования
Мы получили разрешение на использование площадки для переработки промышленных отходов в Бруклине, юго-восточном районе Нью-Йорка. Исследовательская площадка расположена на одном из участков, принадлежащих компании Waste Management (WM). WM предоставляет комплексные услуги по управлению отходами в Северной Америке, начиная от сбора и утилизации и заканчивая переработкой и производством возобновляемой энергии. В рамках своей приверженности « поддержке клиентов и сообществ» WM согласилась разместить исследовательский проект по изучению крыс в Нью-Йорке на территории Фордхэмского университета. Завод по переработке отходов представляет собой закрытое полузакрытое здание, которое, как и любая промышленная зона, обеспечивает достаточное укрытие для грызунов. Работа ведется круглосуточно, шум от грузовиков и экскаваторов, а также может быть вредным из-за собранного мусора. Для мелких животных обеспечен постоянный приток потенциальной пищи, поскольку централизованные операции требуют доставки, сортировки, переработки и утилизации мусора или его вывоза. Площадка не оборудована системой климат-контроля, и животные могут рыть норы под полами и глубоко внутри стен, поэтому температура внутри может меняться в зависимости от местоположения. Наша основная колония крыс (рис. 1 ) расположена в основном в неосвещенном месте, где днем и ночью либо слабое освещение, либо темнота. Основной источник пищи находится непосредственно к западу от главных нор.
Рисунок 1
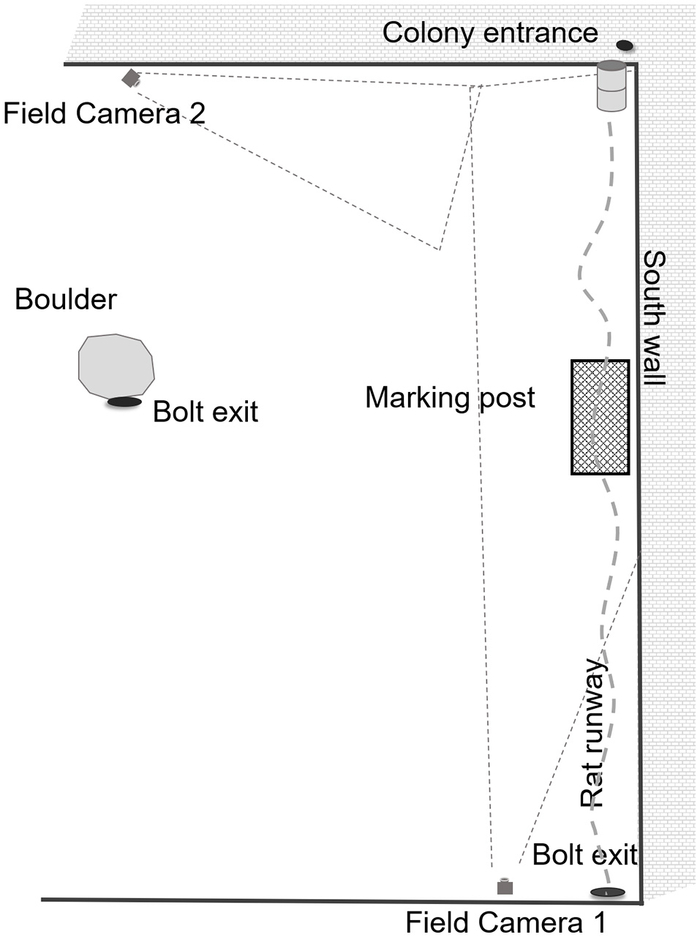
Экспериментальная установка для проведения испытаний с участием диких кошек в полузакрытом промышленном здании в Нью-Йорке в период с 27 декабря 2017 года по 28 мая 2018 года.
Экспериментальный дизайн
В настоящее время мы проводим исследование поведения крыс на нескольких площадках в Нью-Йорке, оценивая их реакцию на запахи и составляя этограммы и бюджеты активности для документирования поведения городских крыс в естественной среде. В ходе последних этапов наших предыдущих экспериментов (неопубликованные данные Parsons et al.) несколько кошек проникли на территорию нашего исследования и оставались там на протяжении всего периода. Вместо того чтобы останавливать эксперименты, мы разработали исследование для количественной оценки влияния кошек на поведение и передвижения крыс.
Наши методы отлова живых животных и анестезии были описаны ранее (Parsons et al.,2015,2016(Парсонс и др., неопубликованные данные). Также был установлен стационарный центральный процессор RFID с регистратором данных (PADAR; UID, Чикаго, Иллинойс, США) (Парсонс и др.,2016Колония крыс (рис. 1 ) насчитывала, по оценкам, 120–150 особей, исходя из 37 случаев отлова живых животных (большинство из которых были использованы в другом исследовании (Парсонс и др., неопубликованные данные)). Средний вес составлял 337 ± 13,6 г, а средняя длина — 375 ± 4,2 мм (включая хвост). Кроме того, животные из этой популяции активны большую часть дня и вечера, при этом некоторые особи активно регистрируют свое присутствие с помощью RFID-датчиков в течение дня и ночи.
Мы установили две инфракрасные полевые камеры (Browning Strike Force Elite HD) со скоростью срабатывания затвора 0,4 с, направленные с северо-востока на юго-восток и с запада на восток, по активной, хорошо сформировавшейся колонии крыс внутри городского промышленного центра (рис. 1 ). Мы записывали эти данные с 29 декабря 2017 года, когда были обнаружены первые кошки, по 28 мая 2018 года. Мы подсчитали количество срабатываний от кошек, крыс и людей и перевели их в процент от общего количества записанных видео в качестве основных переменных отклика, используя день в качестве периода наблюдения. Мы записывали только те интервалы, когда была зафиксирована хотя бы одна крыса или кошка, чтобы учесть ложные срабатывания исследователей, ветра и птиц в дни, когда не наблюдалось никакой активности животных.
Мы также исследовали, влияет ли присутствие кошки, замеченной накануне, на присутствие крысы, замеченной на следующий день. Мы рассчитали период задержки для кошек, вычитая 1 день из времени поимки каждой крысы (например, мы рассчитали количество срабатываний на крыс в качестве зависимой переменной на графике и использовали период задержки для кошек в качестве предиктора). Мы также рассчитали период задержки для крыс, чтобы определить, предсказывает ли их присутствие на следующий день количество кошек.
Все видеозаписи были просмотрены и оценены вручную одним и тем же наблюдателем. В первую очередь мы отметили, что вход в колонию (убежище) находился на востоке (рис. 1 ) вдоль южной стены, а выход через засов (резервный вход, когда основной вход заблокирован) — на севере. Типичные источники пищи и воды, которые использовали животные, находились на западе, в виде мусорных куч. При оценке видеозаписей с кошками мы фиксировали все случаи восьми основных поведенческих реакций кошек (рис. 2 ): ходьба, бег, преследование, погоня (хищничество), обнюхивание, кормление, сидение, мечение территории (или чрезмерное мечение) и направление движения животного. Поскольку продолжительность этих видеозаписей варьировалась, некоторые длились всего несколько секунд, мы фиксировали только отдельные события, а не продолжительность поведения. При оценке видеозаписей с крысами (рис. 3 ) мы фиксировали все случаи ходьбы, бега, замирания, питья, кормления, сидения и преследования/хищничества. Аналогично, мы фиксировали направление движения животного. Данная работа выполнена в соответствии с рекомендациями Комитета по этике использования животных в научных исследованиях (IACUC) по этичному поведению при уходе за животными и их использовании в исследованиях (Fordham IACUC JMS 17-01).
Рисунок 2

Рисунок шерсти пяти диких кошек и последующая этограмма, использованная для оценки их поведения в полузакрытом промышленном здании в Нью-Йорке в период с 27 декабря 2017 года по 28 мая 2018 года. Оценка поведения кошек (модифицировано по данным Stanton et al.).
2015Включает ходьбу (w): движение вперед, при котором животное движется медленно. Бег (r): животное использует движение вперед быстрым шагом. Преследование (s): медленное движение вперед в присевшей позе, с низко опущенной головой и взглядом, сфокусированным на потенциальной добыче. Хищничество (p): охота или убийство, приводящие к прямой смерти. Обнюхивание (s): кошка обнюхивает пол или предмет, вдыхая воздух через нос. Кормление (f): животное проглатывает пищу, обычно с пола. Сидение (s): животное неподвижно в вертикальном положении. Метка (m): стоя с вертикально поднятым хвостом, кошка выпускает струю мочи назад на поверхность предмета. Хвост может дрожать при выделении мочи.
Рисунок 3
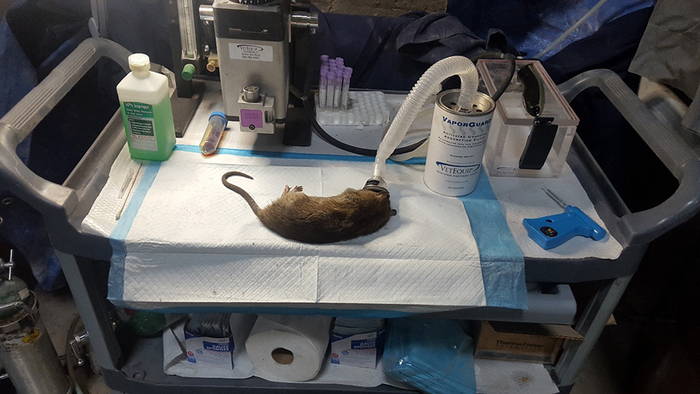
Для оценки поведения крыс в полузакрытом промышленном здании в Нью-Йорке с 27 декабря 2017 года по 28 мая 2018 года использовалось анестезиологическое оборудование и типичная крыса весом более 330 г, пойманная вместе с этограммой. Оценивались следующие поведенческие характеристики крыс: ходьба (w): движение вперед, когда животное движется медленно. Бег (r): движение вперед, когда животное движется быстрее, вытянув предплечья. Замирание/остановка (f): крысы прекращают активность и становятся неподвижными, обычно в ответ на предполагаемую угрозу. Питье (d): животное пьет воду или другой источник жидкости, обычно из луж. Обнюхивание (s): животное обнюхивает объект, вдыхая воздух через нос. Кормление (f): животное проглатывает частицы пищи, часто с поверхности пола. Сидение (s): животное находится в расслабленной позе, неподвижно, может стоять на задних лапах с вытянутыми предплечьями.
Статистика
Мы провели бинарную логистическую регрессию, чтобы оценить, связана ли доля срабатываний «крысы» с долей срабатываний «кошки» в течение дня, связана ли доля наблюдений кошек за предыдущий день с долей наблюдений крыс в текущий день, связана ли доля наблюдений крыс за предыдущий день с долей наблюдений кошек в текущий день, и связана ли доля срабатываний «люди» с долей крыс в течение дня. Мы также использовали бинарную логистическую регрессию для определения взаимосвязи между кошками и 8 распространенными типами поведения крыс (за исключением преследования/хищничества, которое наблюдалось только 3 раза) и направлением их передвижения. Статистическая значимость (альфа) была установлена на уровне P ≤ 0,05, и все описательные и инференциальные анализы проводились с использованием Minitab V. 17 (State College, PA).
Результаты
Всего было оценено 306 видеозаписей, сделанных в течение 79 дней. Пять отдельных кошек были идентифицированы на основе цвета, отметок и рисунка шрамов (Рисунок 2 ). Эти животные впервые появились 27 декабря и оставались на протяжении всего исследования. В пяти случаях три кошки появлялись в один и тот же день. Было три попытки нападения со стороны двух кошек (по одной успешной попытке) и одна неудачная попытка. Все пять кошек участвовали в событиях преследования (20).
В период присутствия кошек (с 27 декабря по 28 мая) количество кошек в конкретный день влияло на вероятность обнаружения крыс в тот же день (Рисунок 4 , ОР = 0,01; 95% ДИ [0,00, 0,02], p < 0,001). При каждом увеличении количества кошек на 1% в конкретный день вероятность срабатывания камеры в этот день уменьшается в 100 раз. Кроме того, количество кошек в предыдущий день обратно пропорционально предсказывало количество крыс ( ОР = 0,15; 95% ДИ [0,06, 0,34], p < 0,001). Каждое увеличение количества кошек, замеченных в предыдущий день, на 1% приводит к тому, что вероятность обнаружения крысы в этот день уменьшается в 6,6 раза. Количество крыс, замеченных в определенный день, не предсказывало количество кошек, замеченных на следующий день (ОР = 0,62, p > 0,1), равно как и количество людей, замеченных в определенный день, не влияло на вероятность обнаружения крыс в тот же день (ОР = 0,44, p > 0,1).
Рисунок 4
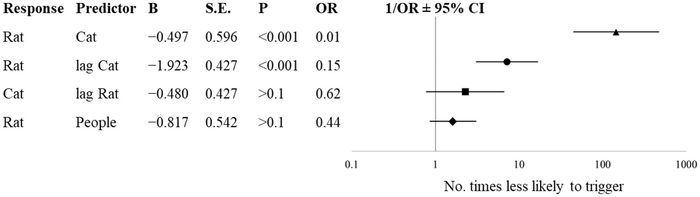
Диаграмма Фореста и оценки вероятности, полученные в результате бинарной логистической регрессии с различными предикторами в полузакрытом промышленном здании в Нью-Йорке в период с 27 декабря 2017 г. по 28 мая 2018 г. Задержка относится к наблюдениям в предыдущий день.
Поведение и использование пространства
Было зафиксировано 259 случаев поведения кошек (таблица 1 ). Среди этих случаев 46,3% (120) включали ходьбу в восточном или западном направлении, начинающуюся или заканчивающуюся возле входа в крысиный питомник. Два видео завершились тем, что голова кошки приблизилась к входу в крысиный питомник. Мы обнаружили, что 27,4% зафиксированных случаев поведения кошек (71) включали обнюхивание. Это поведение выполнялось по открытому пространству и включало два объекта, которые часто метились слишком сильно. Две кошки (рисунок 1 ) метили центральный объект на снимке (корпус RFID) и иногда садились на него, в то время как три другие кошки метили ведро рядом с входом в питомник. Было зафиксировано 20 попыток преследования и 3 попытки хищничества, хотя кошки и крысы появлялись в одном видео менее 10 раз. Две попытки нападения хищника оказались успешными, когда кошки вытащили животное из-под укрытия (рядом с меткой), единственная попытка нападения на открытой местности провалилась, когда крыса остановилась, а кошка также прекратила погоню, лишь уставившись на животное. Поскольку количество замеченных крыс в определенный день не предсказывало количество замеченных кошек на следующий день, мы не оценивали количество наблюдений крыс как предикторы какого-либо конкретного поведения кошек.
Таблица 1
Идентификатор кошкиЧерныйСерыйШрамыЧерно-белыйБелыйНеизвестныйОбщийПОВЕДЕНИЕХодьба1446621276120Бег2103208Преследование13295020Хищничество0101103Нюхание8452610071Кормление0601108Сидя3300208Маркировка73245021НАПРАВЛЕНИЕСевер131514226Юг2201117Восток8931414149Запад336454354
Зафиксированные случаи поведения кошек в полузакрытом промышленном здании в Нью-Йорке в период с 27 декабря 2017 года по 28 мая 2018 года.
Под направлением понимается перемещение животного, находящегося вне поля зрения .
С другой стороны, поскольку количество кошек действительно предсказывало появление крыс, мы изучили, как количество появлений кошек менялось в зависимости от индивидуального поведения крыс. Количество кошек в конкретный день влияло на количество крыс, движущихся в восточном направлении к своей колонии (Таблица 2 ; OR = 1,19; 95% CI [1,00, 1,41], p < 0,05). Ходьба, более консервативный показатель передвижения, чем потенциальная встреча с хищником, была единственным другим видом поведения, который положительно коррелировал с количеством появлений кошек.
Таблица 2
ПоведениеКоэф .ЮВЗПИЛИНижеВерхняяХодить0,1910,0882.170,0301.211.021.44Бегать−0,0600,086−0,700,4830,940,801.11Заморозить−0,1440,159−0,900,3660,870,631.18Напиток−17,465, 568.95−0.000,9970.000.00*Нюхнуть0,03020,0950,320,7511.030,861.24Кормить−17.5213, 990.70−0.000,9960.000.00*Сидеть−17.4665, 568.95−0.000,9970.000.00*Север0.1220.1071.140,2541.130,921.39Юг0.1600.1191.340.1811.170,931.48Восток0.1710,0861.980,0471.191.001.41Запад0,1760,0931.900,0581.190,991.43
Оценки вероятности в бинарной логистической регрессии с количеством наблюдений кошек в качестве предиктора поведения крыс в полузакрытом промышленном здании в Нью-Йорке в период с 27 декабря 2017 г. по 28 мая 2018 г.
Направление указывает на ориентацию, в которой движется крыса при выходе из видео. Жирным шрифтом выделено число кошек, повлиявших на это поведение при p < 0,05.
*
Обозначает верхний предел, слишком большой для отображения, например, > 1000 .
Обсуждение
Это исследование — первое, документирующее прямое и косвенное влияние идентифицируемых диких кошек на частично идентифицированную популяцию городских крыс. Эта частично огороженная городская экосистема находилась под сильным давлением кошек, о чем свидетельствует количество присутствующих кошек и количество попыток преследования со стороны каждой кошки на протяжении всего исследования. Эти события происходили в любое время дня и вечера. Тем не менее, из 259 случаев поведения, зафиксированных кошками за 79 дней/ночей, только 3 раза кошка преследовала крысу — и только один раз по открытой местности. Мы были удивлены, что в районе, где в течение одних и тех же 24 часов патрулировали до трех кошек, было так мало прямых попыток хищничества. Хотя полевые камеры часто имеют период задержки и не идеально фиксируют все виды поведения, камеры имели высокую скорость срабатывания затвора (0,4 с), а непрерывный характер погони обеспечивает высокую вероятность фиксации хотя бы части процесса на видео.
Поначалу нас удивило, что такой низкий уровень хищничества может привести к значительным изменениям в использовании пространства и времени активности. Коэффициенты вероятности были очень высокими при прогнозировании обнаружения крыс, когда кошка была замечена либо в течение данного дня ( OR = 0,01), либо в предыдущий день ( OR = 0,15). Мы могли бы ожидать, что крысы привыкнут к кошкам примерно так же, как, по-видимому, привыкли к посещению людей. Это несмотря на то, что люди регулярно отлавливали и выпускали крыс, а также когда велась их дезинсекция по распоряжению городских властей. Однако также хорошо известно, что все животные, особенно потенциальная добыча, переоценивают свои риски, используя «простые эмпирические правила» (Боускила и Блюмштейн,1992Лучше ошибочно предположить, что организм опасен (ложноположительный результат), чем предположить, что опасное животное безвредно (ложноотрицательный результат) (Джонсон и др.).2013Это происходит главным образом потому, что невосприимчивость к хищничеству может привести к окончательной потере приспособленности из-за хищничества (Боускила и Блюмштейн,1992).
Вероятно, два случая успешного хищничества произошли с участием более мелких особей, вес которых не превышал 250 г (диапазон, определенный Чайлдсом).1986К сожалению, у этих двух особей не было микрочипов, поэтому мы не смогли измерить их вес. Кроме того, оба животных были пойманы в укрытии рядом или под местами, где кошки обычно метят территорию. Таким образом, возможно, что крысы были больны или стали жертвами отравления бромадиолоном. Хотя исследователи имеют долгосрочный доступ к месту исследования, владельцы участка по закону обязаны продолжать использовать приманки, даже при поддержке научных исследований. Использование приманок является нормальной частью городских экосистем в городах по всему миру, и это необходимо учитывать при любых экологических исследованиях.
На первый взгляд казалось нелогичным, что скорость передвижения значительно и положительно зависит от количества замеченных кошек. Однако на рисунке 1 показано, что крысы могли либо переходить свою тропу, используя укрытие, двигаясь на восток, либо потенциально попадать в засаду. В этом контексте ходьба является более консервативным способом передвижения. Животные чаще двигались на восток во время интенсивного наблюдения за кошками, поскольку именно там находится основное убежище и вход в колонию (рисунок 1 ).
Мы были несколько разочарованы тем, что станция RFID и данные о ранее чипированных крысах оказались не более полезными для нашего исследования хищничества. Тем не менее, система RFID все же позволила получить некоторое представление о количестве животных, присутствующих в активной колонии, и показать, что их совокупный вес значительно превышает верхний предел в 250 г (Чайлдс,1986Если бы антенны были обмазаны аттрактантом, крысы, возможно, с большей вероятностью активировали бы свои идентификационные метки до того, как вызвать/выполнить определенное поведение. Так уж получилось, что мы как раз занимались оценкой предполагаемого отпугивающего запаха (Парсонс и др., неопубликованные данные).
Исходя из полученных результатов, мы можем лишь отметить, что сохраняющаяся путаница между крысами и мышами ( Mus musculus и Peromyscus spp .) может способствовать распространению некачественной, но рискованной (Woinarski et al.,2017) подход к борьбе с крысами. Очевидно, что, хотя крысы реже встречались в местах, где были установлены наши камеры (открытые пространства), они все же сохраняли свою популяцию в густонаселенной колонии. Их пространственное поведение указывает на то, что они просто перемещались в другие места в поисках пищи, не прибегая к попыткам выслеживания.
Хотя наши результаты очевидны, их следует воспроизвести в районах с нестабильным снабжением пищей. Учитывая высокую скорость размножения городских крыс (Дэвис,1951При достаточном количестве пищи одна беременная самка может родить от 6 до 10 пометов по 12 детенышей в год. В районах с меньшим количеством пищи, где, следовательно, крысы размножаются реже, возможно, прямое воздействие хищничества кошек может быть более значительным. Для проверки этого явления и дальнейшего изучения влияния кошек на смертность крыс необходимы дополнительные экспериментальные исследования с участием кошек (например, данные с присутствием и без присутствия кошек).
Given that small prey over-estimate risks from predators, further experimentation is also required as to how the presence of cats, or their scents, indirectly influences the feeding rate (Wernecke et al., 2016) and fecundity (Voznessenskaya, 2014) of rats, and whether this has any tangible benefit as an adjunct to rat control (Himsworth et al., 2013). Such information is essential if we are to understand whether the influence of feral cats on rats is remotely worth the risks to native urban wildlife. Our results at a waste recycling facility, however, suggest that city rats can persist in high density, simply by altering their movements, despite the presence of hunting cats.
Statements
Data availability statement
The authors hope our study will lead to future efforts to quantify the impact of cats on rats. Thus, our data will be made available free of charge on request.
Author contributions
MP designed the study and wrote the article. MP and MD collected all data. PB and JM-S assisted the writing and edits.
Funding
This work was funded by the National Pest Management Association.
Acknowledgments
This work was sponsored by the Pest Management Foundation, Fairfax, Virginia. In particular, we thank Jim Fredericks for his assistance throughout the project. We thank Tom Jordan of Arrow Exterminating Company, Inc for MD's time and collaboration. We thank Rudy Fulmer (Arrow) for his ongoing support in provided urban trapping and wildlife monitoring advice. We thank Matt Ruiter and Craig Jordan of UID Identification Solutions for designing the all-weather RFID (PADAR) system for identification of urban rats. Bob Schrock of Vet Equip assisted design of the mobile, anesthesiology unit. We thank Eddie Valentin, Site Manager, for Waste Management (WM) for being extremely helpful in accommodating this research in spite of obligations to ongoing extermination of rats.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
ApfelbachR.ParsonsM. H.SoiniH. A.NovotnyM. V. (2015). Are single odorous components of a predator sufficient to elicit defensive behaviors in prey species?Front. Neurosci.9:263. 10.3389/fnins.2015.00263
2
BouskilaA.BlumsteinD. T. (1992). Rules of thumb for predation hazard assessment: predictions from a dynamic model. Am. Nat.139, 161–176. 10.1086/285318
3
CartheyA. J.BanksP. B. (2018). Naïve, bold, or just hungry? An invasive exotic prey species recognises but does not respond to its predators. Biol. Invasions1–13. 10.1007/s10530-018-1782-4
4
ChildsJ. E. (1986). Size-dependent predation on rats (Rattus norvegicus) by house cats (Felis catus) in an urban setting. J. Mammal.67, 196–199. 10.2307/1381025
5
CombsM.PuckettE. E.RichardsonJ.MimsD.Munshi-SouthJ. (2018). Spatial population genomics of the brown rat (Rattus norvegicus) in New York City. Mol. Ecol.27, 83–98. 10.1111/mec.14437
6
DaviesH. F.McCarthyM. A.FirthR. S.WoinarskiJ. C.GillespieG. R.AndersenA. N.et al. (2017). Top-down control of species distributions: feral cats driving the regional extinction of a threatened rodent in northern Australia. Divers. Distrib.23, 272–283. 10.1111/ddi.12522
7
DavisD. E. (1951). A comparison of reproductive potential of two rat populations. Ecology32, 469–475. 10.2307/1931724
8
EmbarK.RavehA.BurnsD.KotlerB. P. (2014). To dare or not to dare? Risk management by owls in a predator–prey foraging game. Oecologia175, 825–834. 10.1007/s00442-014-2956-0
9
FitzgeraldB.KarlB.VeitchC. (1991). The diet of feral cats (Felis catus) on Raoul Island, Kermadec Group. New Zealand J. Ecol. 15, 123–129.
10
FrankA. S.JohnsonC. N.PottsJ. M.FisherA.LawesM. J.WoinarskiJ. C.et al. (2014). Experimental evidence that feral cats cause local extirpation of small mammals in Australia's tropical savannas. J. Appl. Ecol.51, 1486–1493. 10.1111/1365-2664.12323
11
GlantonD. (2017). Chicago's Rat Problem Gnaws at Feline Fears, Chicago Tribune. Chciago, IL.
12
GriffithD. A.WongD. W. (2007). Modeling population density across major US cities: a polycentric spatial regression approach. J. Geogr. Syst.9, 53–75. 10.1007/s10109-006-0032-y
13
HermanC. S.ValoneT. J. (2000). The effect of mammalian predator scent on the foraging behavior of Dipodomys merriami. Oikos91, 139–145. 10.1034/j.1600-0706.2000.910113.x
14
HimsworthC. G.FengA. Y.ParsonsK.KerrT.PatrickD. M. (2013). Using experiential knowledge to understand urban rat ecology: a survey of Canadian pest control professionals. Urban Ecosyst.16, 341–350. 10.1007/s11252-012-0261-4
15
HurstJ. L.BeynonR. J. (2004). Scent wars: the chemobiology of competitive signalling in mice. Bioessays26, 1288–1298. 10.1002/bies.20147
16
JeschkeJ. M.KoppM.TollrianR. (2002). Predator functional responses: discriminating between handling and digesting prey. Ecol. Monogr.72, 95–112. 10.1890/0012-9615(2002)072[0095:PFRDBH]2.0.CO;2
17
JohnsonD. D.BlumsteinD. T.FowlerJ. H.HaseltonM. G. (2013). The evolution of error: Error management, cognitive constraints, and adaptive decision-making biases. Trends Ecol. Evol.28, 474–481. 10.1016/j.tree.2013.05.014
18
KarlB. J.BestH. A. (1982). Feral cats on Stewart Island; their foods, and their effects on kakapo. New Zealand J. Zool.9, 287–293. 10.1080/03014223.1982.10423857
19
KauhalaK.TalvitieK.VuorisaloT. (2015). Free-ranging house cats in urban and rural areas in the north: useful rodent killers or harmful bird predators?Folia Zool.64, 45–55. 10.25225/fozo.v64.i1.a6.2015
20
KelliherK. R.WersingerS. R. (2009). Olfactory regulation of the sexual behavior and reproductive physiology of the laboratory mouse: effects and neural mechanisms. ILAR J.50, 28–42. 10.1093/ilar.50.1.28
21
KikillusK. H.ChambersG. K.FarnworthM. J.HareK. M. (2017). Research challenges and conservation implications for urban cat management in New Zealand. Pac. Conserv. Biol.23, 15–24. 10.1071/PC16022
22
LossS. R.MarraP. P. (2017). Population impacts of free-ranging domestic cats on mainland vertebrates. Front. Ecol. Environ. 15:1633. 10.1002/fee.1633
23
LossS. R.WillT.MarraP. P. (2013). The impact of free-ranging domestic cats on wildlife of the United States. Nat. Commun.4:1396. 10.1038/ncomms2380
24
MontagueM. J.LiG.GandolfiB.KhanR.AkenB. L.SearleS. M.et al. (2014). Comparative analysis of the domestic cat genome reveals genetic signatures underlying feline biology and domestication. Proc. Natl. Acad. Sci. U.S.A.111, 17230–17235. 10.1073/pnas.1410083111
25
NaidenkoS. V.NaidenkoS. V.ClarkL.VoznessenskayaV. V. (2003). Predator Presence Affects the Reproductive Success of Prey in Outdoor Conditions.Lincoln, NE: USDA
26
OrrockJ. L.DanielsonB. J. (2009). Temperature and cloud cover, but not predator urine, affect winter foraging of mice. Ethology115, 641–648. 10.1111/j.1439-0310.2009.01654.x
27
OrrockJ. L.DanielsonB. J.BrinkerhoffR. J. (2004). Rodent foraging is affected by indirect, but not by direct, cues of predation risk. Behav. Ecol.15, 433–437. 10.1093/beheco/arh031
28
ParsonsM. H.BanksP. B.DeutschM. A.CorriganR. F.Munshi-SouthJ. (2017). Trends in urban rat ecology: a framework to define the prevailing knowledge gaps and incentives for academia, pest management professionals (PMPs) and public health agencies to participate. J. Urban Ecology3:jux005. 10.1093/jue/jux005
29
ParsonsM. H.BlumsteinD. T. (2010). Feeling vulnerable? Indirect risk cues differently influence how two marsupials respond to novel dingo urine. Ethology116, 972–980. 10.1111/j.1439-0310.2010.01810.x
30
ParsonsM. H.SarnoR.DeutschM. (2015). Jump-starting urban rat research: conspecific pheromones recruit wild rats into a behavioral and pathogen-monitoring assay. Front. Ecol. Evol.3:146. 10.3389/fevo.2015.00146
31
ParsonsM. H.SarnoR. J.DeutschM. A. (2016). A detailed protocol to enable safe-handling, preemptive detection, and systematic surveillance of rat-vectored pathogens in the urban environment. Front. Public Health4:132. 10.3389/fpubh.2016.00132
32
PillayK. R.StreicherJ.DownsC. T. (2018). Home range and habitat use of feral cats in an urban mosaic in Pietermaritzburg, KwaZulu-Natal, South Africa. Urban Ecosyst.1–11. 10.1007/s11252-018-0766-6
33
PuckettE. E.ParkJ.CombsM.BlumM. J.BryantJ. E.CacconeA.et al. (2016). Global population divergence and admixture of the brown rat (Rattus norvegicus). Proc. R. Soc. B283:20161762. 10.1098/rspb.2016.1762
34
RozziR.LomolinoM. V. (2017). Rapid dwarfing of an insular mammal–the feral cattle of amsterdam island. Sci. Rep.7:8820. 10.1038/s41598-017-08820-2
35
Schmidt-NielsenB.O'DellR. (1961). Structure and concentrating mechanism in the mammalian kidney. Am. J. Physiol.200, 1119–1124. 10.1152/ajplegacy.1961.200.6.1119
36
SiegelP. S.StuckeyH. L. (1947). The diurnal course of water and food intake in the normal mature rat. J. Comparative Physiol. Psychol.40:365. 10.1037/h0062185
37
StantonL. A.SullivanM. S.FazioJ. M. (2015). A standardized ethogram for the felidae: a tool for behavioral researchers. Appl. Anim. Behav. Sci.173, 3–16. 10.1016/j.applanim.2015.04.001
38
TakácsS.GriesR.ZhaiH.GriesG. (2016). The sex attractant pheromone of male brown rats: identification and field experiment. Angew. Chem. Int. Edn.55, 6062–6066. 10.1002/anie.201511864
39
Themb'alilahlwaA.MonadjemA.McCleeryR.BelmainS. R. (2017). Domestic cats and dogs create a landscape of fear for pest rodents around rural homesteads. PLoS ONE12:e0171593. 10.1371/journal.pone.0171593
40
VoznessenskayaV. V. (2014). Influence of cat odor on reproductive behavior and physiology in the house mouse (Mus musculus). Neurobiol. Chem. Commun. 389–406. 10.1201/b16511-15
41
VoznessenskayaV. V.NaidenkoS. V.FeoktistovaN. Y.KrivomazovG. J.MillerL. A.ClarkL. (2003). Predator Odours as Reproductive Inhibitors for Norway Rats. USDA National Wildlife Research Center-Staff Publications. Boca Raton, FL: CRC Press.
42
WerneckeK. E.BrüggemannJ.FendtM. (2016). Predator odor exposure increases food-carrying behavior in rats. Physiol. Behav.154, 15–19. 10.1016/j.physbeh.2015.11.005
43
WoinarskiJ.MurphyB.LeggeS.GarnettS.LawesM.ComerS.et al. (2017). How many birds are killed by cats in Australia?Biol. Conserv.214, 76–87. 10.1016/j.biocon.2017.08.006
44
YipS. J.RichM.-A.DickmanC. R. (2015). Diet of the feral cat, Felis catus, in central Australian grassland habitats during population cycles of its principal prey. Mammal Res.60, 39–50. 10.1007/s13364-014-0208-7
45
ZhouN.FridleyD.KhannaN. Z.KeJ.McNeilM.LevineM. (2013). China's energy and emissions outlook to 2050: Perspectives from bottom-up energy end-use model. Energy Policy53, 51–62. 10.1016/j.enpol.2012.09.065
46
ZielinskiW. J.vom SaalF. S.VandenberghJ. G. (1992). The effect of intrauterine position on the survival, reproduction and home range size of female house mice (Mus musculus). Behav. Ecol. Sociobiol.30, 185–191. 10.1007/BF00166702
Summary
Keywords
city rats, Felis catus, New York city rat, Norway rats, Rattus norvegicus, urban wildlife interactions
Citation
Parsons MH, Banks PB, Deutsch MA and Munshi-South J (2018) Temporal and Space-Use Changes by Rats in Response to Predation by Feral Cats in an Urban Ecosystem. Front. Ecol. Evol. 6:146. doi: 10.3389/fevo.2018.00146
Received
23 June 2018
Accepted
04 September 2018
Published
27 September 2018
Volume
6 - 2018
Edited by
G. Darrel Jenerette, University of California, Riverside, United States
Reviewed by
Rafal Stryjek, Institute of Psychology (PAN), Poland; Adam James Bates, Nottingham Trent University, United Kingdom
Updates
Check for updates
Copyright
© 2018 Parsons, Banks, Deutsch and Munshi-South.
Ну как хорошо ловят крыс?
