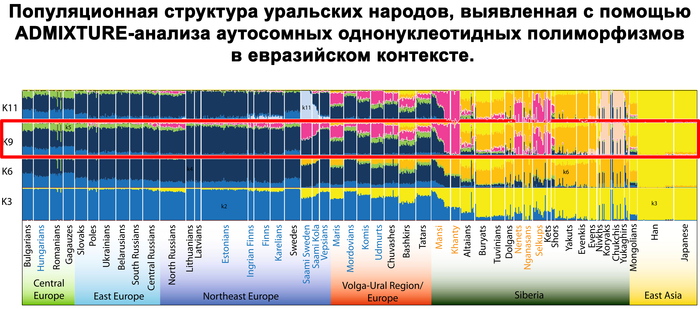
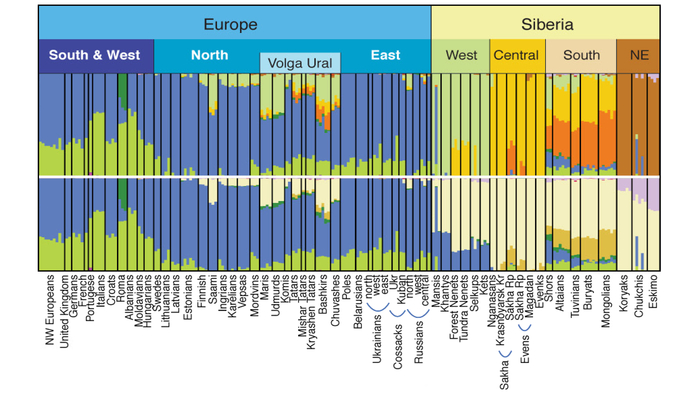
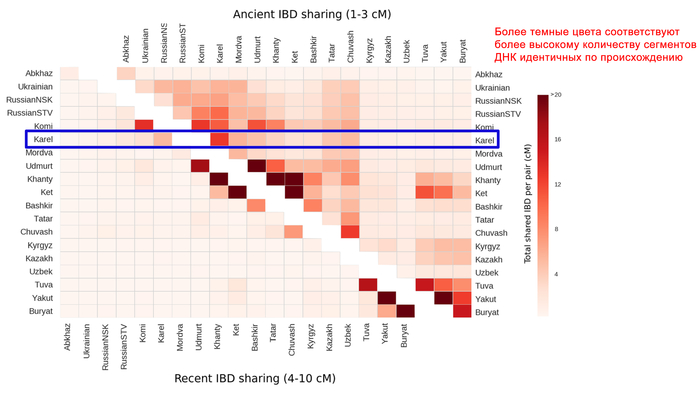
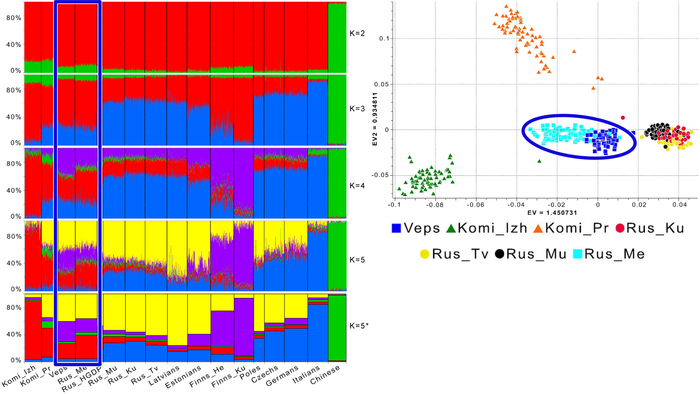
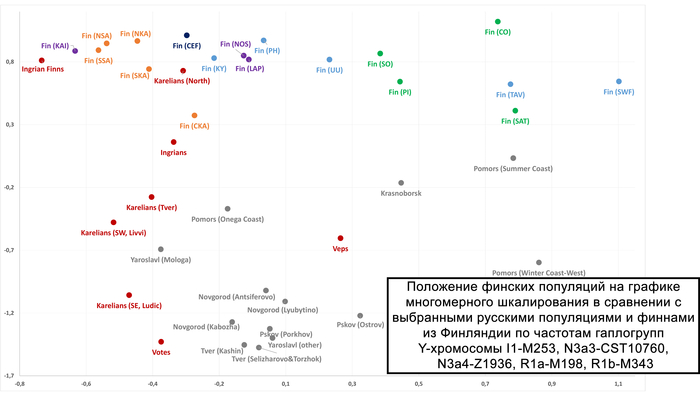
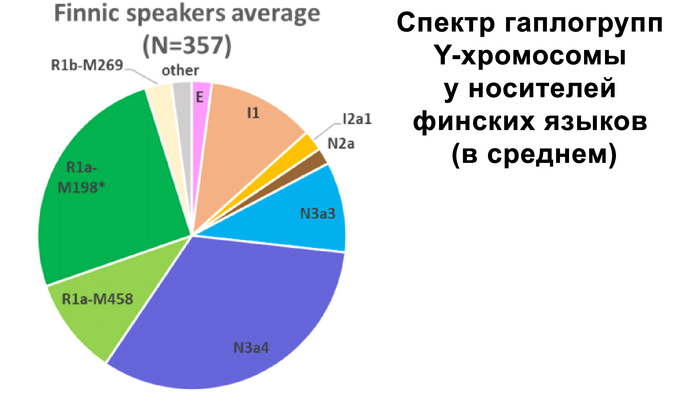
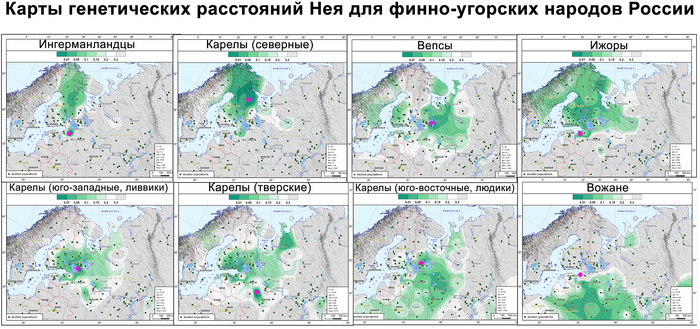
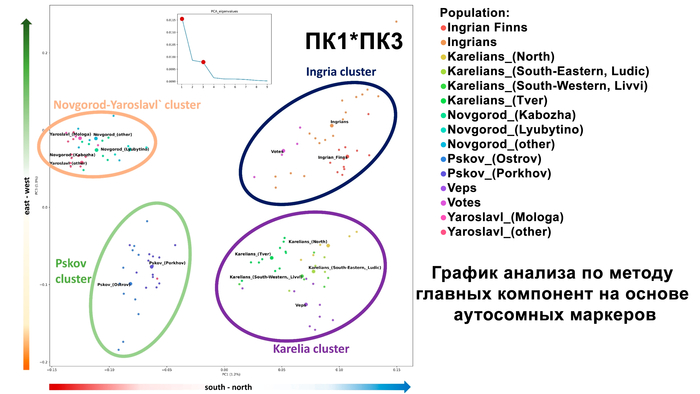
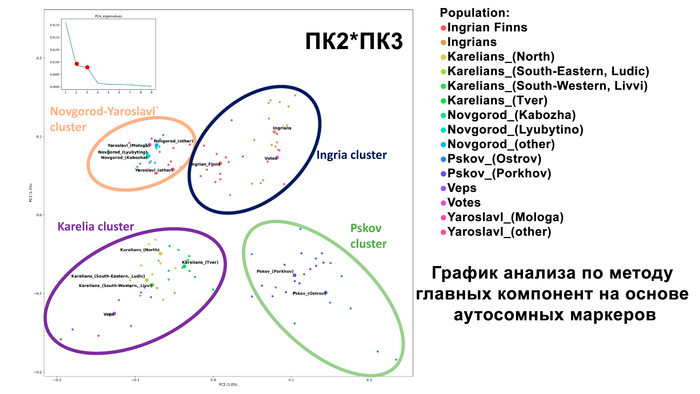
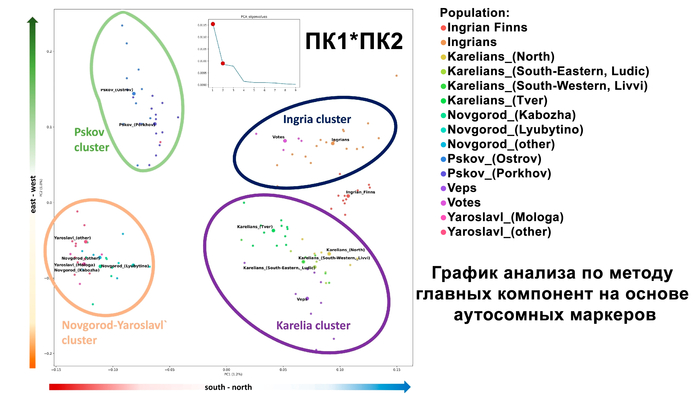
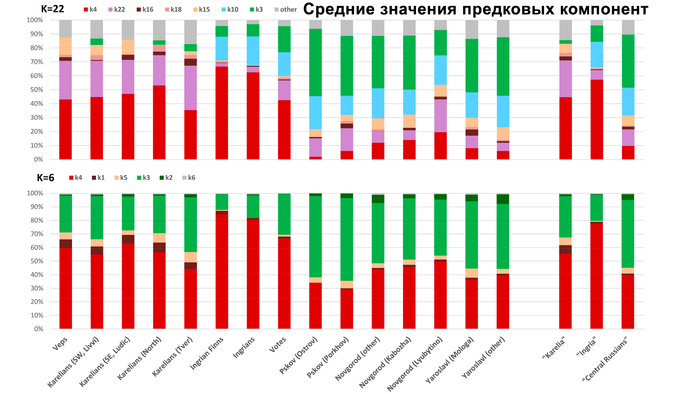
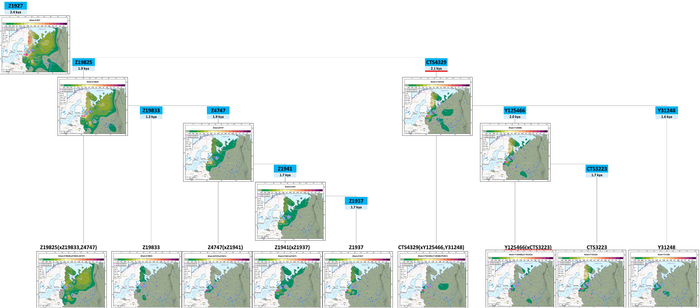
Зорич -сын ПриРоды
Тварь я дрожащая или право имею? Природа уже знает ответ.
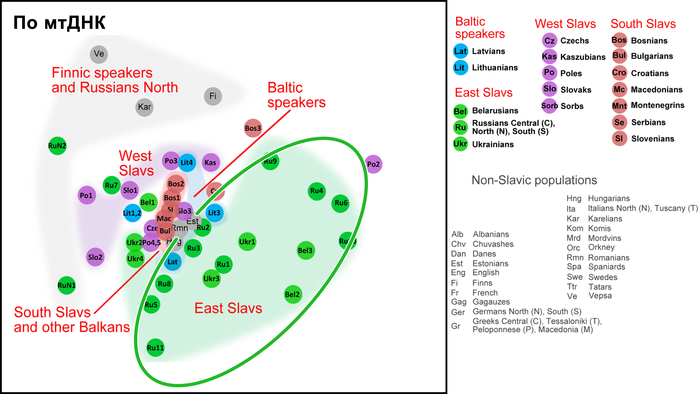
Её прозвали «Чёрным Кашлем». Вирус, вырвавшийся из лабораторных глубин павшего мира, не оставлял шансов. Люди гибли, захлёбываясь собственной кровью, их лёгкие отказывали за считанные дни. Цивилизация рухнула, скрытая чёрной пеленой смерти. Отдалённая Вепская земля с её древними лесами и забытыми деревнями стала одним из последних островков жизни. Но волна смерти докатилась и сюда.
Часть 1: Тихое проклятие
Сначала Зорич, страж Урочища, заметил лишь неестественную тишину. Птицы умолкли, звери ушли вглубь чащи. Потом с предгорий потянуло смрадом горящих тел.
Она не вошла, а рухнула на порог его жилища, подкошенная страшной хворью, словно тростник под ударом топора. Её тело, ещё недавно сильное и ловкое, безвольно распласталось на поросшем мхом полу. Голова с влажными от пота волнами тёмных волос откинулась, обнажив тонкую, бледную шею, где уже проступали зловещие тёмные прожилки. Красота её была подобна подрубленному дубу — величественной даже в падении. Черты лица, отточенные и ясные, казались высеченными из мрамора, но мрамора больного, с тем самым «багряным чумным румянцем», что пылал на щеках алым стягом смерти. Длинные ресницы, мокрые от испарины, лежали на синеватых тенях под глазами. Из полуоткрытых, потрескавшихся губ вырывался хриплый, прерывистый звук — не крик, а предсмертный хрип, который был страшнее любого вопля. Её пальцы, сильные и умелые, которыми она ещё вчера, возможно, плела кужель, судорожно впились в мох, будто ища опоры в ускользающем мире. От неё пахло не жизнью — не хлебом и дымом, а смертельным смрадом хвори — смесью горящего железа и увядшей полыни.— Чёрный… Кашель… — выдохнула она, падая. Увидев Богдану на пороге, Зорич на мгновение окаменел. Затем его сознание заработало с чёткостью боевого механизма. Он не стал переносить её далеко, чтобы не трясти в лихорадке, а бережно перетащил в сторону, на ложе из мягких шкур и высушенного мха. Он не сразу начал поить её снадобьями. Сперва Зорич развёл прямо у входа малый очистительный костёр из можжевельника и бросил в него щепотку сушёного зверобоя. Зорич и Богдана не были чужими. За годы до Чащерного Урочища и Чёрного Кашля их судьбы уже пересекались на пыльной улице маленькой вепской деревни. Они сидели за одной партой в начальной школе. Он — замкнутый мальчик, чьи взгляды постоянно ускользали через окно в сторону леса. Она — живая, яркая девочка с смехом, который звенел на всю классную комнату, дочь уважаемых в деревне людей. Её отец был кузнецом, чья кузница стояла на отшибе. Зорич, тогда ещё просто мальчишка, часто завороженно наблюдал, как тот, могучий и потный дядька, рождает из огня и металла полезные вещи — подковы, топоры, ножи. А мать Богданы трудилась на ткацкой фабрике в соседнем посёлке. Она приносила домой яркие обрезки ткани, из которых Богдана шила наряды своим куклам. Они были из разных миров: он — из мира тишины и тайн леса, она — из мира огня, металла и грохота станков. И вот теперь их миры столкнулись вновь. На пороге его убежища лежала не просто красивая незнакомка, а девочка из его прошлого, чей отец когда-то выковал его первый охотничий нож. Это знание придавало его действиям новую силу — это был долг. Долг перед памятью о общем детстве, перед её родителями, чей труд был так же честен и суров, как труд его деда.
«Дымом-стеной, огнём-щитом, Обходи хворь стороной. Не для тебя этот порог, Не для тебя этот урок.» Шептал он заговоры, окуривая помещение, он создавал невидимый барьер, отсекая «дух болезни» от её источника. Он знал, что «Чёрный Кашель» пожирает лёгкие изнутри, вызывая гниение. Нужно было остановить это. Он взял добрую горсть серо-зелёной Уснеи Бородатой, этого лесного антибиотика, и бросил в слабый раствор горячего спирта. Пока он настаивался, Зорич растирал в ступе сухую Цетрарию (Исландский мох) в мелкую пыль. Он будет нужен позже, чтобы смягчить и заживить. Когда настой Уснеи остыл до тёплого, Зорич бережно приподнял голову Богданы и ложкой влил ей глоток целительного зелья.
«Уснея-борода, сила твоя крепка. Пронзи хворь густую, как туман,
Выжги скверну из белых костей. Верни лёгким дыхание, а телу — покой.».
Он вливал в неё не просто настой , а своё намерение, заклинание, обращённое к духу травы. Он видел не просто больную девушку а ту самую девочку, которая когда-то смеялась на школьном дворе. Его заговоры стали более личными. Шепча «Цепляйся за этот свет, Богдана», он мог мысленно добавлять: «Помнишь, как мы бежали с уроков? Помнишь запах металла из кузницы твоего отца? Вот за это и цепляйся». Его решимость укрепилась. Он не мог позволить болезни отнять у мира дочь кузнеца и ткачихи. Спасти её значило спасти частицу того старого, нормального мира, который они когда-то знали. Лихорадка сводила Богдану с ума. Зорич развёл в миске густую пасту из растёртой коры осины (природный аналог аспирина) Лобазника и прохладной глины. Этой массой он обмазал её виски, запястья и ступни — места, где кровь течёт близко к коже, — чтобы «вытянуть огонь». Он готовил сложное снадобье, «Чай четырёх сил» Зорич действовал как часовой, отмеряющий каждый шаг ритуала. Свет от пробивающегося сквозь хвою утреннего солнца падал на его неподвижную фигуру у низкого очага. Он расстелил на скамье чистый холст и разложил на нем четыре берестяных туеска. Открывая каждый, он нашептывал его силу: Из первого туеска он извлек Пармелию — серые, кружевные ломтики лишайника. «Сила Огня. Жги заразу», — прошептал он, отмеряя три щепоти в глиняную чашу-мешалку. Из второго посыпались мелкие листочки Чабреца. «Сила Ветра. Гони хворобу из костей». Две щепоти. Из третьего — темно-зеленые, сморщенные листья Малины. «Сила Воды. Уйми внутренний пожар». Две щепоти. Из четвертого — соцветия Таволги, пахнущие медом и горьким миндалем. «Сила Земли. Усмири боль, даруй покой». Две щепоти. Он не смешивал их, а лишь соединил в одной чаше, позволив им познакомиться. В небольшой, почерневший от дыма медный котелок он налил ровно два полных ковша родниковой воды — 400 мл. Вода плескалась, подхватывая первый солнечный луч. «Вода-сестра, стань проводником, донеси силу до каждой клети, до каждой жилки. Котелок был поставлен на треногу над уже подготовленными углями — без открытого пламени, но с ровным жаром. Зорич сел на корточки и не сводил глаз с воды, замерший в ожидании. Он ловил момент «белого ключа» — когда со дна начинают подниматься первые пузыри, похожие на жемчужные россыпи, а на поверхности вода мутнеет, но еще не кипит. Это был знак. Одним движением он опрокинул в котелок всю смесь трав. Шипения не было — был лишь глухой звук и мгновенное распространение густого, горьковато-сладкого аромата, который ударил в нос, как боевой клич. Он тут же снял котелок с огня, накрыл его деревянной крышкой и, не медля, укутал его в свой старый, пропахший дымом и травами плащ, словно укачивая младенца. Образовался теплый кокон. Зорич отнес его в самый темный угол жилища и поставил на дубовый пень.
«Теперь ваша воля. Четыре духа, сойдитесь воедино. Дважды по девять и еще девять дыханий.» Он сел рядом, в позу сторожа, и погрузился в полную неподвижность, следя за внутренними часами. Ровно 27 минут он не шевелился, лишь его губы беззвучно шептали заговор, вплетая его в самую суть настоя. Когда время истекло, он развернул плащ. Крышку он снял медленно, выпуская наружу концентрированный, живительный дух. Настой имел цвет темного янтаря, а на его поверхности переливалась радужная пленка эфирных масел. Он взял лоскут чистого, грубого льна и процедил жидкость в другую глиняную чашу. Травяная гуща осталась на ткани — свою работу она сделала. В чаше теперь плескалось снадобье «Чай четырёх сил» — не просто отвар, а квинтэссенция воли целителя и духов леса, готовая вступить в бой за жизнь Богданы. Оно было горьким, терпким, но за этой горечью скрывался аромат хвойного леса, летнего луга и надежды. Ночь стала самым тяжёлым временем. Богдану бил такой кашель, что казалось, её лёгкие вот-вот разорвутся. Зорич поставил рядом с ней плоский камень, накалил на огне докрасна другой камень-валунчик и капал на него отвар чаги, смешанный с пихтовой смолой. Густой, едкий, целебный пар поднимался к потолку. Он заставлял Богдану дышать этим дымом, пробивая заторы в её лёгких. Сидя у её изголовья, он не спал. Он клал руку на её горячий лоб и тихо, монотонно, как шаман, читал главный заговор — заговор на волю к жизни: Это превращало лечение из акта милосердия в акт верности памяти и искупления. Он спасал не просто жизнь — он спасал живое напоминание о своей собственной, другой, человеческой жизни.
Первые лучи солнца, бледные и осторожные, пробились в жилище Зорича, словно пытаясь разглядеть — кто одержал верх в ночной битве. Воздух внутри был тяжёлым, пропитанным запахом целебных дымов, пота и горьких трав. Над остывшим очагом всё ещё висела лёгкая дымка — призрак вчерашнего сражения. Богдана лежала на своём ложе из шкур. Страшный, багряный румянец, пылавший на её щеках, уступил место мертвенной, но уже природной бледности. Синева под её глазами не исчезла, но стала менее тёмной, как тучи после прошедшей бури. Самое главное — её дыхание изменилось. Оно больше не было тем хриплым, рвущимся на части предсмертным свистом. Теперь это были глубокие, ровные, хоть и ослабленные вздохи. Грудь поднималась и опускалась спокойно, без той судорожности, что терзала её всю ночь. Она спала. Но это был не тот тяжёлый, безпамятственный сон, в который она провалилась накануне. Теперь это был целительный покой, время, данное телу на восстановление. Иногда её веки чуть вздрагивали, а пальцы шевелились — признаки того, что сознание потихоньку возвращается из глубин. Зорич сидел у её изголовья, в той же позе, в которой провёл всю ночь. Его спина была прямой, но плечи выдали глубочайшую усталость. Тени под его глазами были густыми, как чернила. Одна его рука всё ещё лежала на шкуре рядом с её головой, будто он и во сне продолжал охранять её покой. Рядом стояла почти полная чаша остывшего «Чая четырёх сил» — свидетельство того, что в самые тяжёлые часы ночи он без устали поил её, капля за каплей вливая в неё жизнь. На столе лежали остатки глиняной пасты от жара, а в воздухе витал лёгкий, едва уловимый аромат исландского мха — следующее лекарство, уже приготовленное и ждущее своего часа. Когда солнце поднялось выше и луч упал прямо на лицо Богданы, она слабо зашевелилась. Не проснулась, а лишь глубже вдохнула, повернув голову, уходя от света. И на её лице не было гримасы боли — лишь умиротворение уставшего, но живого человека. В этот миг Зорич медленно выдохнул. Он не улыбнулся. Не было триумфа. Было лишь молчаливое, выстраданное облегчение. Он откинулся спиной на теплую стену своей сосны-крепости и впервые за много часов позволил векам сомкнуться. Битва была выиграна. Путь к выздоровлению — только начат. Но самое страшное осталось позади. Тишина утра была уже не зловещей, а мирной, и её нарушал лишь ровный, живой звук дыхания Богданы.
— Никого… не осталось… Бежать было некуда. Уроки Горазда и Радмилы стали его щитом: «Твой долг — беречь жизнь, какая есть». Зорич объявил войну невидимому врагу Он силой и угрозой заставил оставшихся в живых жить поодиночке в избах на окраинах. Между домами натянул верёвки с колокольчиками — для передачи еды и снадобий без контакта. На всех подходах к Урочищу развесил связки резко пахнущих трав — полыни, пижмы, можжевельника — как психологический и возможный физический барьер для чужаков. В его жилище закипела работа по старым рецептам. Совершал ежедневные обходы границ, взывая к духам леса о защите. Жёг ночами костры из можжевельника и шалфея, окутывая долину очищающим дымом. Не всех удалось спасти. Каждую потерянную жизнь Зорич воспринимал как личное поражение. Тех, кого не удавалось спасти, Зорич не хоронил в земле. Памятуя о стремительности и заразности Чёрного Кашля, он совершал очистительное сожжение на специальном погребальном костре, сложенном особым образом из осиновых ветвей — дерева, считающегося чистым и отпугивающим нечисть. Этот обряд был не актом жестокости, а суровой необходимостью и последней мерой защиты живых, чтобы болезнь не продолжила своё шествие по земле. Он провожал каждого словами: «Огонь, прими плоть. Ветер, развей пепел. Земля, очистись. Дух, иди к своим.» Когда эпидемия отступила, Урочище стало убежищем для горстки выживших. Зорич, поседевший за эти месяцы, продолжил свой дозор. Он понимал: его миссия не в победе над смертью, а в сохранении жизни — любой ценой. И пока в Чащерном Урочище теплился огонь в очаге, надежда для этого мира была жива.
Когда последний погребальный костер угас, а смрад смерти был окончательно вытеснен горьковатым дымом полыни и можжевельника, в Чарном Урочище воцарилась хрупкая, выстраданная тишина. Горстка выживших, спасенная ценою седины на висках Зорича и десятков безмолвных ночных дозоров, робко начала возвращаться к жизни. Они уже смотрели на него не как на чудаковатого отшельника, а как на Стража. На ту непоколебимую скалу, о которую разбилась сама Смерть. Однажды вечером, стоя на той самой поляне, где когда-то он призывал духов Исподни против браконьеров, Зорич смотрел на просыпающиеся звезды. В его памяти всплывали все те, кого он не смог спасти. Их лица, их последние хрипы, холодный пепел от их погребальных костров. Он вспомнил суровые уроки Горазда: «Сила не в мышцах, а в выдержке. Дерево гнётся, но не ломается, потому что умеет слушать ветер». И тихие наставления Радмилы: «Каждая травинка — это буква в великой книге исцеления. Надо только суметь её прочитать». Вся его жизнь — это диалог. Диалог с лесом, с духами, с болезнью, со смертью. Он не приказывал стихиям, он договаривался. Не покорял природу, а слушал её. И в этом слушании рождалась сила, перед которой отступил «Чёрный Кашель». Он не спас всех. Но он спас жизнь. Саму возможность жизни в этом Урочище. Он взял на себя тяжелейшее бремя выбора — кого изолировать, кому отдать последнюю горсть целебных кореньев, кого предать очищающему огню, чтобы спасти остальных. И тогда, в безлунную ночь, глядя на тлеющие угольки своего костра, Зорич нашел ответ на вопрос, отголосок которого преследовал его с тех пор, как он осознал тяжесть своей власти над жизнью и смертью.
«Тварь я дрожащая или право имею?»
Тишина Урочища, вся природа вокруг него, уже знала ответ. Он не был дрожащей тварью, ибо дрожащий не выстоял бы против чумы. Но он и не присвоил себе право, не вырвал его у мира силой. Это право было дано ему. Дедом, учившим слушать молчание. Бабкой, показавшей язык трав. Самим лесом, признавшим в нем своего голос и свою защиту. Право иметь — это не право брать. Это право отвечать. Отвечать за вверенный тебе клочок земли, за каждую жизнь в нем — от исполинской сосны до заблудившегося в чаще человека. Это не право на тиранию, а тяжелейшая обязанность хранителя, которую он добровольно взвалил на свои плечи.
И Зорич, Страж Чарного Урочища, сын приРоды понял это. Он не имел права распоряжаться жизнями по своей прихоти. Но он имел право — нет, был ОБЯЗАН — защищать великий Круговорот, даже если цена этой защиты — седина в волосах, шрамы на душе и вечное бремя одинокого решения. Он поднял голову. Звезды холодно сияли в черной вышине. Где-то в чаще пел свой последний перед сном напев дрозд. Воздух был чист и свеж. Природа уже знала ответ. И он тоже.