Популяционная генетика послеледниковой Евразии.
2 поста
Современные прибалтийско-финские народы населяют Северо-Восточную Европу и представляют западную ветвь уральской языковой семьи. В частности, финны, эстонцы и небольшая этническая группа ливвиков в основном проживают в Финляндии, Эстонии и Латвии соответственно. А карелы, вепсы, ижоры, вожане и ингерманландцы проживают на северо-западе России, в том числе в Республике Карелия, а также в Ленинградской, Архангельской, Вологодской и Тверской областях.
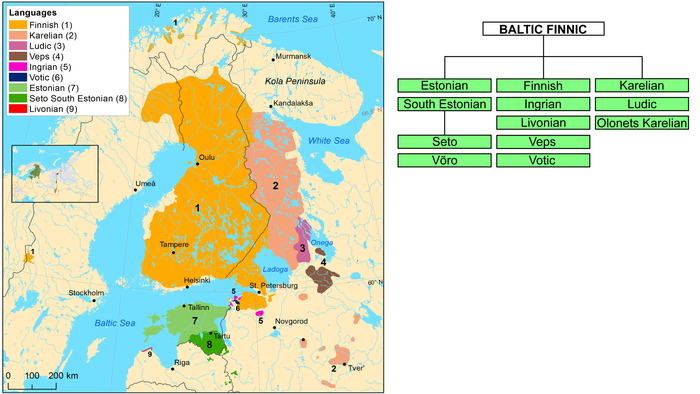
Согласно переписи населения России 2020 года, на долю прибалтийско-финских народов в России приходится менее 1% всех носителей финно-угорских языков, однако они занимают не менее трети территории, на которой говорят на финно-угорских языках.
История западных (или прибалтийских) финских народов тесно связана с Северо-Восточной Европой. Как языковое сообщество, протофинноязычные группы сформировались около 3000 лет назад.
К началу бронзового века генофонд на юге Северо-Восточной Европы (в регионе Восточной Балтики) претерпел значительные структурные изменения. Вклад мезолитических восточных охотников-собирателей (EHG) превзошёл вклад западных охотников-собирателей (WHG).
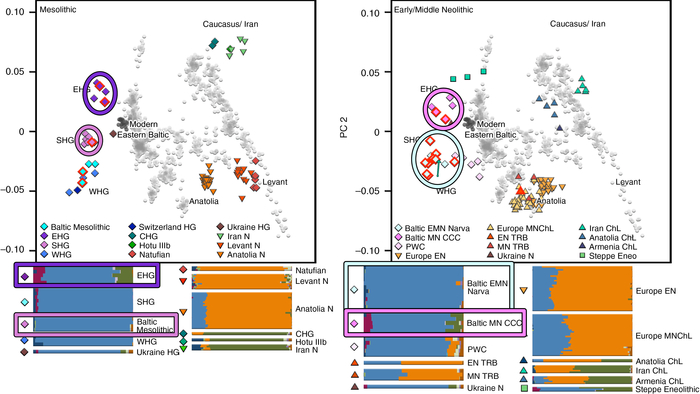
Кроме того, новые генетические компоненты, характерные для носителей культуры шнуровой керамики, были привнесены в результате миграций из Причерноморско-Каспийской степи, которые привели к переходу к земледелию и животноводству, а также к распространению индоевропейских языков в регионе.
Прежде чем продолжить, сразу отмечу, что названия гаплогрупп меняются по мере увеличения количества образцов и корректировки филогении. Значительную роль в изменении названий играет Международное сообщество генетической генеалогии (ISOGG). Поэтому в комментариях можно встретить возмущённые замечания от людей, которые запомнили обозначения гаплогрупп из более ранних работ. В данной работе также использованы более ранние обозначения гаплогрупп. Прошу это учитывать и ориентироваться на цифровые и буквенные обозначения после указания основной линии. Постарался адаптировать текст, чтобы не было путаницы.

Кроме того, люди часто не понимают смысл гаплогрупп и наделяют их какими-то удивительными свойствами. А в погоне за упрощением происхождения их носителей отталкиваются от места возникновения первой мутации, которую учёные условно назвали новой буквой, и пытаются поместить прародину популяций, у которых эта линия встречается, не в место формирования генофонда популяции, а в место, где обнаружены первые образцы с этой условно-названной линией. Хотя эта линия всего лишь продолжение предыдущей, а её обозначение весьма условно. Часто такое встречаю в комментариях касательно гаплогруппы R, которая также, как и N, является продолжением гаплогруппы K, и так далее в прошлое, вплоть до A, т. е. в начало филогении Y-хромосомы человека.
Но продолжим…
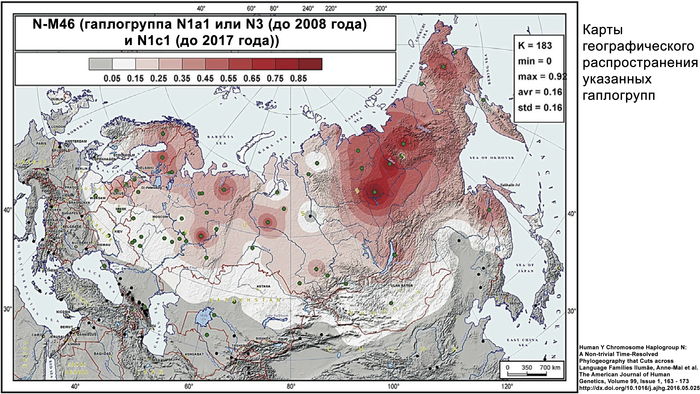
Самые ранние генетические следы миграций из Западной Сибири были обнаружены на севере Кольского полуострова.
Эти следы были выявлены как по аутосомным данным (сибирская компонента), так и по данным Y-хромосомы N-M46 (гаплогруппа N1a1 или N3 (до 2008 года) и N1c1 (до 2017 года)). Этот период относится примерно к 3,5 тысячам лет назад и совпадает с распространением носителей уральских языков. До 2000 лет назад эта волна миграции достигла Скандинавии, где её влияние прослеживается от эпохи до викингов (1–749 гг. н. э.) и до наших дней. Интересно, но в образцах из Восточной Прибалтики бронзового века не было обнаружено сибирского вклада в генетику населения.
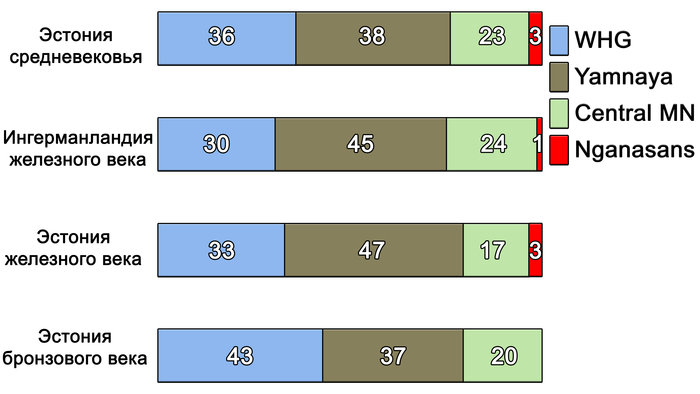
Однако в железном веке, не позднее 2,5 тысяч лет назад, когда финно-угорские языки начали активно разделяться, у представителей культуры каменных могильников на территории современной Эстонии были выявлены сибирская аутосомная компонента и гаплогруппа Y-хромосомы N-TAT или M46. Эта культура характеризуется каменными могильниками с оградками – тарандами и может упоминаться как культура тарандов.
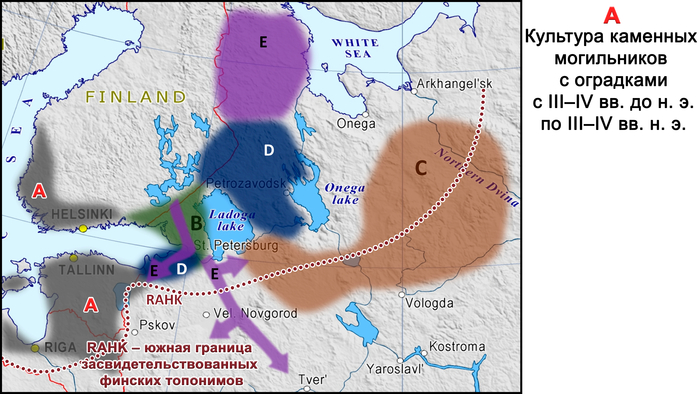
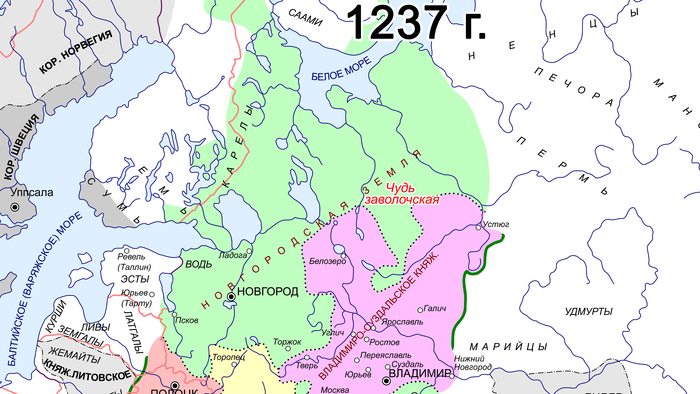
Сообщества культуры каменных могильников имели фиксированную среду обитания, отчетливую материальную культуру, существовали за счет местных ресурсов и, вероятно, пережили демографический подъем и значительное увеличение плотности населения. К настоящему времени обнаружено гораздо больше могильников с оградками культуры каменных могильников, чем захоронений с каменными ящиками в том же ареале. Археологический период каменных могильников с I по IV вв. н. э., характерных для Эстонии, Северной и Западной Латвии, Юго-Западной Финляндии и Северо-Западной России в эпоху римского железного века, можно назвать «золотым веком» протофинской цивилизации. Финны, эстонцы, ливвики и вожане — все они появились в пределах географических границ культуры каменных могильников. В частности, вожане начали отделяться от северных протоэстонских предков в I в. н. э. и гораздо позже стали частью Новгородской республики. В середине I тысячелетия н. э. (с VII-VIII вв. н. э. по XII – XIV вв. н. э.) некоторые потомки носителей культуры каменных могильников с юга современной Финляндии переселились на Карельский перешеек, где позже появилось средневековое племя Корела.
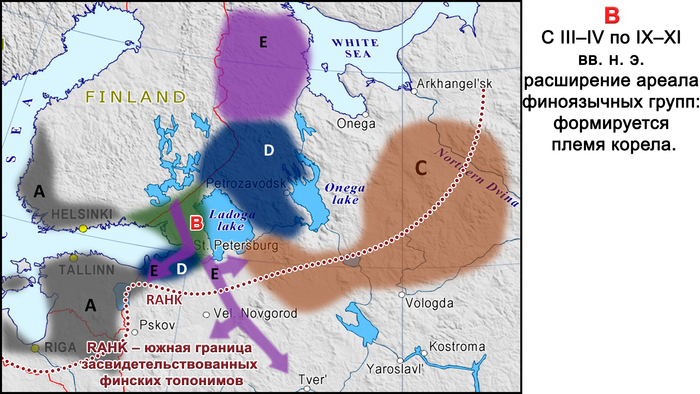
К IX–XI векам нашей эры миграции финноязычных племён усилились, что привело к дальнейшему этническому разделению. Население Приладожской «чудской» курганной культуры из южно-ладожских и белозерских регионов средневекового племени весь дало начало современным вепсам. В XI–XII веках родственные им племена расселились дальше на восток, в бассейн Северной Двины. В XII–XIV веках (особенно до XIV века) эти племена упоминались в русских летописях как чудь заволочская, а к XIX веку они перешли на русский язык.
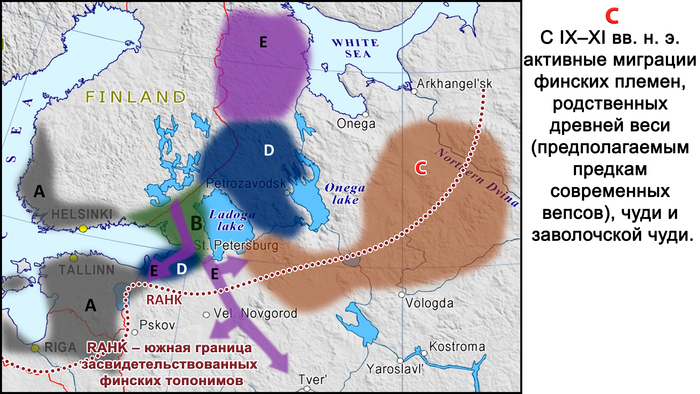

Другие миграции связаны с разделением племён карелов и вепсов.

Во-первых, с конца 1-го тысячелетия нашей эры (и особенно с XII по XIII века) и не позднее XVII века ижоры отделились от южных племён корела, которые жили на юге Карельского перешейка, и распространились на юг, вниз по реке Оредеж и вдоль южного побережья Финского залива и далее в район нижнего течения реки Луга.
Во-вторых, восточная часть племён корела вместе с одноимённым городом, современным Приозерском, которая осталась в составе Древнерусского государства (Новгородской земли) по Ореховскому договору 1323 года со Швецией, дала начало современным карелам (в частности, современным носителям южно-карельского диалекта).
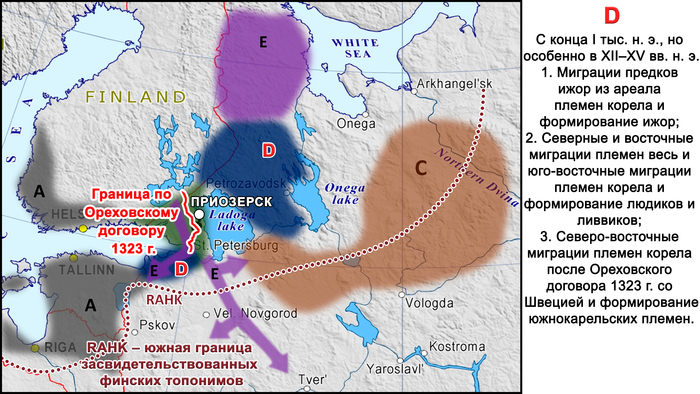
Современные карелы – ливвики и людики, проживающие в районе между Ладожским и Онежским озёрами, являются результатом смешения этих карельских племён, двигавшихся на юго-восток, с предками вепсов, двигавшихся на северо-восток.
В XVII веке после Тявзинского мирного договора 1595 года и особенно после Столбовского договора 1617 года Швеция присоединила к себе территории к северу и западу от Ладожского озера, и некоторые местные группы переселились на восток вглубь Русского царства.
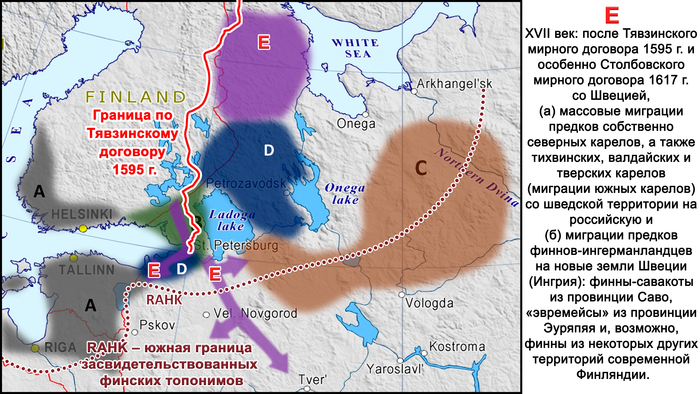
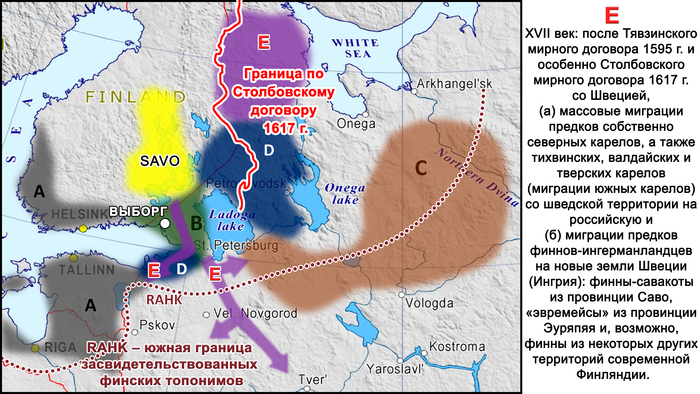
В частности, многие карелы из этих районов мигрировали сначала на север и восток, что привело к появлению современных носителей собственно северного карельского диалекта, а затем на юг, что привело к появлению современных «островков» карельского диалекта карельского языка в Тихвинском районе Ленинградской области, Валдайском – Новгродской и в Тверской области.
Происхождение финнов-ингерманландцев относится к тому же периоду. После Столбовского мирного договора финны-савакоты из провинции Саво в Финляндии, расположенной к северу от Выборга, и «эвремейсы» из провинции Эуряпяя, жившие к западу от Выборга, а также, по-видимому, финны из некоторых других финских провинций, расположенных севернее, были переселены шведскими властями на юг. Эти новые регионы включали в себя южные районы Карельского перешейка, юг бассейна Невы и далее до Сойкинского и Кургальского полуостровов, и их целью было заселение этой территории после того, как их покинуло православное население.
Подводя итог, можно сказать, что исторически прибалтийско-финские народы можно разделить на две основные группы по их миграционной активности: (а) относительно оседлая группа и (б) группа активных мигрантов. Финны, эстонцы, ливвики и вожане относятся к первой группе: они жили в пределах географических границ своей языковой родины не менее 2000 лет. К группе активных мигрантов относятся предки современных карелов, а также ижоры, ингерманландцев, вепсов и средневековые финноязычные общины Русского Севера, которые ассимилировались к XIX веку.


Все эти народы сформировались в результате миграционных волн с их языковой родины за последние 1500 лет. Сегодня оседлая группа вожан и большая часть мигрировавших народов (за исключением северных миграций финнов Финляндии) проживают в Российской Федерации.
Генофонды карелов, вепсов и ингерманландцев были проанализированы в исследовании распространения уральских языков в Северной Европе.
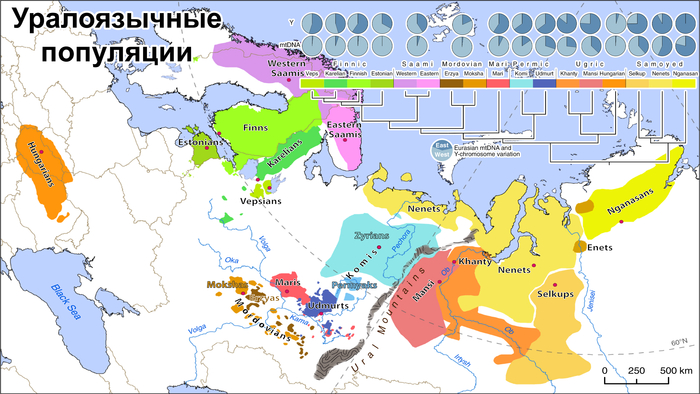
Предковая компонента, характерная для носителей уральских языков, встречается у 11–15% карелов, вепсов и ингерманландцев, что соответствует северным русским (13%), но ниже, чем у саамов (~50%), и выше, чем у эстонцев (5%). По всему спектру предковых компонент карелы, вепсы и ингерманландцы имеют наибольшее сходство с населением Финляндии.
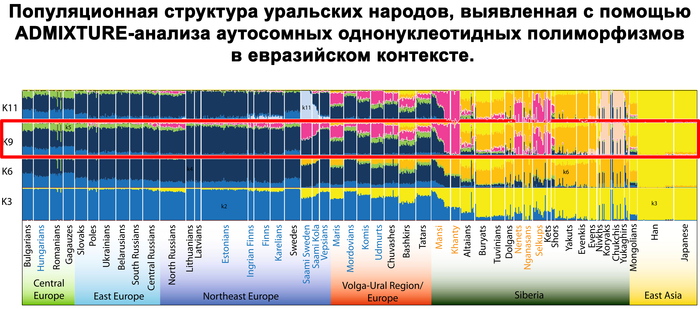
Согласно исследованиям всего генома, уральская компонента встречается в современных финноязычных популяциях с той же частотой, что и сибирская компонента, около 10% у карелов, вепсов и ингерманландцев.
В ходе исследования генофондов различных этнических групп, проживающих от Прибалтики до озера Байкал, было выявлено небольшое сходство между карелами и населением Волго-Уральского региона. Это сходство проявилось в количестве коротких сегментов ДНК от 1 до 3 сантиморганид, имеющих общее происхождение (IBD).
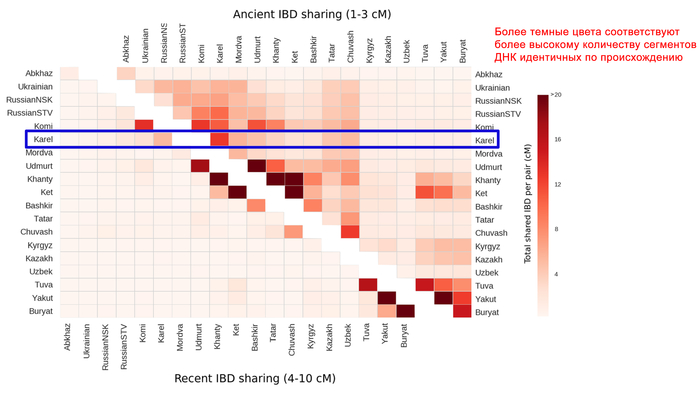
Генофонд вологодских вепсов имеет наибольшее сходство с генофондом северных русских из Мезенского района Архангельской области. Анализ полных геномов тех же популяций вепсов и карелов с помощью D-статистики показал, что у карелов, вепсов, северных русских и коми есть сибирская компонента, у карелов и вепсов около 10%. Однако у популяций, проживающих южнее, эта компонента отсутствует.
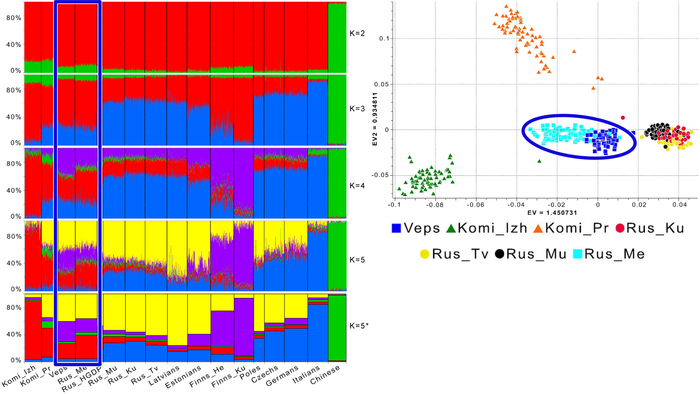
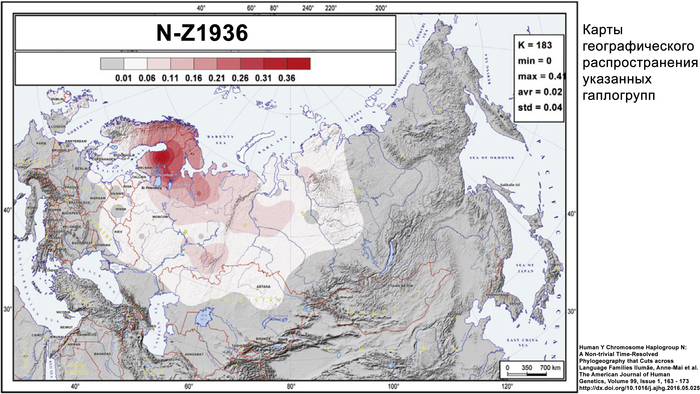
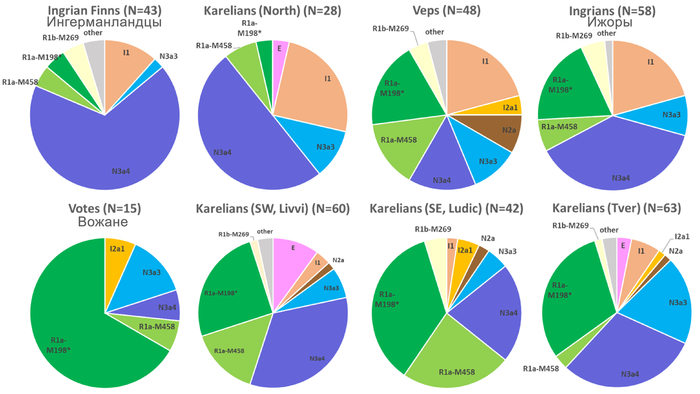
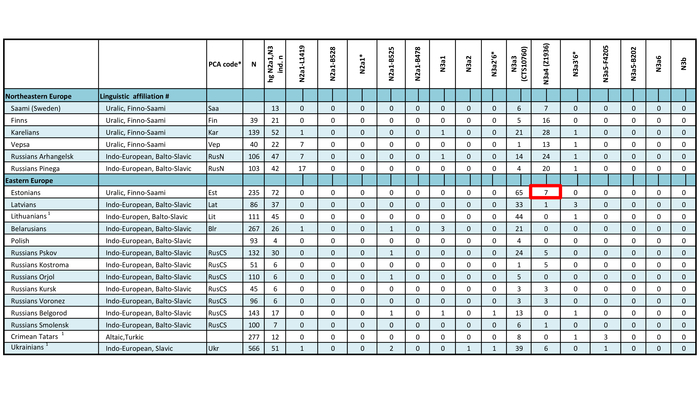
Набор гаплогрупп Y-хромосомы у финноязычных народов характеризуется повышенным присутствием гаплогруппы N с двумя её ветвями: N-Z1936, преобладающей у карелов (20%), вепсов (33%) и финнов (41%), и N-CTS10760, более распространённой у эстонцев (28%). Из пяти линий N преобладает N-Z1934 (B535), которая появилась около 4000 лет назад и встречается в популяциях Северо-Восточной Европы. Её частота достигает ~20% у карелов, ~33% у вепсов, что немного ниже, чем у финнов (около 44%), а также 20–30% у саамов и 19–23% у русских Архангельской области.
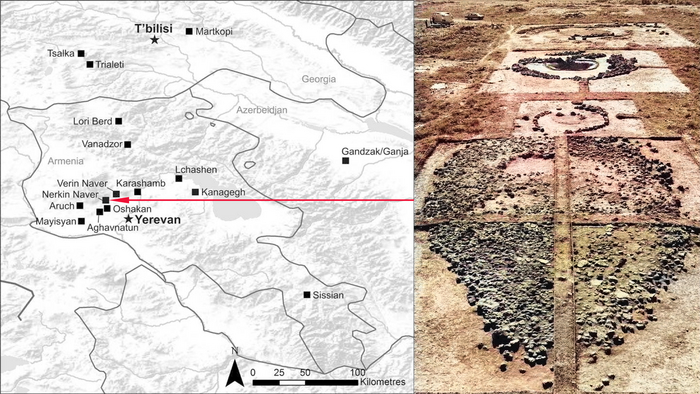
В данной работе описываются карелы в наибольшем многообразии, а именно северные и тверские карелы, людики и ливвики, а также вепсы и малоизученные популяции ижор, ингерманландцев и вожан. При этом совокупность Y-хромосом тверских карелов из предыдущей работы продемонстрировала их генетическое родство с южными популяциями карелов и вепсов.
Всего исследование охватывает восемь финно-угорских популяций, в настоящее время проживающих на территории Российской Федерации. В частности, авторы изучали северных карелов Калевальского района, ливвиков Олонецкого района, людиков Пряжинского района Республики Карелия и тверских карелов Тверской области, а также вепсов Прионежского района Карелии, ингерманландцев Гатчинского муниципального округа, ижорцев Сойкинского полуострова и нижнего течения реки Луга Кингисеппского района Ленинградской области, и западных вожан из района бывшей деревни Краколье в пределах посёлка Усть-Луги Кингисеппского района Ленинградской области.
В этом исследовании упор также сделан на локальное сравнение финно-угорских групп с соседними популяциями русских.
Кстати, здесь позволю себе сделать некоторое отступление, потому как по опыту знаю, что, когда в исследовании упоминаются финно-угорские народы и русские, часто в комментариях появляются люди, распространяющие искаженные данные более ранних работ учёных, которые некоторые СМИ упорно распространяли около 20 лет, каждый раз подавая их как новые сенсационные открытия того, что «русские оказались финнами» и, следовательно, «русские — не славяне». Те, кому нравились такие выводы, в это верили, не проверяя, несмотря на то, что сами учёные, на которых ссылались авторы подобных статей, полностью опровергли подобные выводы и указали на ошибки и подмену понятий, карт и данных. Не стану подробно рассказывать, что «исконно русских» или каких-то ещё генов попросту не существует, потому как это надолго и может отвлечь от обзора нового исследования, а сразу сообщу основные сведения, о которых известно уже около 10 лет и более.
Так, по всем генетическим маркерам, а именно по Y-хромосоме, митохондриальной ДНК и полногеномным аутосомным данным, русские центральных и южных областей, украинцы и белорусы группируются в свой собственный кластер, который находится близко к кластеру западных славян, но не перекрывается с ним.
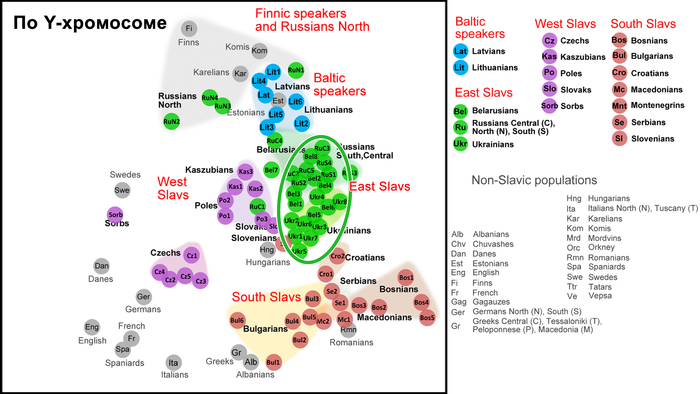
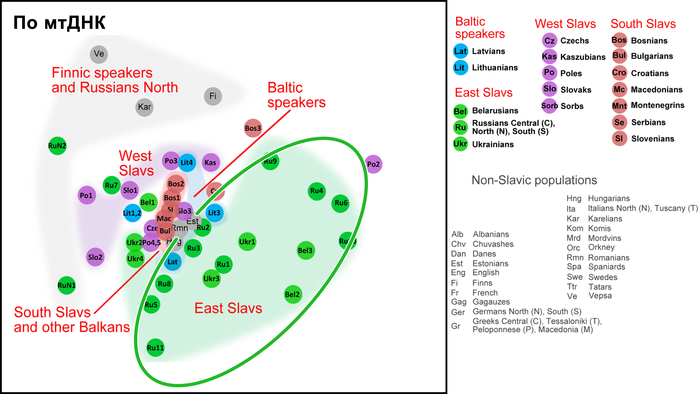
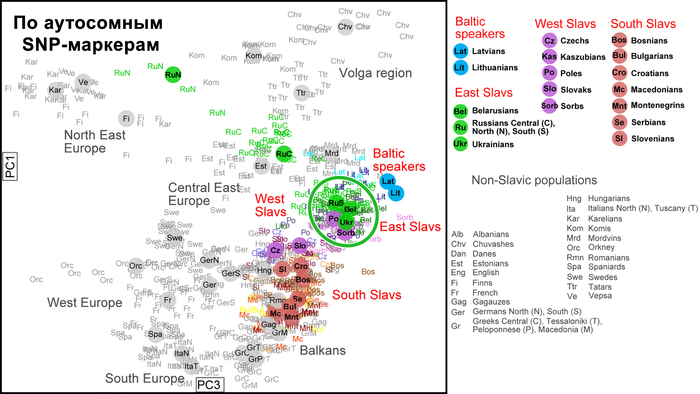
А внутри этого восточнославянского кластера генофонды русских и украинцев «образуют два облака, которые соприкасаются и частично переходят друг в друга по краям без четкой границы. Что касается белорусов, то часть белорусских популяций Полесья генетически неотличима от украинцев, а другая часть — от русских». Однако исключением являются северные русские Архангельской области, особенности которых часто, но ошибочно распространяют на всех русских. Они также упоминаются и в работе, которой посвящён этот обзор. Эти популяции русских генетически отдаляются от остальных восточных славян, тяготея к финно-угорским популяциям, при этом не переставая быть русскими, как минимум по самоопределению и своей истории. При этом всё население Архангельской области, по данным 2024 года, составляет чуть менее 1 миллиона человек. Однако, если убрать региональную привязку к Архангельской области, получаются такие вот горе-сенсации.
Аутосомный состав популяций Карелии (вепсов и карелов) более однороден, чем их отцовские линии. Генофонд Карелии по линиям Y-хромосомы в значительной степени сформирован тремя генетическими компонентами и характеризуется межпопуляционными различиями в разнообразии ветвей N.
Различия между северными и южными популяциями карелов, а именно между людиками и ливвиками, можно объяснить, как их географической изоляцией друг от друга (около 400 км), так и историей их формирования. Они также могут указывать на генетическое родство между карелами-людиками и исчезнувшей древней популяцией, оставившей свой след в генофонде центральных русских.
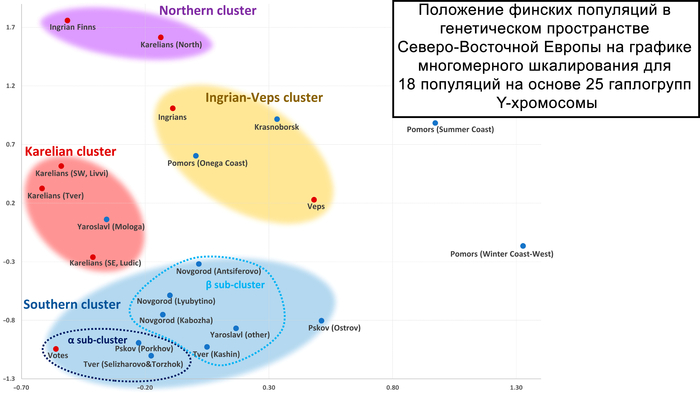
На уровне Y-хромосом большое генетическое расстояние между вепсами и тремя популяциями карелов, но близость вепсов к северным русским может указывать на то, что вепсы и финноязычные народы Русского Севера, ассимилировавшиеся в XVI–XIX веках, имеют общее генетическое происхождение. Действительно, исчезнувший народ заволочская чудь из района Поморья часто связывали с вепсами.
В течение последних трёх столетий тверские карелы жили примерно в 500 километрах от других карельских народов. Более раннее исследование показало, что генофонды тверских карелов и карелов с вепсами из Карелии схожи.
Используя расширенную панель гаплогрупп Y-хромосомы и панель SNP всего генома, исследование демонстрирует генетическое родство тверских карелов с ливвиками и, в некоторой степени, с людиками и ижорой. Это противоречит историческим данным, свидетельствующим об общем происхождении северных и тверских карелов.
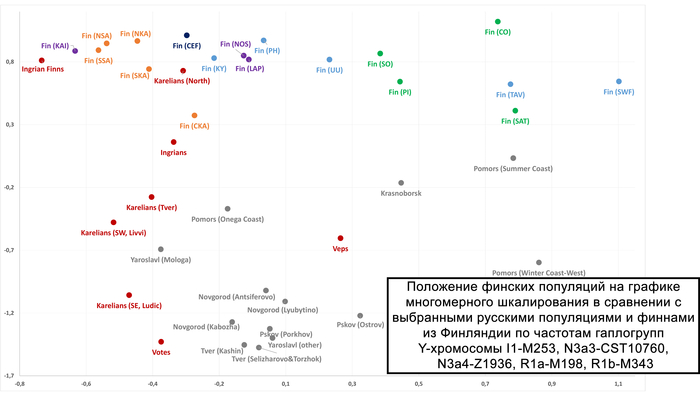
Анализ Y-хромосомы показывает, что ингерманландцы наиболее близки к северным карелам и восточным финнам, в том числе к финнам из провинций Северное и Южное Саво, которые исторически считаются источником миграции савакотов, предков ингерманландцев. Наибольшее разнообразие линий в пределах N наблюдается у ингерманландцев. А наличие этих линий у ижоры и карелов говорит об их общем происхождении.
Анализ аутосомных данных показывает генетическую близость между ингерманландцами, ижорой и вожанами, что могло быть результатом браков между ингерманландцами и женщинами ижоры и вожан или того факта, что значительная часть современного генофонда исторической Ингерманландии происходит от коренного местного населения (возможно, от вожан или местных славянских племён). Учитывая, что вожане и ижора были православными, как и русские, смешанные браки между этими тремя группами исторически были распространены, в то время как их браки с лютеранскими ингерманландцами до Второй мировой войны были редки, но всё же были.
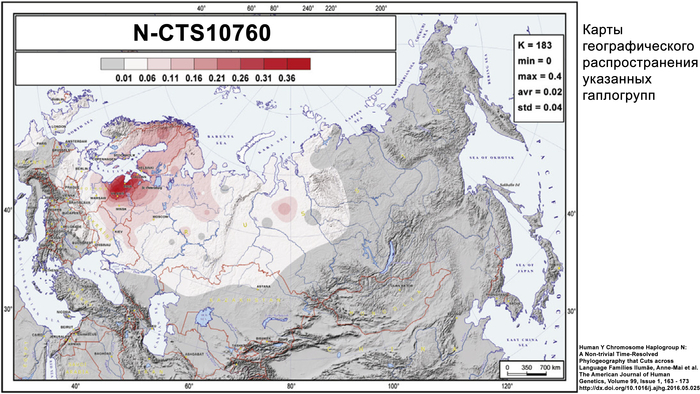
В генофонде анализируемых финно-угорских народов России насчитывается 18 полиморфных гаплогрупп Y-хромосомы. Наиболее распространёнными из них являются I1, N-CTS10760, N-Z1936 и R1a.
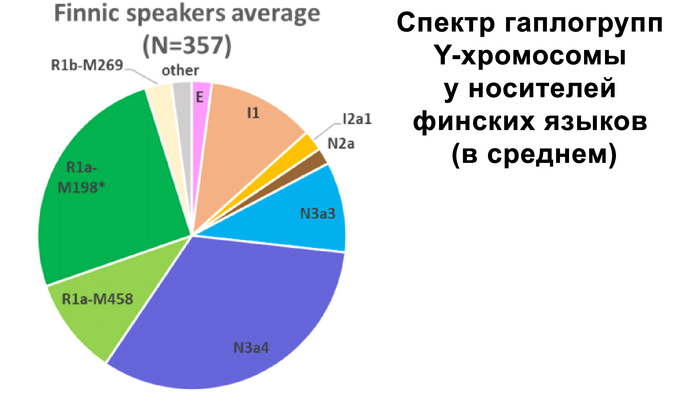
Вклад гаплогруппы I1 максимален у северных карелов, вепсов и ижор (21–25%) и высок у ингерманландцев (12%). Гаплогруппа N-CTS10760 составляет пятую часть всего разнообразия Y-хромосом тверских карелов (19%) и встречается с высокой частотой у ингерманландцев, вожан, вепсов и северных карелов (9–13%). Гаплогруппа N-Z1936 составляет треть генофонда Y-хромосом финноязычных народов России, преобладая у ингерманландцев (67%) и северных карелов (50%). Её вклад в генофонды вепсов, ливвиков, тверских карелов и ижор составляет от 21% до 38%. Гаплогруппа R1a, включая её ветвь R1a-M458, является доминирующей гаплогруппой у вожан (73%) и людиков (60%) и составляет треть разнообразия Y-хромосом у ливвиков, тверских карелов, вепсов и ижор.
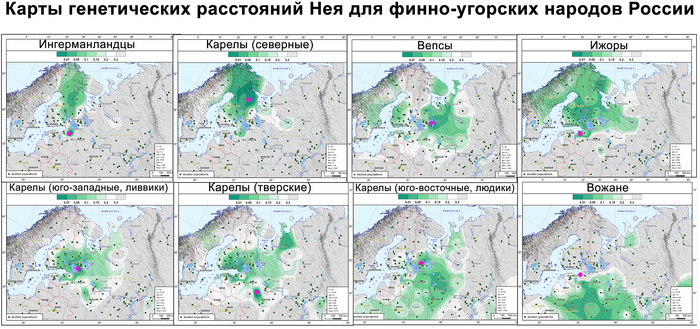
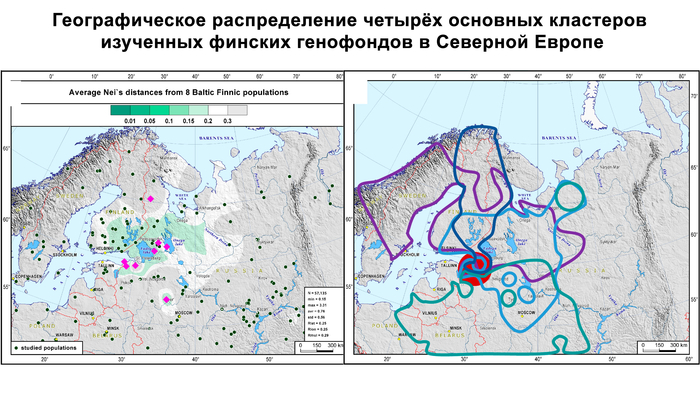
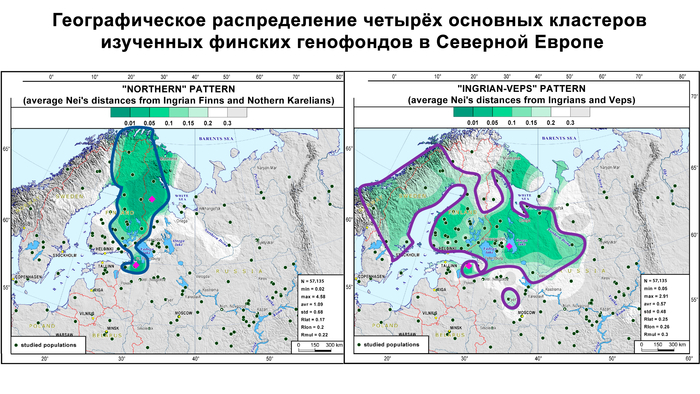
Изученные популяции заметно отличаются по разнообразию четырёх основных гаплогрупп и их частотам как друг от друга, так и от усреднённой выборки, объединяющей все изученные финно-угорские группы. Генофонды финских популяций сравнивались друг с другом и с некоторыми соседними популяциями русских по всему спектру гаплогрупп Y-хромосом с помощью матрицы попарных генетических расстояний Нея.
Данные были визуализированы на графике многомерного шкалирования и картах генетических расстояний Нея.
На графике вожане и ижоры занимают противоположные точки генетического пространства, несмотря на географическую близость. При этом генофонд вожан по отцовским линиям резко отличается от генофонда всех остальных финно-угорских групп из-за значительного вклада гаплогруппы R1a. Вожане образуют отдельный отдалённый кластер вместе с населением Центральной России. Причём только с современными русскими из Порхова Псковской области, который был в составе Новгородской республики, ближайшими географическими соседями вожан из всех рассмотренных здесь новгородских русских популяций. Такая же степень сходства наблюдается между вожанами и русскими, проживающими на западе Тверской области России, в Селижаровском и Торжском районах.
Это может быть связано с генетическим дрейфом, небольшим размером популяции или изначальным различием между генофондами вожан и ижор. Последнее предположение о различиях может быть правдоподобным, учитывая отсутствие миграционной активности у вожан.
Здесь стоит учитывать, что отличие по отцовским линиям, по которым многие комментаторы ставят клеймо на людей, не означает отличие по всему геному, т. е. по большинству предков, а по ним вожане и ижоры являются частью кластера Ингерманландии вместе с финнами-ингерманландцами. При этом внутрипопуляционное разнообразие ижор поражает как с точки зрения аутосомных данных, так и гаплогрупп Y-хромосомы. По мужским линиям ижоры, находясь в пределах кластера ижор и вепсов, тяготеют и к карельскому, и к северному кластерам ингерманландцев и северных карелов.
Это объяснимо, потому как выборка ижор из исследования состоит из двух подгрупп: представителей Сойкинского полуострова и нижнего течения реки Луга, которые имеют совершенно разную этническую историю, которую авторы опишут в следующей своей работе.
Помимо этого, по линиям Y-хромосомы были выявлены связи вепсов, поморов Онежского берега и русских Красноборского района Архангельской области с популяциями заволочской чуди.
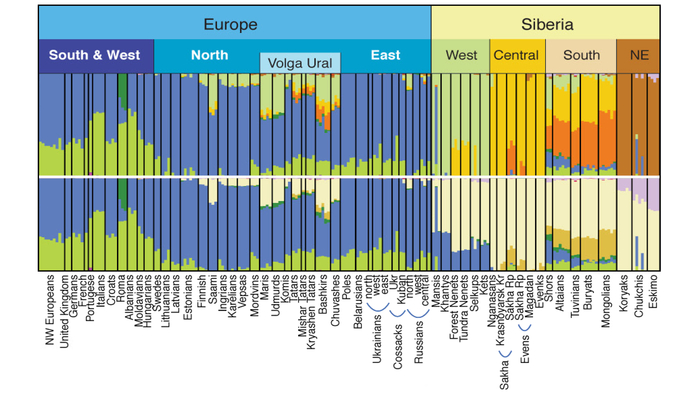
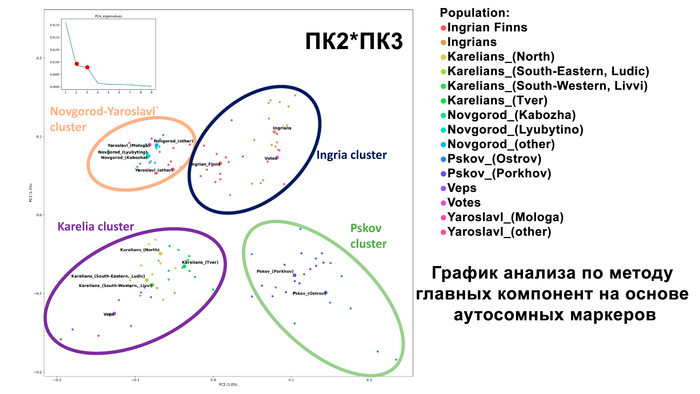
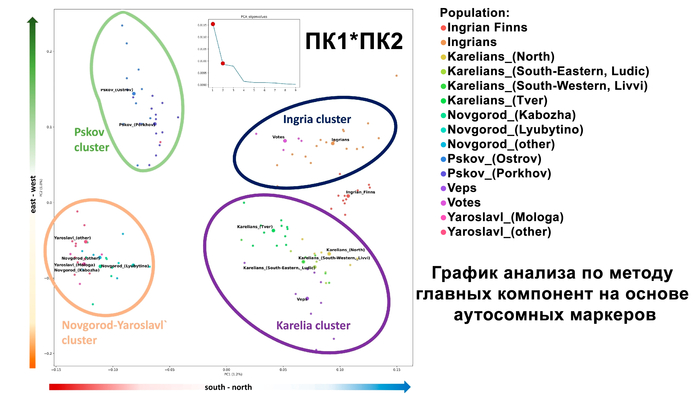
Структура аутосомного и Y-хромосомного генофондов финских популяций России существенно отличается. Генофонд Y-хромосомы финно-угорских популяций более разнообразен, чем аутосомный. Его можно разбить на два больших блока в зависимости от их географической близости:
Популяции исторической Ингерманландии (вожане, ижоры и финны-ингерманландцы).
Популяции Карелии (вепсы и все популяции карелов).
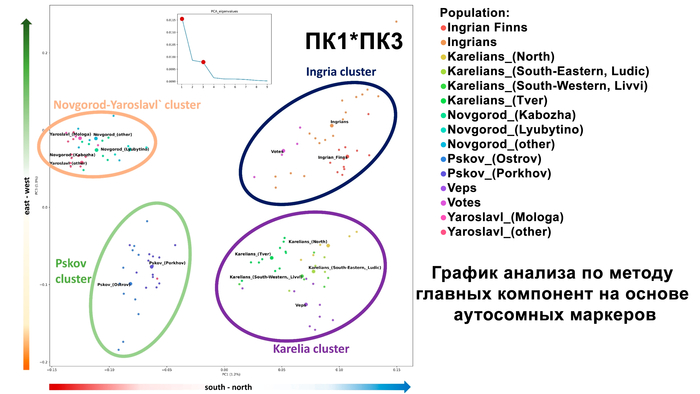
Такая структура обнаруживается на базовом уровне моделирования с участием 6 предковых компонент (K = 6), что косвенно указывает на её древнее происхождение.
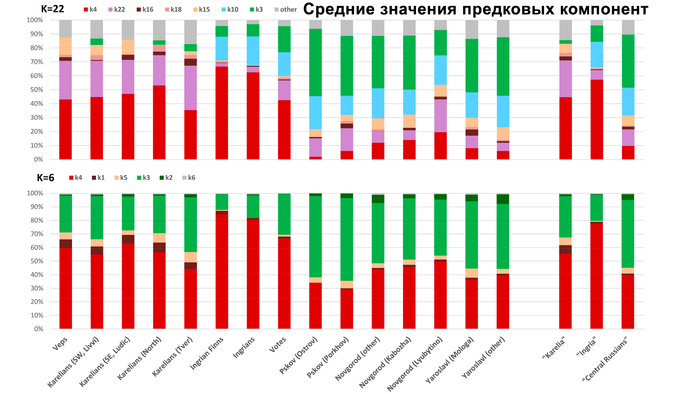
Эта структура сохраняется и в более детальной модели с участием 22 предковых компонент, в этой модели наблюдаются более резкие различия между финно-угорскими народами и русскими соседних районов. В аутосомном генетическом пространстве русские центральных областей отделены от финно-угорских групп, хотя в пространстве Y-хромосом в кластер к русским попадают вожане. А вместе финно-угорские народы и соседние популяции русских образуют четыре кластера: «Карелия» (все карельские популяции и вепсы), «Ингерманландия» (ижора и вожане), «Псков» (русские Островского и Порховского районов Псковской области) и «Новгород-Ярославль» (русские Новгородской и Ярославской областей). При этом ингерманландцы в зависимости от выбранной главной компоненты либо попадают между кластерами «Карелия» и «Ингерманландия», либо в кластер «Ингерманландия» к ижорам и вожанам.
Аутосомный генофонд вепсов наиболее близок к генофонду ливвиков и людиков. Однако, согласно гаплогруппам Y-хромосомы, вепсы довольно далеки от ливвиков, людиков и тверских карелов, но еще более далеки от северных карелов.
Как уже упоминалось, структура аутосомного генофонда зависит от значительно большего числа предков человека, чем структура генофонда по гаплогруппам Y-хромосомы и мтДНК. Однако, хотя генофонд по гаплогруппам Y-хромосомы более подвержен влиянию генетического дрейфа, он лучше сохраняет древние предковые линии на протяжении тысячелетий, даже в тех случаях, когда в аутосомном генофонде от этих древних предков не осталось и следа. Здесь главное — грамотно и умело пользоваться этими однородительскими маркерами.
Гаплогруппа N-Z1936 является основной гаплогруппой для популяций трёх из четырёх кластеров в генетическом пространстве Y-хромосомы. В генофонде финно-угорских народов она представлена в основном субкладой N-Z1927 (её предковая ветвь Z-1934 была обнаружена с высокой частотой у карелов, вепсов, финнов, саамов и северных русских). Это означает, что большинство носителей N-Z1936 среди финно-угорских народов, а также среди некоторых народов Северной и Центральной России происходят от общего предка, жившего примерно 2400 лет назад. Гаплогруппа N-Z1927 представлена ветвями N-Z19825 и N-CTS4329, которые появились около 2 тыс. лет назад.
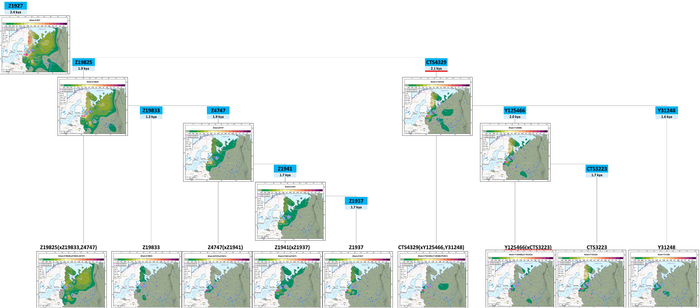
Обе ветви встречаются минимум в одной карельской популяции и по крайней мере в одной популяции из Ингерманландии. Эти ветви также распространены по всей Финляндии с высокой частотой: 21,1% и 16,6% соответственно.
Ветвь N-Z19825* является корневой кластерной (полная номенклатура: Z19825(xZ19833, Z4747)) и, предположительно, включает в себя группу ещё не изученных подветвей. Это предположение основано на обширной географии N-Z19825*, которая встречается не только у карелов и вепсов, но и у русских из Архангельской и Вологодской областей. Дальнейшее изучение этой ветви может привести к обнаружению её линий, общих для северных русских и населения Карелии, а также региональных вариантов, появившихся за последние 2000 лет, то есть за всё время существования линии N-Z19825.
Субгаплогруппа N-CTS4329, возрастом около 2100 лет, более характерна для финно-угорских народов, в частности для северных карелов (31%), ингерманландцев (19%), ижор (16%) и ливвиков (8%). Среди рассмотренных в данной работе русских она наиболее часто встречается у ярославских русских из Мологи (5%) и северных русских. Также N-CTS4329 встречается с частотой от 1% до 3% у народов Поволжья – эрзян, марийцев и татар-мишарей.
В пределах N-CTS4329 авторы проанализировали три ветви: основную ветвь N-Y125466 и две редкие ветви N-Y31248 и N-CTS4329*. Географическое разнообразие N-Y125466, возрастом около 2000 лет, в значительной степени представлено её субкладой N-CTS3223, возрастом около 1700 лет, которая чаще всего встречается у северных карелов (31%), ингерманландцев (8%), ливвиков (4%) и ижор (4%). Субклада N-Y125466 (xCTS3223) встречается у ингерманландцев (5%), поморов Зимнего берега (3%) и у эрзян (2%). Редкая ветвь N-Y31248, возрастом около 1700 лет, ограничена карелами-ливвиками, ингерманландцами и русскими Мологи (3–5%). А ветвь N-CTS4329 (не считая сублинии Y125466 и Y31248) встречается только у ижор (11%) и у северных русских из Красноборского района Архангельской области (3%).
Таким образом, разнообразие N-Z1936 в изученных финноязычных популяциях в значительной степени представлено девятью параллельными ветвями, которые появились около 1300-2100 лет назад. Четыре из них имеют обширное географическое распространение (Z19825*, Z1941*, Z1937 и CTS3223), а остальные встречаются в виде изолированных островков.
Филогенетическое древо указывает на рост популяции носителей N-Z1927, поскольку в каждой из её ветвей есть новые линии. Линии CTS3223 и Z1941 появились около 1700 лет назад и встречаются как в Карелии, так и среди населения исторической Ингерманландии, а также в Финляндии. Таким образом, в то время как предок N-Z1927 жил около 2400 лет назад, быстрый рост численности населения среди его финноязычных потомков, по-видимому, начался только 1700–2000 лет назад.
Этот рост хронологически соответствует I–IV вв. н. э., периоду археологической культуры каменных могильников – «золотому» веку протофиннской цивилизации. Однако эта культура типична для территории современной Эстонии, в то время как гаплогруппа Y-хромосомы N-Z1936 относительно редко встречается у эстонцев.
Таким образом, предполагаемый рост численности населения среди носителей N-Z1927 не имеет чёткой связи с распространением культуры каменных могильников.
Однако усреднённая карта генетических расстояний охватывает ареалы обитания финских групп в России и на востоке Финляндии.
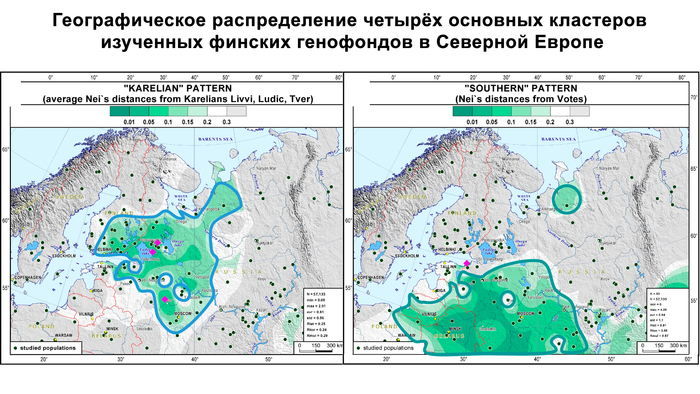
А пересечение четырёх кластеров Y-хромосомы в самой восточной части распространения культуры каменных могильников может указывать на восточную часть прародины предкового генофонда прибалтийских финнов. Дальнейшие исследования древней ДНК представителей культуры каменных могильников в этом самом восточном уголке прародины прибалтийских финнов могут пролить свет на возможную связь этих людей с распространением Y-хромосомы N-Z1927 в тот период.
Это исследование представляет собой первый комплексный анализ генофонда прибалтийско-финских народов России с использованием двух наиболее информативных систем генетических маркеров. Генофонд Y-хромосомы изучаемых финно-угорских народов более разнообразен, чем аутосомный, и состоит из четырёх генетических кластеров:
Северный кластер — включает ингерманландцев и северных карелов.
Кластер ижор и вепсов — включает одноимённые народы.
Карельский кластер — объединяет ливвиков, людиков и тверских карелов.
Южный кластер — включает вожан (кстати, в этот кластер входят и некоторые популяции русских).
Однако аутосомный генофонд исследованных финно-угорских народов, можно разделить на 2 кластера:
Кластер Карелии — включает вепсов, северных карелов, людиков, ливвиков и тверских карелов.
Кластер Ингерманландии — включающий ижор, вожан и ингерманландцев.
При этом популяции русских попадают в два отличающихся кластера — новгородско-ярославский и псковский.
В более детальной модели (построенной с помощью метода ADMIXTURE), с участием 22 предковых компонент, компоненты k4 и k22 встречаются во всех популяциях, но преобладают в финно-угорских. Компонента k10 в равной степени характерна как для «Ингерманландии», так и для групп Центральной России, у которых в генофонде преобладает компонента k3.
Филогеографический анализ гаплогруппы N-Z1927 позволяет предположить, что у исследованных финно-угорских групп и некоторых северных и центральных русских, около 2400 лет назад был общий предок, но рост численности носителей N-Z1927 произошёл гораздо позже, примерно 1700–2000 лет назад, в период культуры каменных могильников, когда цивилизация финноязычных народов в целом процветала.
Однако тот факт, что территория Эстонии была одним из центров культуры каменных могильников, а гаплогруппа N-Z1936 редко встречается у современных эстонцев, указывает на то, что необходимы дополнительные исследования для изучения возможной связи между предполагаемым ростом населения и распространением культуры каменных могильников.
Новые данные, которые указали на высокую внутреннюю неоднородность ингерманландцев, относительно изолированное положение вепсов среди изученных прибалтйиско-финских групп, а также на сходство между вепсами и русскими поморами, требуют дальнейшего междисциплинарного изучения всех этих групп и сравнения восточно-финских популяций с более обширным набором образцов, в который также вошли бы и сибирские популяции.
Источник:Agdzhoyan, A.; Ponomarev, G.; Pylev, V.; Autleva, Z.; Gorin, I.; Evsyukov, I.; Pocheshkhova, E.; Koshel, S.; Kuleshov, V.; Adamov, D.; et al. The Finnic Peoples of Russia: Genetic Structure Inferred from Genome-Wide and Y-Chromosome Data. Genes 2024, 15, 1610. https://doi.org/10.3390/genes15121610
По русским: https://генофонд.рф/?page_id=30373
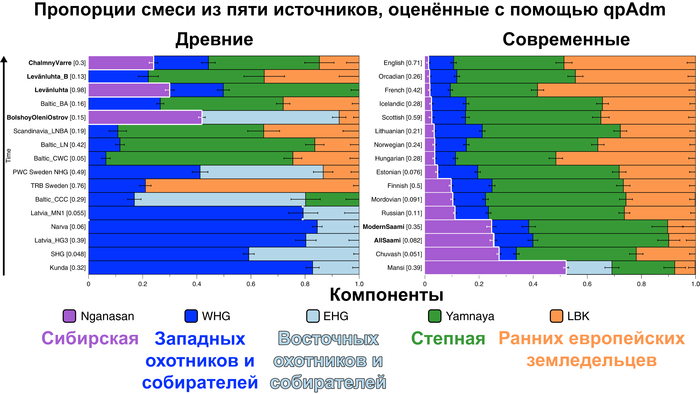
Пять тысяч лет назад в Евразии происходили масштабные миграции людей, которые существенно изменили генетический состав её населения. Эти волны миграций были связаны с кочевыми скотоводами ямной культуры раннего бронзового века. Они распространились на обширные территории, оставив заметный след как в культуре, так и в генетике.
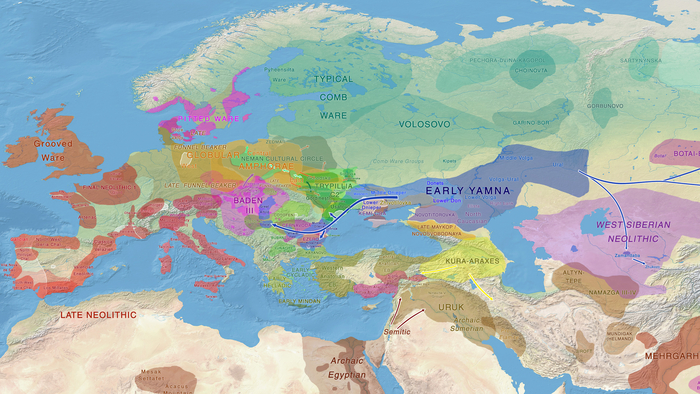
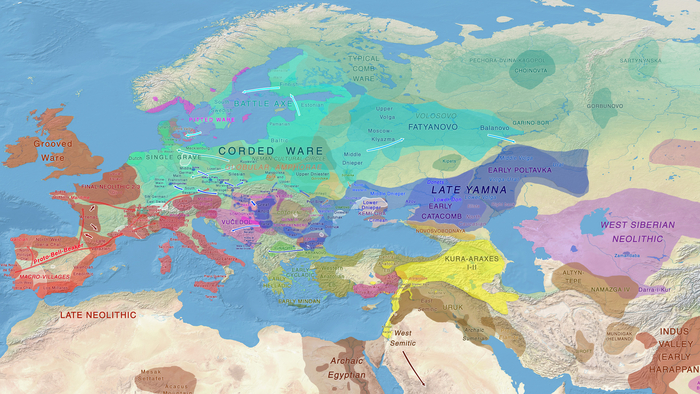
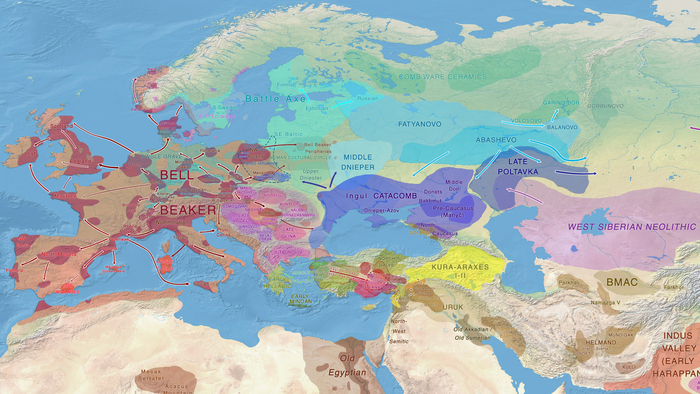
Эти миграции также, вероятно, сыграли ключевую роль в распространении индоевропейских языков. Однако степное происхождение распространялось в различных регионах Западной Евразии разными путями и в разное время.
В различных регионах Европы вклад от степных скотоводов был связан с популяциями культур шнуровой керамики (КШК) (5000–4350 лет назад) и колоколовидных кубков (4800–4300/3800 лет назад).
Однако степень, в которой подобная динамика проявлялась в популяциях Восточного Средиземноморья и прилегающих территорий Западной Азии, остается менее изученной.
Хотя ранее степная родословная была обнаружена у древних жителей современных Испании, Италии, Греции и Кавказа, вопросы о взаимосвязи этих популяций в контексте Средиземноморского региона в целом остаются открытыми.
Распространение степных народов тесно связано с разделением индоевропейского праязыка на исторически зафиксированные подгруппы.
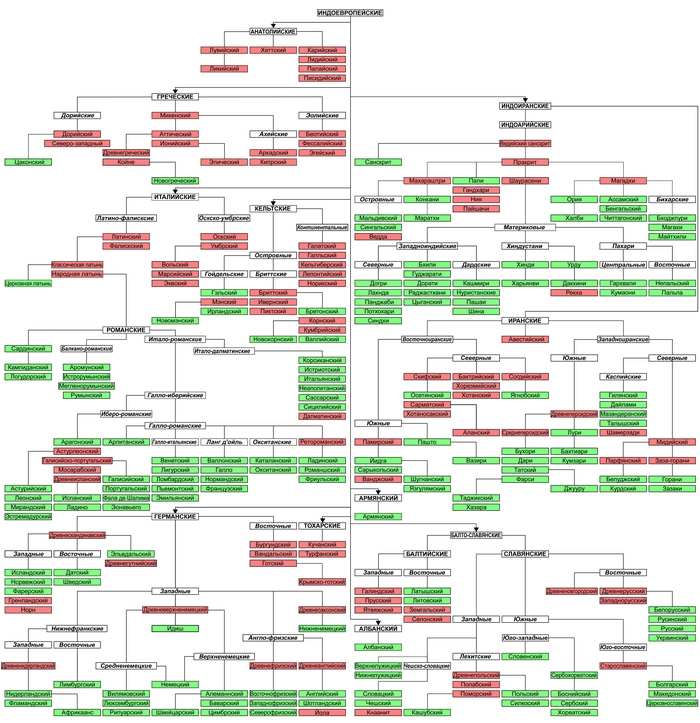
В Средиземноморье важными индоевропейскими языками классической античности были галльский, латинский и греческий, а армянский, был распространён на Южном Кавказе и на востоке полуострова Малая Азия. Для них было предложено несколько конкурирующих филогенетических лингвистических моделей. Так называемая индо-греческая гипотеза объединяет греческий язык, а также близкородственный ему фригийский с индоиранскими языками, в то время как конкурирующая греко-армянская гипотеза предполагает, что греческий образует подгруппу с армянским, возможно, также включая албанский («балканский индоевропейский»). Аналогичным образом, италийская индоевропейская подгруппа, от которой произошёл латинский язык, по-разному объединялась с кельтской и германской подгруппами, что привело к появлению традиционно популярных итало-германской и контрастной итало-кельтской гипотез. Хотя касательно греко-армянской и итало-кельтской гипотез существует относительный лингвистический консенсус, они не являются бесспорными.
Основным препятствием для междисциплинарного изучения распространения индоевропейских языков является отсутствие чёткой лингвистической модели, которая могла бы объяснить, как эти языки разделились.
В своей работе авторы изучили различные источники степного происхождения, расположенные вдоль северной границы Средиземноморья, чтобы разработать наиболее подходящую модель разделения индоевропейских языков в этом регионе.
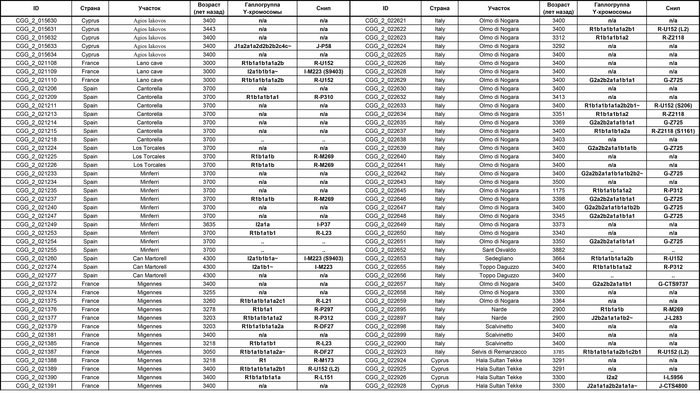
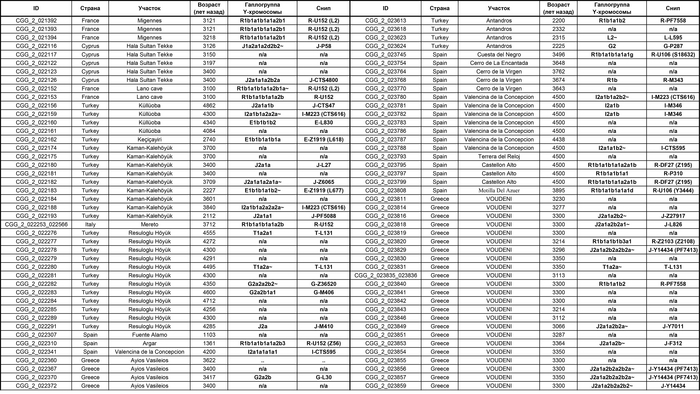
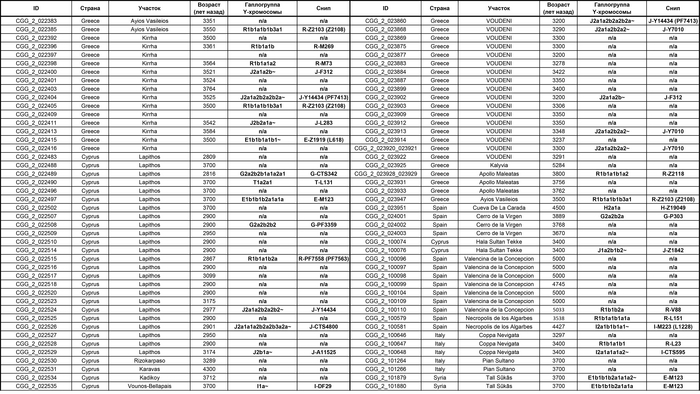
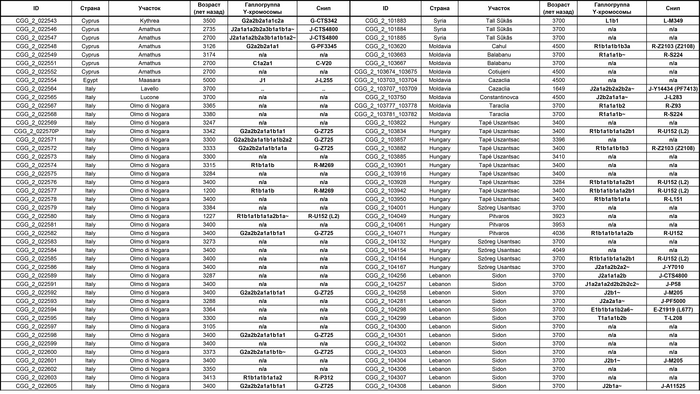
Они представили новые данные о полных геномах 314 древних людей из различных частей Европы и Восточного Средиземноморья, включая Испанию, Францию, Италию, Венгрию, Молдову, Грецию, Кипр, Турцию, Сирию и Ливан. Большинство из этих людей жили в бронзовом веке, но их возраст варьируется от 5200 до 2100 лет.
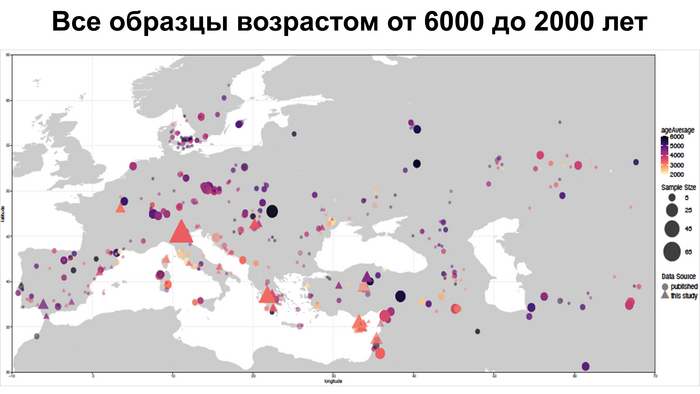
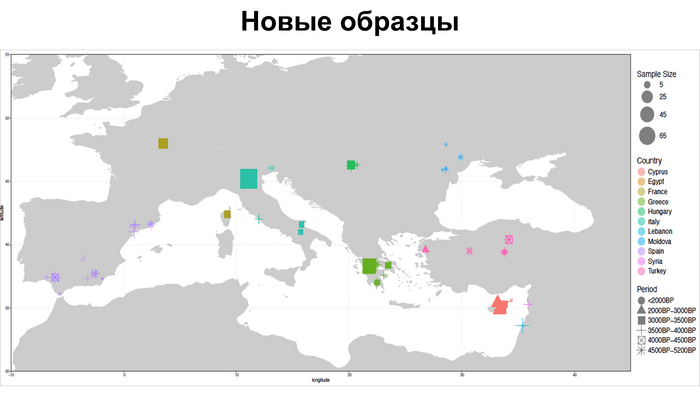
Авторы также предоставили изотопные данные для 224 человек и радиоуглеродный возраст для 144 человек. Используя сегменты ДНК, идентичные по происхождению и моделирование примесей с конкретными исходными популяциями, они получили более точное представление о генетическом происхождении этих людей. Кроме того, они объединили изотопные и генетические данные, чтобы лучше понять, как люди перемещались с течением времени.
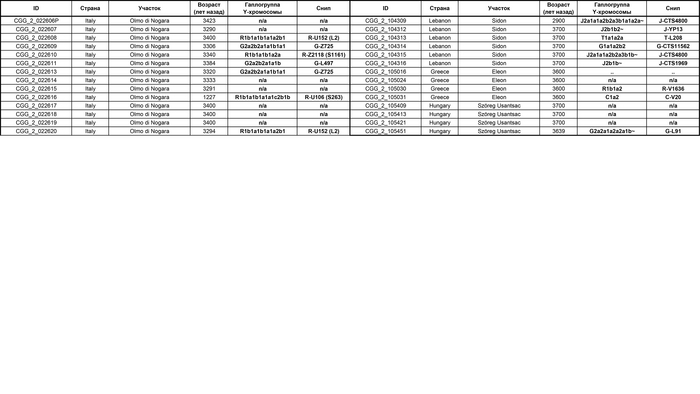
Используя кластерный анализ, учёные выделили шесть основных кластеров:
1. Кластер земледельцев. Этот кластер охватывает популяции, связанные с земледелием и включает в себя четыре субкластера.
1.1. Европейские земледельцы. Этот субкластер охватывает популяции, которые занимались земледелием на территории Европы.
1.2. Средиземноморье 3000-4000 лет назад. Этот субкластер включает в себя популяции, проживавшие в Средиземноморье в период с 3000 по 4000 лет назад.
1.3. Ранние земледельцы. Этот субкластер охватывает популяции, занимавшиеся земледелием в более ранние периоды.
1.4. Италия бронзового века. Этот субкластер включает в себя популяции, проживавшие в Италии в эпоху бронзы.
2. Европейские охотники-собиратели. В этот кластер входят популяции, проживавшие на территории Европы в эпоху охотников и собирателей.
3. Кавказ – Иран. Этот кластер объединяет популяции, проживавшие в Кавказском регионе и Иране.
4. Степной кластер. Этот кластер включает популяции, которые были связаны со степными регионами Евразии и подразделяется на три субкластера.
4.1. Россия-Центральная Азия. Этот субкластер включает в себя популяции, расселившиеся на территории России и Центральной Азии.
4.2. Культура колоколовидных кубков. Этот субкластер охватывает популяции, связанные с культурой колоколовидных кубков.
4.3. Культуры шнуровой керамики и ямной. Этот субкластер охватывает популяции, относящиеся к культурам шнуровой керамики и ямной.
5. Центральная Азия – Сибирь. Этот кластер включает в себя популяции, обитавшие в Центральной Азии и Сибири эпохи бронзы.
6. Марокканские охотники-собиратели. Этот кластер охватывает популяции, населявшие Марокко в эпоху охотников и собирателей.
Особенно подробно исследователи изучили кластеры земледельцев и степной.
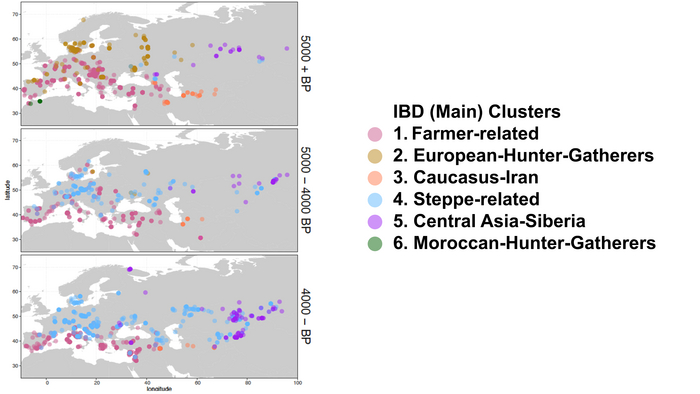

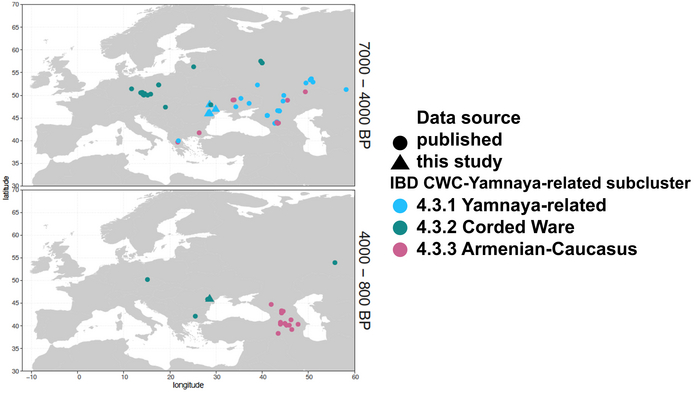
Появление этих субкластеров связано с распространением родственных ямной культуре предков как на восток, так и на запад, что привело к формированию новых генетических профилей по всей Европе и Центральной Азии.
Сосредоточившись конкретно на временных рамках этого исследования, можно наблюдать чёткую закономерность в распределении этих степных кластеров от 5 до 4 тыс. лет назад. Распространение людей из кластера, связанного с культурами шнуровой керамики и ямной, из Причерноморско-Каспийской степи, в Центральную и Восточную Европу, а также на Север Греции происходило до 4 тыс. лет назад. Этот кластер сохранялся к западу от Черного моря и в Армении после 4 тыс. лет назад, тогда как кластер связанный с культурой колоколовидных кубков стал преобладающим в Европе.
На юге Греции в этот период наблюдается смещение границы между степным и земледельческим кластерами.
Первый случай приграничных взаимодействий относится к концу второго тысячелетия до нашей эры. В Греции были обнаружены два образца бронзового века, которые относятся к кластеру, связанному с культурами шнуровой керамики и ямной. Этот кластер, в свою очередь, подразделяется на более мелкие субкластеры. Один из образцов относится к субкластеру ямной культуры, а другой — к армяно-кавказскому.
В восточной части этой пограничной зоны люди из Молдовы попадают в субкластеры, связанные со степным кластером. В то же время, все жители Восточного Средиземноморья, включая греков, входят в субкластер европейских земледельцев.
Образцы раннего бронзового века из Молдовы относятся к субкластеру, связанному с ямной культурой, а образцы среднего бронзового века — к субкластеру культуры шнуровой керамики.
Помимо этого, все жители Малой Азии, Кипра и Леванта, жившие в период между 3 и 4 тысячами лет назад, относятся к средиземноморскому кластеру этого периода, за исключением одного человека из раннего бронзового века и двух человек с Кипра, которые были отнесены к кластеру ранних земледельцев и к кластеру, связанному с культурой колоколовидных кубков.
Чтобы лучше понять, как сформировалась генетическая основа индоевропейской языковой семьи и как произошло её разделение, авторы изучили вклад наиболее вероятных предковых популяций из различных регионов Средиземноморья. В том числе из регионов, в которых исторически были распространены италийский, кельтский, греческий и армянский языки.
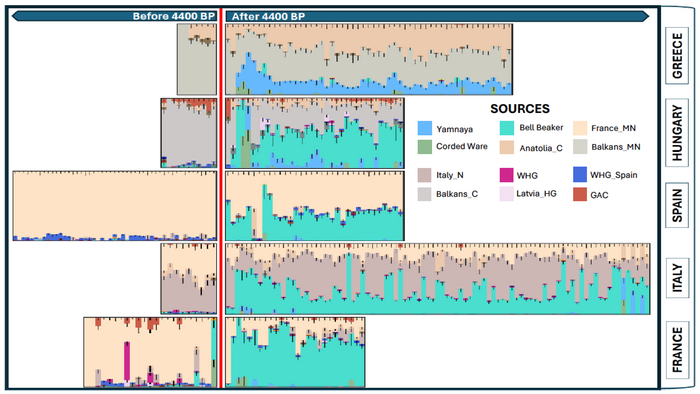
Согласно сегментам ДНК, идентичным по происхождению и моделированию примесей с предполагаемыми степными предковыми популяциями, подтверждаются существенные различия между восточными и западными индоевропейскими популяциями Средиземноморья. Благодаря наличию степных предковых линий, связанных как с ямной культурой у одних, так и с культурой колоколовидных кубков у других.
Что также подтверждает греко-армянскую и итало-кельтскую лингвистические гипотезы. При этом опровергая другие, альтернативные гипотезы: индо-греческую и итало-германскую. Которые предполагали тесную связь степных популяций, говоривших на германских и индоиранских языках с культурой шнуровой керамики.
Стоит отметить, что полученные результаты, согласуются и с изотопными данными, где среди 224 древних людей из Кипра, Греции, Италии и Испании было выявлено 56 человек как потенциально неместных.
А теперь более подробно…
До романизации Италия отличалась значительным языковым разнообразием, на её территории проживали представители как индоевропейских, так и неиндоевропейских языковых семей.
Италийские языки, в том числе латинский, оскский, умбрский и, возможно, венетский, имеют индоевропейское происхождение, так же, как и более отдаленные цизальпинский галльский, мессапский и греческий.
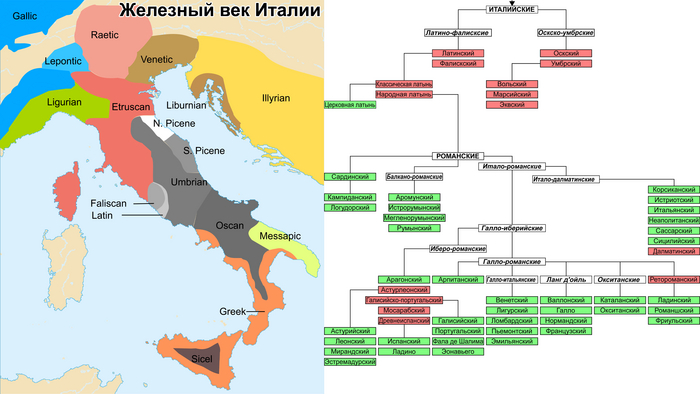
С другой стороны, этрусский и ретский языки относились к так называемой тирренской языковой семье. Из-за этого многообразия проследить распространение италийских языков, которые впоследствии стали доминирующими на Апеннинском полуострове, представляется непростой задачей.
Археологические интерпретации по-разному связывают это распространение с культурами ремеделло, ринальдоне, террамар, протовилланова и вилланова. Однако самые ранние прямые доказательства присутствия италийского были обнаружены в надписях на древнелатинском и умбрском языках, возрастом 2650 лет.
Предыдущие генетические исследования датировали появление степной родословной в Северной Италии примерно в 4000 году, а в Центральной Италии — около 3600 года. По данным из этого исследования, степная родословная появилась в Центральной Италии столетием ранее, у двух недавно обнаруженных людей из Лация (Пиан-Султано), предположительного эпицентра распространения латинского языка.

Согласно сегментам ДНК, идентичным по происхождению и моделированию примесей, степная родословная этих людей, наряду с родословной опубликованных представителей позднего бронзового века из Гротта Регина Маргерита и Топпо Дагуццо, генетически ближе к представителям культуры колоколовидных кубков, чем к ямной, подобно кельтоязычным популяциям Западной Европы.
Вышесказанное, а также широкое распространение этой родословной на протяжении бронзового и железного веков, согласуется с итало-кельтской гипотезой.
Вопрос о том, когда и как греческий язык появился в Греции, традиционно вызывает множество споров. В Греции позднего бронзового века письменность впервые появляется в виде минойского линейного письма А (около 3800-3400 лет назад) и микенского линейного письма Б (около 3350-3150 лет назад).
На Кипре около 3500–3300 лет назад появляется кипро-минойская слоговая письменность, которая представляет собой местную версию линейного письма А, и производная от нее кипрская слоговая письменность, примерно 3000–2300 лет назад.
В то время как линейное письмо А остается в значительной степени нерасшифрованным, линейное письмо Б, а также кипрская слоговая письменность, как было показано, представляют собой самые ранние письменные свидетельства греческого языка.
Ранее было установлено, что степная родословная уже была на севере Греции 4200 лет назад, указывая на связь с Причерноморско-Каспийской степью. При этом анализ показал, что эта степная родословная происходит непосредственно от популяций, связанных с ямной культурой, и отличается от популяций культуры шнуровой керамики, которые сформировались благодаря контакту ямной культуры с культурой шаровидных амфор и широко распространились по большей части Европы. Кстати, по мнению Дэвида Райха, основная часть генофонда основателей культуры шнуровой керамики была заменена благодаря женщинам ямной культуры и шаровидных амфор. Сохранив при этом гаплогруппу Y-хромосомы R1a.
Интересно, что у людей, живших примерно до 3800 года, в том числе у мужчины из Арголиды на Пелопоннесе, который жил примерно в то же время, доля предков, связанных с ямной культурой, была выше.
Это открытие является самым убедительным доказательством проникновения на Пелопоннес степных групп, которые могли быть носителями греческого языка.
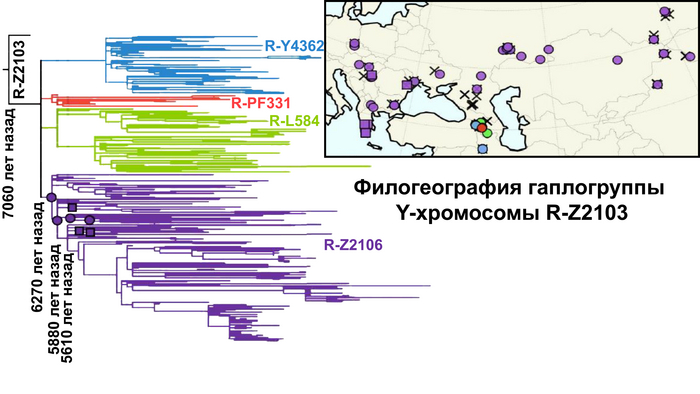
Таким образом, появление степных предков в этом регионе предшествовало возникновению древнейших прямых свидетельств существования греческого языка в виде линейного письма Б, которое к тому времени уже достигло своего начального уровня развития.
В то же время дописьменные контакты с местным населением, возможно, нашли своё отражение в том, что греческий язык обогатился так называемой «догреческой» лексикой, которая свидетельствует о взаимодействии с языками, не относящимися к индоевропейской семье.
На Кипре аркадо-кипрский диалект древнегреческого языка был распространен, по крайней мере, с раннего железного века, наряду с финикийским и одним или несколькими неизвестными языками, засвидетельствованными в кипро-минойской и кипрской слоговой письменности.
Генетические связи между населением Кипра в эпоху бронзы и раннего железного века с Левантом и Эгейским регионом также свидетельствуют о разнообразии населения.
Степное происхождение, обнаруженное у некоторых жителей Хала Султан Текке и Лапитоса на Кипре, указывает на принадлежность к позднеэлладским (то есть микенским) популяциям Пелопоннеса.
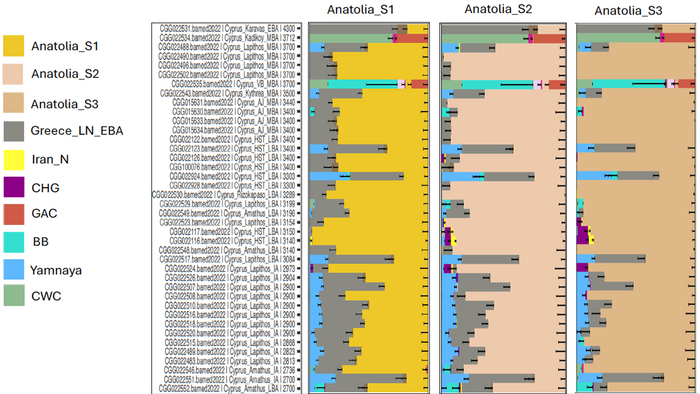
Это согласуется с появлением микенской керамики, импортируемой в позднем бронзовом веке, и с лингвистической классификацией аркадо-кипрского диалекта древнегреческого языка как потомка того же южно-греческого диалекта, что и микенский, в отличие от других греческих диалектов, таких как дорийский и ионический.
Однако еще один человек из Лапитоса, хотя и был захоронен 4100–4000 лет назад, уже группируется с популяциями Греции позднего бронзового века, что предполагает связь с популяциями Эгейского моря, подвергшимися влиянию степей ещё до возникновения микенской цивилизации.
Армянский язык, впервые засвидетельствованный около 1550 лет назад, исторически был распространен на Южном Кавказе и в восточной части Малой Азии. Ранее считалось, что он возник там примерно 3100 лет назад.
В позднем железном веке большая часть территории этого региона входила в состав древнего государства Урарту.
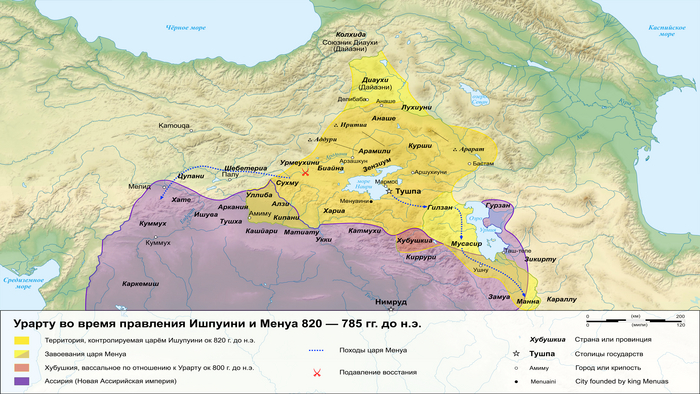
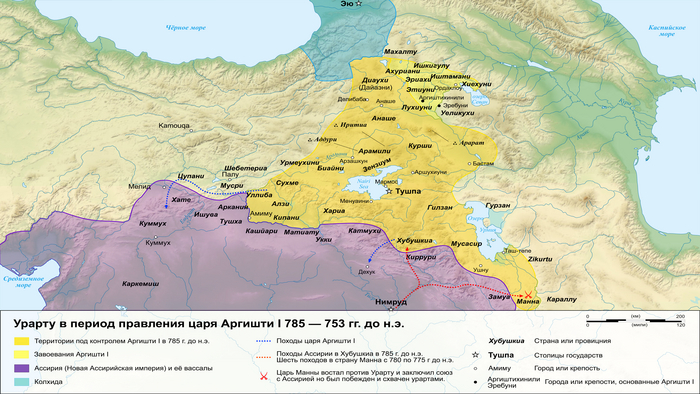
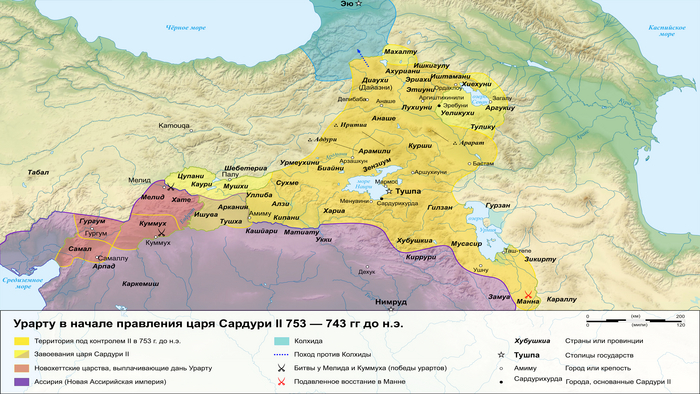
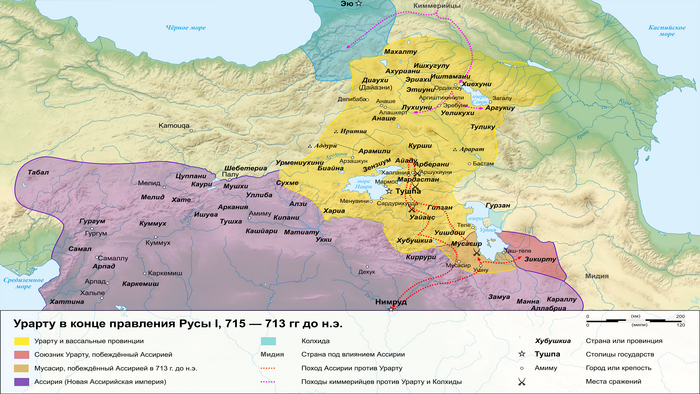
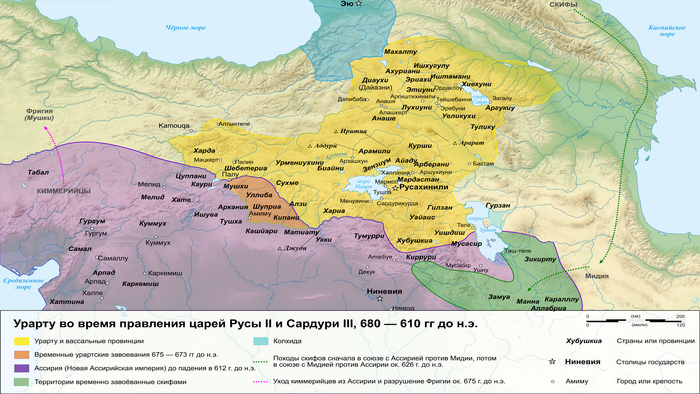
Это государство отличалось культурным разнообразием и, возможно, включало в себя элементы армянского языка. Об этом свидетельствует обмен лексикой между урартским и ранним армянским языками. Это предположение подтверждается наличием степной родословной у людей, захороненных в доурартских и урартских контекстах.
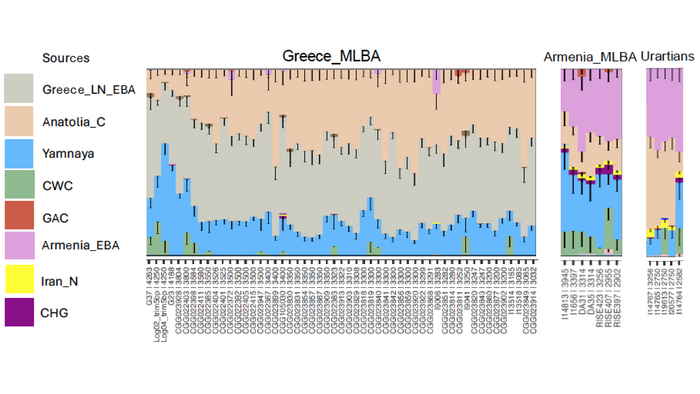
Ранее на Южном Кавказе была обнаружена степная родословная в эпоху средней бронзы, что совпадает с переходом от куро-араксской к триалетской культуре, который произошел в конце III тысячелетия до нашей эры.
Теперь учёные подтверждают, что эти люди, как и их предки в Урарту, имели степное происхождение от популяции западной ямной культуры, как и люди, населявшие Эгейский регион во II тысячелетии до н. э. Эти результаты подтверждают лингвистическую греко-армянскую гипотезу и позволяют предположить, что предшественник армянского языка появился на Кавказе в конце III тысячелетия до н. э.
Кроме того, важно отметить, что гипотезы Геродота о непосредственных связях армян с народами Балканского полуострова не нашли подтверждения в ходе нескольких исследований. Однако был выявлен факт смешивания с левантийско-раннеземледельческим населением, которое происходило до, во время или после окончания позднего бронзового века.
В эпоху средней бронзы возникли различные культуры, преобладал кочевой образ жизни, а также были установлены активные связи с Малой Азией и Эгейским регионом. Поздний бронзовый век, напротив, характеризовался интенсивным культурным взаимодействием с соседними народами, включая хурритов, хеттов и жителей Месопотамии.
Популяции культуры колоколовидных кубков оказали значительное влияние на генетический состав и культуру Пиренейского полуострова. Однако в Италии ситуация была более сложной.
На северо-востоке Италии группы культуры колоколовидных кубков, вероятно, были немногочисленны, но некоторые из них были захоронены в курганах, что привлекло внимание исследователей. Однако в период раннего бронзового века и на рубеже среднего бронзового века появились доказательства связи между Центральной Европой и Италией. Эти доказательства проявляются в распространении кинжалов треугольной формы, которые часто встречаются в кладах. Такие кинжалы были распространены по всей Италии, и их появление совпало с широким распространением родословной, связанной с культурой колоколовидных кубков, по всему Апеннинскому полуострову примерно 3800–3500 лет назад.
В долине реки По, в регионе Террамаре, наблюдался значительный рост населения, что свидетельствует о притоке людей из соседних регионов. Археологические находки указывают на тесные связи с Венгрией и регионами к северу от Альп, хотя эти связи стали очевидными только позже, в железном веке. Это свидетельствует о том, что Северная Италия выступала в роли культурного «плавильного котла», который впоследствии оказал влияние на другие части Италии.
После 1200 года до нашей эры археологически зафиксирован заметный отток населения, особенно из южной части региона Террамаре.
Дальше на юг вдоль побережья Адриатического моря, в центральной и южной частях итальянского побережья, три образца демонстрируют схожесть с популяциями на Балканах. Это указывает на продолжительные контакты между обеими сторонами Адриатики, которые сохранялись на протяжении III и II тысячелетий до нашей эры.
Эти связи не ограничивались только Адриатикой, а простирались до Эгейского моря, о чём свидетельствует присутствие микенской керамики и других товаров в поселениях вдоль итальянского побережья.
Генетические профили людей также указывают на небольшие миграции, возможно, с участием ремесленников и торговцев. Или же практика экзогамии могла способствовать наблюдаемой генетической изменчивости в этом регионе.
В Греции наблюдается иная картина. Около 3800 лет назад как на континенте, так и на Пелопоннесе можно было заметить следы степного культурного влияния, которое стало доминирующим в местах захоронений в последующие столетия.
Появление микенской культуры и греческого языка было интерпретировано по-разному. Предлагались различные сценарии: от постепенного мирного проникновения до более быстрого установления политического контроля. Однако новые генетические данные не опровергают ни один из сценариев.
После того как раннеэлладское общество распалось в короткий среднеэлладский период, с севера прибыли новые мигранты. Они укрепили свой политический контроль с наступлением позднеэлладского периода и формированием ранней микенской культуры около 3700 лет назад.
Этот процесс также сопровождался культурными и языковыми столкновениями между пришлым и местным населением.
Со временем первоначальный степной вклад стал менее значительным из-за смешения с местным населением, которое в основном происходило от ранних европейских земледельцев. Археологические находки указывают на тесные связи с генетическим источником, расположенным в Молдове и прилегающем Карпатском регионе. Эти связи нашли отражение в появлении степных лошадей и колесниц, а также в торговле серебром из карпатских рудников.

Таким образом, давний спор о происхождении микенской культуры может быть решен, по крайней мере частично. Генетические связи с Малой Азией и Критом сохранились, отражая культурные влияния, которые сформировали микенское общество. Более того, микенская культура демонстрирует удивительное сходство с более ранней триалетской культурой Южного Кавказа, которая, вероятно, способствовала процессам, лежащим в основе последующего этногенеза армян.
В бронзовом веке на Южном Кавказе заметно появление новой элиты в современной Грузии и Армении, регионах, богатых полезными ископаемыми. Эта элита была связана как со степными традициями погребения в курганах, так и с хеттской цивилизацией в Малой Азии.
Богатые курганные захоронения триалетской культуры демонстрируют параллели как с хеттскими городами-государствами, так и ранними микенскими шахтовыми гробницами.
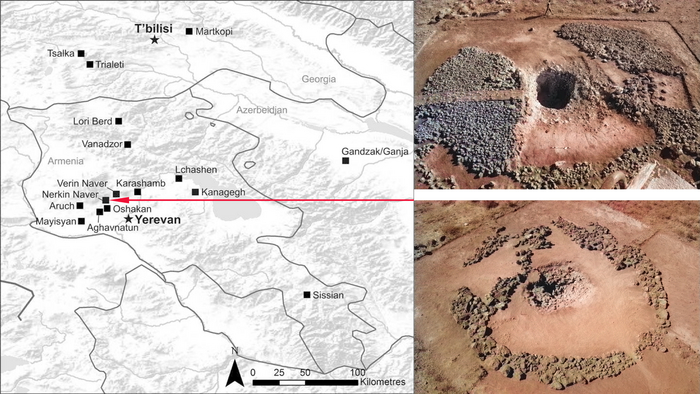
Это свидетельствует о формировании новых торговых и военных связей между степными народами и ближневосточными цивилизациями, что теперь подтверждается и генетическими данными.
Генетические данные, полученные на Кипре, подчёркивают его важную роль в качестве торгового центра благодаря обилию медных рудников, существовавших с эпохи энеолита до железного века.

Более того, эти данные свидетельствуют о тесных связях острова с Западной Малой Азией и Эгейским регионом. Поздний бронзовый век стал пиком добычи меди на Кипре и торговли ею по всему Средиземноморью, способствуя процветанию культуры в этом обширном регионе. Что также отражено и в генетических данных, которые указывают на связи между Кипром и регионами Восточного и Западного Средиземноморья.
К началу II тысячелетия до нашей эры археологические свидетельства также указывают на тесные связи между унетицкой культурой и Восточным Средиземноморьем, включая Кипр. Эта связь может быть подтверждена генетическими данными, но пока она основана лишь на анализе ДНК одного человека, и её историческое значение остается неясным.
В исследовании впервые удалось последовательно объединить данные древней ДНК, изотопов стронция, лингвистики и археологии, чтобы подтвердить двойственную модель расхождения италийского, греческого и армянского языков в эпоху бронзы.
В частности, было показано, что степное происхождение населения Испании, Франции и Италии было опосредованным через популяции колоколовидных кубков в Западной Европе, что, вероятно, способствовало появлению италийских и кельтских языков. В отличие от них, армянские и греческие популяции приобрели степное происхождение непосредственно от ямной культуры Восточной Европы. Эти результаты согласуются с греко-армянской и итало-кельтской лингвистическими гипотезами, объясняющими происхождение большинства средиземноморских индоевропейских языков античности.
Однако, в отличие от этого, результаты не подтверждают конкурирующие итало-германскую и индо-греческую гипотезы, поскольку степное происхождение у населения исторически германо- и индоираноязычных областей было охарактеризовано как связанное в первую очередь с культурой шнуровой керамики. Таким образом, выводы авторов согласуются с конкретными моделями лингвистического разделения для индоевропейской языковой семьи, в то же время противореча другим. Это подчеркивает важность древней ДНК в раскрытии доисторического разнообразия человеческих популяций и языковых сообществ.
Источник:
Ancient genomics support deep divergence between Eastern and Western Mediterranean Indo-European languages. In bioRxiv. https://doi.org/10.1101/2024.12.02.626332
Yediay, F. E., Kroonen, G., Sabatini, S., Frei, K. M., Frank, A. B., Pinotti, T., Wigman, A., Thorsø, R., Vimala, T., McColl, H., Moutafi, I., Altinkaya, I., Ramsøe, A., Gaunitz, C., Renaud, G., Mederos Martin, A., Demeter, F., Scorrano, G., Canci, A., … Willerslev, E. (2024).
Дополнительно:
Demographic history and genetic variation of the Armenian population
Hovhannisyan, Anahit et al. https://doi.org/10.1016/j.ajhg.2024.10.022
The American Journal of Human Genetics, Volume 112, Issue 1, 11 – 27
Если ютуб не работает видео из ВК:
Ашкеназские евреи, по названию Германии в средневековой раввинской литературе, представляют большинство ныне живущих евреев и появились в результате расселения средиземноморских евреев на севере Рейнской области в раннем средневековье (земли Рейнланд-Пфальц и Северный Рейн-Вестфалия). Между XIII и XV веками популяция ашкеназских евреев существенно расширилась как географически, сначала в Восточной Европе, а после и за её пределами, так и численно, около 10 миллионов в настоящее время.
Кстати, этот выпуск сделан при поддержке потомков семьи Каценеленбоген, жившей в XIX - начале ХХ века в местечке Любавичи, и чей дальний родственник по мужской линии упоминается в исследовании как "Эрфурт 14847".
Начиная с первых дней развития генетики человека, у ашкеназских евреев были выявлены десятки патогенных вариантов в генах, связанных с аутосомно-рецессивными заболеваниями, что привело к разработке успешных программ скрининга на этапе планирования беременности. Значительная часть этих вариантов чрезвычайно редка за пределами групп ашкеназских евреев и появляется на фоне давно распространённых гаплотипов, что подразумевает, что ашкеназские евреи произошли от небольшой предковой популяции - основателей.
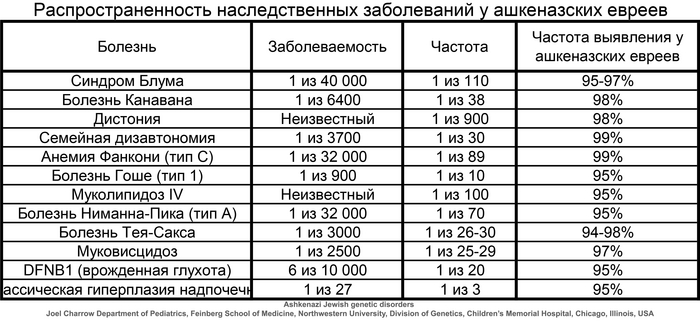
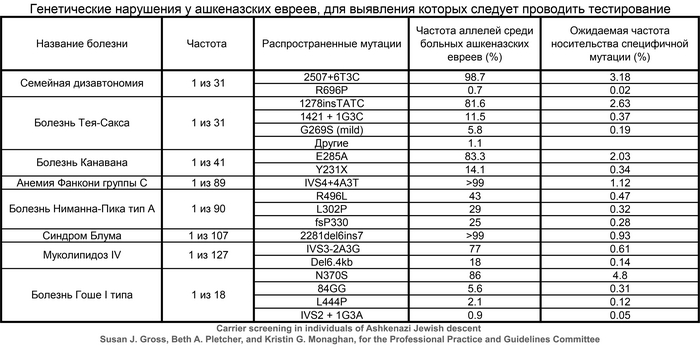
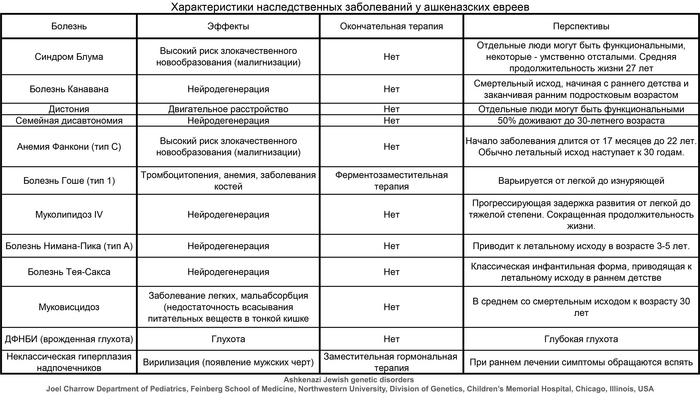

Совсем недавно исследования выявили высокий уровень родства у ашкеназских евреев на основе сегментов ДНК идентичных по происхождению (IBD), присутствующих у не связанных близким родством людей, что является признаком происхождения от довольно небольшой группы. О чём также свидетельствует наличие 4 митохондриальных линий у 40% ашкеназских евреев, которые происходят из доисторической Европы, как и большая часть менее распространённых линий.
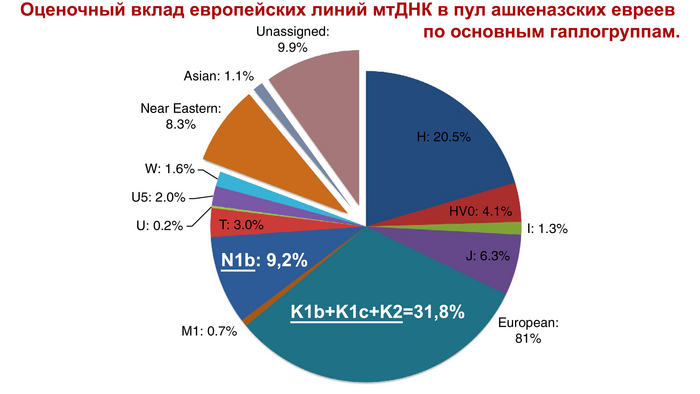
Таким образом, подавляющее большинство ашкеназских предков-основателей по материнской линии были не из Леванта или Кавказа, а из Европы.
Помимо этого, количественное моделирование показало, что ашкеназские евреи пережили резкое сокращение численности (эффект «бутылочного горлышка»), уже после формирования популяции, в позднем средневековье и что эффективный размер их популяции после этого исчислялся сотнями. Здесь стоит учитывать и эффекты основателя, и эффект бутылочного горлышка, которые произошли последовательно.
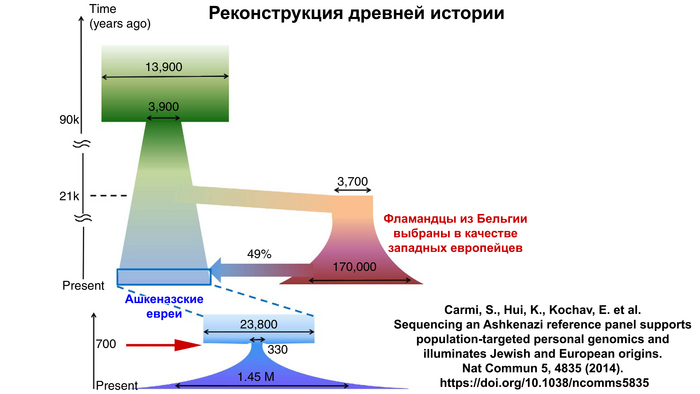
Происхождение ранних ашкеназских евреев, а также история событий смешения, которые сформировали их генофонд, являются предметом споров.
Генетические данные подтверждают смешанное ближневосточное и европейское происхождение ашкеназских евреев, указывая на то, что они имеют промежуточную родословную, между населением Ближнего востока и Европы, около 40% ближневосточной и 60% европейской.
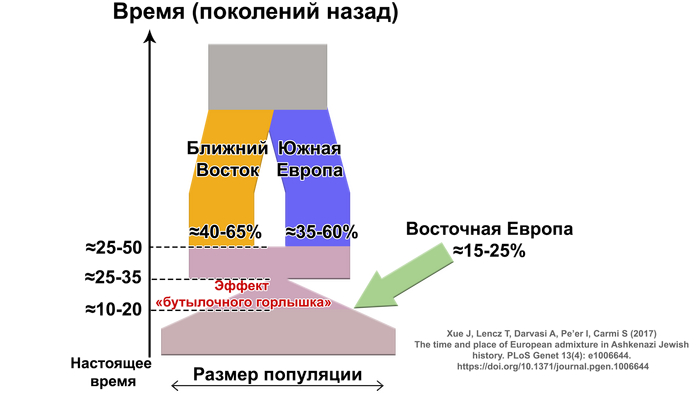
Это основано на однородительских маркерах в виде гаплогрупп из обоих регионов, а также на аутосомных данных.
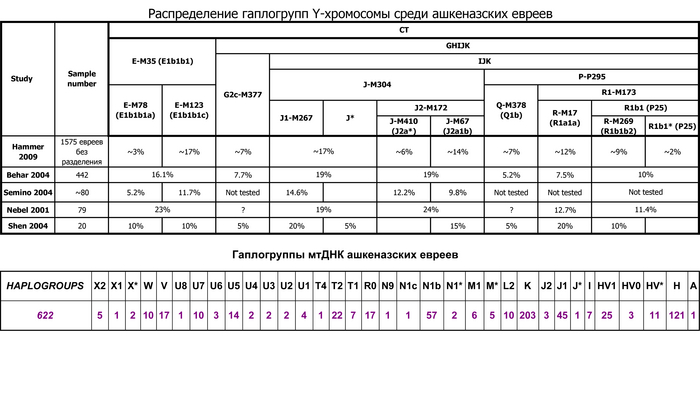
Эти и другие аутосомные исследования также показали, что ашкеназские евреи генетически отличаются от других. Недавнее моделирование показало, что ≈60–80%, т.е. большая часть, из 50-70% европейской родословной у ашкеназских евреев согласуется с южноевропейскими источниками.
Популяция ашкеназских евреев в целом генетически довольно однородна, с небольшими различиями в зависимости от современной страны проживания, что условно разделяет их на восточноевропейских и западноевропейских.

При этом, несмотря на недавний прогресс в изучении генофонда ашкеназских евреев, некоторые демографические вопросы всё же остаются открытыми, в том числе о времени, месте основания популяции и маршрутах распространения после сокращения численности, а также о времени, месте и источниках примеси. Изучение геномов ашкеназских евреев, живших ближе ко времени образования их популяции, может пролить свет на эти вопросы. В своей работе авторы исследовали ДНК средневековых ашкеназских евреев XIV века из Эрфурта в Германии, где они проживали с конца XI века по 1454 год, с небольшим перерывом после, мягко говоря, печальных событий в Страсбурге 1349 года и последующих погромов до 1351 года в странах Средневековой Европы во время чумы.
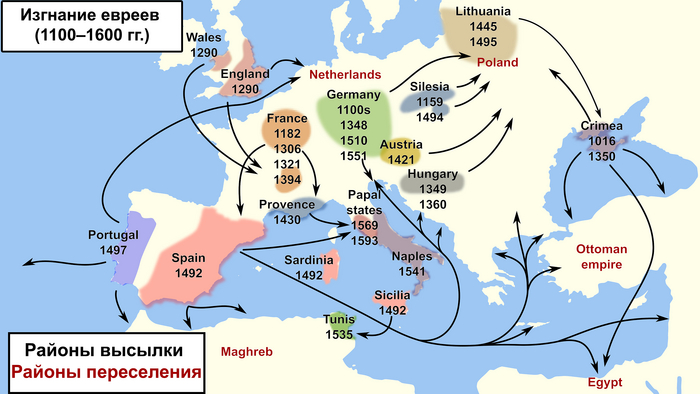
Первая еврейская община Эрфурта, существовавшая до 1349 года, была старейшей в Тюрингии, а её кладбище также использовалось жителями соседних городов. Во время погрома 1349 года большинство евреев Эрфурта и близлежащих общин были убиты или изгнаны. Евреи вернулись в Эрфурт в 1354 году, чтобы основать вторую общину, которая была одной из крупнейших в Германии. После очередного изгнания евреев в 1454 году, на еврейском кладбище было построено зернохранилище (в 1465-1467 годах). В 2013 году зернохранилище пустовало, и город разрешил переоборудовать его в парковку. Это потребовало дополнительного строительства и проведения археологических спасательных раскопок. Люди, которых исследовали в данной работе, были похоронены в юго-западной части средневекового еврейского кладбища. Археологические находки позволяют предположить, что это место использовалось второй общиной.

Радиоуглеродный анализ десяти человек показал, что все они жили примерно между 1270 и 1400 годами, но данные не были информативными относительно того, использовалось ли это место первой или второй общиной. Предполагаемый возраст умерших варьировался от 5 до 60 лет, при этом 14 из 33 (42%) были моложе 20 лет. Причину смерти точно удалось установить только для одного человека (I14904), который был убит несколькими ударами по голове острым предметом.
Исследование предков представителей современных религиозных групп предполагает определённые трудности. Еврейские законы, которым следовали ашкеназские евреи Эрфурта, запрещают беспокоить умерших, включая эксгумацию для большинства целей. Но исследователям удалось найти компромисс с раввинами при условии, что будут использованы только отдельные зубы, взятые из уже извлечённых на тот момент 38 останков. Однако данные по всему геному, прошедшие контроль качества, были получены для 33 человек: 19 женщин и 14 мужчин, со средним количеством однонуклеотидных полиморфизмов (SNP) в 383 тыс., охваченных, по крайней мере, одной последовательностью. Обратите внимание, что это значительно больше, чем по половым хромосомам.
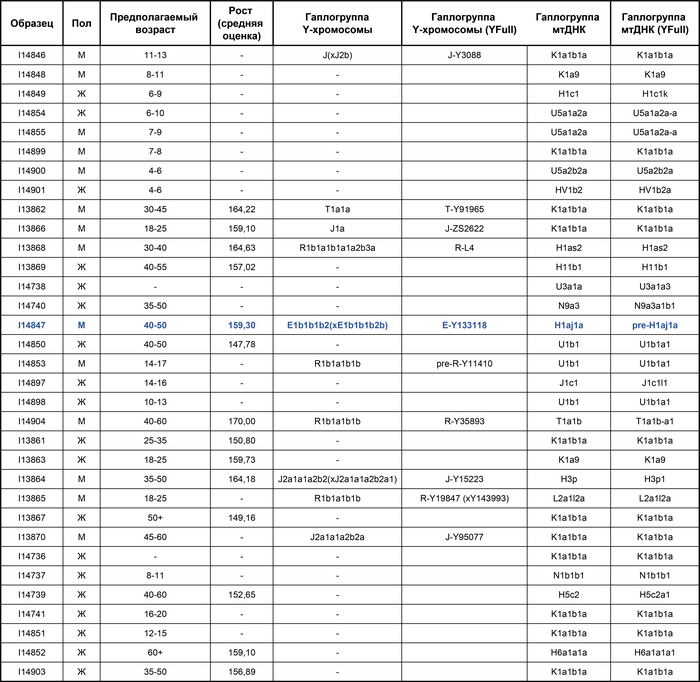
При этом следов бактерий чумной палочки (Yersinia pestis) в последовательностях ДНК не было обнаружено.
Помимо этого, в исследовании также были выявлены патогенные варианты генов.
Среди изученных людей было выявлено две группы близких родственников, одна включала мать с сыном и дочерью, где дети были похоронены в трёх рядах от матери, ориентация могилы которой была противоположной всем остальным захоронениям. А вторая группа родственников состояла из похороненных рядом, упомянутого ранее мужчины, убитого ударом по голове, и его дочери.
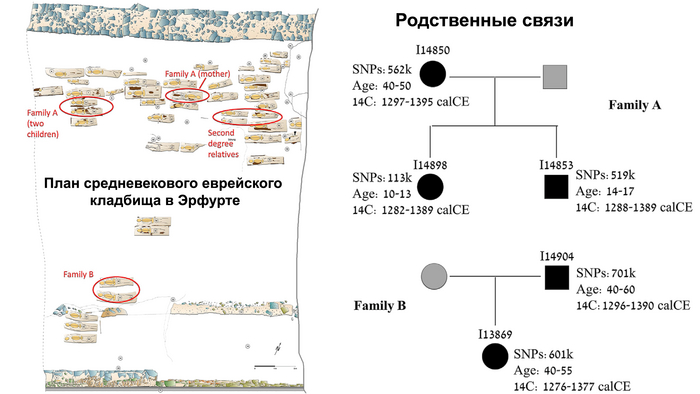
Также было выявлено три похороненных рядом человека, степень родства которых точно установить не удалось из-за низкого количества полученных ДНК-маркеров. Однако у двоих из них была определена линия мтДНК U5a1a2a, не встречавшаяся у других людей из выборки.
На графике анализа главных компонент ашкеназские евреи из Эрфурта пересеклись с современными ашкеназскими евреями, а также с другими европейскими средиземноморскими народами.
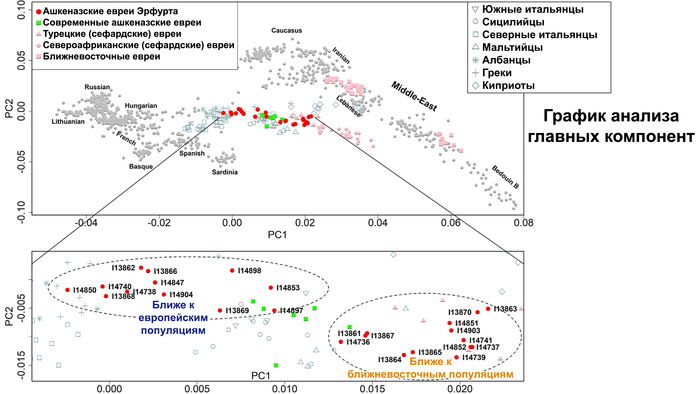
Интересно, что средневековые ашкеназские евреи из Эрфурта отличались от современных ашкеназских евреев большим разбросом по линии Европа – Ближний Восток. Авторы разделили ашкеназских евреев Эрфурта на две группы в зависимости от близости к европейским и ближневосточным популяциям. Дополнительные анализы, методика которых описана в оригинале работы, подтвердили обоснованность такого разделения на 2 отличающиеся группы в отличие от непрерывного градиента изменчивости. Поочерёдное сравнение двух групп ашкеназских евреев из Эрфурта с современными ашкеназскими евреями восточноевропейского или западноевропейского происхождения показало, что современные западноевропейские их популяции почти полностью пересекались со средневековыми популяциями Эрфурта из группы, приближённой к ближневосточным популяциям, которые также пересекались и сефардскими евреями Турции.
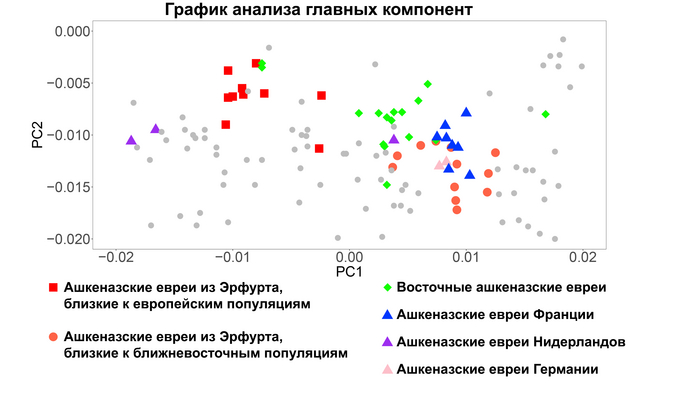
А современные ашкеназские евреи восточноевропейского происхождения занимали промежуточное положение между двумя группами средневековых евреев Эрфурта, но ближе к группе, приближённой к ближневосточным популяциям.
Дальнейшие тесты, включая f4 статистику, показали, что ашкеназские евреи из Эрфурта, близкие к европейским популяциям, имеют больше европейской родословной, особенно восточноевропейской, чем средневековые ашкеназские евреи из Эрфурта, близкие к ближневосточным популяциям и современные ашкеназские евреи. Изотопный анализ эмали зубов подтвердил различия между двумя группами. Они были значительными для соотношения стабильных изотопов кислорода-18, что свидетельствует о различиях в источниках воды у представителей этих двух групп Эрфурта в детстве.
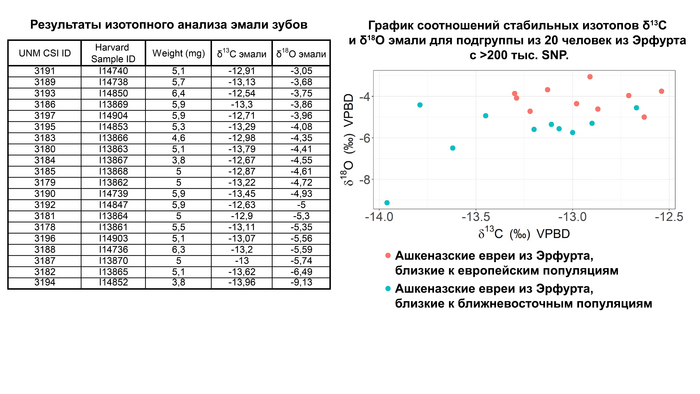
Тем не менее, несмотря на различия не наблюдается признаков культурного или временного разделения на свой — чужой.
При этом обе группы из Эрфурта ближе к современным ашкеназским евреям, чем к какой-либо другой еврейской группе.
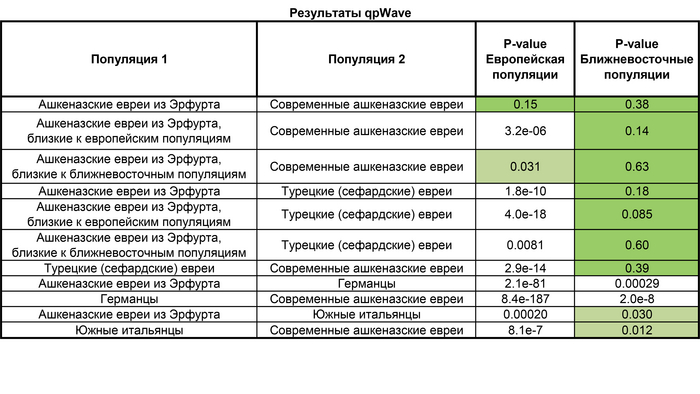
Это генетическое сходство несмотря на то, что они жили с разницей в 600–700 лет, предполагает высокую степень эндогамии в течение этого периода. То есть браки они заключали преимущественно в рамках своей субэтнической группы. А близость к популяциям Восточной Европы средневековых евреев Эрфурта из группы близкой к европейцам, может быть связана с исторически зафиксированной миграцией семей из Богемии, Моравии и Силезии во вторую общину Эрфурта после 1354 года.
Примечательно, что согласно данным анализа ДНК шести человек, найденных на дне средневекового колодца в британском Норвиче, которые были одними из жертв эпизода антисемитского насилия 6 февраля 1190 года, они генетически были сильно близки к современным ашкеназским евреям, а их родословная моделировалась как смесь ∼33% сефардских евреев и ∼67% сицилийцев, без восточноевропейской примеси, что также предполагает её появление у ашкеназских евреев не ранее 13 века.
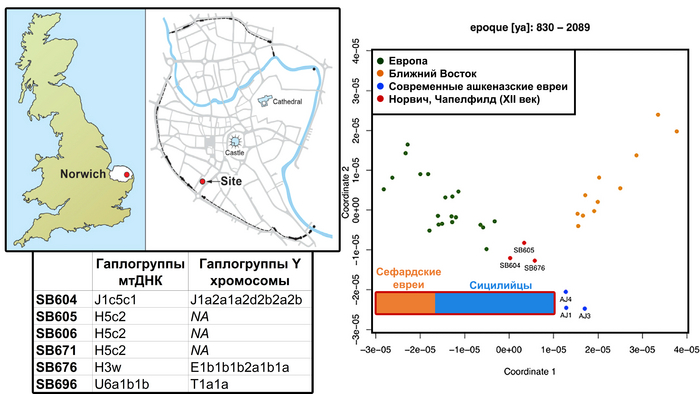
А любое гипотетическое событие смешения между ашкеназскими евреями и жителями Восточной Европы за последние ≈20 поколений должно было быть ограничено заменой не более 2%–4% от их общего генофонда, т.е. не более 0,2% на поколение.
Помимо этого, тестовые сравнения популяций ашкеназских евреев как средневековых, так и современных с популяциями Ближнего Востока, включающих и турецких евреев, позволяют предположить, что эти группы имеют схожие источники ближневосточного происхождения.
В работе авторы рассмотрели модель, в которой средневековые ашкеназские евреи из Эрфурта представляют собой смесь следующих источников: южноевропейского (включающего южных или северных итальянцев), ближневосточного (включающего друзов, египтян, бедуинов, палестинцев, ливанцев, иорданцев, сирийцев, или саудовцев) и восточноевропейского (представленного русскими).
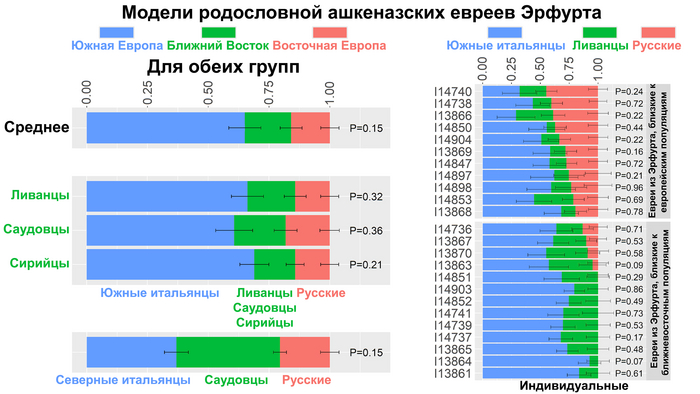
В качестве источников авторы использовали современное население, поскольку моделирование с использованием древних источников было неудачным. Поэтому эти модели надо адекватно воспринимать, они отражают генетические связи между предковыми источниками ашкеназских евреев и современных популяций.
Многочисленные модели с участием южных итальянцев оказались правдоподобными, что согласуется с историческими данными, указывающими на Итальянский полуостров как на источник популяций ашкеназских евреев.
Средние пропорции примесей у обеих групп представителей средневековья по всем вероятным моделям, составили 65% вклада популяций Южной Италии, 19% Ближнего Востока и 16% Восточной Европы.
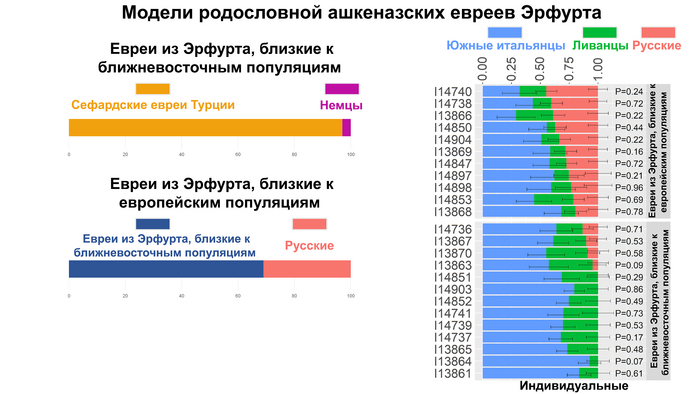
А по отдельности, ашкеназских евреев Эрфурта, близких к популяциям Ближнего Востока, можно смоделировать как смесь 97% сефардских евреев из Турции и 3% немцев, а представителей группы средневековых евреев близкой к европейским популяциям, можно смоделировать как смесь 69% эрфуртских ашкеназских евреев, близких к популяциям Ближнего Востока и 31% русских. При этом в индивидуальных моделях величина восточноевропейского вклада была сильно изменчива у ашкеназских евреев Эрфурта. В группе близкой к популяциям Европы он в среднем составлял 33%, но не был обнаружен у 9 из 13 средневековых евреев из группы, близкой к ближневосточным популяциям.
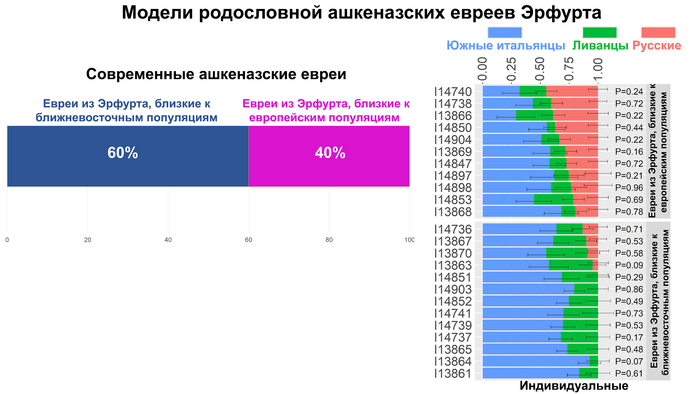
В свою очередь, современных ашкеназских евреев можно было смоделировать как смесь 60% средневековых ашкеназских евреев Эрфурта, близких к популяциям Ближнего Востока, и 40% представителей другой группы, близкой к европейским популяциям.
В совокупности результаты показывают, что группа Эрфурта, близкая к популяциям Ближнего Востока, генетически близка и к сефардским евреям, а группа, близкая к Европейским популяциям, получила дополнительную восточноевропейскую примесь. А современные ашкеназские евреи сформировались в результате почти равномерной смеси этих двух групп.
Лингвистические, культурные и ономастические, т.е. изучающие любые собственные имена, исследования выявили различия между ашкеназскими евреями из Западной Европы, представляющими ранние общины Рейнланда, и ашкеназскими евреями из Центральной и Восточной Европы. Эти группы называли по их способу произношения восьмой буквы еврейского алфавита «Хет».
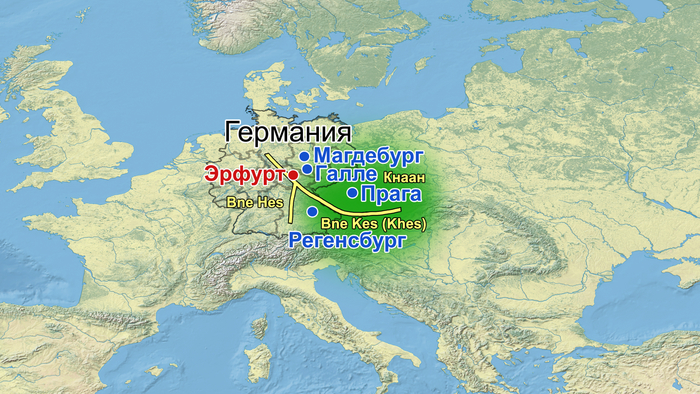
Западные группы, вероятно, произошли в основном от евреев, населявших Северную Францию и Италию непосредственно за столетия до этого. Они придерживались уникального религиозного обряда и носили имена, отличные от имён евреев, живших на Востоке, и в течение первых столетий второго тысячелетия говорили на немецких диалектах, структурно схожих с диалектами христианского большинства. В исследовании представителями западной общины, возможно, были эрфуртские евреи с большим ближневосточным происхождением и современные евреи западноевропейского происхождения. А восточные евреи населяли юго-восточную Германию, Австрию, Богемию, Моравию и Силезию, а первыми крупными поселениями были Регенсбург, Прага, Магдебург и Галле. В первые века второго тысячелетия к большей части этой территории применялся древнееврейский географический термин "Кнаан", а старочешский был основным языком, на котором говорили местные евреи, кроме самой западной части района, охватывающего Регенсбург, Австрия, и западные общины Восточной Германии, где Евреи говорили на немецких диалектах. Если эта группа представлена эрфуртскими евреями с большим европейским происхождением, то они ранее смешивались с местным нееврейским населением. Эрфурт при этом, возможно, находился на границе между двумя общинами ашкеназских евреев. Исследования имён, диалектов и религиозных обрядов современных ашкеназских евреев позволяют предположить, что западная и восточная общины в итоге объединились и сформировали единую ашкеназскую культуру, определяемую единым религиозным обрядом и языком идиш. Доступные списки еврейских мучеников из Эрфурта (1221 год) и соседнего Вайсензея (1303 год) показывают наличие имён, типичных для западных ашкеназских евреев.


Эти евреи или их предки мигрировали в Тюрингию из Рейнской области. Тем не менее, в источниках из Эрфурта второй половины XIV века многочисленные славянские имена указывают на присутствие там восточных ашкеназских евреев. Эти соображения могут объяснить, почему выборка только из одного участка кладбища в Эрфурте позволила выявить структуру средневековых ашкеназских евреев.
При этом негенетические различия почти исчезли к XVI веку, что согласуется и с отсутствием генетических различий у современных ашкеназских евреев.
Однако, рассматривая модели родословной, стоит учитывать, что по составу современные и древние популяции отличались и пропорции примесей не следует считать точными. Модели с южноитальянским происхождением имеют правдоподобную историческую основу. Однако эти модели предполагают, что только около 20% предков ашкеназских евреев из Эрфурта имеют ближневосточные корни, меньше, чем предыдущие оценки. Это также может быть истолковано как подразумевающее, что современные ашкеназские евреи получили лишь малую часть своей родословной от древних иудеев. И если это так, то большинство ашкеназских евреев обязаны своим происхождением новообращённым европейцам. Хотя это одно из возможных объяснений, потому как сами современные итальянцы имели гораздо более высокую долю ближневосточной примеси, по крайней мере, со времён Римской империи в Европе, и это особенно характерно для современной Южной Италии.
Таким образом, альтернативное объяснение этим наблюдениям состоит в том, что истинная доля ближневосточной родословной у ашкеназских евреев выше, чем в модели, около 30-50%, а фактический вклад итальянцев не так велик, как предполагает этот анализ. Согласно обширному набору изученных моделей, ближневосточное происхождение эрфуртских ашкеназских евреев оценивается в диапазоне от 19% до 43%, а европейско-средиземноморское — в диапазоне от 37% до 65%.
В частности, уровни ближневосточного происхождения в Италии исторически были различными, а популяции Ближнего Востока также были изменчивы за последние два тысячелетия, особенно в плане африканской примеси. В противоположном сценарии ашкеназские евреи могут вообще не иметь предков из древнего Леванта. Это могло бы иметь место, если бы не итальянское население (с большей примесью от популяций Леванта, чем у современных южноитальянцев) было источником левантийских предков, наблюдаемых у ашкеназских евреев. Авторы считают, что все возможные сценарии вероятны.
Таким образом, результаты из исследования следует интерпретировать только как предположение о том, что предки ашкеназских евреев имеют связи с населением, проживающим сегодня в Средиземноморской Европе и на Ближнем Востоке.
Хорошее соответствие моделей (qpAdm) родословной ашкеназских евреев из Эрфурта, в которых её источником была Италия, особенно Южная, обеспечивает некоторую поддержку теории происхождения там их предков. Южная Италия - одно из немногих мест в Европе, где есть свидетельства еврейской демографической и культурной преемственности от позднеримского периода до раннего Средневековья и далее. В течение этого времени еврейские общины Южной Италии находились в непосредственном контакте с еврейскими общинами Палестины в византийский и раннеисламский периоды, от которых они получали литургические традиции и распространяли по Европе. А позже они появились в сборнике молитв ашкеназских евреев. Также поддерживались связи с еврейскими общинами в других частях Восточного Средиземноморья в связи с тем, что Южная Италия в конце XI века была частью Византии.
Все имеющиеся на сегодняшний день свидетельства указывают на то, что в римский период и в раннем Средневековье евреи были тесно интегрированы в сообщества Южной Италии с историческими свидетельствами того, что в Южной Италии существовал, по крайней мере, некоторый обмен генами между евреями и неевреями, поскольку в позднеримский и раннесредневековый период имперские и церковные власти пытались предотвратить практику смешанных браков между евреями и христианами, а также предотвратить обращения неевреев в иудаизм. Когда со временем авторитетные евреи с развитыми связями из Южной Италии начали переезжать на север, к ним присоединились другие евреи из Центральной и Северной Италии. Например, известно, что еврейская семья Калонимус с Итальянского полуострова оказала большое влияние на жизнь ашкеназских евреев в Майнце и Шпейере X века. Это было многослойное миграционное наследие, которое может быть отражено в генетической родословной, связанной с Южной Европой, которую можно наблюдать в моделях генофонда эрфуртских евреев. Тем не менее, греки также могут подойти для модели, основанной на средиземноморско-европейском происхождении. В эпоху эллинизма, начиная с IV века до н. э., евреи двинулись на север, в Малую Азию и Грецию, включая такие острова, как Де́лос, который в то время был крупным торговым центром. Хотя поначалу отношения между евреями и греками были напряжёнными, в римский период они активно взаимодействовали. Существует множество археологических и письменных свидетельств существования активных еврейских общин как в Греции, так и на западном побережье Малой Азии в городах, которые возникли как греческие колонии. Позже, начиная с периода ранней Римской империи, еврейские общины существовали на юге Италии и на Сицилии, особенно в Та́ранто (Апулия) и Сиракузах (Сицилия).
А данные о том, что около 15% предков ашкеназских евреев имеют восточноевропейское происхождение, согласуется с результатами предыдущего исследования. Однако эти популяции могли происходить из обширных регионов Центральной или Восточной Европы, особенно учитывая зарегистрированную миграцию в Эрфурт из Богемии, Моравии и Силезии.
Генетические данные свидетельствуют о высокой степени эндогамии среди ашкеназских евреев на протяжении последних 700 лет. Исторические свидетельства указывают на то, что практика смешанных браков между евреями и христианами не одобрялась средневековыми еврейскими и христианскими властями. Генетические данные также свидетельствуют, что на практике в еврейскую общину с этого периода поступало очень мало генов извне. Это говорит о том, что большая часть генофонда, связанного с Восточной Европой, возникла ещё до XIV века.
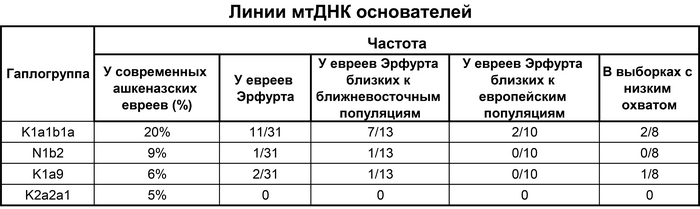
Предыдущие анализы сегментов ДНК, идентичных по происхождению, и патогенные варианты предположили, что средневековые ашкеназские евреи пережили снижение численности популяции, так называемый «эффект бутылочного горлышка». Однако детали этого события ещё не полностью выяснены. Сравнение линий мтДНК K1a1b1a, одной из четырёх наиболее распространённых линий у ашкеназских евреев, которая почти отсутствует у людей нееврейского происхождения, но была определена у 11 неродственных средневековых жителей Эрфурта, показало, что время до самого последнего общего предка составляет около 1500 лет назад, но с большими диапазонами. Причём эта линия преобладала у ашкеназских евреев Эрфурта с большим ближневосточным происхождением, что согласуется с более высокой её частотой у ашкеназских евреев западноевропейского происхождения.
В целом, результаты анализа мтДНК свидетельствуют о том, что средневековые и современные евреи испытали общее событие сокращения численности популяции. Это также подтверждает анализ участков гомозиготности (ROH) и наличие аллелей-основателей у ашкеназских евреев.
Однако с определением деталей и времени сокращения численности популяции оказалось не всё так просто. Моделирование предполагает, что сокращение численности популяции произошло около 1000 лет назад, в момент формирования ранних сообществ ашкеназских евреев. То есть его можно соотнести с эффектом основателя. Однако фактическое количество участков гомозиготности (ROH) у средневековых евреев из Эрфурта превышало ожидаемые в такой простой модели.
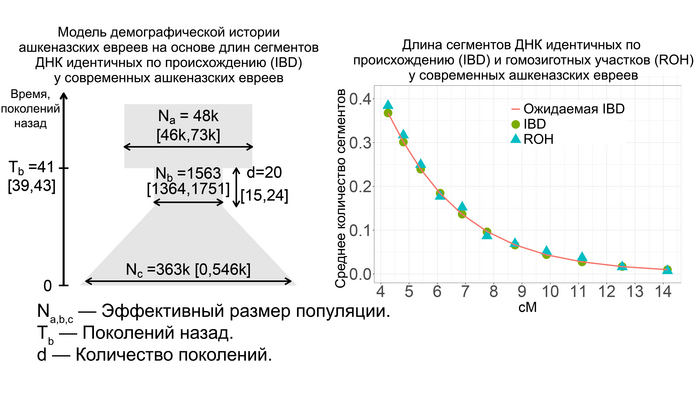
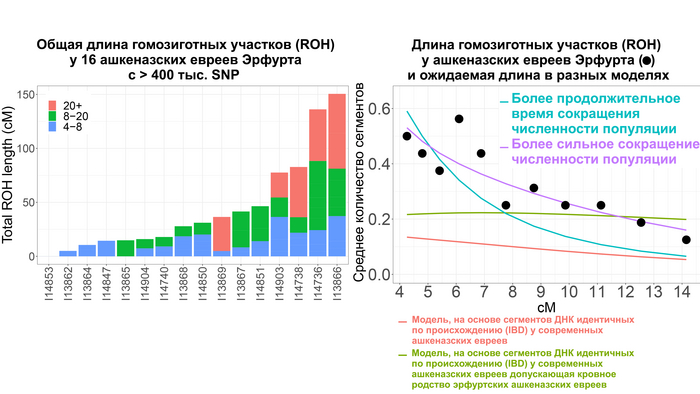
Поэтому, учитывая, что евреи Эрфурта представляют собой только один средневековый участок, авторы допустили существование ещё одной популяции ашкеназских евреев, численность которой в момент сокращения как предковой для Эрфурта популяции, так и самой эрфуртской популяции с IX по XV вв. существенно не менялась, но сократилась в конце этого процесса при слиянии с другой группой ашкеназских евреев.
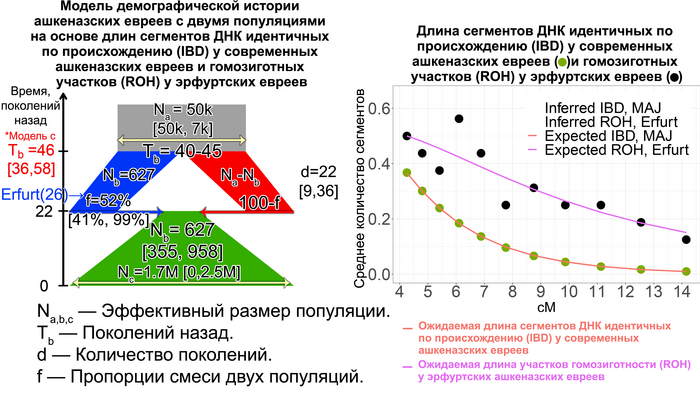
Обе группы впоследствии внесли почти равный вклад в формирование генофонда современных ашкеназских евреев. Такая модель хорошо соответствует как современным, так и древним данным.
При этом разделение между этими популяциями совпадает с началом сокращения численности предковой популяции для евреев из Эрфурта 40-45 поколений назад, или приблизительно 1000-1200 лет назад. Этот период начался задолго до того, как в конце 11-го века преследования евреев в Рейнской области стали массовыми. Сокращение численности предковой популяции на ранних этапах формирования общины ашкеназских евреев могло отражать исторические свидетельства того, что первоначальные поселенцы состояли всего из нескольких десятков семей, которые не всегда были желанными гостями и не имели преимуществ полностью развитой еврейской общины. То есть это событие можно связать с эффектом основателя. Однако увеличение численности ашкеназских евреев в моделях произошло 20-25 поколений назад или примерно 500-700 лет назад. Это подтверждает исторические данные о постепенном демографическом росте еврейской общины на землях Германии. Этот рост трудно поддаётся количественной оценке, но, особенно начиная с 1300-х годов, он, по-видимому, был значительным, учитывая быстрое увеличение числа городов, в которых проживали еврейские общины. А продолжительность времени, за которое сокращалась численность популяции около 20 поколений или 500 лет, соответствует и эффекту основателя, и эффекту бутылочного горлышка, и, возможно, повторному эффекту основателя после смешения с другой популяцией с общим первоначальным эффектом основателя.
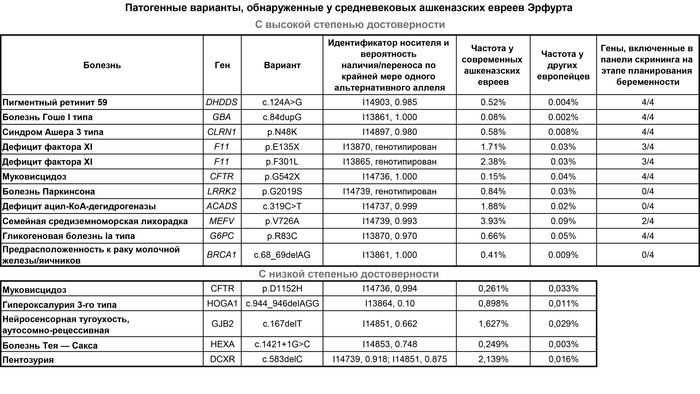
В геномах ашкеназских евреев Эрфурта было обнаружено 11 патогенных вариантов с высокой степенью достоверности и 5 с низкой. Все варианты с высокой степенью достоверности присутствовали у восьми человек из Эрфурта. Шесть из обнаруженных вариантов были ранее датированы с использованием современных геномных данных, при этом их предполагаемое время происхождения соответствует присутствию их уже в 14-м веке. Большинство этих вариантов находятся в генах, включённых в панели скрининга на этапе планирования беременности в наши дни.
Помимо этого, авторы оценили варианты, связанные с выбранными немедицинскими или полигенными фенотипами. Варианты, связанные с восприимчивостью лактозы во взрослом возрасте, пигментацией глаз, волос и (предположительно) риска чумы имели схожую частоту у средневековых и современных ашкеназских евреев. Интересно, что по сравнению с европейцами аллель восприимчивости лактозы во взрослом возрасте был менее распространён у ашкеназских евреев, 10% у современных и 11,7% у средневековых, против 60% у европейцев.
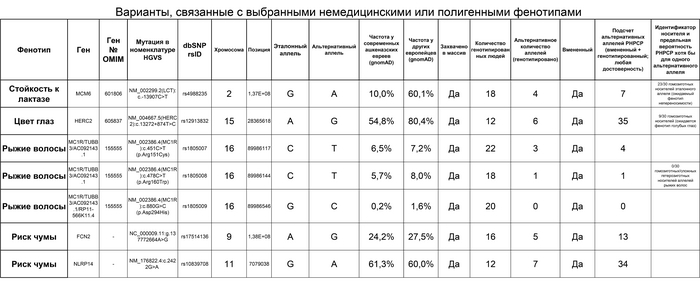
Но это различие может отражать градиент в Европе, где его частота составляет 69,8% у северо-западных европейцев и только 33,0% у южноевропейцев.
Анализируя геномные данные средневековых жителей Эрфурта, авторы работы дополнили и уточнили наше понимание происхождения ранних ашкеназских евреев. Их родословная была тесно связана с родословной современных ашкеназских евреев, что свидетельствует о высокой степени её стабильности за последние ≈700 лет. Эндогамия и смешение в пределах субэтнической группы евреев со времён средневековья, способствовали формированию наблюдаемой однородной генетической структуры ашкеназских евреев. Однако средневековые ашкеназские евреи Эрфурта имели более разнообразную генетическую структуру, чем современные, и были разделены на группы. Одна была ближе европейским популяциям, демонстрируя незначительную примесь от популяций, связанных с Восточной Европой, а другая имела родословную, близкую к таковой у современных западноевропейских ашкеназских евреев и современных сефардских евреев. Изотопные данные также указали на разное географическое происхождение этих средневековых групп. Вероятно, в позднем Средневековье население ашкеназских евреев было лингвистически и культурно разделено по оси запад-восток. Группа, близкая к ближневосточным популяциям, вероятно, представляла собой потомков первых ашкеназских евреев, поселившихся в Рейнской области. В одном источнике прямо упоминается, что Эрфурт находится на границе между двумя группами, и в XIV веке имена, данные в Эрфурте, были типичны как для западных, так и для восточных ашкеназских евреев. Эти моменты, наряду с данными о миграции во вторую общину с Востока, могут объяснить, почему в Эрфурте были обнаружены люди, принадлежащие к обеим генетическим группам.
Помимо этого, результаты не исключают ещё большего разнообразия средневековых групп ашкеназских евреев за пределами Эрфурта.
Многочисленные данные также свидетельствуют о том, что популяция эрфуртских ашкеназских евреев уже сталкивалось с общим для современных ашкеназских евреев событием сокращения численности популяции и генетического разнообразия. В соответствии с предыдущими исследованиями авторы датируют начало роста популяции ашкеназских евреев примерно в 20-25 поколений назад или 500-700 лет назад.
При этом средневековые ашкеназские евреи из Эрфурта в своей популяционной истории столкнулись с более серьёзным сокращением численности, чем современные, но их предки, вероятно, были генетически более разнообразными.
Источник:
Genome-wide data from medieval German Jews show that the Ashkenazi founder event pre-dated the 14th century Author: Shamam Waldman, Daniel Backenroth, Éadaoin Harney, Stefan Flohr, Nadia C. Neff, Gina M. Buckley, Hila Fridman, Ali Akbari, Nadin Rohland, Swapan Mallick, Iñigo Olalde, Leo Cooper, Ariel Lomes, Joshua Lipson, Jorge Cano Nistal, Jin Yu, Nir Barzilai, Inga Peter et al. Published: November 30, 2022 DOI: https://doi.org/10.1016/j.cell.2022.11.002
Европейский железный век к северу от Альп характеризуется двумя ключевыми археологическими культурами – гальштатской (800–450 гг. до н. э.) и латенской (после 450 г. до н. э. и до начала римского периода около 50 г. до н. э.), которые в разной степени описываются как «кельтские».


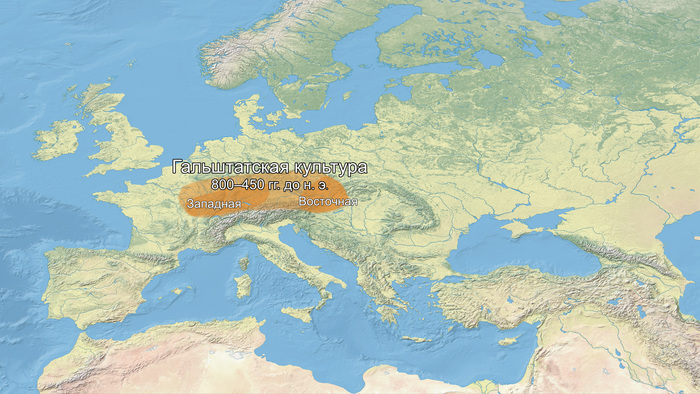
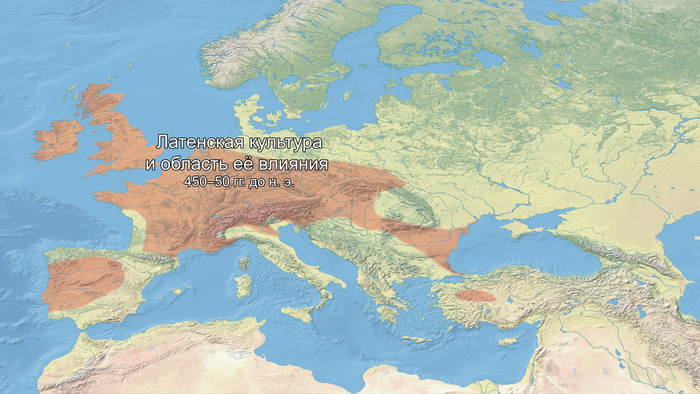
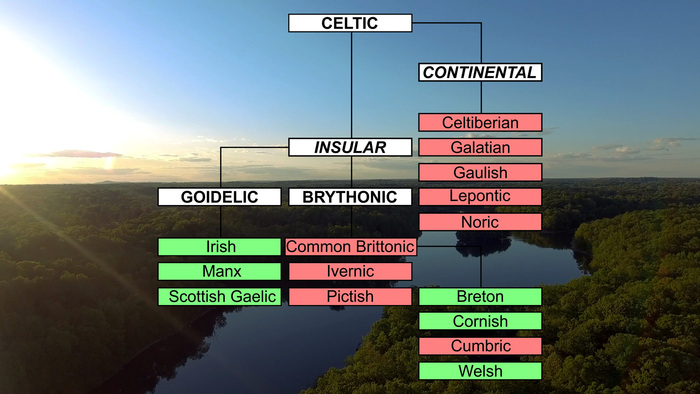
Кельты впервые упоминаются в греческих источниках конца VI в. до н. э. и это название широко используется в античных источниках для обозначения обществ, связанных с латенской культурой. Помимо этих исторических данных и их связи с гальштатской и латенской культурами, существуют также лингвистические свидетельства существования кельтских языков на значительной части Европы. Действительно, общеевропейские модели и лингвистические свидетельства культурных связей того времени сложны и охватывают обширные регионы от Пиренейского полуострова и Британских островов по всей Центральной Европе и вплоть до Малой Азии в III веке до нашей эры. В то время как более ранние исследования предполагали возникновение этого более позднего общеевропейского явления исключительно в относительно узко определенной области к северо-западу от Альп, новые перспективы предполагают модель полицентрического возникновения на обширной территории между атлантическим побережьем и юго-западной Германией.
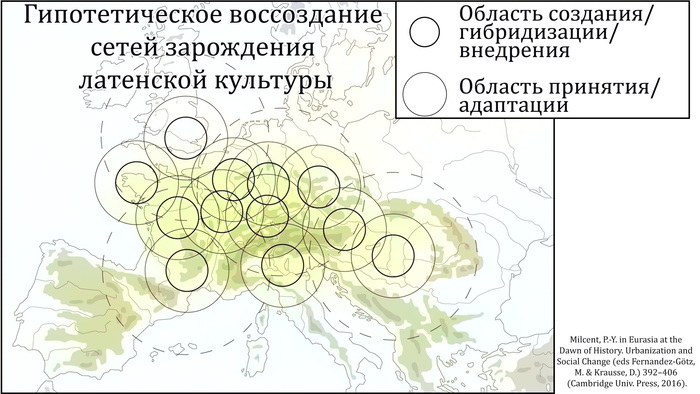
Один из этих основных регионов находился на территории современной восточной Франции, Швейцарии и юго-западной Германии. Между 600 и 400 гг. до н. э. (западногальштатская и латенская культуры) эта территория выделяется своей археологической значимостью, о чем свидетельствуют богатые захоронения «Фюрстенгребер», в переводе – Княжеские могилы.





Эти захоронения характеризуются монументальными курганами и роскошным погребальным инвентарем, таким как церемониальные повозки, элементы мебели, золотые украшения, греческие и этрусские импортированные предметы либо напитки и еда. Такой богатый погребальный инвентарь обычно указывает на высокий социальный статус погребённых. На протяжении раннего железного века воинские и сакрально-религиозные представления в этих богатых захоронениях все больше сливались воедино, объединяя мирскую и духовную власть, что, возможно, больше соответствовало сакрализации королевской власти, т.е. правителей обожествляли, в противовес простым вождям. После смерти их хоронили под внушительными памятниками и почитали как героических предков. А некоторым поклонялись как богам в больших церемониальных комплексах, таких как погребальные памятники возле Глауберга в Гессене, воздвигнутые в ранний период латенской культуры около 400 г. до н. э.
Соответственно, эти монументальные так называемые княжеские захоронения представляли собой проявление династических систем власти, в которых политическое господство, по крайней мере частично, основывалась на биологически унаследованных привилегиях, что было отличительной чертой ранних сложных обществ. Хотя природа ранней кельтской политической системы и особенно важность биологического родства, по сей день вызывает большие споры. Некоторые ученые интерпретируют этих умерших как "поселковых старейшин", которые приобрели свой высокий социальный статус при жизни благодаря личным достижениям, а не унаследовали его. Однако наличие необычайно богатых детских захоронений, свидетельствующих о высокой социальной власти и престиже, по-видимому, противоречит этой гипотезе самостоятельно приобретенного престижа, поскольку эти молодые люди вряд ли могли достичь такого статуса за свою короткую жизнь, а вместо этого должны были унаследовать его. Аргумент в пользу наследования статуса среди семей элит дополнительно подкрепляется повторяющимся сочетанием символов, ассоциирующихся с ритуальной властью, в погребениях сыновей и дочерей правителей, которые включают золотые украшения, драгоценные сосуды для питья и повозки.
Ключевым аспектом династической системы наследственной власти является биологическое родство. Хотя существуют и другие формы родства, в том числе социальные, подразумевающие наличие приёмных детей, что, как известно, трудно выявить только на уровне археологических раскопок погребений, биологическое родство может быть окончательно реконструировано с использованием генетических данных. Таким образом, древняя ДНК является уникальным инструментом для решения этого вопроса, ответить на который до сих пор не удавалось. Но в этом научном исследовании учёные предоставляют полногеномные данные от представителей раннего кельтского общества юго-западной Германии, свидетельствующие о его генетической структуре и политической организации в VI и V веках до нашей эры.
Авторы исследования на 7 участках с элитными погребениями получили полногеномные данные от 31 человека (20 мужчин и 11 женщин), как из высокостатусных захоронений, так и из вторичных, под вторичными в данном случае подразумеваются более поздние по времени захоронения в уже существовавших погребальных комплексах.
В выборку вошли:
· Семнадцать человек из большого кургана Магдалененберг земли Баден-Вюртемберг на юго-западе Германии;
· четыре человека из Хохдорфа;
· три из Графенбюля близ Асперга;
· три человека из Людвигсбурга-Рёмерхюгеля;
· два человека из захоронений городища Хойнебург;
· один из захоронения «княжеского» ранга в районе Шёкинген города Дитцинген;
· И ещё один из ритуального места Альте Бург в общине Лангенэнслинген, примерно в 9 км к северо-западу от кельтского городища Хойнебурга гальштатской культуры и раннего периода латенской.
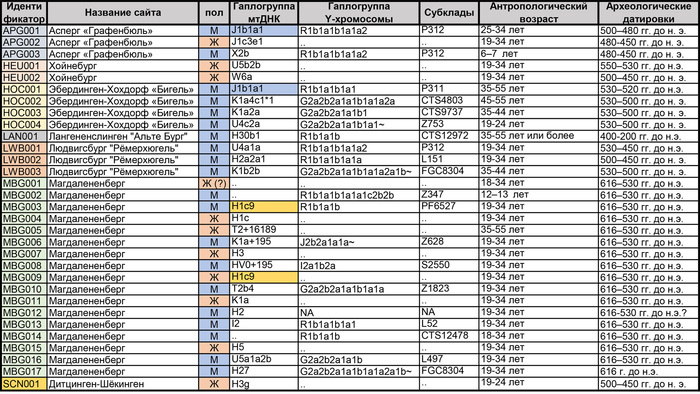
При этом три центральных захоронения Магдалененберга (Магдалененберг017), Хохдорфа (Хохдорф001) и Графенбюля (Графенбюль001) были мужскими, а в центральном захоронении Дитцинген-Шёккингена была похоронена молодая девушка (Шёкинген 001), что подтверждается и остеологическими данными.
В дополнение к полногеномным были получены и изотопные данные, чтобы реконструировать закономерности индивидуальных миграций.
Среди изученных людей авторы выявили несколько тесных родственных связей.
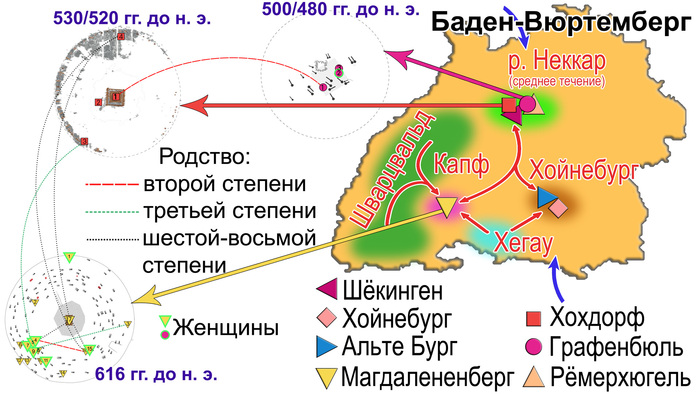
Примечательно, что родство выявлено между мужчинами из самых богатых захоронений доисторической эпохи Европы — центральных могил Хохдорфа (Хохдорф001) и Графенбюля (Графенбюль001), для которых авторы определили родство второй степени. Оба мужчины имеют один и тот же гаплотип мтДНК – J1b1a1 (с двумя частными мутациями), что предполагает родство по материнской линии. С вероятностью в 86% мужчина из Хохдорфа (Хохдорф001) был дядей захороненного в Графенбюле (Графенбюль001).
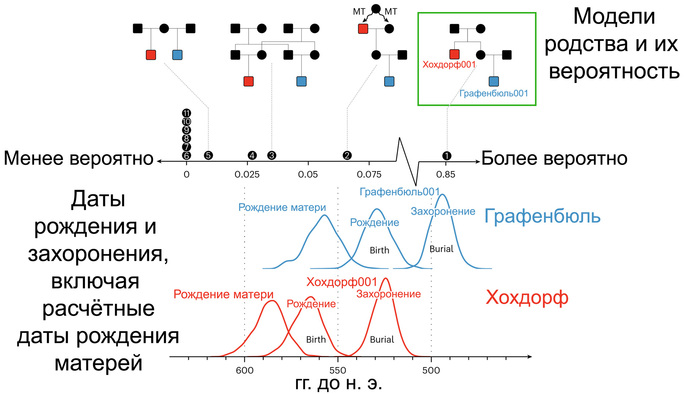
Их изотопные данные также очень похожи и согласуются с жизнью в регионе среднего течения реки Неккар, указывая на местное происхождение обоих мужчин. Эти результаты согласуются с предыдущими предположениями об их связи, основанными на датировках захоронений и археологических данных. Тесная биологическая связь между ними может также объяснить их исключительный рост – у дяди (Хохдорф001) он составлял 180-184см, племянник был чуть ниже. Они были самыми высокими в том числе и по сравнению с зарегистрированными представителями железного века Южной Германии. Хотя в целом мужчины из элитных захоронений были выше мужчин из вторичных.
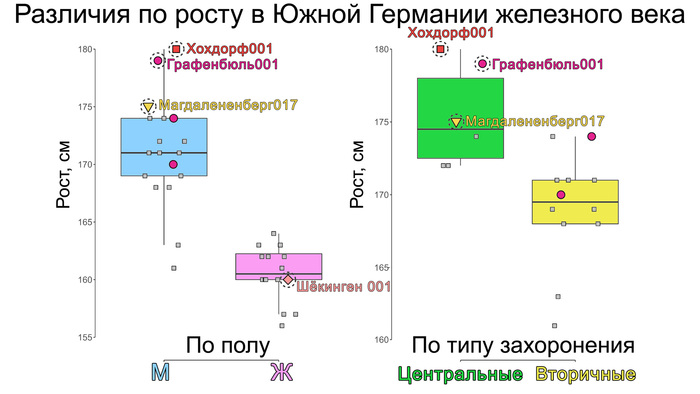
Это подчеркивает вероятность того, что, помимо лучшего питания, генетическое родство также могло способствовать этой социальной дифференциации по росту.
Вторая уникальная находка - это биологическое родство третьей степени (возможно, как у прабабушки с правнуком) между женщиной Магдалененберг009 из богато обставленной могилы Магдалененберга и мужчиной Хохдорф003 из вторичного захоронения из Хохдорфа на удалении в 100 км друг от друга и разницей во времени захоронения в 100 лет. По изотопным данным мужчина вырос в районе Капфа, поселения, связанного с Магдалененбергом, хотя происхождение к северу от Хохдорфа могло бы тоже возможно.
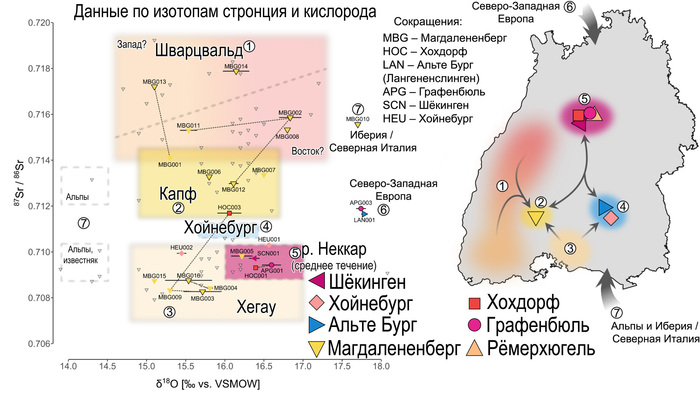
Такая тесная связь между поселениями на большом географическом расстоянии чрезвычайно редка в археогенетических записях из того времени. Внутри этой группы Магдалененберга авторы дополнительно выявили родственников третьей степени между женщиной Магдалененберг009 и молодым взрослым мужчиной Магдалененберг003, оба имели один и тот же гаплотип мтДНК – H1c9, что указывает на то, что близкое родство, вероятно, было по материнской линии.
Третья группа родственников на удалении, состоит из родственников второй степени - взрослой женщины Магдалененберг004 и взрослого мужчины Магдалененберг016, которые были более дальними родственниками мужчины Магдалененберг017 из центрального захоронения Магдалененберга и молодого мужчины из вторичного захоронения «княжеских могил» Хохдорфа – Хохдорф004. У этих людей отмечены значительные фрагменты ДНК идентичные по происхождению, типичные для родственников шестой-восьмой степени, что указывает на то, что у этих людей не так давно был общий предок. Примечательно, что погребения женщины Магдалененберг004 и взрослого мужчины Магдалененберг016 Магдалененберга, которые были более близкими родственниками, являются нетипичными для участка. В то время как могила женщины была довольно богато обставлена, могила мужчины была скудна и является единственной могилой, которая имеет нетипичную ориентацию и перекрывается с другим захоронением. Хотя, как мужчина, так женщина были захоронены в кургане в раннюю фазу и потенциально были связаны с семьей-основателя. Помимо этого, женщина Магдалененберг004 похоронена в непосредственной близости от другой неродственной ей женщины Магдалененберг005, которая по изотопным данным была из района среднего течения реки Неккар, где расположены памятники Хохдорфская гробница, Графенбюль и Дитцинген-Шёкинген.
Авторы отмечают, что обнаруженное биологическое родство между центральным и вторичным погребениями согласуется с интерпретациями Магдалененберга как кургана «родственной группы» для «большой семьи».
Интересно, что последняя упомянутая группа родственников демонстрирует значительно большее южноевропейское происхождение, чем другие проанализированные люди, что отражено в большем вкладе от ранних европейских земледельцев (EEF) (55,6 ± 0,9% против 48,4 ± 1,1%). Это может указывать на неместное, южноевропейское происхождение предков элиты из Магдалененберга.
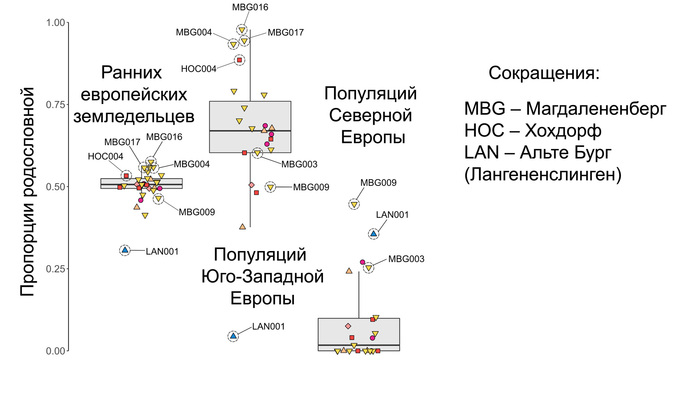
Сравнив геномы Магдалененберг004, Магдалененберг016, Магдалененберг017 и Хохдорф004 с ~ 5660 ранее опубликованными древними геномами, авторы определяют их происхождение от популяций Северной Италии, в то время как другие исследованные представители гальштатской культуры происходят из районов к северу от Альп, недалеко от их мест захоронения. Примечательно, что у людей, связанных по происхождению с Северной Италией, маркером которого является увеличение доли ранних европейских земледельцев, этот вклад был больше по Х-хромосоме по сравнению с аутосомами (83,5 ± 9,9% против 55 ± 1,1%).
А это указывает на то, что примесь ранних европейских земледельцев была получена преимущественно благодаря женщинам южноевропейского происхождения. Чего не выявлено у других проанализированных людей гальштатской культуры.
Увеличивая масштаб каждого участка, авторы реконструировали несколько биологических связей третьей-четвертой степени между вторичными захоронениями Магдалененберг001 и Магдалененберг013, а также тремя захоронениями Магдалененберг002, Магдалененберг011 и Магдалененберг012, которые по изотопным данным были местными из районов Магдалененберга и Шварцвальда.
А вот в Графенбюле и Хохдорфе ни одно из вторичных захоронений не связано с центральными могилами. Также интересно, что в Графенбюле двое захороненных во вторичном двойном захоронении – взрослая женщина Графенбюль002 и мальчик Графенбюль003, помимо отсутствия родства с центральным захоронением даже не были родственны друг другу, что возможно указывает на случай воспитания/усыновления, но точно установить затруднительно.
В целом в выборке из исследования, мужчины и женщины демонстрируют схожие значения стабильных изотопов стронция и кислорода, что контрастирует с миграциями в Южной Германии раннего и среднего бронзового века, где было обнаружено значительно больше неместных женщин, чем мужчин.
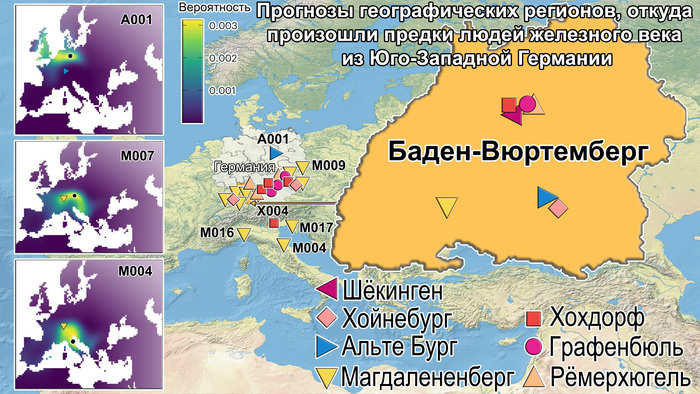

Помимо этого, не было выявлено связи погребального инвентаря с происхождением людей как по ДНК, так и по изотопным показателям.
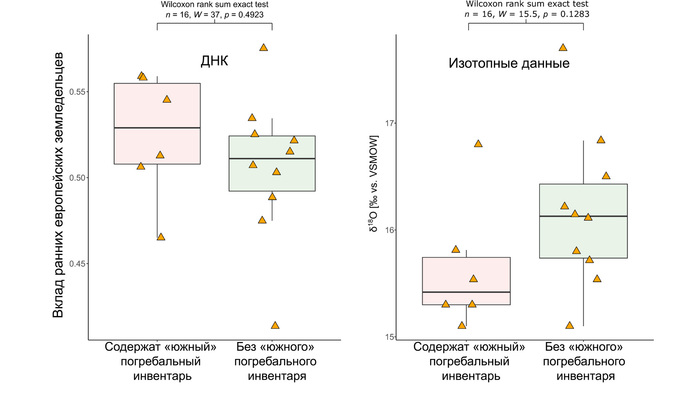
Т.е. наличие неместных артефактов из Северной Италии и Юго-восточной части Альп, которые присутствовали в 6 из 16 погребений Магдалененберга не сочеталось с происхождением захороненного и, следовательно, такой погребальный инвентарь не является надежным маркером южноевропейского происхождения в кельтской популяции Магдалененберга.
В работе авторы также проанализировали людей на наличие протяженных участков гомозиготности (ROH), которые указывают на близкое биологическое родство родителей.
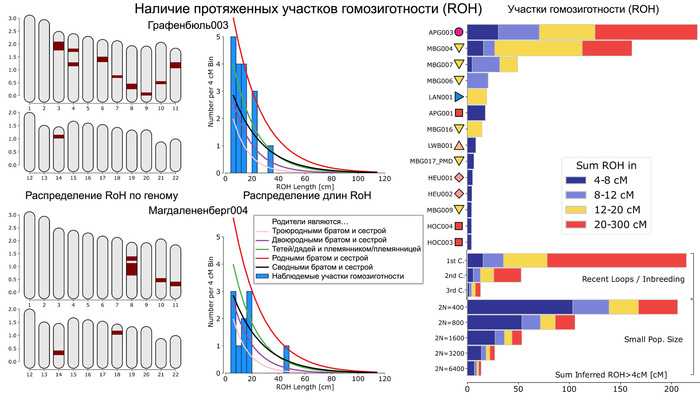
И несмотря на небольшую выборку в 30 человек, такие участки были обнаружены у взрослой женщины Магдалененберга (Магдалененберг004) и у ребёнка из Графенбюля (Графенбюль003). Протяженность участков указывает на то, что их родители могли быть двоюродными братом и сестрой. При этом наличие в такой небольшой выборке двух случаев близкородственной связи, может указывать на то, что кровное родство было более распространенным среди гальштатской элиты на юго-западе Германии, чем в других древних обществах, о которых свидетельствуют археогенетические данные.
В работе авторы сравнили полногеномные данные представителей раннего железного века с современными людьми, преимущественно из европейских популяций. На графике анализа главных компонент видно, что представители железного века генетически отличаются от современных немцев и смещены в сторону современных французов и других жителей Южной Европы.
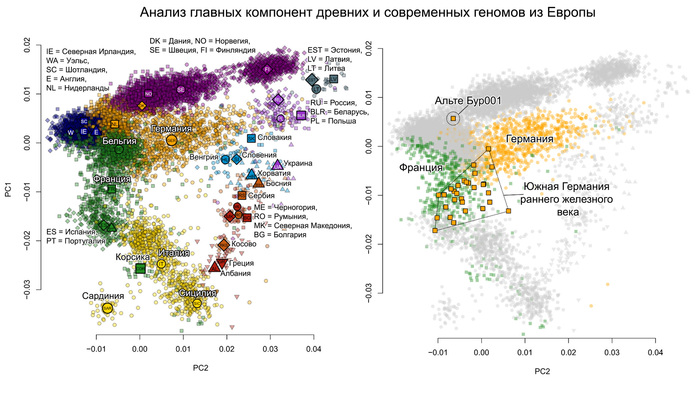
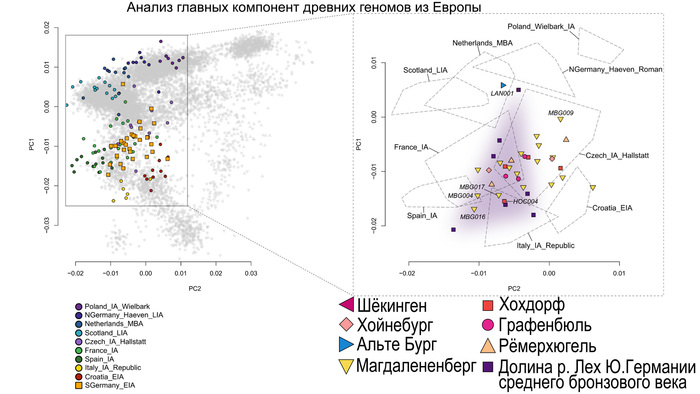
Представители западной гальштатской культуры занимают промежуточное положение между образцами железного века из современной Франции и Чехии и образцами среднего бронзового века из долины реки Лех в Баварии, в пределах разнообразия современных французов.
Здесь ещё раз хочу напомнить про адекватное восприятие метода главных компонент. От союза немца с француженкой кельты не получатся, но их дети будут на графике рядом древними жителями Германии…
Отличия представителей кельтов и современных жителей Германии подтверждаются и другими методами, как на уровне популяции, так и на уровне отдельно взятых людей. При этом родство между представителями гальштатской культуры из Южной Германии и жителями с территории современной Франции бронзового и железного веков является частью более широкого непрерывного генетического сходства от Пиренейского полуострова до Балкан благодаря общей предковой генетической компоненте (показана зелёным цветом на графике).
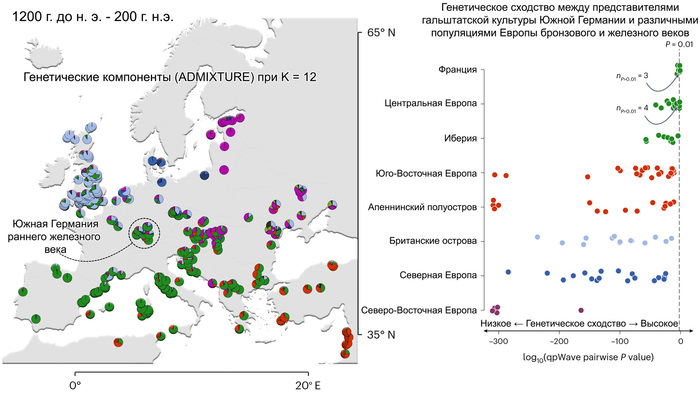
Генофонд людей этой значительной части Европы характеризуется общим демографическим процессом, который отражен в пропорциях предковых компонент. В частности, со времен культуры колоколовидных кубков позднего неолита заметно увеличение предковых компонент ранних европейских земледельцев и сокращение компонент скотоводов древней степи, характерных для представителей ямной и полтавкинской культур.
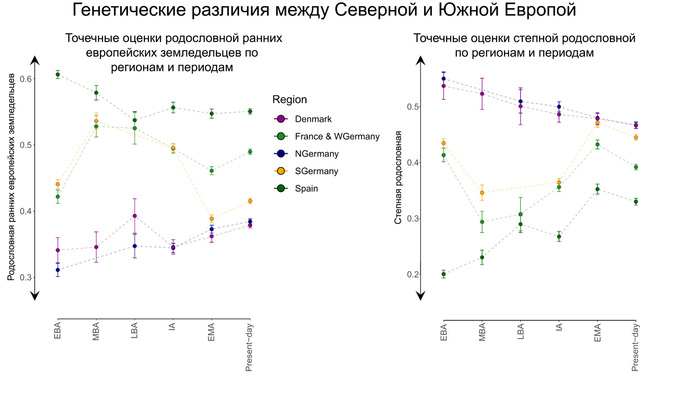
Что, вероятно, произошло благодаря постоянному смешению с сосуществующими группами из других регионов, преимущественно из Южной Европы, у которых было меньше вклада от степных популяций. Что совпадает и с археологическими данными, указывающими на активизацию культурного обмена, особенно в период культуры полей погребальных урн позднего бронзового века, предшественницы гальштатской культуры.
Датировки этой примеси попадают в этот диапазон и указывают на продолжающееся смешение, а не на краткосрочный импульс. Авторы смоделировали генофонд представителей гальштатской культуры как смесь жителей долины Лех Южной Германии раннего бронзового века с представителями популяций Юго-Западной Европы, особенно с Пиренейского полуострова и Италии. Не обошлось без исключений, так женщина Магдалененберг004 и мужчина Магдалененберг016 из вторичных захоронений Магдалененберга продемонстрировали южноевропейское происхождение, а пожилой мужчина Альте Бург001 из Альте Бурга получил большую часть своей родословной из более северных европейских источников, наиболее тесно связанных с населением Нидерландов и Саксонии-Анхальт бронзового и железного веков, что согласуется и с изотопными данными. Миграции людей с большей долей североевропейского происхождения в латенский период также можно наблюдать и в опубликованных данных из соседней Чехии, где было выявлено ранее неописанное разнообразие генофонда в зависимости от предковых компонент североевропейского происхождения из соседних регионов от гальштатского периода к латенскому.
На юге Германии, в Баден-Вюртемберге и Баварии, увеличивающийся приток людей с севера Европы приводит к значительному генетическому обмену между железным веком и ранним средневековьем.
Этот обмен выражен в резком сокращении предковых компонент ранних европейских земледельцев и в существенном увеличении степной родословной, т.е. после притока родословной ранних европейских земледельцев с уменьшением степной родословной, начался противоположный процесс. Поэтому популяции гальштатского периода продемонстрировали наибольшую близость к современным французам, испанцам и бельгийцам, а раннесредневековые жители Южной Германии, как алеманы и баварцы демонстрирует наибольшее сходство с современными датчанами, северными немцами, голландцами и скандинавами и генетически неотличимы от населения железного века и средневековья Северной Германии и Скандинавии.
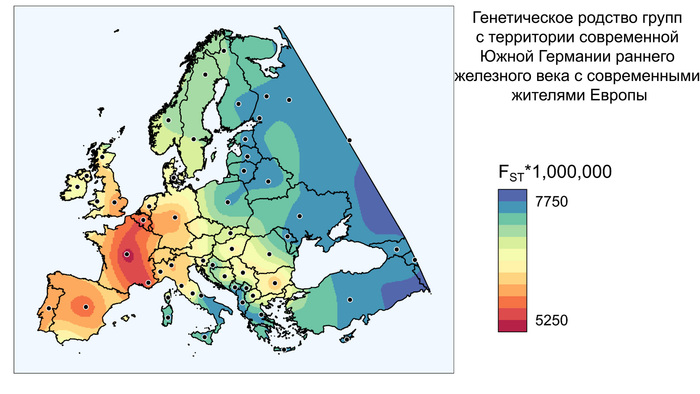
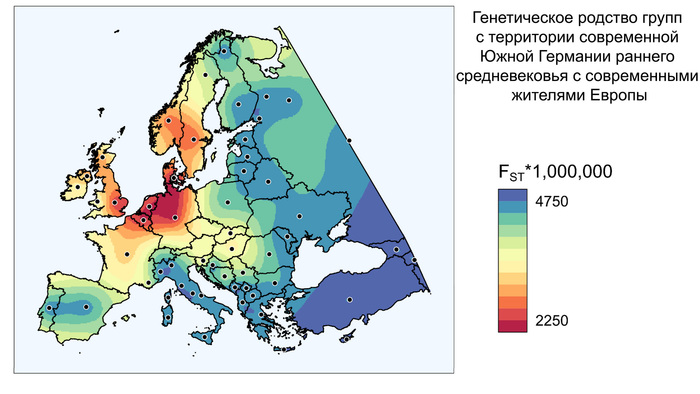
Авторы работы аргументированно утверждают, что это результат значительного притока генов из указанных регионов, как следует из анализа примесей.
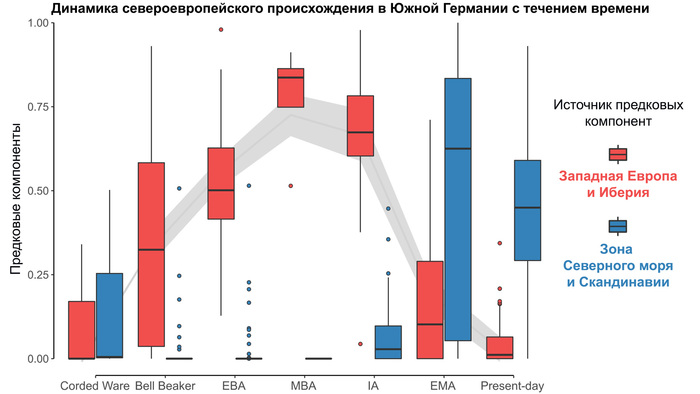
В северных регионах Германии (Саксонии-Анхальт, Нижней Саксонии, Мекленбурге-Передней Померании и Шлезвиг-Гольштейне) процессы формирования генофонда сильно отличались от таковых в Южной Германии.
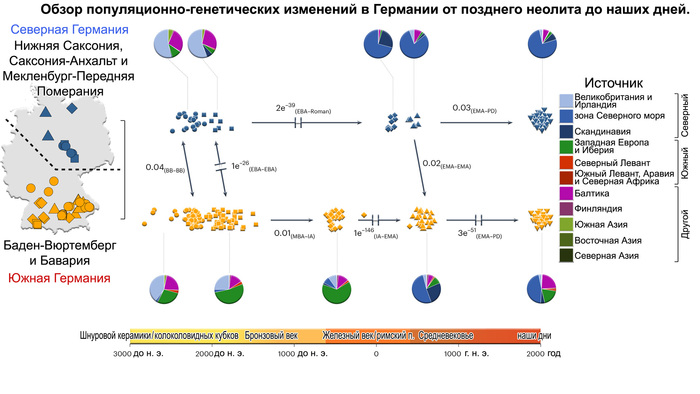
В то время как население бронзового и железного веков на севере также получило дополнительную родословную ранних европейских земледельцев, она была значительно меньше, чем та, что попала в Южную Германию, образуя генофонд, обогащенный предковыми компонентами, связанными со степью, схожий с генофондом современных популяций Дании, Швеции и Норвегии.
А миграция из Северной Германии привела к тому, что в Южную Германию попало меньше предковых линий ранних европейских земледельцев, что привело к увеличению среднего числа североевропейских предковых линий с 2,8% в железном веке до 62,5% в раннем средневековье, а также к появлению новых отцовских линий в виде гаплогрупп Y-хромосомы I1-M253, наиболее характерных для популяций Скандинавии и Северо-Западной Европы.
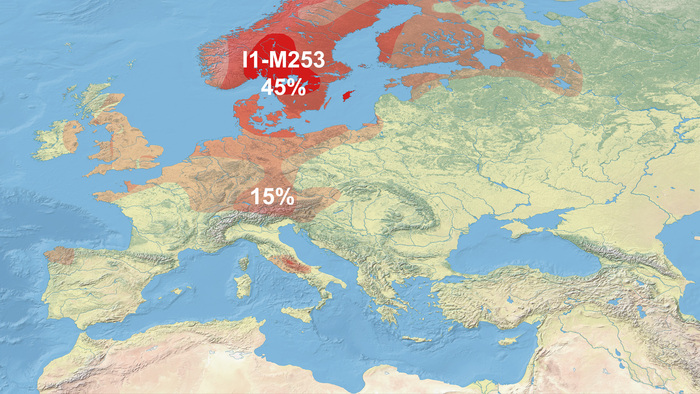
Хотя исследователи не смогли точно датировать эту миграцию, данные о римлянах и популяциях позднего железного века из Баварии и Тюрингии указывают на то, что часть генофонда раннего железного века в Южной Германии не менялась до четвертого или пятого века нашей эры (при этом среднее значение вклада от популяций северной Европы в этих выборках не превышало 8%). А в целом, эти отличающиеся траектории движения людей, вызвавшие изменения в генофонде, по-видимому, являются частью более масштабных миграций повлиявших на пропорции предковых линий североевропейского происхождения у раннесредневековых жителей Англии, Венгрии, Италии и Испании.
Большинство современных немцев попадает в промежуток между представителями гальштатской культуры и раннесредневековыми группами из Южной Германии, что указывает на очередной приток людей с большим вкладом от ранних европейских земледельцев, особенно на юг Германии.
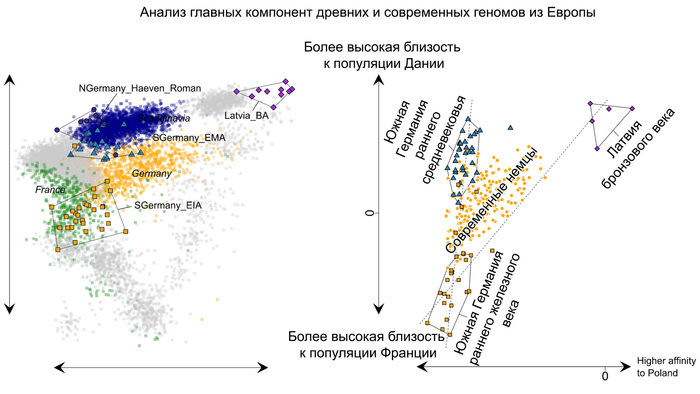
На это также указывают однородительские маркеры в виде гаплогрупп Y-хромосомы. У носителей гальштатской культуры преобладают линии R1b-M269 и G2a-P303, причем на субкладу G2a-L497 приходится 37% гаплотипов в выборке. Примечательно, что мужчины с гаплогруппой G2a-L497 как, Магдалененберг017, Магдалененберг016 и Хохдорф004, по аутосомному геному имеют значительно больший южноевропейский вклад, чем носители линии R1b-M269, как Хохдорф001, Графенбюль001 и Магдалененберг003. Хотя G2a чрезвычайно редка в современной Европе к северу от Альп, G2a-L497 по-прежнему встречается в западных районах распространения гальштатской культуры связанной с протокельтскими племенами, а именно в Восточной Франции, Южной Германии, Швейцарии и Северной Италии, что является дополнительным свидетельством восстановления родословной схожей с таковой у носителей гальштатской культуры железного века в этих регионах.
А большинство современных немцев можно смоделировать как трехстороннюю смесь, в составе которой:
· 54,5 ± 2% от групп южной части современной Германии раннего железного века.
· 33,8 ± 2,5% Популяций Северной Германии римского периода.
· 11,7 ± 1,2% дополнительного вклада, представленного в моделях популяциями Латвии бронзового века. Этот поток генов был после первоначального события смешения и потенциально связан со средневековой миграцией славяноязычных популяций в Восточную Германию.
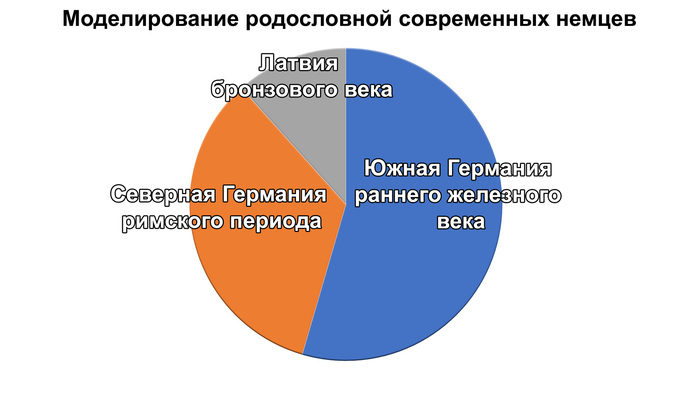
Наследование власти описывается как один из ключевых аспектов ранних исторически сложившихся обществ по всему миру, но факт передачи статуса по наследству трудно доказать только с помощью археологических данных. В описываемой научной работе авторы объединили однородительские и аутосомные данные и смогли доказать тесную биологическую связь (дядя-племянник) между двумя центральными княжескими захоронениями Хохдорфа (Хохдорф001) и Графенбюля (Графенбюль001). Этот наиболее вероятный сценарий родства соответствует более поздним римским историческим свидетельствам, описывающим практики авункулата среди ранних кельтов пятого или четвертого века до н.э. В наше время матрилинейно организованные общества представляют лишь 12–17 % населения мира, а большинство обществ организовано на основе патрилинейной системы родства, что также очевидно из исследований древней ДНК из сообществ неолита и бронзового века Европы.
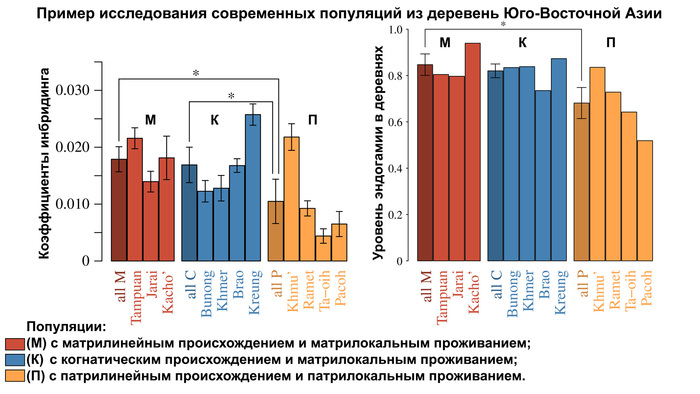
Все же известны примеры доисторических обществ, где наследование власти передавалось по женской линии из поколения в поколение, как в Этрурии и Древнем Риме, к примеру.
Показано, что авункулат возникает в популяциях, в которых распространены внебрачные связи и/или уверенность в отцовстве низка, так что мужчины, скорее всего, генетически более тесно связаны с детьми своих сестер, чем с детьми своих собственных жен, что в конечном итоге способствует инвестированию в детей сестер.

В этом контексте показательны рождённые от союзов между двоюродными братьями и сестрами взрослая женщина из Магдалененберга (Магдалененберг004) и ребёнок из Графенбюля (Графенбюль003). Согласно данным древней ДНК, потомки от союзов между двоюродными братьями и сестрами встречаются довольно редко, с частотой менее 3%.
Тем не менее, авторы подчеркивают, что эта система наследования может быть ограничена южной частью Германии и не распространяться на остальную территорию распространения гальштатской культуры. Кроме того, могут существовать различия между элитой и более широкими слоями населения.
Недавние генетические данные, полученные от захороненных в курганном могильнике раннего железного века Долге Ньиве Словении, не согласуются ни со строго матрилинейной, ни с патрилинейной структурой родства между захороненными и могут указывать на более сложную систему наследования как по мужской, так и по женской линиям, которая потенциально также включала усыновление или воспитание.
Здесь можно отметить захороненных в Графенбюле во вторичном двойном захоронении, а именно взрослую женщину Графенбюль002 и мальчика Графенбюль003, которые не были родственно связаны как с погребённым в центральном захоронении, так и между собой, что совместно с данными изотопов стронция может представлять собой пример воспитания/усыновления. В письменных источниках также описывается воспитание неродных детей среди континентальной и островной кельтской элиты.
Помимо этого, были представлены дополнительные доказательства родственных связей разной степени между людьми из Магдалененберга (Магдалененберг017), Хохдорфа (Хохдорф001) и Графенбюля (Графенбюль001). Такие связи на расстоянии более 100 км и временном интервале до 140 лет свидетельствуют о высокой степени социальной сложности и формировании иерархии регионального масштаба.
В целом, изотопные данные захороненных в Магдалененберге указывают на высокую мобильность по всему континенту в течение их жизни и могут свидетельствовать о наличии структур брачных союзов и опеки, которые связывали отдаленные центры кельтской элиты и формировали широкие и устойчивые социально-экономические сети.
Ранняя кельтская элита, сформировавшая эти сети, возникла в результате длительного популяционно-генетического процесса, выраженного в постоянном смешении с сосуществовавшими группами в Южной Европе, у которых был меньший вклад степных популяций бронзового века. Здесь авторы акцентируют внимание на том, что по результатам генетического анализа взрослого мужчины, из самой ранней в регионе центральной могилы Магдалененберга (Магдалененберг017), датируемого 616 годом до н. э., а также его родственников 5/6 степени (Хохдорф004, Магдалененберг016), их происхождение связано с регионами к югу от Альп. Что может свидетельствовать о ведущей роли этой связи в первоначальном формировании ранних кельтских популяций гальштатской культуры. Связи с регионами к югу от Альп сохранялись на протяжении веков и в материальной культуре, включая погребальный инвентарь. Однако эти сложные политические структуры распались в V и IV веках до н. э.
Люди, генетически отличающиеся от основной массы кельтов из этого и ранее опубликованных исследований, свидетельствуют о том, что впоследствии, в разгар кельтских миграций в IV и III веках до н. э., мигрировали не только «кельты», но и, по крайней мере, ограниченное число людей из северной части Центральной Европы, которые возможно были связаны с такими древнегерманскими племенами как кимвры и тевтоны.
Исторические и археологические данные указывают на то, что формирование культур и популяций в Юго-Западной Германии было изменчивым по времени, особенно в III-I веках до нашей эры. Если от бронзового к железному веку в Южной Германии наблюдается относительная генетическая преемственность на протяжении 2000 лет, то в эпоху поздней античности и раннего средневековья наблюдается резкое увеличение степной родословной. С точки зрения популяционной генетики это согласуется с прибытием германоязычных племен из северной Германии или Дании в период миграции, о чем также свидетельствуют надписи на памятниках VI–VII вв. в Баден-Вюртемберге и Баварии.
Источник: Gretzinger, J., Schmitt, F., Mötsch, A. et al. Evidence for dynastic succession among early Celtic elites in Central Europe. Nat Hum Behav (2024). https://doi.org/10.1038/s41562-024-01888-7
Со времён династии Шан правители Китая обычно пользовались титулом Ван, что-то по типу европейского царь или король, однако в 221 году до нашей эры Ин Чжэн или Цинь Шихуанди «великий император основатель царства Цинь», провозгласил себя “Первым императором”. И в последствии на протяжении более 2000 лет, вплоть до последнего императора государства Цин – Айсиньгёро Пуи, императоры Китая были наделены символическим значением. Император считался “Сыном Неба”, и воспринимался в качестве божественного ставленника, наделенного “Небесным мандатом”, для правление государством, который считался источником легитимации правящей династии. А всего в истории Китая было 83 феодальных династии за 2132 года.
Из них – царство Вэй эпохи Троецарствия (220-266 гг. н.э.), государство эпохи Шести династий – Цзинь (266-420 гг.), а далее с наложением эпохи Шестнадцати варварских государств 304-439 гг., был период Южных и Северных Династий (420-589 гг.) во время которого часто менялась политическая власть, а на севере и юге Китая правили разные императоры. Историки севера отмечают, что в этот период интеграция между кочевыми и земледельческими группами достигла своего апогея, что привело к китаизации некоторых кочевых групп. В течение этих столетий императоры также часто сменялись по мере того, как династии взлетали и падали.
И вот среди этих китайских императоров одной из примечательных фигур был третий император китайско-сяньбийской династии Северной Чжоу - У-ди (Юйвэнь Юн). Это было сяньбийское/тобасское государство периода Северных и Южных Династий в Северном Китае в 557—581 годах. В годы правления императора У-ди Северная Чжоу в 577 году уничтожила своего соперника — империю Северная Ци, присоединив её территорию, что легло в основу дальнейшего объединения Китая во времена императора Вэнь-ди основателя династии Суй (581—618 гг.) и бывшего полководца Северной Чжоу.
Сяньби играли важную роль в истории Китая как минимум на протяжении 7 столетий, поэтому довольно интересно была ли генетическая разница между императором У-ди и другими представителями древнемонгольских кочевников, исследованных ранее. Также немалый интерес как академического сообщества, так и общественности вызывает его генетический профиль, а также физические характеристики, включая внешний вид и предрасположенность к болезням учитывая, что У-ди внезапно скончался в возрасте 36 лет, а его сын умер в возрасте 21 года.
Единственный сохранившийся полный портрет У-ди включен в знаменитый свиток Тринадцати императоров, приписываемый китайскому художнику периода Тан, Янь Либэню (601–673 гг. н.э.). Используя самое современное программное обеспечение для реконструкции лица, авторы исследования попытались нарисовать портрет бывшего императора Северной Чжоу на основе антропологических и генетических данных.

В результате выяснилось, что император был кареглазым брюнетом с кожей от тёмного до промежуточного цвета, что соответствует фенотипам современных жителей Восточной или Северо-Восточной Азии. Что касается причин смерти императора У-ди, которая сильно изменила траекторию последующей истории Китая, то существуют две основные гипотезы: по одной причиной была язва с осложнениями, а по другой его попросту отравили враги. Последняя гипотеза на заре кампании против тюрков, походит на теории заговора. Это когда простые люди могут умирать сколько угодно от болезней и прочих случайных причин, а известные обязательно по злому умыслу. В качестве альтернативы в «Книге Чжоу» сказано, что у императора было расстройство речи, низкое положение верхнего века, проблемы со зрением и хромота на одну ногу. А это уже могли быть симптомы инсульта, учитывая ещё и то, что один из шести локусов риска, связан с инсультом, наряду с повышенным риском подагры и хронического лимфолейкоза.
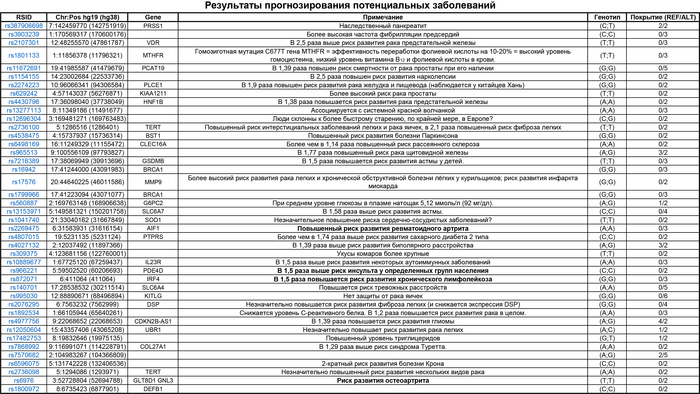
Примечательно, что поскольку вокруг внешности представителей сяньби бытуют споры реконструированная внешность императора У-ди не сильно отличалась от таковой у основного населения Северо-Восточной Азии, что идет вразрез некоторым описаниям сяньби как светловолосых людей с густыми бородами и выпуклыми «высокими» носами.
Помимо этого, предыдущие исследования связали высокий репродуктивный успех по мужской линии с богатством и с социальным статусом. Поэтому считалось, что древние королевские/императорские семьи оказали глубокое влияние на генофонд последующих популяций. Однако гаплогруппа Y-хромосомы C2a1a1b1a-F3830+, F8497 императора У-ди встречается с низкой частотой (<5%) среди населения Восточной и Северо-Восточной Азии. Как и мтДНК C4a1a + 195. Но пока ещё мало данных от таких именитых представителей чтобы подтверждать или опровергать такую теорию.
Генетические данные императора У-ди важны и в контексте происхождения представителей племён сяньби. Так, была обнаружена, наиболее тесная связь императора Северной Чжоу с древними киданями и представителями племени хэйшуй народа мохэ, а также с современными даурами и монголами.
Также авторы исследования смогли сделать вывод о вероятных случаях смешения жителей из Северо-Восточной Азии и долины реки Хуанхэ. В реконструированной генеалогии У-ди его бабушка была из народа когурё в Северо-Восточном Китае, поэтому почти треть его родословной, связанной с популяциями реки Хуанхэ, может быть объяснена последовательными смешанными браками между знатными семьями сяньби и местными китайскими аристократами. Однако у его супруги – императрицы Ашины, как уже было сказано в предыдущем ролике, преобладали компоненты древних северо-восточных азиатов (ANA), без влияния ханьцев связанных с популяциями реки Хуанхэ, что указывает на ограниченное генетическое смешение представителей древнетюркского правящего рода с китайцами. Однако геномные данные самого императора У-ди напрямую свидетельствуют о процессе смешения местной и неместной аристократии.
Помимо этого, в анализе почти у всех представителей сяньби доминировала родословная, связанная с древними северо-восточными азиатами (ANA). При этом группы сяньби продемонстрировали неоднородный генетический состав, тесно связанный с географией. Самые южные представители племен имели до 32% вклада от популяций Хуанхэ, самые западные группы близко к Северному Синьцзяну и горным регионам Алтая имели до ~7% родословной связанной с популяциями западных степных скотоводов, а у сяньби от восточной Монголии до бассейна Амура, вклад древних северо-восточных азиатов был максимален, до 96%.
А генетический профиль Юйвэнь Юна может быть смоделирован как смесь около 62% древних северо-восточных азиатов, 32% китайцев, как представители популяций хуанхэ и 6% западных степных скотоводов. Интересно, что представитель киданей – кочевых монгольских племен – живший примерно через 460 лет после императора У-ди, имел почти тот же генетический состав.
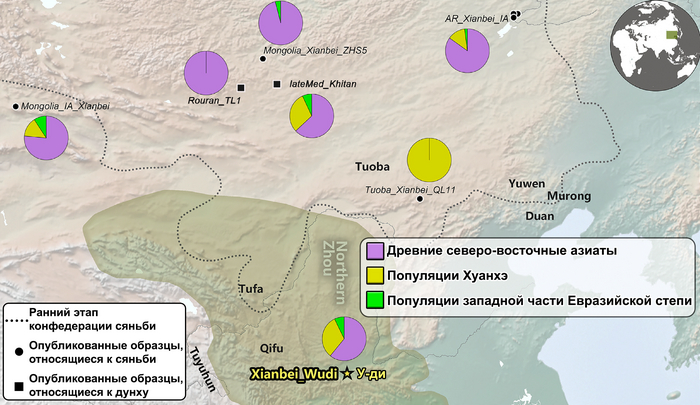
Наблюдаемый генетический ландшафт, вероятно, отражает крупномасштабную миграцию населения из Северо-Восточной Азии и последующее смешение мигрирующих групп древних северо-восточных азиатов с местным населением.
Du P, Zhu K, Qiao H, Zhang J, Meng H, Huang Z, Yu Y, Xie S, Allen E, Xiong J, Zhang B, Chang X, Ren X, Xu Y, Zhou Q, Han S, Jin L, Wei P, Wang CC, Wen S. Ancient genome of the Chinese Emperor Wu of Northern Zhou. Curr Biol. 2024 Apr 8;34(7):1587-1595.e5. doi: https://doi.org/10.1016/j.cub.2024.02.059
В статье речь пойдёт о генетических данных представительницы древних тюрков – принцессы, а после брака с императором китайско-сяньбийской династии Северной Чжоу - У-ди, императрицы – Ашины из одноимённого рода, дочери Мукан-кагана, третьего кагана древних тюрков – Ашина Сыцзинь, при котором Тюркский каганат расширил свои границы от Жёлтого до Чёрного моря, став крупнейшей империей того времени.
Кстати, о происхождении и внешности супруга Ашины - третьего императора китайско-сяньбийской династии Северной Чжоу - У-ди (Юйвэнь Юна) будет в следующей статье.
А в данном исследовании авторы, проанализировав генетический профиль императрицы Ашины, пытаются ответить на три ключевых вопроса:
1. Какое было происхождение у древних тюрков?
2. Есть ли генетическая связь между Ашиной и другими тюркскими народами, а также различными популяциями Центральной и Восточной степи после железного века?
3. Есть ли генетическая связь между древними тюрками и современными тюркоязычными народами?
Первый расцвет тюркских народов пришелся на период с VI по VIII век, когда возникла могущественная кочевая империя в виде Тюркского каганата, основанного правителями из рода Ашина, простиравшегося от Монгольского плато до Каспийского моря. Появившись вслед за хунну, сяньби и другими кочевыми конфедерациями, влияние тюрков ощущалось по всей Евразии, что повлияло на языковой ландшафт значительной её части в течение следующих столетий. Тюркский язык, состоящий из более чем 40 языков, является самой многочисленной группой в гипотетической алтайской языковой семье, и около 200 миллионов человек говорят на тюркских языках более чем в 10 странах.
К сожалению, исторические сведения о Тюркском каганате носят разрозненный характер и часто противоречивы. Происхождение древних тюрков является предметом споров, но если не брать мифические истории с тотемными предками по типу появления от хуннского царевича и волчицы, их происхождение может быть сведено к трем конкурирующим гипотезам:
1. Представители племени Ашина выходцы из одноимённого племени хунну в Северо-Восточной Азии.
2. Их происхождение связано с Причерноморско-Каспийской степью или Центральной Азией с последующей миграцией на восток.
3. Сложносоставное происхождение в окрестностях Пинляна или Гаочана в Турфанской впадине, охватывающее как восточные, так и западные евразийские этнические группы.
Поэтому происхождение представителей правящего рода из племени Ашина вызывает большой интерес и является ключом к разгадке происхождения древних тюрков. Тем более с учётом того, что среди тюркской знати была широко распространена кремация.
По данным мтДНК императрица Ашина принадлежала к гаплогруппе F1d, распространенной в Северо-Восточной Азии. А по результатам полногеномных анализов она группировалась с древними популяциями Северо-Восточной Азии и востока Монгольского плато, но особенно, с древними северо-восточными азиатами, как представители неолита из пещеры Чёртовы Ворота, возрастом около 6700 лет и носители бойсманской культуры среднего неолита из археологического памятника Бойсмана-2, Приморья, представителями неолита Амура и Северной Монголии, а также с кочевниками Восточной степи после железного века, включая сяньбийцев, жужаней, киданей и части монгольского населения. Из современных популяций она была генетически близка с современными тунгусо-маньчжурскими и монголоязычными народами.
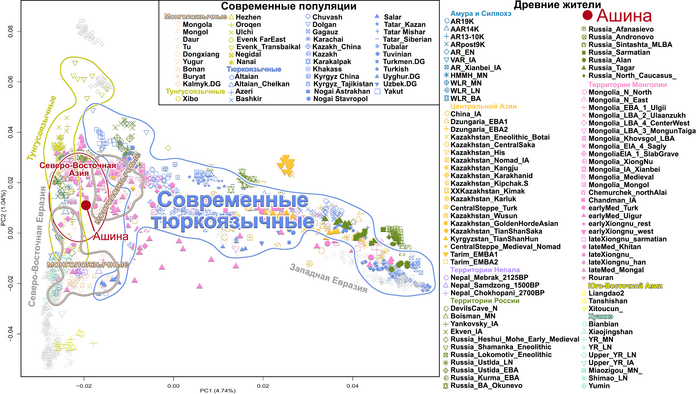
При этом вклада от древних китайцев, связанных с жителями реки Хуанхэ у Ашины не было обнаружено. Однако отмечен небольшой вклад от жителей Западной Евразии – 2,3-3,9%, включающий степную родословную с примесью анатолийских земледельцев, который произошел у предков Ашины за 1566 ± 396 лет до её жизни. В целом генетические данные опровергают гипотезы о западно-евразийском и множественном происхождении представителей знатного рода тюркских каганатов – Ашина в пользу восточноазиатского происхождения.
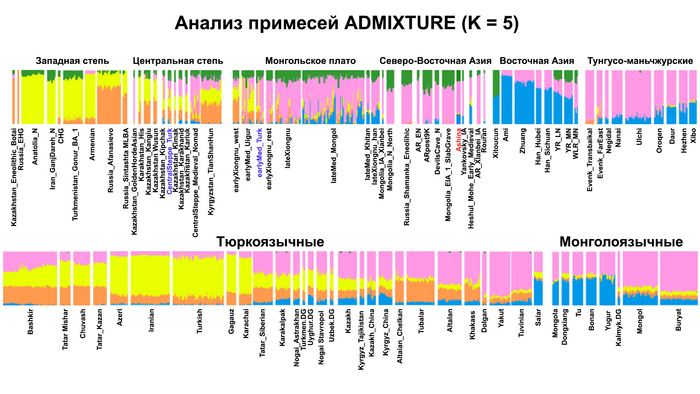
Однако стоит отметить, что у других образцов в тюркский период наблюдалось смешение компонент древних северо-восточных азиатов, которых в среднем было более половины, с компонентами западноевразийских популяций Бактрийско-Маргианского археологического комплекса и кочевников степи железного века, что также соответствует заметному увеличению гаплогрупп Y-хромосомы R и J, в течение тюркского периода в Монголии.
Начиная с железного века в Восточной Степи возникали и рушились, выходящие далеко за её пределы, различные государственные образования кочевников: хунну, сяньби, жужани, тюрки, уйгуры, кидани, кыргызы и – наиболее агрессивные и успешные из всех – монголы. Соседняя средняя часть Евразийской степи или Центральная степь, также была свидетелем перемен в судьбах кочевых образований: гунны, усуни, кангюи, карлуки, кимаки, караханиды и кыпчаки.
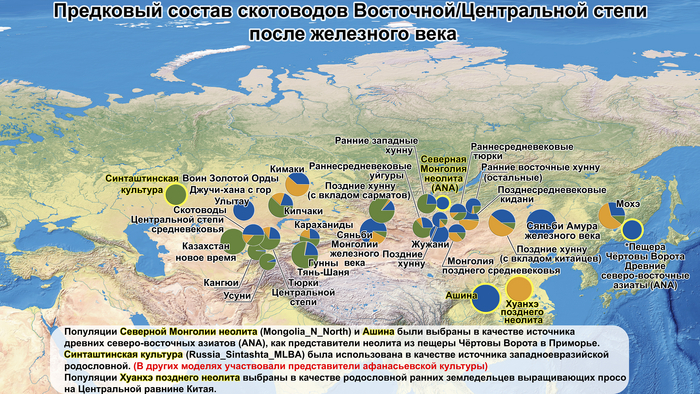
Однако представительница элиты Тюркского каганата продемонстрировала генетическую связь только с древнемонгольскими племенами сяньбийцев, киданей и жужаней, а также с представителями тунгусо-маньчжурских племён мохэ. А вот монголы, уйгуры, карлуки, кимаки, кипчаки, а также караханиды были отдалены от кластера Ашины. Хунну вообще был союз разных по происхождению племён, они сильно отличались между собой по географии и по времени. Некоторые западные их представители вообще на 68,4% имели компоненты как у людей из афанасьевской культуры.
Кстати в моделях надо адекватно воспринимать компоненты, они подразумевают схожие генофонды, а не непосредственных представителей культур.
Среди скотоводов Центральной степи усуни, кангюи и гунны Тянь-Шаня большую часть своего происхождения (62,4–73%) ведут от западных степных групп афанасьевской культуры.
А тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, имеют 35–50,6% компонент как у афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но доля восточно-евразийского происхождения увеличилась у средневековых скотоводов Центральной степи с 67,3% до 82,5%, за некоторым исключением. Однако стоит отметить, что ещё в новое время на территории Казахстана встречались люди с генетическим профилем на 75,5% происходящем от западных степных скотоводов. При этом Ашина не продемонстрировала близкого генетического родства с тюрками Центральной степи или раннего средневековья. Но в отличие от Ашины, тюрки Центральной Степи и раннесредневековые тюрки продемонстрировали высокую, но вариативную степень западно-евразийского происхождения, что указывает на существование генетической подструктуры ещё у древних тюркоязычных народов.
В скотоводческих образованиях Восточной Степи утверждалось, что распространение монгольских языков было связано с сяньбийскими, жужаньскими, киданьскими и монгольскими группами, в то время как хунну, тюрки и уйгуры считались связанными с распространением тюркских языков. В последнем случае были выдвинуты гипотезы о двух волнах распространения: булгаро-тюркской диффузии, начавшейся в гуннский период, спровоцированной более ранней экспансией хунну, и последующей демографической экспансии, связанной с Тюркским каганатом. При этом во втором и третьем веках нашей эры Центральная степь была населена ираноязычными группами, которые постепенно переходили на тюркские языки.
Поэтому генетические данные не подтверждают связи между древними и современными тюркоязычными группами и указывают на значительные различия между тюркоязычными популяциями и популяциями, связанными с распространением тюркских языков.
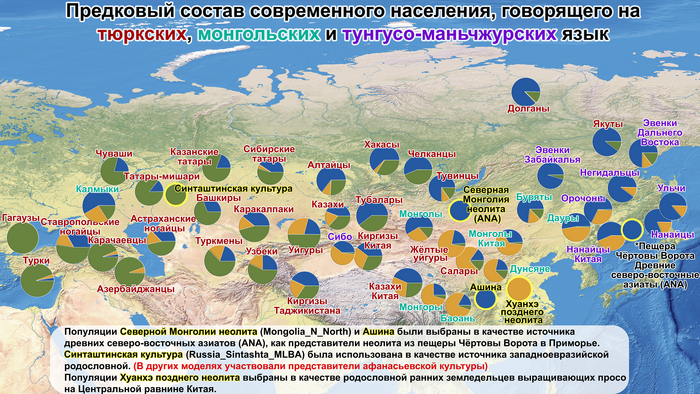
А пропорции предков древних северо-восточных азиатов, которые составляли основу древних тюрков, варьировались у тюркоязычных групп и даже полностью отсутствовали у некоторых самых западных их представителей. Это указывает на то, что распространение тюркских языков было в основном обусловлено культурными факторами, а не демографической диффузией и интеграцией населения. Поэтому натягивать современные тюркоязычные популяции на тюркизированные древние в качестве доказательства тюркоязычности этих древних популяций абсолютно не имеет смысла и противоречит всем данным.
А подвести итог хочу простым языком, отклонившись от строгой научной линии повествования. Ещё раз: это не научная часть!
Древние тюрки, и тем более из рода Ашина, по происхождению были близки к древним северо-восточным азиатам, с незначительным вкладом от западных степных скотоводов. По реконструкциям понятно даже как они выглядели. Это характерные отличия.
Древние и современные тюрки имеют разное происхождение. И многие современные тюркоязычные народы объединяет лишь язык.
Отдельные представители тюркских народов в поисках своих предков почему-то обращаются к древним тюркам, однако схожих с ними компонент у них не так много, а у некоторых вообще почти нет. Их генофонд состоит из смеси различных древних популяций, говоривших на разных языках, при этом у разных народов и разный генетический коктейль и своя интересная история. Но если древних тюрков начинать искать от современных, то можно выйти на разные популяции, которые внесли свой вклад в современные и намеренно или искренне заблуждаясь, объявить их тюрками.
Характерным примером таких манипуляций выглядит якобы доказательство тюркоязычности скифов, основанное только на словах из исследования 2017 года, что современные потомки западно-скифских групп встречаются среди различных современных групп на Кавказе и в Центральной Азии, в то время как сходство с восточно-скифскими группами встречается более широко, но почти исключительно среди современных тюркоязычных групп, особенно из кыпчакской ветви тюркского языка, которые в прошлом были кочевниками. Что не удивительно с учётом огромного генетического разнообразия этих групп, значительно большего чем у всех народов Европы вместе взятых. Трудно промахнуться в стадо слонов из дробовика. К тому же ещё и с учётом того, что сами тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, как уже было сказано выше, имеют 35–50,6% предков от афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но они же по определению тюрки, и мы не называем кыпчаков индоевропейцами или монголами с китайцами! С такой логикой если в Анадыре с вами поздоровался и разговаривает чукча на чисто русском языке, то его предки по мнению таких вот специалистов в кавычках, должны быть исключительно русскими.
Работа о скифах 2017 года: Unterländer, M., Palstra, F., Lazaridis, I. et al. Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat Commun 8, 14615 (2017). https://doi.org/10.1038/ncomms14615
И это исследование 2017 года в комментариях, не цитировал разве что только ленивый защитник тюркского происхождения скифов и не только. И продолжат это делать, вот увидите. При этом само исследование они не читали и вряд ли понимают…
Толкование скифских слов из тюркского несостоятельно хотя бы потому, что фонетика засвидетельствованных скифских слов противоречит фонетическим характеристикам пратюркского (распад которого по глоттохронологическим данным относится к I в. до н. э., т. е. много позже появления на исторической арене скифов – соответственно, сравнивать скифские слова имело бы смысл только с пратюркскими, а не с их рефлексами в отдельных древних и современных тюркских языках). Так, пратюркское слово не могло начинаться на сонанты *m
и *l, тогда как нам известны скифские имена Мадий и Липоксай; невозможными в пратюркском были и скопления согласных в анлауте (начале слова), засвидетельствованные в самоназвании скифов и таких именах, как, например, Спаргапиф. Одним словом, попытки объявить скифов тюрками представляют интерес разве что с точки зрения современной этнопсихологии. Сергей Кулланда http://генофонд.рф/?page_id=3267



Интервью одного из авторов работы о скифах 2017 г., руководителя межинститутского сектора молекулярной палеогенетики Института цитологии и генетики СО РАН, Института археологии и этнографии СО РАН Александра Пилипенко на научно-популярном сайте elementy.ru
К примеру, от союза русского и китаянки положение их детей может быть на том же графике где-то около ногайцев или ближе к казахам.

*Очень грубый пример
И по пропорциям главных компонент они будут на них похожи, но ведь нельзя же будет их ребенка назвать казахом или ногайцем. Часто вижу непонимание этого элементарного биологического аспекта в комментариях. Но когда нужно что-то доказать о себе любимом и, тем более, великом, тут логика ломается.
Или упор часто делают на то, что если кто-то до наших дней сохранил традиции, связанные с кочевым образом жизни, питанием и тому подобно… То только его предки в прошлом могли этим заниматься. Мне вот интересно, чем занимались по такой логике предки тех, кто сейчас компьютерные технологии развивает или в космос ракеты запускает? А так и бомжам можно сказать, что они продолжатели славных традиций охотников и собирателей… Уж пусть не обижаются на меня современные кочевники и скотоводы, а также их потомки. В сохранении традиций нет ничего плохого, если не сочинять себе историю…
Примечание: текста много, тема сложная и состоит из двух постов. Разделы – "Социокультурные изменения" и "Итоги" (Появление и сокращение генетического разрыва), во второй части.
Генетическое разнообразие человеческих популяций Западной Евразии в значительной степени сформировалось в результате трех основных доисторических миграций:
1. Миграции охотников-собирателей, а именно анатомически современных людей, находящихся на ранней стадии социально-экономического развития, которые занимали территорию, примерно, с 45 тыс. лет назад. При этом вклад в последующие популяции внесли выжившие их линии после последнего ледникового максимума около 25-19 тыс. лет назад.
2. Далее, гораздо ещё более существенный вклад, внесли ранние неолитические земледельцы, пришедшие с Ближнего Востока примерно с 11 тыс. лет назад.
3. Степные скотоводы, распространившиеся из Причерноморско-Каспийской степи около 5000 лет назад, привнесли третью основную генетическую компоненту в популяции Западной Евразии.
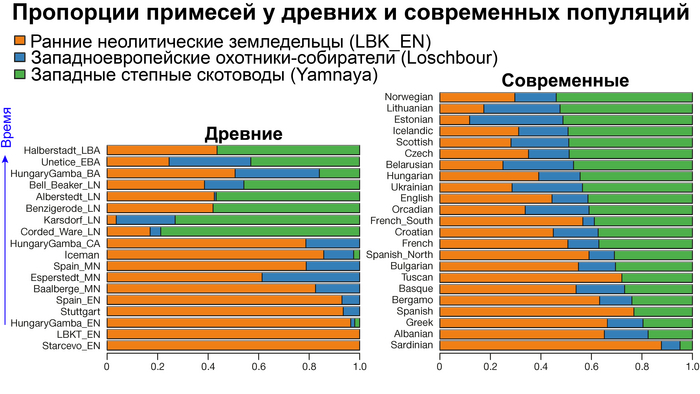
Аутосомный генофонд практически всех европейских популяций по большей части состоит из различных пропорций этих предковых компонент, при этом независимо какие у них гаплогруппы.
Европейский континент заселялся людьми современной анатомии в несколько волн, при этом не все линии этих людей были предками современных. Первые «сапиенсы» Европы (более 40 тыс. лет назад) пересекались и скрещивались с неандертальцами, пока те совсем не вымерли. Об этой примеси свидетельствуют геномы людей из пещеры Бачо Киро в Болгарии, связанных с археологической культурой начального верхнего палеолита, и со стоянки Пештера-ку-Оасе в Румынии. Другие люди того периода, такие как женщина из пещеры Златый кунь Чехии и человек из Усть-Ишима России, не несут в своих геномах значительного количества неандертальского вклада, чем другие неафриканские группы. При этом люди, жившие в Европе более 40 тыс. лет назад, не оставили существенных следов в генетическом составе современных евразийских популяций, что говорит о том, что они в значительной степени вымерли или были ассимилированы последующими группами людей современной анатомии.
В те времена, на демографию древних людей влияли различные факторы, в том числе и климатические. Пик похолодания во время последнего ледникового максимума (LGM) был около 21 тыс. лет назад, когда Северная Европа была в значительной степени покрыта льдом, а южная граница ледниковых щитов проходила через Германию и Польшу. Многие участки Европы стали холодными пустынями с тундростепями и лесостепями на побережье Средиземного моря. А люди и животные переместились в южные широты, о чем свидетельствует, например, солютрейская культура на Пиренейском полуострове и на юге Франции. Другими предложенными рефугиумами или климатическими убежищами в тот период были Апеннинский полуостров, Балканы и юго-восточная часть Европейской равнины.
А в результате послеледниковой колонизации, палеогенетический анализ выявил разделение популяций охотников-собирателей на западных и восточных. Источники родословной западных охотников-собирателей (WHG) связаны с носителями эпиграветтской, азилийской и других эпипалеолитических культур генетического кластера Виллабруна, появление которого обусловлено миграциями в конце ледникового периода и появлением новой генетической компоненты, связанной с Ближним Востоком того времени, начиная с бёллинг-аллерёдского потепления около 14 тыс. лет назад, что археологически коррелирует с культурными переходами.
А родословная восточных охотников-собирателей (EHG) демонстрирует дальнейшую примесь с верхнепалеолитическими популяциями Сибири - древними северными евразийцами (ANE).
Предковый состав западных охотников-собирателей (WHG) был регионально изменчивым в мезолитических популяциях. Если охотники и собиратели из Британии и северо-западной континентальной Европы продемонстрировали однородный состав от палеолита и мезолита, а также вплоть до неолитической экспансии около 6000 лет назад, то их коллеги из Испании, представляли собой смесь людей из двух рефугиумов, когда к генетическому профилю кластера Виллабруна, добавляется примесь от более ранних местных пиренейских охотников-собирателей, связанных с солютрейской и мадленской культурами верхнего палеолита.
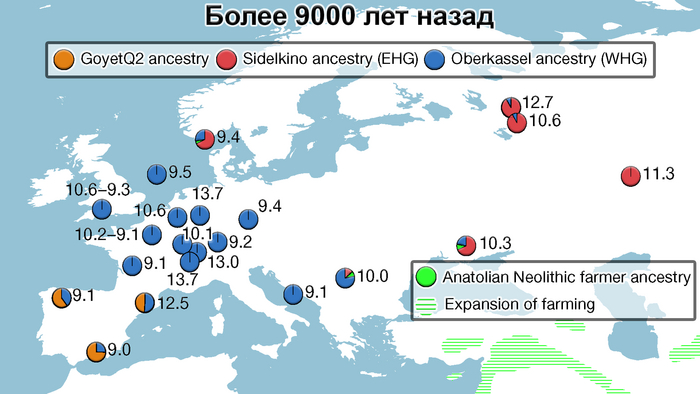
Этот генетический профиль сохранился во время последнего ледникового максимума в Юго-Западной Европе около 25-19 тыс. лет назад.
А время появления примеси сформировавшей восточных охотников-собирателей (EHG), оценивается в 15–13 тыс. лет назад, однако её пропорции в популяциях, по-видимому, соответствуют клине, которая в целом коррелирует с географией, так охотники-собиратели Прибалтики и Украины демонстрируют большее сходство с предками кластера Виллабруна, чем охотники и собиратели западной части современной России, у которых было больше вклада от древних северных евразийцев (ANE).
В свою очередь скандинавские охотники и собиратели продемонстрировали различные пропорции смеси западных и восточных охотников и собирателей (WHG и EHG) в позднем мезолите.
Стоит отметить, что охотники и собиратели мезолита и тем более палеолита, были очень разнообразны и неоднородны, но из-за ограниченного количества данных, наши знания о мезолитической структуре населения и процессах демографического смешения долгое время были скудны и содержали существенные хронологические и географические пробелы в информации. Отчасти это объясняется малым количеством хорошо сохранившихся мезолитических человеческих останков возрастом более 8000 лет, а отчасти и потому, что большинство исследований древней ДНК, посвященных периодам мезолита и неолита, были ограничены преимущественно находками из Европы.


Хотя давно напрашивается более детальная классификация охотников и собирателей по их характерным отличиям. Так, археологические данные указывают на условную границу от восточной Балтики до Черного моря, к востоку от которой сообщества охотников и собирателей существовали гораздо дольше, чем в Западной Европе, несмотря на аналогичное расстояние до центра распространения раннего земледелия на Ближнем Востоке.
Компоненты восточного и западного происхождения у охотников-собирателей в этом пограничном регионе сильно различаются, но более точные представления о демографических процессах, связанных с разделением на восток и запад и о динамике численности населения к востоку от Европы, включая Северную и Центральную Азию за тот же период времени, весьма ограничены. В этих регионах термин «неолит» характеризуется культурными, экономическими и социальными изменениями, а также использованием керамики. К примеру, неолитические культуры степей Центральной Азии и таежного пояса России уже изготавливали керамику, однако продолжали вести прежний образ жизни с присваивающей экономикой, как у предшествующих мезолитических культур охотников-собирателей, с различными инновациями в зависимости от условий среды обитания. Однако пока недостаточно данных для понимания того, как неолитизация отличалась по своим срокам, механизмам и последствиям в разных уголках Евразии.
В Европе, кроме восточной её части, переход от охоты и собирательства к земледелию сопровождался прибытием мигрантов ближневосточного происхождения, практикующим производящее хозяйство на основе одомашненных растений и животных. С последующим значительным ростом населения. В Юго-Восточной, Центральной и Западной Европе анатолийские земледельцы впоследствии в некоторой степени смешались с местными охотниками и собирателями, а на территории Британии, прибывшие с континентальной Европы ранние европейские земледельцы полностью заменили местных охотников и собирателей.
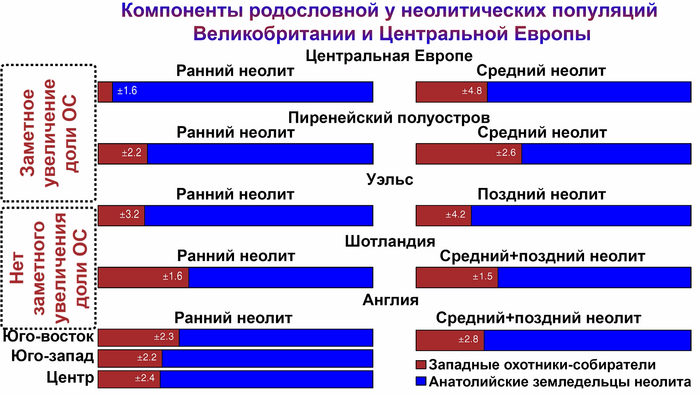
А вот в восточно-балтийском регионе наблюдается заметно иная модель неолитизации, с появлением одомашненных животных только при появлении культуры шнуровой керамики (CWC) около 4800 лет назад. Аналогичным образом в Северном Причерноморье люди с генетическим профилем мезолитических охотников и собирателей тысячелетиями сосуществовали с земледельческими группами, расположенными дальше на запад.
И третий крупнейший генетический пласт в истории европейских популяций связан со степными скотоводами раннего бронзового века, как представители ямной культуры около 5 тыс. лет назад. Этот генетический профиль, быстро распространился по Европе посредством экспансии как носителей культуры шнуровой керамики (CWC), так и родственных ей культур.
Хотя предыдущие исследования выявили эти крупномасштабные миграции в Европу и Центральную Азию, основные аспекты, касающиеся демографических процессов, не решены. Ямная или степная родословная в общих чертах охарактеризована как смесь восточноевропейских (EHG) и кавказских охотников-собирателей (CHG), образовавшаяся в результате гипотетической смеси «северного» степного и «южного» кавказского источников. Однако точное происхождение этих предковых источников не было установлено. Более того, за некоторым исключением, известные гаплогруппы Y-хромосомы представителей ямной культуры не соответствуют гаплогруппам, обнаруженным у европейцев после 5000 тыс. лет назад и происхождение этой отцовской линии также не установлено. К тому же "степная" родословная в Европе до сих пор была выявлена только в смешанной форме, а детали и механизмы, с помощью которых эта родословная впоследствии распространилась вместе культурой шнуровой керамики (CWC), оставались неясными.
В новой работе учёные под руководством Эске Виллерслева из образцов, датированных радиоуглеродным методом при помощи ускорительной масс-спектрометрии, секвенировали геномы 317 человек, 309 из них датируются периодом от 11 до 3 тысяч лет назад.
А совместно с ранее опубликованными данными получилась выборка из 1600 древних геномов, секвенированных с помощью метода дробовика.
Географически 317 отобранных образцов охватывают обширную территорию Евразии, по долготе от Атлантического океана до озера Байкал, а по широте от Скандинавии до Ближнего Востока, и происходят из контекстов, которые включают курганы, пещеры, болота и дно водоёмов.
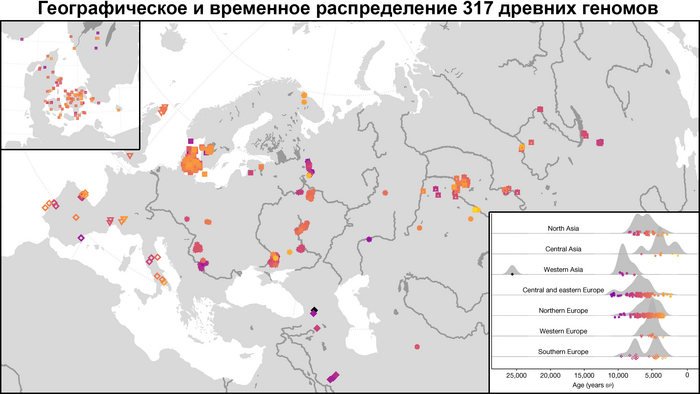
Географически образцы можно разделить на три региона:
1. Центральная, Западная и Северная Европа.
2. Восточная Европа, включая западную часть России, Беларусь и Украину.
3. Урал и Западная Сибирь.
Образцы включают представителей многих ключевых культур Западной Евразии мезолита, неолита и последующих периодов, такие как маглемозе, эртебёлле, воронковидных кубков (TRB) и шнуровой керамики или одиночных погребений в Скандинавии; импрессо или кардиумной керамики в Средиземноморье; кёрёш и линейно-ленточной керамики (LBK) в Юго-Восточной и Центральной Европе; и многие археологические культуры Украины, западной части России и Зауралья (как веретьё, льяловская, волосовская, китойская и др.). Выборка была особенно плотной в Дании, где сотня геномов, охватывала период от раннего мезолита до бронзового века. Обширная выборка была также получена из Украины, Западной России и Зауралья, охватывая период от раннего мезолита до неолита, примерно до 5000 лет назад.
Полученный набор генетических данных позволил при анализе главных компонент использовать за основу древние геномы, и уже на них спроецировать положение современных популяций, а не наоборот, как это было во многих предыдущих исследованиях.
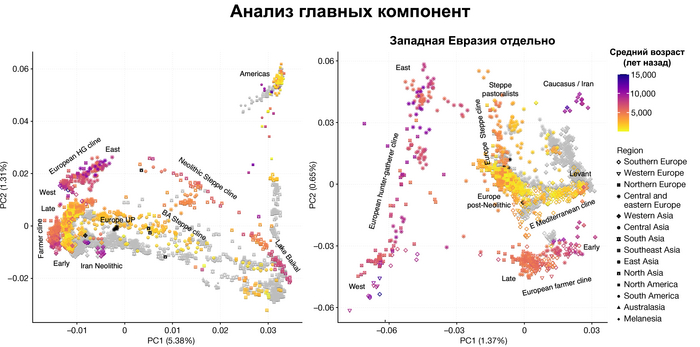
И хотя результаты согласуются с тем, что древние люди сильнее друг от друга генетически отличались, чем современные, такой подход позволил выявить ещё более значительные генетические различия между древними людьми, которые ранее не наблюдались, что отражает меньший эффективный размер их популяции и большую генетическую изоляцию.
Чтобы получить более точную генетическую характеристику в пространстве и времени, авторы использовали попарную идентификацию сегментов ДНК идентичных по происхождению, чтобы сгруппировать анализируемых людей в связанные кластеры сходного генетического происхождения, а не по культурным или территориальным признакам.
В более крупном масштабе они отражали ранее описанные группы западных и восточных охотников-собирателей, а вот в более мелком масштабе они сгруппировались не только в пределах ограниченных пространственно-временных диапазонов и/или археологических контекстов, но также продемонстрировали ранее неизвестные связи в более широких географических диапазонах.
Полученные кластеры далее использовались для моделирования состава предков последующих популяций.
До последнего ледникового максимума, популяции Евразии довольно сильно отличались и в разной степени смешивались с неандертальцами. Некоторые популяции пока остаются гипотетическими, как к примеру, так называемые, «базальные евразийцы» с незначительной или нулевой примесью от неандертальцев, их прямых образцов, подтвержденных генетикой, на данный момент нет, но генетические следы этой популяции были обнаружены у охотников-собирателей Кавказа и Западной Азии верхнего палеолита, а также у более поздних западноевропейских охотников и собирателей.
В данной работе авторы сообщают о генетических данных полученных из зуба женщины верхнего палеолита, жившей около 26-25 тыс. лет назад в окрестностях карстового грота Котиас Клде в современной Грузии, не путать с более молодыми образцами кавказских охотников-собирателей из этого археологического комплекса, у которых было менее 20% этой кавказской родословной верхнего палеолита. А сам геном этой представительницы Кавказа верхнего палеолита представляет собой смесь 76% западноевразийских охотников-собирателей верхнего палеолита (т.е. до максимума последнего оледенения) и 24% «базальных евразийцев». Кстати популяции Ближнего Востока верхнего палеолита, почти наполовину произошли от «базальных евразийцев», которые, практически не имели примеси неандертальцев и отделились от других неафриканских линий ещё до их разделения между собой. При этом западноевропейские охотники-собиратели, жившие после последнего ледникового максимума, моделируются как итоговая, за несколько событий, смесь 46% популяций, связанных с Кавказом верхнего палеолита, 22% европейцев, живших около 33-37 тыс. лет назад, связанных со стоянками Сунгирь и Костёнки и 32% древних популяций, связанных с Сибирью верхнего палеолита, как представитель стоянки Мальта, возрастом 24 тыс. лет, т.е. древних северных евразийцев.
В свою очередь восточноевропейские охотники-собиратели моделируются как смесь 58% популяций, связанных с обитателями стоянки Мальта в Прибайкалье, 28% популяций, связанных с Кавказом верхнего палеолита, как женщина из Котиас Клде (но в разные периоды, сначала 24%, а потом 4%) и 14% представителей Западной Азии верхнего палеолита. Эта смесь также проходила в несколько этапов. Тем самым новые данные подтверждают генетическое разделение охотников и собирателей в Восточной и Западной Европе после максимума последнего оледенения, когда люди снова расселялись по Евразии, при этом эти различия сохранялись на протяжении всего мезолита.
В анализе примесей, эти сложносоставные генофонды представлены отдельными компонентами. А различия между ними, а именно между западноевропейскими и восточноевропейскими охотниками-собирателями мезолита максимально выражены у представителей Италии, возрастом от 15 до 9 тыс. лет и Северо-Запада современной России возрастом от 11 до 8 тыс. лет.
Обратите внимание, что генетический состав одинаков даже у людей с разными гаплогруппами и наоборот, по всему геному люди с одинаковыми линиями гаплогрупп отличаются.
Кстати, в представлении сложных процессов, не стоит забывать о временных промежутках, о которых идёт речь.
Что касается североевропейских охотников и собирателей, то они отличались по пропорциям смеси компонент как у западноевропейских и восточноевропейских охотников-собирателей, за исключением Дании, где популяции продемонстрировали южноевропейское происхождение и были генетически схожи с охотниками-собирателями из Италии, возрастом от 15 до 9 тыс. лет. При чём эта родословная сохранялась в регионе на протяжении 5 тыс. лет.
В целом результаты указывают на то, что существовало по меньшей мере три отчетливые волны миграций охотников и собирателей в Скандинавию:
1. Первая волна связана с миграцией людей южноевропейского происхождения на территорию Дании и прибрежной части Юго-Западной Швеции.
2. Вторая волна в Прибалтику и Юго-восточную Швецию связана с популяциями из Юго-Восточной Европы.
И 3. Третья волна миграций на крайний север Скандинавии, а затем на юг вдоль атлантического побережья Норвегии связана с охотниками и собирателями с северо-запада современной России.
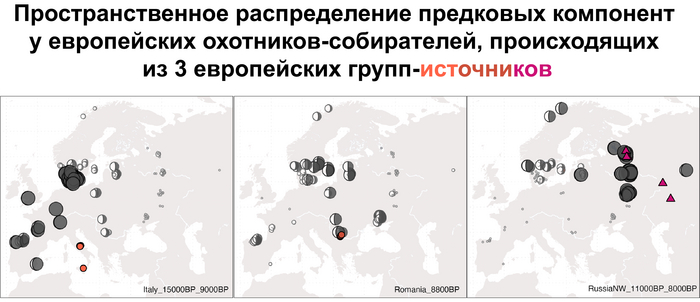
Эти миграции, вероятно, представляют собой послеледниковую экспансию из рефугиумов или районов-убежищ, общих как для людей, так и для многих видов растений и животных.
Стоит отметить, что генофонд европейских охотников-собирателей верхнего палеолита, живших до максимума последнего оледенения был в значительной степени заменён генофондом, происходящим из рефугиума в Южной Италии. На Пиренейском полуострове самые древние образцы из этого исследования, в том числе охотника-собирателя, возрастом около 9500 лет (NEO694) из бухты Санта-Майра на востоке Испании, показали преимущественно южноевропейское происхождение с незначительным вкладом охотников и собирателей верхнего палеолита, который, вероятно, отражает генофонд, связанный с мадленским периодом до максимума последнего оледенения.

А более поздние жители севера Пиренейского полуострова, были ближе к охотникам и собирателям Юго-Восточной Европы, демонстрируя около 40–30% вклада балканских охотников-собирателей.
Поток генов, связанных с охотниками-собирателям Юго-Восточной Европы был направлен и в Восточную Европу после мезолита, на территорию современной Украины и далее. А у двух человек из могильника мариупольского типа раннего энеолита (NEO212, NEO113 ) близ хутора Голубая Криница на юге Воронежской области, возрастом около 7300 лет, генетический профиль состоял уже из смеси более ранних охотников-собирателей с территории Украины с вкладом от 18 до 24% из источника, связанного с охотниками-собирателями Кавказа.
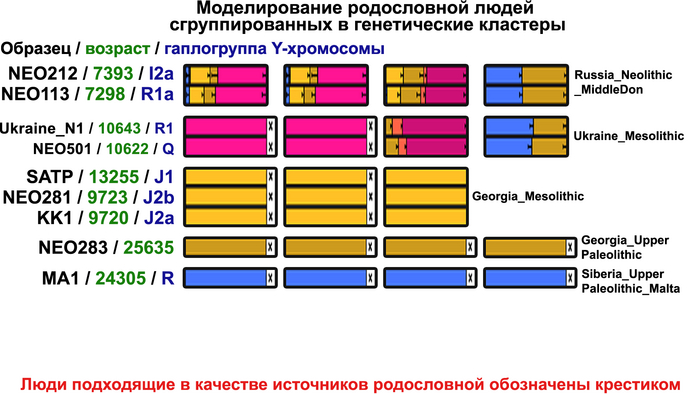
Поэтому на графике анализа главных компонент они отклонены в сторону Ирана и Кавказа. По расчётам эта примесь произошла примерно 8300 лет назад. Результаты указывают на более ранний генетический контакт между популяциями Кавказа и степи ещё до появления культур кочевников в Причерноморско-Каспийской степи. Помимо этого, этот контакт задокументирован западнее, чем предполагалось ранее.
Предыдущие исследования древней геномики задокументировали несколько эпизодов крупномасштабной смены населения в Европе за последние 10 тыс. лет, но 317 новых геномов из данной работы, заполняют важные пробелы в наших знаниях.
Анализ выявил глубокие различия в пространственно-временной динамике неолитизации между восточной и западной частями Европы вдоль пограничной зоны, простирающейся от Черного моря до Балтийского. К западу от этой пограничной зоны, переход к неолиту сопровождался крупномасштабными сдвигами в генетической родословной, от местных охотников-собирателей к земледельцам анатолийского происхождения (Boncuklu_10000BP). Этот переход в зависимости от региона охватывал обширный период времени, более 3 тыс. лет, от самых ранних свидетельств на Балканах около 8700 лет назад до 5900 лет назад в Дании.
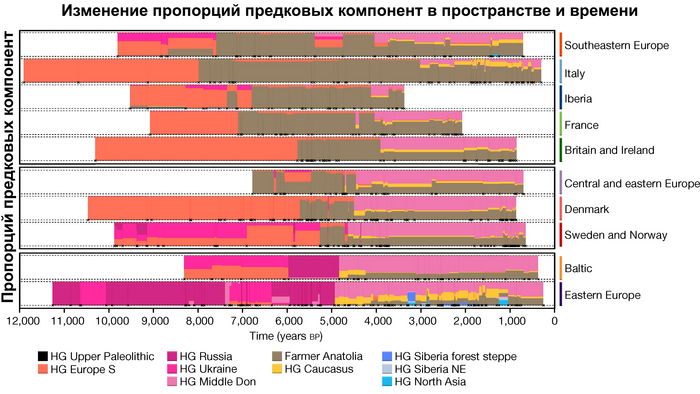
В последующие периоды смешение между ранними европейскими земледельцами и местными охотниками-собирателями привело к частичному увеличению родословной охотников и собирателей во многих регионах Европы, но этот вклад редко превышал 10%, помимо заметных исключений, наблюдаемых у людей из района Железных ворот на границе современных Сербии и Румынии, а также у носителей культур ямочной керамики из Швеции и кардиумной керамики раннего неолита из Португалии, которые, по оценкам, содержат от 27 до 43% вклада от охотников и собирателей, как представители Пиренейского полуострова, возрастом от 9 до 7 тыс. лет (Iberia_9000BP_7000BP).
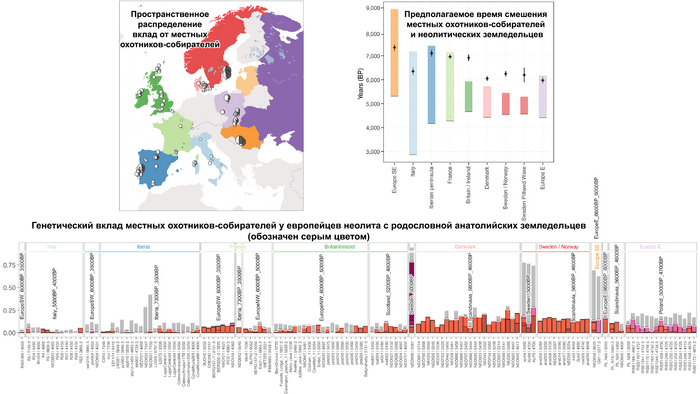
Обитатели эпохи неолита с территории современной Дании продемонстрировали до 25% генофонда охотников-собирателей (EuropeW_13500BP_8000BP), но преимущественно не местных.
Также обнаружены региональные различия в происхождении последующих неолитических групп. Так последующие неолитические популяции Западной Европы происходят от ранних земледельцев Южной Европы. А в последующие неолитические группы Восточной Европы и Скандинавии внесли свой вклад преимущественно ранние земледельцы Центральной Европы.
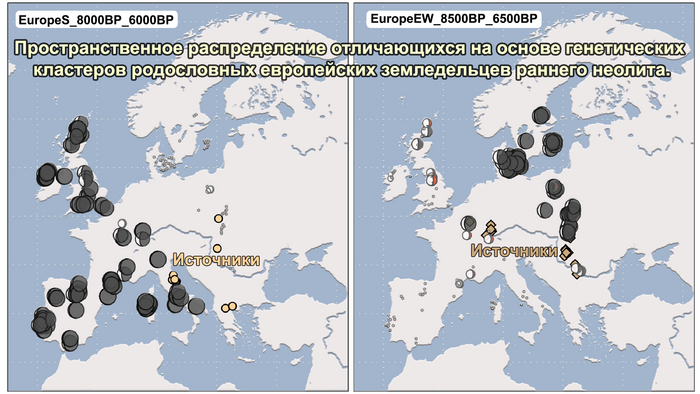
А вот с восточной стороны упомянутой пограничной зоны, на территории Прибалтики, Украины и западной части России, не наблюдается существенных генетических изменений, а предковые линии местных охотников и собирателей, без заметного вклада от анатолийских земледельцев,сохраняются примерно до 5 тыс. лет назад. Эта генетическая преемственность согласуется с археологическими данными, которые свидетельствуют о сохранении в этом обширном регионе групп охотников, рыболовов и собирателей, которые использовали керамику, но не практиковали земледелие и животноводство ещё на несколько тысяч лет.
Но несмотря на то, что влияние степной родословной на популяции Евразии хорошо известно, вопросы о её происхождении оставались не решенными. В данной работе авторы показывают, что состав степной родословной (Steppe_5000BP_4300BP) может быть смоделирован как смесь примерно 65% компонент, связанных с охотниками-собирателями, использующими керамику, из района Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет (MiddleDon_7500BP) и 35% компонент, связанных с охотниками и собирателями Кавказа, возрастом от 13 до 10 тыс. лет (Caucasus_13000BP_10000BP).

Таким образом, охотники-собиратели Среднего Дона, которые уже имели родословную, связанную с кавказскими охотниками-собирателями, являются неизвестным до сих пор ближайшим так называемым северным источником большей части родословной у людей, связанных с ямной культурой. При этом генетический состав людей из могильника Голубая Криница, отличается от такового у других древних людей из могильников мариупольского типа Приднепровья. Однако распространение степной родословной в Европе связно с культурой шнуровой керамики, носители которой представляют собой смесь предковых линий, связанных со степью и неолитическими земледельцами, происхождение которых может быть связано исключительно с культурой шаровидных амфор.
Это наблюдение имеет большое значение для понимания возникновения культуры шнуровой керамики. Хотя уже давно высказывались предположения о стилистической связи между керамикой культур шаровидных амфор и шнуровой керамики, включая использование сосудов в форме амфор и узоры, выполненные шнуром. При этом незадолго до появления самых ранних групп культуры шнуровой керамики восточные группы культуры шаровидных амфор и западные группы ямной культуры обменялись культурными элементами в лесостепной переходной зоне к северо-западу от Черного моря, где керамические амфоры и кремневые топоры культуры шаровидных амфор были включены в погребения ямной, а типичное для ямной использование охры встречалось в погребениях культуры шаровидных амфор, что указывает на тесное взаимодействие между этими группами. Предыдущие генетические анализы нескольких людей из зоны контакта предполагали, что эти контакты были ограничены культурным влиянием, а не примесью. Однако, в свете новых генетических данных, похоже, что эта зона — и, возможно, другие подобные зоны контакта между культурой шаровидных амфор и группами из степи (как из ямной культуры) — были ключевыми в формировании культуры шнуровой керамики, благодаря которой генетические компоненты, связанные со степью, и культурой шаровидных амфор, распространились далеко на запад и север, что привело к регионально разнообразным ситуациям взаимодействия и смешения.
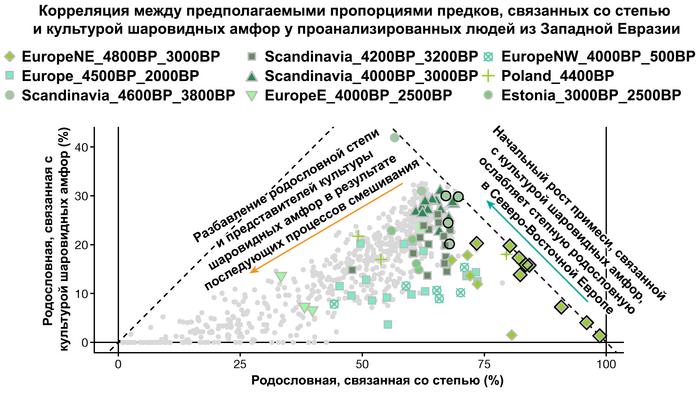
При этом распространение культуры шнуровой керамики происходило преимущественно благодаря связям, которые были выстроены культурой шаровидных амфор в предшествующий период. Но несмотря на сходство по аутосомным данным, различные линии гаплогрупп Y-хромосомы у представителей культур шнуровой керамики и ямной предполагают, что известные на данный момент образцы представителей ямной культуры не подходят в качестве непосредственного источника степной родословной у носителей культуры шнуровой керамики.
Это предположение было подтверждено моделированием предковых компонент у европейцев со степным происхождением возрастом от 3 тыс. лет назад с участием представителей родственных ямной и афанасьевской культур, потому как модели показывают, что представители афанасьевской культуры, хоть и незначительно, но оказались более лучшим источником степной родословной у европейцев, несмотря на разное географическое распространение.
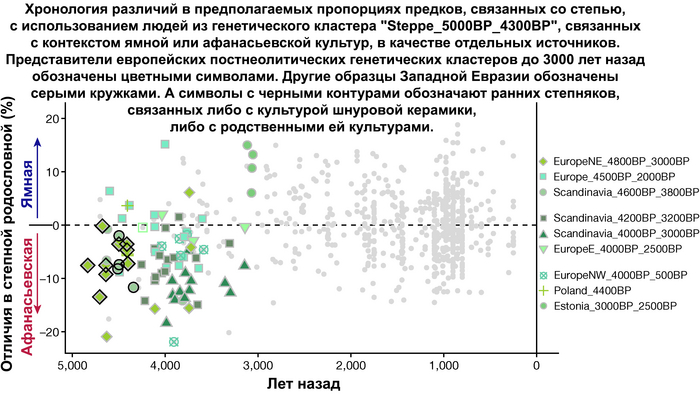
В отличие от значительного числа геномов древних охотников-собирателей из Западной Евразии, которые были изучены до настоящего времени, геномные данные охотников-собирателей к востоку от Урала были скудны. Эти регионы характеризуются ранним появлением керамики из районов, расположенных дальше к востоку, они были населены сложными обществами охотников-собирателей с постоянными, а иногда и укрепленными поселениями. В данной работе сообщается о 38 новых образцах, 28 из которых от охотников-собирателей, использующих керамику 8300-5000 лет назад. Большинство этих геномов попадает на неолитическую степную клину, которая охватывает сибирские лесостепные зоны бассейнов рек Иртыш, Ишим, Обь, Енисей, а также озера Байкал.
Эти охотники и собиратели могут быть смоделированы как смесь компонент из трёх генетических кластеров:
1. Компонента западносибирских охотников-собирателей возрастом от 8300 до 7000 лет (SteppeC_8300BP_7000BP), доминировала в западной части лесостепной зоны.
2. Компонента охотников-собирателей Северо-Восточной Азии возрастом около 7500 лет (Amur_7500BP) или древних северо-восточных азиатов из пещеры Чёртовы ворота в Приморье, была максимально выражена у популяций Байкала.
И 3. Компонента представлена палеосибирской родословной возрастом 9800 лет (Siberian_9800BP) как у обитателя Дуванного Яра в низовьях реки Колымы, она снижались по мере удаления от озера Байкал на запад.


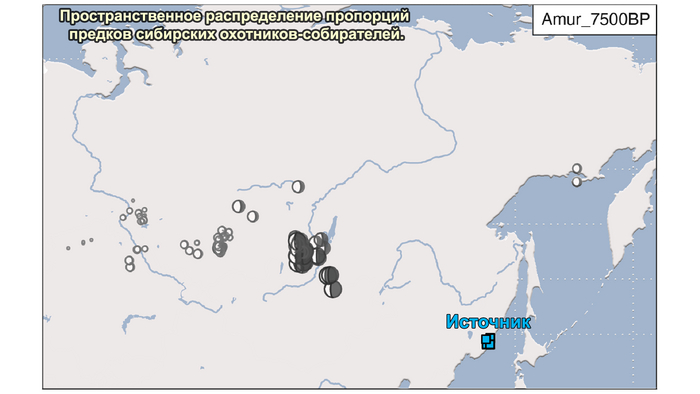
Авторы использовали кластеры также и для исследования пространственно-временной динамики генетического состава в степи и районе озера Байкал после неолита. Результаты подтверждают увеличение компоненты охотников-собирателей лесостепи возрастом от 7000 до 3600 лет (SteppeCE_7000BP_3600BP) у жителей Байкала позднего неолит-ранней бронзы (кластеры Baikal_5600BP_5400BP и Baikal_4800BP_4200BP). Однако родословная, относящаяся к этому кластеру, уже наблюдалась у вновь описанных неолитических охотников-собирателей как у озера Байкал (NEO199 и NEO200), так и вдоль Ангары на севере (NEO843) около 7000 лет назад. У двух мужчин Байкала была определена гаплогруппа Y-хромосомы Q1b1, характерная для последующих групп позднего неолита-ранней бронзы из того же региона. Что совместно с расчётной датой примеси около 7300 лет назад, позволяет предположить, что поток генов между охотниками-собирателями Байкала и лесостепей Южной Сибири уже был во времена раннего неолита, что согласуется и с археологическими данными.
Новые образцы также проливают свет на генетическое происхождение носителей окуневской культуры раннего бронзового века в Минусинской котловине на юге Сибири. Её представителей можно смоделировать как смесь сибирских охотников-собирателей лесостепной зоны из кластеров возрастом 6700-4600 и 7000-3600 лет (Steppe_6700BP_4600BP и SteppeCE_7000BP_3600BP) со степными скотоводами возрастом от 5000 до 4300 лет (Steppe_5000BP_4300BP), связанными исключительно с афанасьевской культурой, время этой примеси датировано около 4600 лет назад.


А примерно 3700 лет назад жители в степном регионе и у озера Байкал демонстрируют уже заметно отличающиеся генетические профили. Ранние стадии этого перехода характеризуются притоком предков, связанных со степью, количество которых снижается со временем, от 70% на начальном этапе.
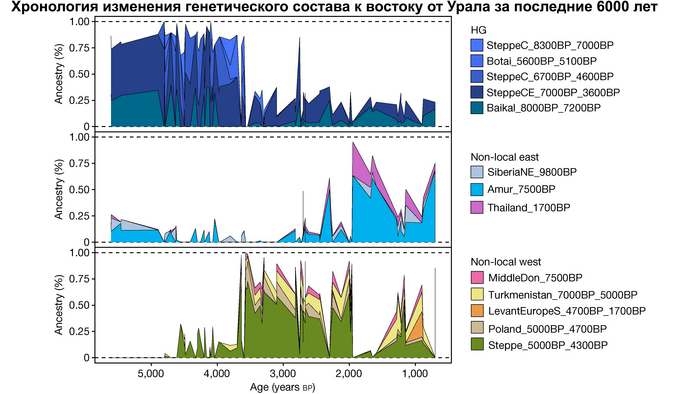
Аналогично динамике в Западной Евразии, происхождение, связанное со степью, здесь соотносится с таковым как у представителей культуры шаровидных амфор с территории современной Польши, возрастом 5000-4700 лет, подтверждая ранее задокументированный поток генов от групп из культуры шаровидных амфор в соседние группы степи и лесостепи, а также поток этой уже смешанной с европейскими земледельцами родословной дальше на восток уже с синташтинской и андроновской культурами в бронзовом веке. Однако у людей из окуневской культуры отсутствует вклад от людей культуры шаровидных амфор, а люди со степным происхождением начиная от 3700 лет назад демонстрируют чуть большую близость к ямной, чем к афанасьевской культуре, что указывает на две волны миграций западных степных скотоводов на восток. Первая волна в раннем бронзовом веке была связана с ямной культурой, а вторая, позже, была связана с синташтинской и андроновской культурами. Более поздние этапы переходного периода характеризуются увеличением родословной, связанной с иранскими земледельцами Центральной Азией (Turkmenistan_7000BP_5000BP) и древними северо-восточными азиатами Приморья (Amur_7500BP).
Суммарно, эти результаты показывают, что на востоке Евразийской степи родословная охотников и собирателей со сложной генетической структурой сохранялась и преобладала значительно дольше, чем в западной Евразии. Пока менее чем 4 тыс. лет назад по степи не прокатились волны миграций, которые привнесли в регион одомашненные породы лошадей с последующим появлением нового снаряжения для верховой езды и колесницы со спицами. Также эти миграции способствовали распространению проса в качестве сельскохозяйственной культуры.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
Это вторая часть с итогами к обеим частям. А первая часть по ссылке
Закономерности распределения сегментов ДНК идентичных по происхождению между людьми, продемонстрировали снижение родства внутри кластеров с течением времени как в западной, так и в восточной частях Евразии. Что согласуется с увеличением эффективного размера популяции в течение этого времени. Однако наблюдаются заметные демографические различия между западной и восточной Евразией. Если на западе во время перехода к неолиту около 9000-6000 лет назад, люди с родословной анатолийских земледельцев демонстрируют снижение доли сегментов ДНК идентичных по происхождению, по сравнению с охотниками-собирателями, то на востоке генетическое родство оставалось высоким примерно до 4000 лет назад, что согласуется с гораздо более длительным существованием небольших локализованных групп охотников-собирателей.
Помимо этого, анализ сегментов гомозиготности (ROH) не выявил значительного числа близкородственных связей, только 29 из 1396. При этом каких-то пространственно-временных или культурных закономерностей в распространенности близкородственных связей обнаружено не было. Зато авторы задокументировали первый случай однородительской дисомии по древнему геному (когда человек получает две копии хромосомы или части хромосомы от одного родителя и ни одной копии от другого родителя). Эта особенность обнаружена у представителя из Темясовских курганов позднесарматской культуры возрастом около 1700 лет.
Также были выявлены люди с различной степенью родства.
После максимума последнего оледенения около 25-19 тыс. лет назад во время расселения древних людей по Евразии, установилось чёткое генетическое разделение между востоком и западом по линии от Черного моря до Балтийского, что отражено и в археологических данных. Это разделение сохранялось на протяжении всего мезолита и неолита. В Восточной Европе распространение земледелия происходило с опозданием примерно на 3000 лет, и эта задержка может быть связана с особенностями окружающей среды, поскольку регионы к востоку от условной границы имеют более континентальный климат и более суровые зимы, возможно, менее подходящие для ближневосточных методов ведения земледелия. На этих территориях сохранились высокоразвитые сообщества охотников-собирателей со стабильными, сложными, а иногда и укрепленными поселениями, с обменом на большие расстояния и крупными кладбищами. Рацион, включающий пресноводную рыбу, зафиксирован как по изотопным данным из этого исследования, так и по анализам остатков липидов в керамике.
В северных лесных районах этой пограничной зоны сообщества охотников-собирателей сохранялись вплоть до появления культуры шнуровой керамики около 5 тыс. лет назад, тогда как в южных и восточных степных регионах охота и собирательство со временем были дополнены животноводством с крупным рогатым скотом и овцами и, возможно, коневодством в Центральной Азии, но этот факт ещё под вопросом. Некоторые из этих групп, такие как представители хвалынской культуры Северного Кавказа и Среднего Поволжья, стали свидетелями появления мужских сообществ, вовлеченных в широкомасштабные обменные связи на основе медных изделий с Восточной и Центральной Европой, а также Кавказом. Поселения в то время были приурочены в основном к равнинным поймам и долинам рек, тогда как степной пояс оставался в значительной степени неиспользованным.
Окончательное исчезновение этой генетической, экономической и социальной границы было вызвано событиями, развернувшимися в степном регионе. Здесь археологически можно наблюдать две временные фазы технологических инноваций: широкое распространение колесных повозок, запряженных волами, около 5500 лет назад и более позднее развитие колесниц со спицами и верховой езды.
Начиная ещё с первой фазы инноваций, в сочетании с возможными изменениями условий окружающей среды, степь стала выгодной экономической зоной, которая позволила группам по типу ямной культуры эксплуатировать степь в качестве кочевых степных скотоводов около 5000 лет назад. А поселения эпохи энеолита вдоль речных долин были заменены этой новой формой мобильной экономики на основе скотоводства, которая окончательно разрушила генетическую границу, существовавшую в предыдущие тысячелетия.
Далее, после второй фазы инноваций около 4 тыс. лет назад, изобретение боевых колесниц и использование проса в качестве продовольственной культуры позволили андроновцам и родственным им группам окончательно распространиться на восток в Среднюю Азию и за ее пределы, оставив глобальное наследие в виде распространения индоиранских языков.
Это исследование внесло свою лепту в наши знания о прошлом, особенно касательно степных миграций.
Во-первых, авторы выявили до сих пор неизвестный источник родословной, который способствовал появлению степных скотоводов, на его роль хорошо подходили охотники-собиратели Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет.
А во-вторых, было определено, что степная родословная, которая распространилась по Европе, благодаря представителям культуры шнуровой керамики, сформировалась в результате культурного и генетического слияния степных групп, родственных ямной культуре и культуре шаровидных амфор. При этом благодаря своему смешанному культурному и генетическому происхождению, носители культуры шнуровой керамики практиковали смешанную экономику, используя различные стратегии выживания в различных условиях. Такая гибкость в значительной степени способствовала их успеху в расселении и адаптации к очень разным экологическим и климатическим условиям за очень короткий период времени.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0