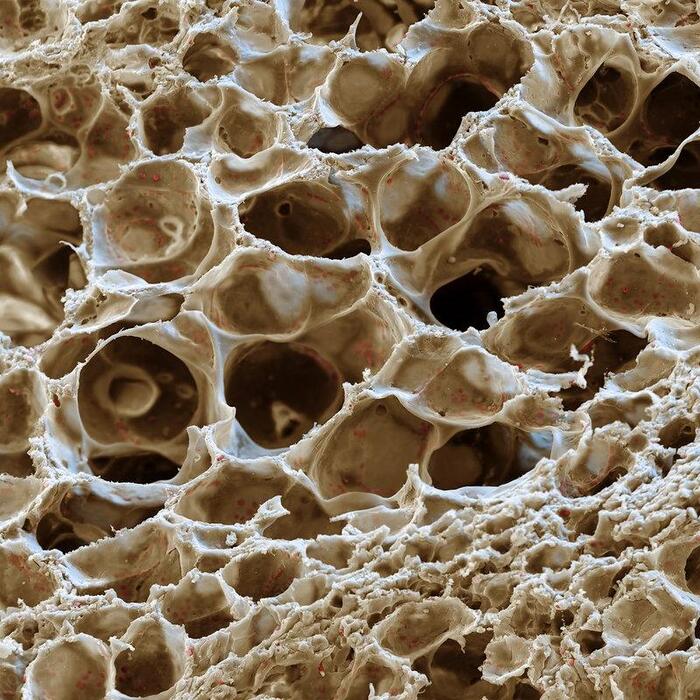
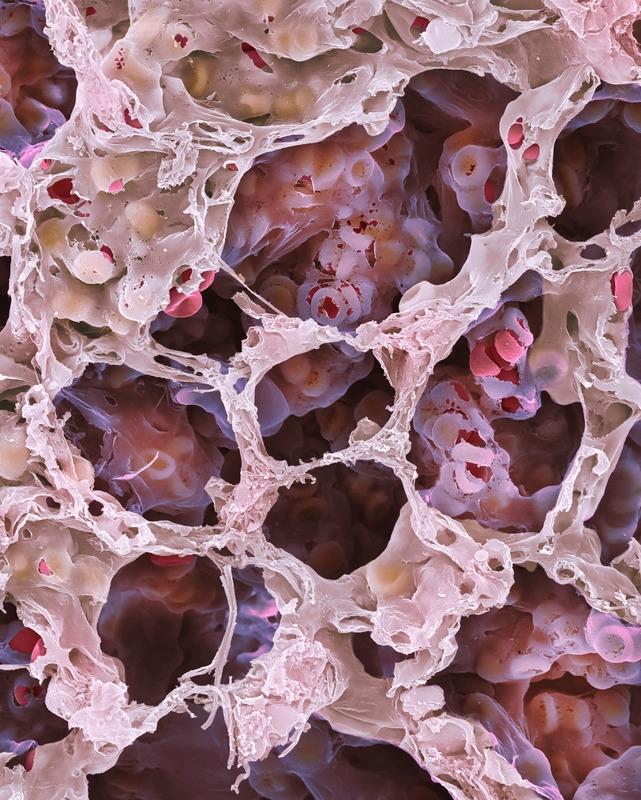
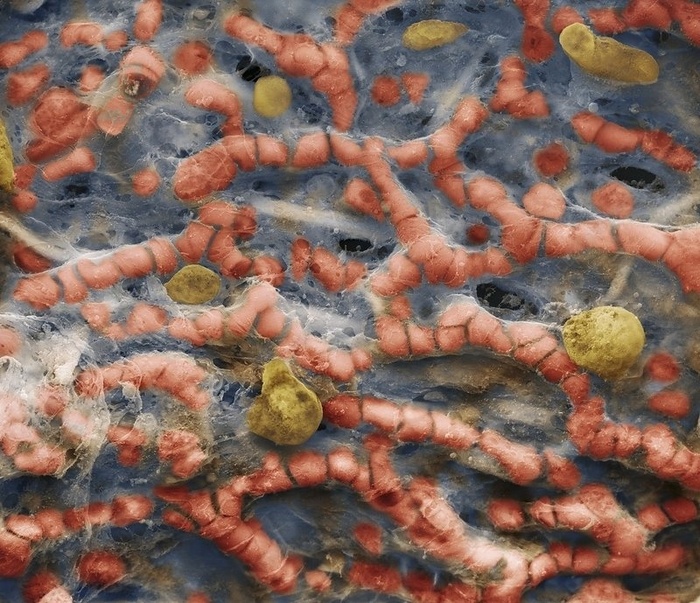
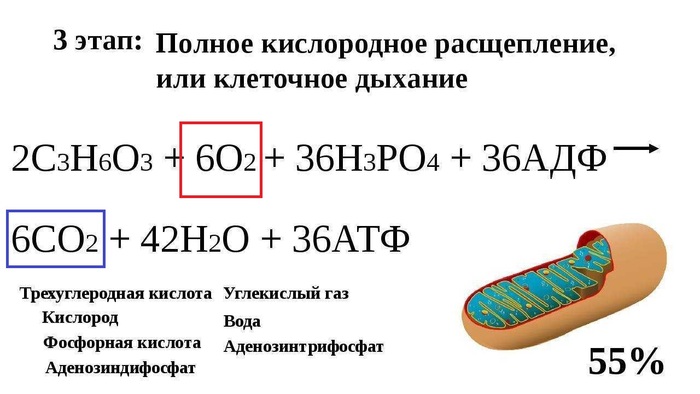
На фотографии представлен снимок лёгких человека. Они состоят из множества ячеистых структур - альвеол.
1/2
Альвеолы лёгких под микроскопом.
В каждой альвеоле происходит газообмен между вдыхаемым воздухом и клетками крови. На фотографиях хорошо видны красные кровяные тельца — эритроциты.
Капилляры в альвеолах. Хорошо видно эритроциты.
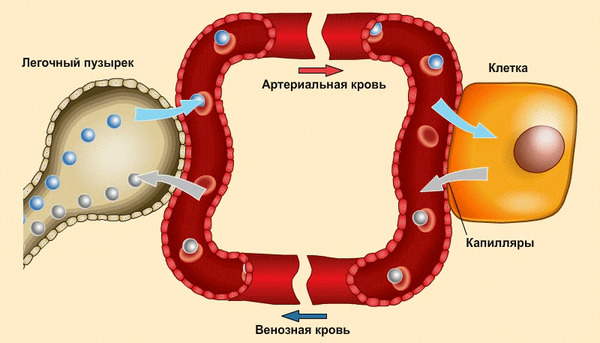
Процесс газообмена в клетках и лёгких.
Когда мы вдыхаем воздух, кислород проходит через стенки альвеол и связывается с гемоглобином, находящимся в эритроцитах. Затем эритроциты в процессе кровообращения доставляют кислород ко всем тканям и органам нашего тела.
Одновременно с этим происходит и обратный процесс: углекислый газ возвращается из клеток в лёгкие и удаляется из организма при выдохе.
Кислород, который поступает в клетку, участвует в реакции получения энергии. В ходе этой реакции выделяется углекислый газ, который является побочным продуктом.
Бонус: Разбираем строение дыхательной системы на примере 3D-модели.
Спасибо, друзья, больше материалов про биологию и микромир Вы можете найти в моём профиле. Подписывайтесь на канал и до скорых встреч.
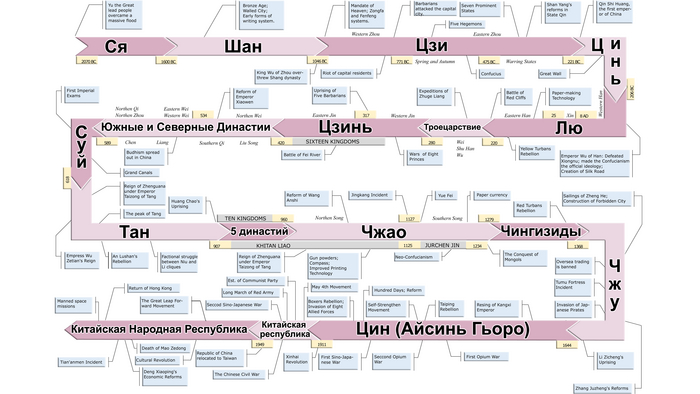
Со времён династии Шан правители Китая обычно пользовались титулом Ван, что-то по типу европейского царь или король, однако в 221 году до нашей эры Ин Чжэн или Цинь Шихуанди «великий император основатель царства Цинь», провозгласил себя “Первым императором”. И в последствии на протяжении более 2000 лет, вплоть до последнего императора государства Цин – Айсиньгёро Пуи, императоры Китая были наделены символическим значением. Император считался “Сыном Неба”, и воспринимался в качестве божественного ставленника, наделенного “Небесным мандатом”, для правление государством, который считался источником легитимации правящей династии. А всего в истории Китая было 83 феодальных династии за 2132 года.
Из них – царство Вэй эпохи Троецарствия (220-266 гг. н.э.), государство эпохи Шести династий – Цзинь (266-420 гг.), а далее с наложением эпохи Шестнадцати варварских государств 304-439 гг., был период Южных и Северных Династий (420-589 гг.) во время которого часто менялась политическая власть, а на севере и юге Китая правили разные императоры. Историки севера отмечают, что в этот период интеграция между кочевыми и земледельческими группами достигла своего апогея, что привело к китаизации некоторых кочевых групп. В течение этих столетий императоры также часто сменялись по мере того, как династии взлетали и падали.
И вот среди этих китайских императоров одной из примечательных фигур был третий император китайско-сяньбийской династии Северной Чжоу - У-ди (Юйвэнь Юн). Это было сяньбийское/тобасское государство периода Северных и Южных Династий в Северном Китае в 557—581 годах. В годы правления императора У-ди Северная Чжоу в 577 году уничтожила своего соперника — империю Северная Ци, присоединив её территорию, что легло в основу дальнейшего объединения Китая во времена императора Вэнь-ди основателя династии Суй (581—618 гг.) и бывшего полководца Северной Чжоу.
Сяньби играли важную роль в истории Китая как минимум на протяжении 7 столетий, поэтому довольно интересно была ли генетическая разница между императором У-ди и другими представителями древнемонгольских кочевников, исследованных ранее. Также немалый интерес как академического сообщества, так и общественности вызывает его генетический профиль, а также физические характеристики, включая внешний вид и предрасположенность к болезням учитывая, что У-ди внезапно скончался в возрасте 36 лет, а его сын умер в возрасте 21 года.
Результаты
Реконструкция внешности
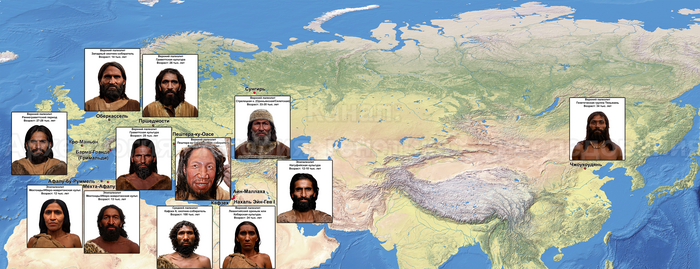
Единственный сохранившийся полный портрет У-ди включен в знаменитый свиток Тринадцати императоров, приписываемый китайскому художнику периода Тан, Янь Либэню (601–673 гг. н.э.). Используя самое современное программное обеспечение для реконструкции лица, авторы исследования попытались нарисовать портрет бывшего императора Северной Чжоу на основе антропологических и генетических данных.
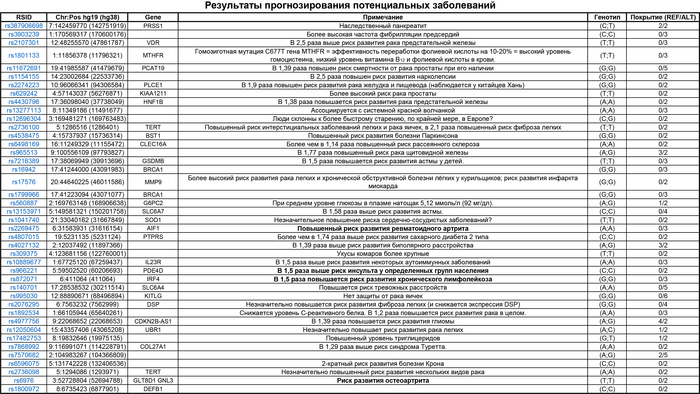
В результате выяснилось, что император был кареглазым брюнетом с кожей от тёмного до промежуточного цвета, что соответствует фенотипам современных жителей Восточной или Северо-Восточной Азии. Что касается причин смерти императора У-ди, которая сильно изменила траекторию последующей истории Китая, то существуют две основные гипотезы: по одной причиной была язва с осложнениями, а по другой его попросту отравили враги. Последняя гипотеза на заре кампании против тюрков, походит на теории заговора. Это когда простые люди могут умирать сколько угодно от болезней и прочих случайных причин, а известные обязательно по злому умыслу. В качестве альтернативы в «Книге Чжоу» сказано, что у императора было расстройство речи, низкое положение верхнего века, проблемы со зрением и хромота на одну ногу. А это уже могли быть симптомы инсульта, учитывая ещё и то, что один из шести локусов риска, связан с инсультом, наряду с повышенным риском подагры и хронического лимфолейкоза.
Примечательно, что поскольку вокруг внешности представителей сяньби бытуют споры реконструированная внешность императора У-ди не сильно отличалась от таковой у основного населения Северо-Восточной Азии, что идет вразрез некоторым описаниям сяньби как светловолосых людей с густыми бородами и выпуклыми «высокими» носами.
Помимо этого, предыдущие исследования связали высокий репродуктивный успех по мужской линии с богатством и с социальным статусом. Поэтому считалось, что древние королевские/императорские семьи оказали глубокое влияние на генофонд последующих популяций. Однако гаплогруппа Y-хромосомы C2a1a1b1a-F3830+, F8497 императора У-ди встречается с низкой частотой (<5%) среди населения Восточной и Северо-Восточной Азии. Как и мтДНК C4a1a + 195. Но пока ещё мало данных от таких именитых представителей чтобы подтверждать или опровергать такую теорию.
Генетическое происхождение
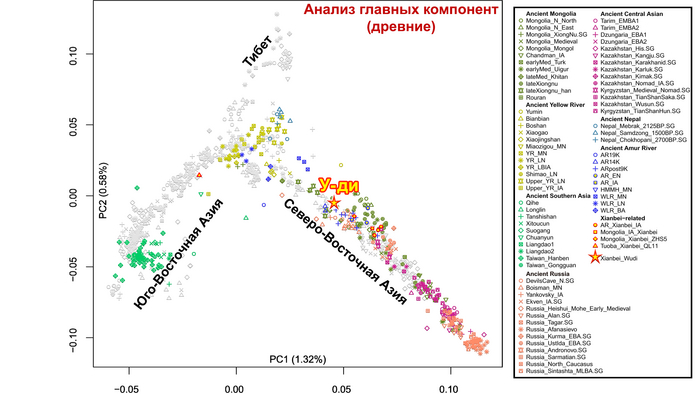
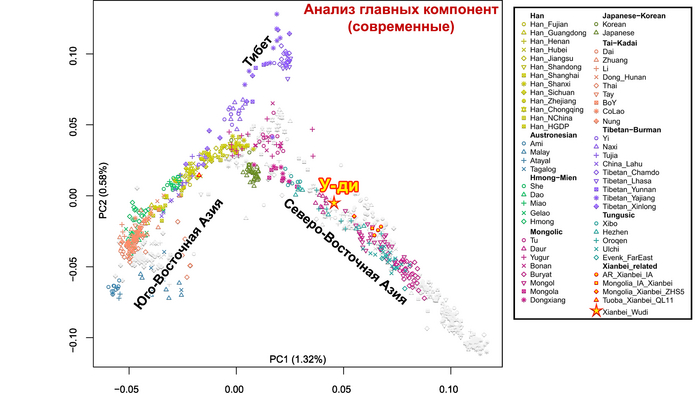
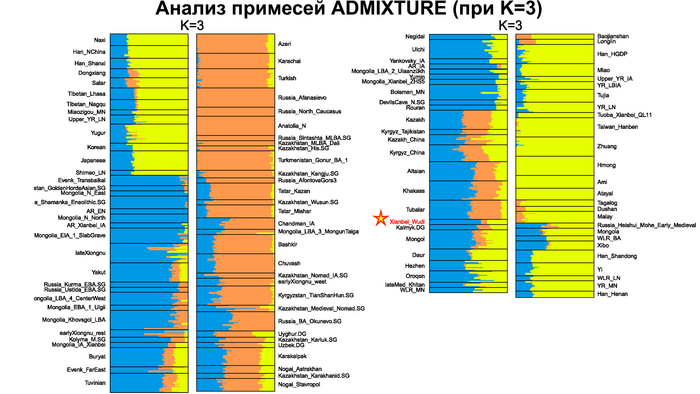
Генетические данные императора У-ди важны и в контексте происхождения представителей племён сяньби. Так, была обнаружена, наиболее тесная связь императора Северной Чжоу с древними киданями и представителями племени хэйшуй народа мохэ, а также с современными даурами и монголами.
1/3
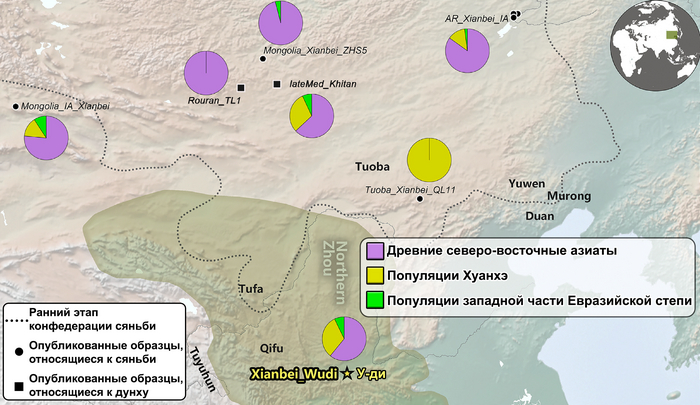
Также авторы исследования смогли сделать вывод о вероятных случаях смешения жителей из Северо-Восточной Азии и долины реки Хуанхэ. В реконструированной генеалогии У-ди его бабушка была из народа когурё в Северо-Восточном Китае, поэтому почти треть его родословной, связанной с популяциями реки Хуанхэ, может быть объяснена последовательными смешанными браками между знатными семьями сяньби и местными китайскими аристократами. Однако у его супруги – императрицы Ашины, как уже было сказано в предыдущем ролике, преобладали компоненты древних северо-восточных азиатов (ANA), без влияния ханьцев связанных с популяциями реки Хуанхэ, что указывает на ограниченное генетическое смешение представителей древнетюркского правящего рода с китайцами. Однако геномные данные самого императора У-ди напрямую свидетельствуют о процессе смешения местной и неместной аристократии.
Помимо этого, в анализе почти у всех представителей сяньби доминировала родословная, связанная с древними северо-восточными азиатами (ANA). При этом группы сяньби продемонстрировали неоднородный генетический состав, тесно связанный с географией. Самые южные представители племен имели до 32% вклада от популяций Хуанхэ, самые западные группы близко к Северному Синьцзяну и горным регионам Алтая имели до ~7% родословной связанной с популяциями западных степных скотоводов, а у сяньби от восточной Монголии до бассейна Амура, вклад древних северо-восточных азиатов был максимален, до 96%.
А генетический профиль Юйвэнь Юна может быть смоделирован как смесь около 62% древних северо-восточных азиатов, 32% китайцев, как представители популяций хуанхэ и 6% западных степных скотоводов. Интересно, что представитель киданей – кочевых монгольских племен – живший примерно через 460 лет после императора У-ди, имел почти тот же генетический состав.
Наблюдаемый генетический ландшафт, вероятно, отражает крупномасштабную миграцию населения из Северо-Восточной Азии и последующее смешение мигрирующих групп древних северо-восточных азиатов с местным населением.
Du P, Zhu K, Qiao H, Zhang J, Meng H, Huang Z, Yu Y, Xie S, Allen E, Xiong J, Zhang B, Chang X, Ren X, Xu Y, Zhou Q, Han S, Jin L, Wei P, Wang CC, Wen S. Ancient genome of the Chinese Emperor Wu of Northern Zhou. Curr Biol. 2024 Apr 8;34(7):1587-1595.e5. doi: https://doi.org/10.1016/j.cub.2024.02.059
Обычно на Пикабу я тестирую лампы, сегодня хочется поговорить про них максимально просто и понятно.
Разбираю я лампочки давно. Прям много лет уже и, честно говоря, это напоминает постоянный поиск компромиссов. На 10 протестированных лампочек приходится только одна, которая более-менее неплохая.
Ну и, казалось бы, это просто лампочка, фиг бы с ним, как-то светит и ладно, зачем париться? А дело в том, что свет не такой уж безопасный как нам кажется. По ссылочке найдете статью, что пугает сбитыми циклами дня и ночи, головными болями, усталостями и провокациями разных очень плохих болячек.
Я не медик, но давайте поговорим про это на бытовом уровне. Вот мы - люди, эволюционно развивались под светом Солнца (вскоре будем эволюционировать под светом монитора, но еще есть время). Вся наша тысячами лет подстраивалась именно под такой спектр излучения.
Вроде логично, значит нам нужны лампы, которые максимально похожи по своим спектральным свойствам на наше светило.
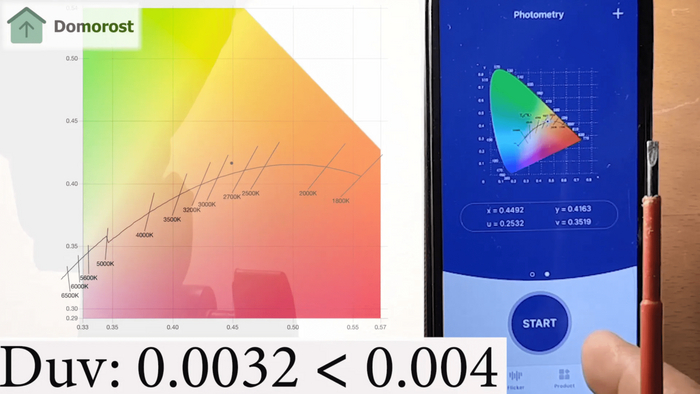
Таким образом (очень упрощенно и именно для нашего контекста), вводится величина которая характеризует похожесть этого вот излучения лампы на излучение солнышка. И она называется индексом цветопередачи - или Cri (Ra), является безразмерной величиной и измеряется от 0-100. Хороший индекс цветопередачи начинается с 90, в лампах же сегодня мы наблюдаем в основном индекс равный 80. Но иногда попадаются и неплохие экземпляры.
Индекс цветопередачи - это конечно хорошо, но наш естественный источник света имеет очень разные свои спектральные характеристики и сам по себе, например, цветовая температура.
И правда, индекс цветопередачи привязан к цветовой температуре, условно он без нее не имеет особенного смысла, так как это значение становится конем в вакууме. И эту штуку тоже я измеряю.
Видите дугу на этой, так называемой, диаграмме цветности? Вот она называется дугой абсолютно черного тела.
Не будем углубляться в понятия, но я всегда находил забавным что спектр абсолютно черного тела теоретический, а наиболее близким к нему физическим объектом является Солнце.
Когда я измеряю спектр излучения лампы (точка на диаграмме), она должна бы попадать на эту дугу, иначе излучение от нее нельзя назвать естественным.
На удивление, тут обычно производители не косячат.
Прекрасно, а что еще в этих лампах есть? Ну например излучение. Оно вообще как бы непрерывное или как лазер из звездных волн запускает импульсы? Вот у Солнышка вполне непрерывное. А у ламп что?
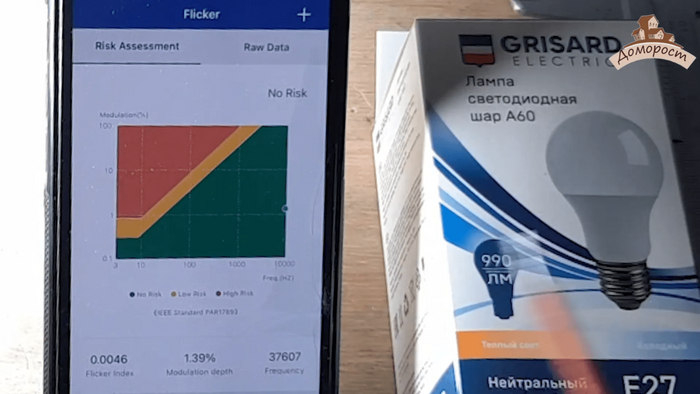
И тут, конечно, проблема. Лампочки светодиодные выдают свет именно условными импульсами (да-да, это бластеры). А задача вашего мозга при этом сгладить такую картинку и сделать из нее удобоваримую. Чтобы все было гладенько. Конечно же это тоже надо измерять - это называется коэффициентом пульсации.
С ним не все так просто, ведь если пульсации большие, но при этом их частота велика, то вроде как мозгу и хорошо, потому придумали целую диаграмму для понимания что хорошо, а что - нет.
Я почти закончил.
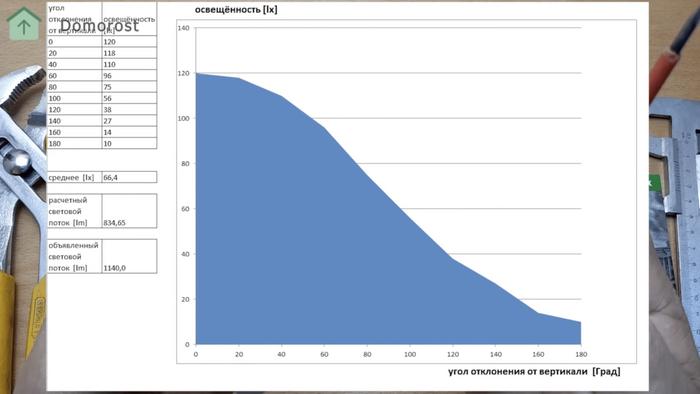
Есть еще такая штука как световой поток. При прочих равных, это то, сколько света даст лампа. фактически представьте что лампочку запихивают в некий объем с датчиками которые понимают сколько света с нее приходят на заданную площадь и потом выдают их интегральную величину.
И эту штуку тоже надо мерить и проверять - тут производители очень часто обманывают.
Вот вроде быстренько мы и прошлись по световым характеристикам ламп. Да, лампочка с виду устройство простое. Оно и правда простое. Но при этом, данная вещь всегда присутствует в нашей жизни. Так уж получилось. Подходите к выбору их с умом.
Зарегистрирован новый солнечный удар по Земле. Пик пришёлся сегодня на 12 часов по московскому времени. В сторону Земли, по некоторым сведениям, ушло не менее 6 крупных выбросов массы. Предварительно к планете дошло пока четыре из них. Магнитная буря держится на уровне G4+ уже более 15 часов. Текущее значение составляет G4.7 (Kp=8.66), что всего на 0.3 балла ниже максимального уровня G5. Промежуточные пики пока были достигнуты ночью между 1 и 5 часами по московскому времени, когда вариации магнитного поля Земли впервые с 2005 года достигли уровня G5.
Шкала магнитных бурь с уровнем от низшего (G1) до высшего (G5) является официально признанной шкалой для классификации событий в магнитосфере Земли. Описание шкалы даёт для событий уровня G4 – G5 cледующие основные признаки:
Магнитные бури уровня G5 Воздействие на энергетические системы: — возможны разрушения энергетических систем и повреждения трансформаторов Воздействие на космические аппараты: — обширный поверхностный заряд; — проблемы с ориентацией; — проблемы со связью и слежением за космическими кораблями.
Воздействие на наземные системы: — токи через трубопроводы достигают сотен ампер; — один или два дня невозможна высокочастотная связь во многих района; — ухудшение точности спутниковых систем навигации; — низкочастотная радио-навигация выходит из строя на несколько часов; — полярные сияния видны вплоть до экватора.
Магнитные бури уровня G4 Воздействие на энергетические системы: — возможны проблемы со стабильностью напряжения; — частичные разрушения энергетических систем и отключение защитных систем.
Воздействие на космические аппараты: — поверхностный заряд и проблемы слежения и ориентации, необходима коррекция Воздействие на наземные системы: — наведенные токи в трубопроводах требуют мер защиты; — спорадическое прохождение ВЧ радиоволн; — ухудшение спутниковой навигации на несколько часов; — отказ низкочастотной радионавигации; — полярные сияния видны до тропиков.
Конкретные последствия конкретных геомагнитных событий индивидуальны. В любом случае, берегите себя и здоровье. Органы чувств человека не способны непосредственно регистрировать переменное магнитное поле и оценить силу воздействия на организм. Доверяйте проверенной информации.
В комментариях к моей предыдущей статье и в комментариях к ролику было много вопросов и некорректных замечаний по поводу парадокса близнецов. Как оказалось, мое объяснение оказалось не настолько понятным, как я надеялся, поэтому в этой статье я решил максимально наглядно, подробно и последовательно объяснить парадокс близнецов и ответить на некоторые другие вопросы.
Для иллюстраций и анимаций я написал интерактивный браузерный визуализатор, где можно двигать ползунки, менять режимы и наблюдать за преобразованиями Лоренца.
Кратко напомню суть парадокса
Берем двух близнецов, сажаем их на маленькую легкую планету (легкую, чтобы не учитывать влияние гравитации), одного оставляем неподвижным, а второго запускаем на ракете полетать и вернуться обратно. При их встрече оказывается, что летавший близнец постарел меньше, чем неподвижный. Парадокс заключается в том, что неочевидно почему именно у летавшего время текло медленнее. Ведь, вроде бы, ситуация симметричная: в системе отсчета летавшего это планета с неподвижным близнецом полетала и вернулась, и это у них должно было натикать меньше времени.
Парадокс близнецов очень важен, т.к. это самый наглядный способ увидеть, что релятивистский эффект замедления времени не просто математический артефакт специальной теории относительности или иллюзия, а вполне реальное физическое явление.
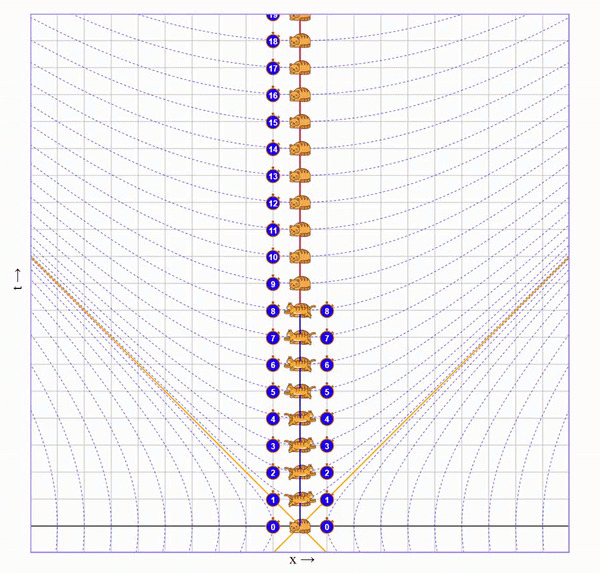
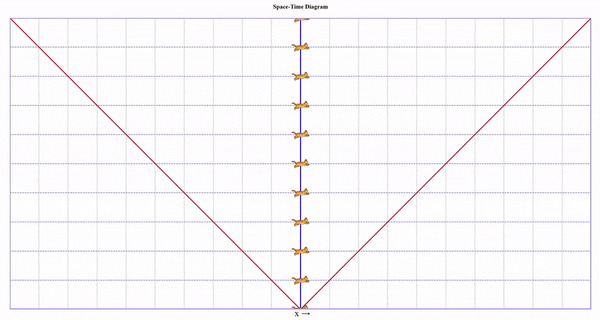
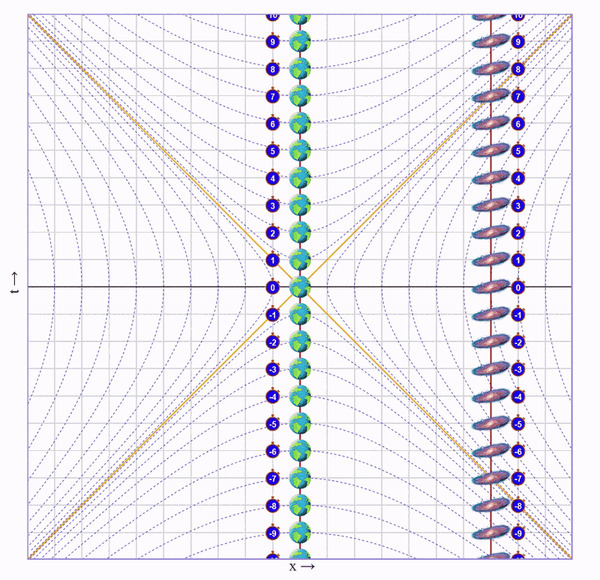
Попросим бегущего кота пробежать вправо со скоростью 75% скорости света, потом развернуться и прибежать с той же скоростью назад. Вот визуализация на диаграмме (по вертикали ось времени, по горизонтали - пространства):
На ней видно, что у бегущего кота натикало меньше времени, чем у неподвижного, но непонятно почему.
Чтобы понять что происходит с каждым из близнецов, нужно посмотреть на ситуацию от лица каждого из них.
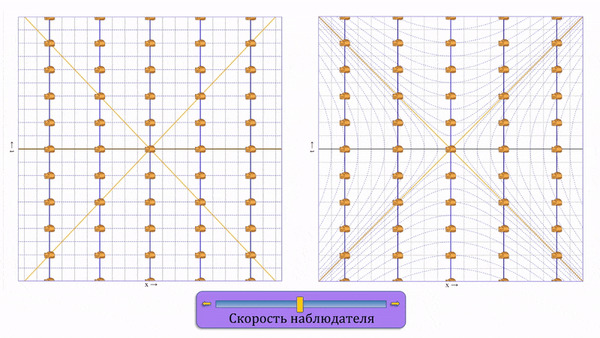
Напомню, что в специальной теории относительности при изменении скорости наблюдателя, точки на диаграмме сдвигаются не только вдоль оси пространства, но еще и вдоль оси времени. Отсюда неизбежно вытекают все релятивистские эффекты (замедление времени, сокращения длин, относительность одновременности).
Слева классическое преобразование Галилея, справа - преобразование Лоренца, которое пришло ему на смену. Желтые прямые иллюстрируют скорость света в обоих направлениях.
Подробнее о том, почему так происходит, я рассказывал в предыдущей статье и видео. Если коротко, то все дело в том, что, согласно экспериментам, один и тот же "пучок" света летит со скоростью 299 792 458 м/с относительно любого наблюдателя. Независимо от того, как быстро и в каком направлении этот наблюдатель движется относительно источника этого света. Иначе говоря, как бы быстро ты ни двигался, свет все равно улетает от тебя со скоростью света.
Этот факт противоречит привычному преобразованию Галилея:
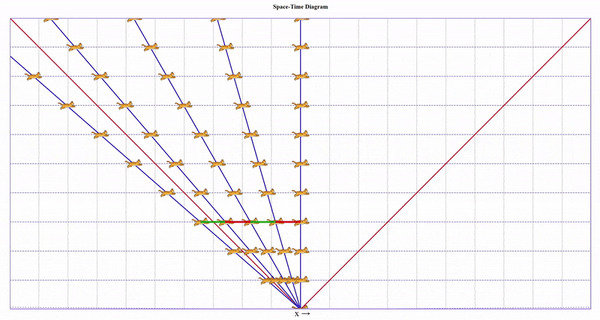
Допустим, мы бежим на коте с огромной скоростью (30% скорости света). Затем мы с поверхности этого кота выбегаем на еще одном коте с такой же скоростью относительно первого кота. Потом делаем так же еще три раза.
В привычном Галилеевском мире получается, что расстояния между котами в каждый момент времени одинаковы, а от первого мы удаляемся быстрее, чем со скоростью света (тут скорость света показана бордовым).
Чтобы примирить факт постоянства скорости света с физикой, пришлось изменить преобразования Галилея (где при изменении скорости наблюдателя, точки на диаграмме смещаются горизонтально) и превратить их в преобразования Лоренца (где точки сдвигаются еще и во времени, ассимптотически приближаясь к линии скорости света). Кстати, математически это является вращением в 4-хмерном пространстве-времени с метрикой Минковского.
Обратите внимание на синие и красные отрезки. В Галилеевском варианте сохраняются их длины, которые считаются как(по теореме Пифагора), а в Лоренцевом - сохраняется интервал, который считается как (метрика Минковского).
Можно сколько угодно раз делать трюк с котом, но первый кот никогда не достигнет бордовой линии. Его линия будет лишь ассимптотически к ней приближаться и растягиваться. Сам свет, при этом, летит со скоростью света относительно любого из котов.
Итак, вернемся к парадоксу близнецов
Представим, что один близнец сидит на Земле, а второго мы попросили слетать в соседнюю галактику и вернуться. Пускай галактика находится на таком расстоянии, чтобы по часам подвижного близнеца на всё путешествие ушло 8 секунд.
Начнем с неподвижного близнеца на Земле:
Тут все просто. Мы ждем 8 секунд и ничего не происходит.
Теперь рассмотрим ситуацию от лица движущегося близнеца.
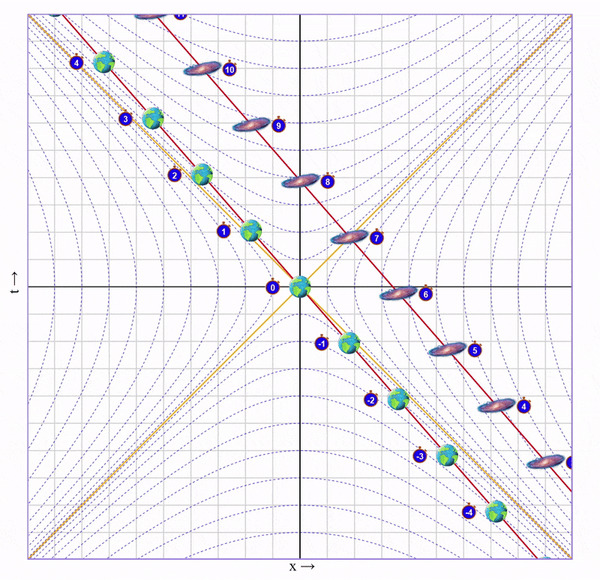
1) Сначала мы быстро набираем скорость (ускоряемся очень быстро, поэтому мы еще не успели пролететь значимое расстояние).
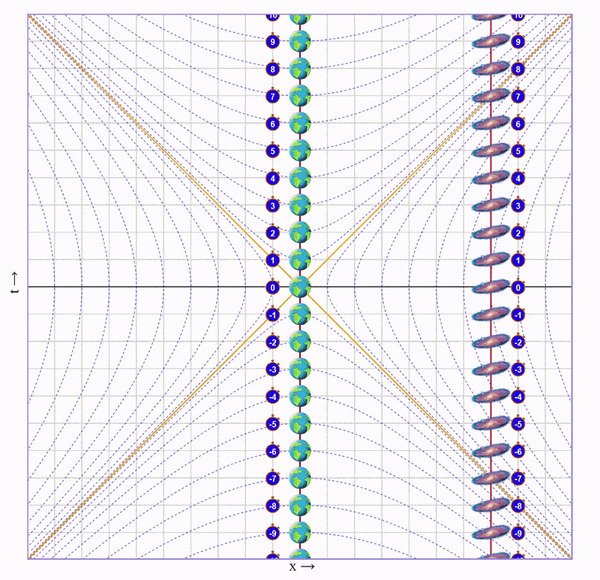
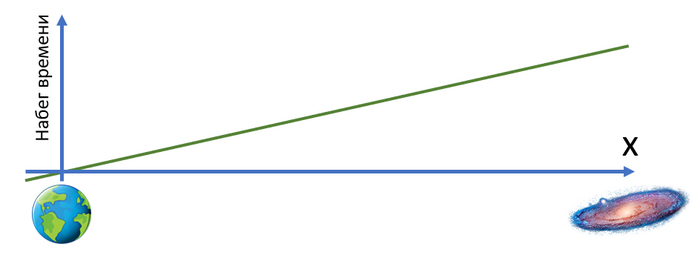
Видно, что время на галактике сместилось в будущее на 6 с лишним секунд. Но мы этого сразу не заметим, ведь это смещение времени увеличивается постепенно вдоль оси от нас к галактике.
График набега времени:
А еще расстояние до галактики уменьшилось (до ускорения между Землей и галактикой было 7 клеток, а стало ~3.5) и ход времени в галактике для нас замедлился (вертикальное расстояние между соседними изображениями галактики стало больше).
В этот момент летящему близнецу недоступна информация о том, что время на галактике сместилось в будущее и что расстояние уменьшилось. Ведь он видит только то, что непосредственно достигло его глаз.
Чтобы не было сомнений, что все по честному, все сдвиги точек на диаграмме происходят только по формулам преобразований Лоренца (это можно проверить по исходникам визуализатора).
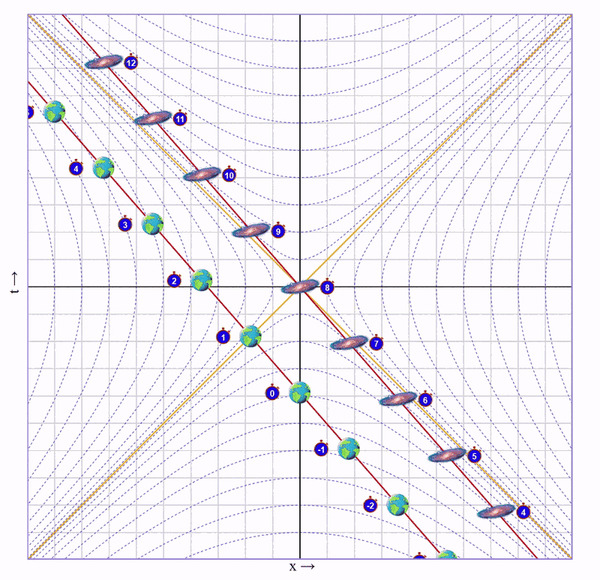
Итак, скорость набрана. Теперь летим с этой скоростью пока не достигнем галактики. На это уйдет 4 секунды (для этого я добавил в визуализатор ползунок "Wait"):
Пока мы летели мы собирали этот ускоренный и сжатый свет от галактики и по ходу движения наблюдали ее эволюцию в быстрой перемотке. В итоге, в точке назначения мы видим, что по нашим часа прошло 4 секунды, а на галактике - 8.
Важный момент: Мы летим к галактике, поэтому ее время для нас замедлено. Но глазами мы видим ее наоборот ускоренной, потому что собираем испущенный от нее сплющенный свет. Иначе говоря, расстояние до галактики скукожилось сильнее, чем замедлилось ее время.
Теперь осталось развернуться, с такой же скоростью полететь обратно на Землю:
И остановиться:
Что мы в итоге видим? Пока подвижный близнец бегал 8 секунд, у неподвижного прошло 16.
Когда подвижный близнец летит без ускорений, ситуация симметричная. Каждый из них считает, что время замедлено у другого. Но именно во время ускорений подвижного близнеца "съедается" время неподвижного, что можно видеть на анимациях.
Итог: Парадокс близнецов разрешен тем, что ускорение одного из котов вносит асимметрию в систему и приводит разнице их возраста. Этот эффект действительно существует, он измерен экспериментально и явно следует из преобразований Лоренца, как и все остальные релятивистские эффекты специальной теории относительности.
Нужна ли общая теория относительности для объяснения парадокса близнецов?
Нет, ОТО нужна там, где нужно учитывать гравитацию. В нашем случае в этом нет необходимости.
В начале 1980-х инженер Вик Тэнди устроился на работу в компанию по производству медицинского оборудования в Уорике, графство Уорикшир, Англия. В примитивной лаборатории, состоявшей из двух стальных гаражей, соединенных вентиляционными трубами, царила обычная обстановка - негромкий гул вентиляторов и помп от оборудования. Ничто не предвещало чего-либо из ряда вон выходящего, пока не появился призрак.
Однажды утром Тэнди застал перепуганную уборщицу, выбегавшую из лаборатории. Она не могла объяснить, что произошло, кроме объявшего её чувства ужаса и отчетливого ощущения присутствия чего-то потустороннего. Тэнди списал это на то, что женщина отработала ночную смену в одиночестве в старом скрипучем здании. Однако в последующие дни Тэнди и его два скептически настроенных коллеги заметили необъяснимую тяжелую атмосферу в лаборатории. Тэнди описывал это как "гнетущее" чувство и что ему вдруг становилось холодно. Случались и другие странности — однажды кто-то из работников работал за верстаком и явственно почувствовал присутствие кого-то за спиной, но, обернувшись, никого не обнаружил. В другой раз, когда Тэнди был один, ему явственно привиделся смутный серый силуэт, но стоило ему повернуть голову, чтобы рассмотреть внимательнее, фигура исчезла. Коллеги решили, что все дело в переутомлении.
Однажды Тэнди зажал в тисках ручку своей фехтовальной рапиры, оставив лезвие торчать наружу. Отойдя на минуту, он вернулся и обнаружил, что лезвие яростно колеблется вверх-вниз под действием невидимой силы. Тэнди схватил рапиру вместе с тисками и колебания прекратились. Но стоило отпустить их и лезвие вновь заколебалось. Тэнди решил обойти лабораторию с рапирой, и обнаружил, что лезвие неподвижно у стен, но колебания усиливаются к центру помещения, особенно возле верстака — как раз там, где ему ранее причудился серый силуэт.
Будучи опытным инженером, Тэнди сделал вывод, что в комнате подвергается концентрированному воздействию звуковых волн, заставляющих рапиру вибрировать и вызывающие у сотрудников чувство присутствия человека. Исследуя эту гипотезу, он выяснил, что недавно в лаборатории был установлен новый вытяжной вентилятор. Это совпало по времени со случаем с уборщицей. Вентилятор в сочетании с геометрией помещения создавал стоячую волну на частоте чуть менее 19 Гц — в области инфразвука, недоступной для человеческого уха, но близкой к резонансной частоте человеческого глаза, что и вызывало едва заметные колебания глазных яблок сотрудников, порождавшие оптические иллюзии. Когда вентилятор заменили, "призрак" исчез, и лаборатория вернулась к нормальному режиму работы.
Считается, что инфразвук может вызывать схожие паранормальные феномены и в других ситуациях, например, когда органные трубы порождают низкочастотные звуковые волны, создавая ощущение присутствия. Наблюдения Тэнди привели к публикации научной статьи об этом явлении, и до самой смерти в 2005 году он посвятил себя разоблачению паранормальных утверждений на основе своей случайно обретенной экспертизы.
Примечание: текста много, тема сложная и состоит из двух постов. Разделы – "Социокультурные изменения" и "Итоги" (Появление и сокращение генетического разрыва), во второй части.
Вступление
Генетическое разнообразие человеческих популяций Западной Евразии в значительной степени сформировалось в результате трех основных доисторических миграций:
1. Миграции охотников-собирателей, а именно анатомически современных людей, находящихся на ранней стадии социально-экономического развития, которые занимали территорию, примерно, с 45 тыс. лет назад. При этом вклад в последующие популяции внесли выжившие их линии после последнего ледникового максимума около 25-19 тыс. лет назад.
2. Далее, гораздо ещё более существенный вклад, внесли ранние неолитические земледельцы, пришедшие с Ближнего Востока примерно с 11 тыс. лет назад.
3. Степные скотоводы, распространившиеся из Причерноморско-Каспийской степи около 5000 лет назад, привнесли третью основную генетическую компоненту в популяции Западной Евразии.
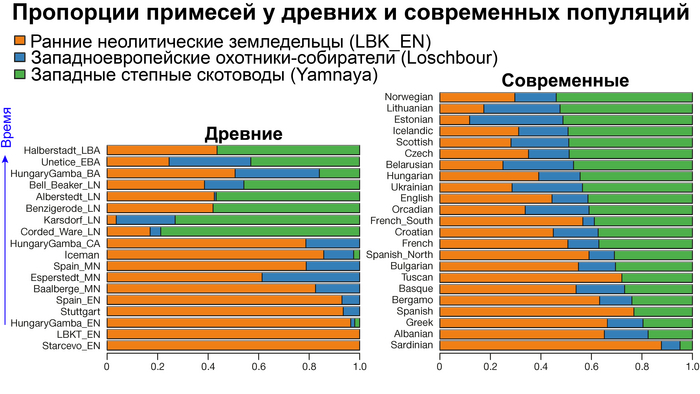
Аутосомный генофонд практически всех европейских популяций по большей части состоит из различных пропорций этих предковых компонент, при этом независимо какие у них гаплогруппы.
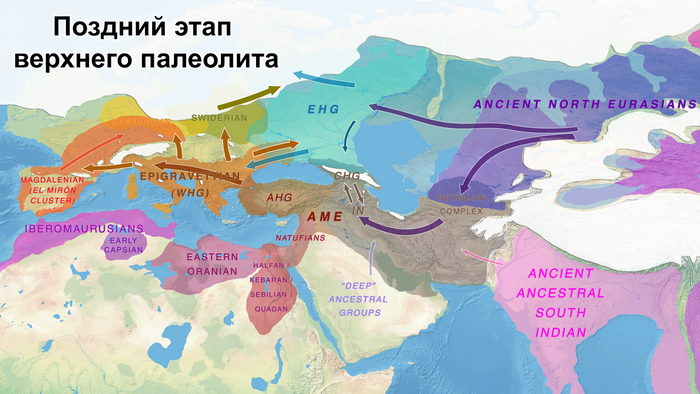
Европейский континент заселялся людьми современной анатомии в несколько волн, при этом не все линии этих людей были предками современных. Первые «сапиенсы» Европы (более 40 тыс. лет назад) пересекались и скрещивались с неандертальцами, пока те совсем не вымерли. Об этой примеси свидетельствуют геномы людей из пещеры Бачо Киро в Болгарии, связанных с археологической культурой начального верхнего палеолита, и со стоянки Пештера-ку-Оасе в Румынии. Другие люди того периода, такие как женщина из пещеры Златый кунь Чехии и человек из Усть-Ишима России, не несут в своих геномах значительного количества неандертальского вклада, чем другие неафриканские группы. При этом люди, жившие в Европе более 40 тыс. лет назад, не оставили существенных следов в генетическом составе современных евразийских популяций, что говорит о том, что они в значительной степени вымерли или были ассимилированы последующими группами людей современной анатомии.
В те времена, на демографию древних людей влияли различные факторы, в том числе и климатические. Пик похолодания во время последнего ледникового максимума (LGM) был около 21 тыс. лет назад, когда Северная Европа была в значительной степени покрыта льдом, а южная граница ледниковых щитов проходила через Германию и Польшу. Многие участки Европы стали холодными пустынями с тундростепями и лесостепями на побережье Средиземного моря. А люди и животные переместились в южные широты, о чем свидетельствует, например, солютрейская культура на Пиренейском полуострове и на юге Франции. Другими предложенными рефугиумами или климатическими убежищами в тот период были Апеннинский полуостров, Балканы и юго-восточная часть Европейской равнины.
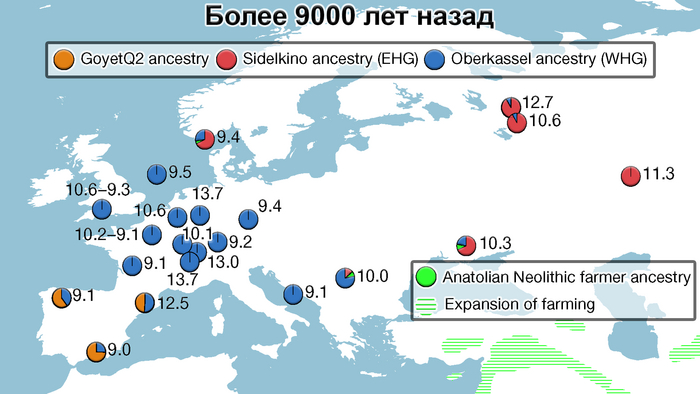
А в результате послеледниковой колонизации, палеогенетический анализ выявил разделение популяций охотников-собирателей на западных и восточных. Источники родословной западных охотников-собирателей (WHG) связаны с носителями эпиграветтской, азилийской и других эпипалеолитических культур генетического кластера Виллабруна, появление которого обусловлено миграциями в конце ледникового периода и появлением новой генетической компоненты, связанной с Ближним Востоком того времени, начиная с бёллинг-аллерёдского потепления около 14 тыс. лет назад, что археологически коррелирует с культурными переходами.
А родословная восточных охотников-собирателей (EHG) демонстрирует дальнейшую примесь с верхнепалеолитическими популяциями Сибири - древними северными евразийцами (ANE).
Предковый состав западных охотников-собирателей (WHG) был регионально изменчивым в мезолитических популяциях. Если охотники и собиратели из Британии и северо-западной континентальной Европы продемонстрировали однородный состав от палеолита и мезолита, а также вплоть до неолитической экспансии около 6000 лет назад, то их коллеги из Испании, представляли собой смесь людей из двух рефугиумов, когда к генетическому профилю кластера Виллабруна, добавляется примесь от более ранних местных пиренейских охотников-собирателей, связанных с солютрейской и мадленской культурами верхнего палеолита.
Этот генетический профиль сохранился во время последнего ледникового максимума в Юго-Западной Европе около 25-19 тыс. лет назад.
А время появления примеси сформировавшей восточных охотников-собирателей (EHG), оценивается в 15–13 тыс. лет назад, однако её пропорции в популяциях, по-видимому, соответствуют клине, которая в целом коррелирует с географией, так охотники-собиратели Прибалтики и Украины демонстрируют большее сходство с предками кластера Виллабруна, чем охотники и собиратели западной части современной России, у которых было больше вклада от древних северных евразийцев (ANE).
В свою очередь скандинавские охотники и собиратели продемонстрировали различные пропорции смеси западных и восточных охотников и собирателей (WHG и EHG) в позднем мезолите.
Стоит отметить, что охотники и собиратели мезолита и тем более палеолита, были очень разнообразны и неоднородны, но из-за ограниченного количества данных, наши знания о мезолитической структуре населения и процессах демографического смешения долгое время были скудны и содержали существенные хронологические и географические пробелы в информации. Отчасти это объясняется малым количеством хорошо сохранившихся мезолитических человеческих останков возрастом более 8000 лет, а отчасти и потому, что большинство исследований древней ДНК, посвященных периодам мезолита и неолита, были ограничены преимущественно находками из Европы.
1/3
Хотя давно напрашивается более детальная классификация охотников и собирателей по их характерным отличиям. Так, археологические данные указывают на условную границу от восточной Балтики до Черного моря, к востоку от которой сообщества охотников и собирателей существовали гораздо дольше, чем в Западной Европе, несмотря на аналогичное расстояние до центра распространения раннего земледелия на Ближнем Востоке.
Компоненты восточного и западного происхождения у охотников-собирателей в этом пограничном регионе сильно различаются, но более точные представления о демографических процессах, связанных с разделением на восток и запад и о динамике численности населения к востоку от Европы, включая Северную и Центральную Азию за тот же период времени, весьма ограничены. В этих регионах термин «неолит» характеризуется культурными, экономическими и социальными изменениями, а также использованием керамики. К примеру, неолитические культуры степей Центральной Азии и таежного пояса России уже изготавливали керамику, однако продолжали вести прежний образ жизни с присваивающей экономикой, как у предшествующих мезолитических культур охотников-собирателей, с различными инновациями в зависимости от условий среды обитания. Однако пока недостаточно данных для понимания того, как неолитизация отличалась по своим срокам, механизмам и последствиям в разных уголках Евразии.
В Европе, кроме восточной её части, переход от охоты и собирательства к земледелию сопровождался прибытием мигрантов ближневосточного происхождения, практикующим производящее хозяйство на основе одомашненных растений и животных. С последующим значительным ростом населения. В Юго-Восточной, Центральной и Западной Европе анатолийские земледельцы впоследствии в некоторой степени смешались с местными охотниками и собирателями, а на территории Британии, прибывшие с континентальной Европы ранние европейские земледельцы полностью заменили местных охотников и собирателей.
А вот в восточно-балтийском регионе наблюдается заметно иная модель неолитизации, с появлением одомашненных животных только при появлении культуры шнуровой керамики (CWC) около 4800 лет назад. Аналогичным образом в Северном Причерноморье люди с генетическим профилем мезолитических охотников и собирателей тысячелетиями сосуществовали с земледельческими группами, расположенными дальше на запад.
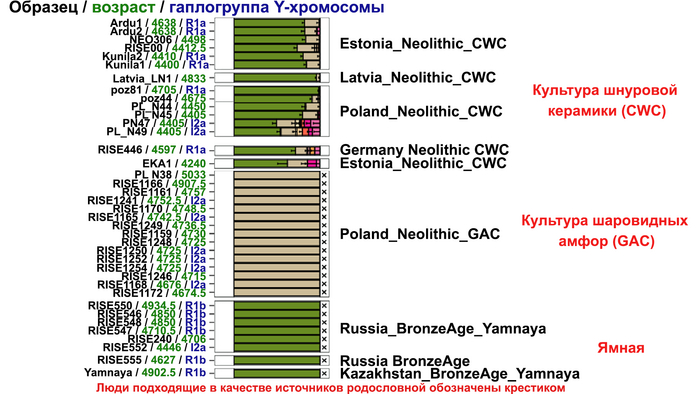
И третий крупнейший генетический пласт в истории европейских популяций связан со степными скотоводами раннего бронзового века, как представители ямной культуры около 5 тыс. лет назад. Этот генетический профиль, быстро распространился по Европе посредством экспансии как носителей культуры шнуровой керамики (CWC), так и родственных ей культур.
Хотя предыдущие исследования выявили эти крупномасштабные миграции в Европу и Центральную Азию, основные аспекты, касающиеся демографических процессов, не решены. Ямная или степная родословная в общих чертах охарактеризована как смесь восточноевропейских (EHG) и кавказских охотников-собирателей (CHG), образовавшаяся в результате гипотетической смеси «северного» степного и «южного» кавказского источников. Однако точное происхождение этих предковых источников не было установлено. Более того, за некоторым исключением, известные гаплогруппы Y-хромосомы представителей ямной культуры не соответствуют гаплогруппам, обнаруженным у европейцев после 5000 тыс. лет назад и происхождение этой отцовской линии также не установлено. К тому же "степная" родословная в Европе до сих пор была выявлена только в смешанной форме, а детали и механизмы, с помощью которых эта родословная впоследствии распространилась вместе культурой шнуровой керамики (CWC), оставались неясными.
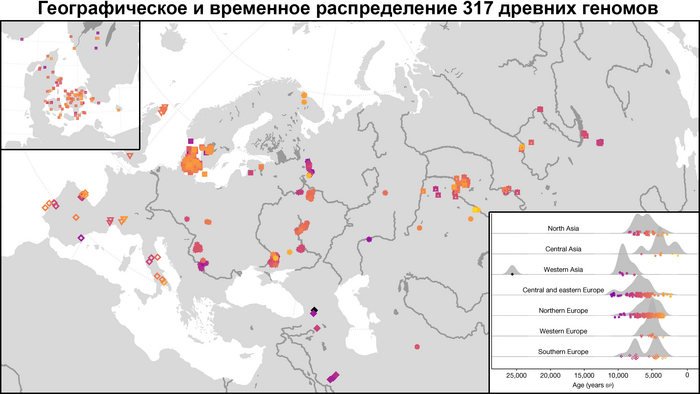
В новой работе учёные под руководством Эске Виллерслева из образцов, датированных радиоуглеродным методом при помощи ускорительной масс-спектрометрии, секвенировали геномы 317 человек, 309 из них датируются периодом от 11 до 3 тысяч лет назад.
А совместно с ранее опубликованными данными получилась выборка из 1600 древних геномов, секвенированных с помощью метода дробовика.
Географически 317 отобранных образцов охватывают обширную территорию Евразии, по долготе от Атлантического океана до озера Байкал, а по широте от Скандинавии до Ближнего Востока, и происходят из контекстов, которые включают курганы, пещеры, болота и дно водоёмов.
Географически образцы можно разделить на три региона:
1. Центральная, Западная и Северная Европа.
2. Восточная Европа, включая западную часть России, Беларусь и Украину.
3. Урал и Западная Сибирь.
Образцы включают представителей многих ключевых культур Западной Евразии мезолита, неолита и последующих периодов, такие как маглемозе, эртебёлле, воронковидных кубков (TRB) и шнуровой керамики или одиночных погребений в Скандинавии; импрессо или кардиумной керамики в Средиземноморье; кёрёш и линейно-ленточной керамики (LBK) в Юго-Восточной и Центральной Европе; и многие археологические культуры Украины, западной части России и Зауралья (как веретьё, льяловская, волосовская, китойская и др.). Выборка была особенно плотной в Дании, где сотня геномов, охватывала период от раннего мезолита до бронзового века. Обширная выборка была также получена из Украины, Западной России и Зауралья, охватывая период от раннего мезолита до неолита, примерно до 5000 лет назад.
Результаты
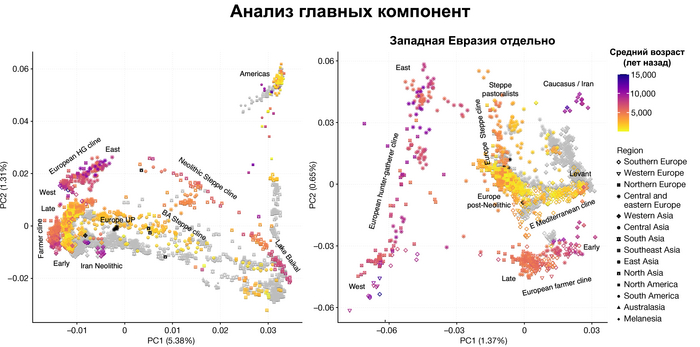
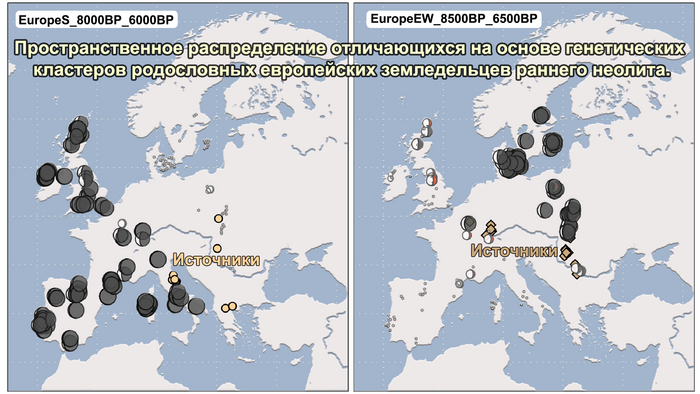
Полученный набор генетических данных позволил при анализе главных компонент использовать за основу древние геномы, и уже на них спроецировать положение современных популяций, а не наоборот, как это было во многих предыдущих исследованиях.
И хотя результаты согласуются с тем, что древние люди сильнее друг от друга генетически отличались, чем современные, такой подход позволил выявить ещё более значительные генетические различия между древними людьми, которые ранее не наблюдались, что отражает меньший эффективный размер их популяции и большую генетическую изоляцию.
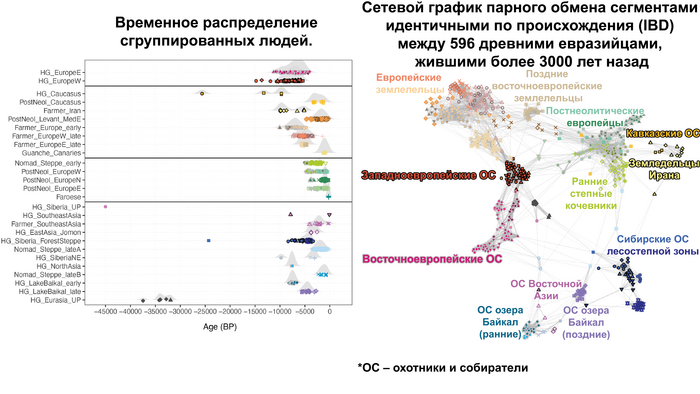
Чтобы получить более точную генетическую характеристику в пространстве и времени, авторы использовали попарную идентификацию сегментов ДНК идентичных по происхождению, чтобы сгруппировать анализируемых людей в связанные кластеры сходного генетического происхождения, а не по культурным или территориальным признакам.
В более крупном масштабе они отражали ранее описанные группы западных и восточных охотников-собирателей, а вот в более мелком масштабе они сгруппировались не только в пределах ограниченных пространственно-временных диапазонов и/или археологических контекстов, но также продемонстрировали ранее неизвестные связи в более широких географических диапазонах.
Полученные кластеры далее использовались для моделирования состава предков последующих популяций.
Популяционная структура охотников и собирателей после максимума последнего оледенения (LGM)
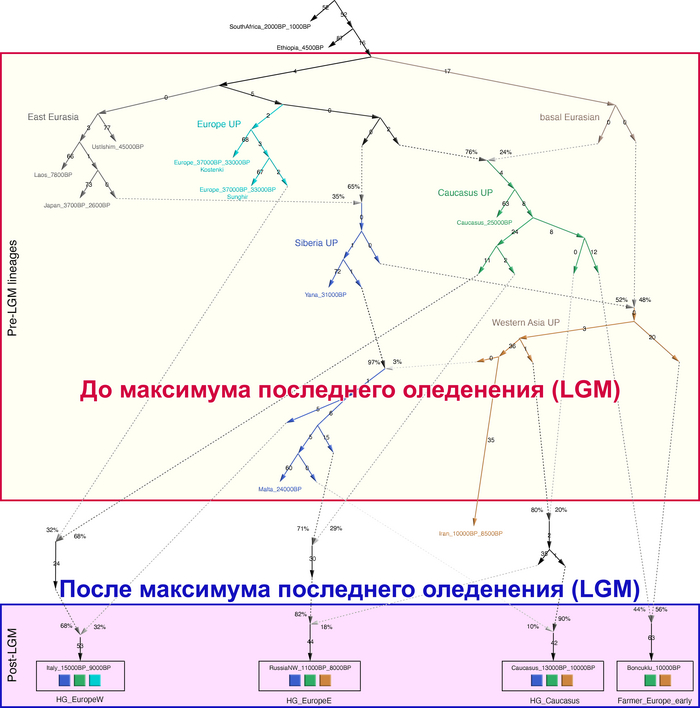
До последнего ледникового максимума, популяции Евразии довольно сильно отличались и в разной степени смешивались с неандертальцами. Некоторые популяции пока остаются гипотетическими, как к примеру, так называемые, «базальные евразийцы» с незначительной или нулевой примесью от неандертальцев, их прямых образцов, подтвержденных генетикой, на данный момент нет, но генетические следы этой популяции были обнаружены у охотников-собирателей Кавказа и Западной Азии верхнего палеолита, а также у более поздних западноевропейских охотников и собирателей.
В данной работе авторы сообщают о генетических данных полученных из зуба женщины верхнего палеолита, жившей около 26-25 тыс. лет назад в окрестностях карстового грота Котиас Клде в современной Грузии, не путать с более молодыми образцами кавказских охотников-собирателей из этого археологического комплекса, у которых было менее 20% этой кавказской родословной верхнего палеолита. А сам геном этой представительницы Кавказа верхнего палеолита представляет собой смесь 76% западноевразийских охотников-собирателей верхнего палеолита (т.е. до максимума последнего оледенения) и 24% «базальных евразийцев». Кстати популяции Ближнего Востока верхнего палеолита, почти наполовину произошли от «базальных евразийцев», которые, практически не имели примеси неандертальцев и отделились от других неафриканских линий ещё до их разделения между собой. При этом западноевропейские охотники-собиратели, жившие после последнего ледникового максимума, моделируются как итоговая, за несколько событий, смесь 46% популяций, связанных с Кавказом верхнего палеолита, 22% европейцев, живших около 33-37 тыс. лет назад, связанных со стоянками Сунгирь и Костёнки и 32% древних популяций, связанных с Сибирью верхнего палеолита, как представитель стоянки Мальта, возрастом 24 тыс. лет, т.е. древних северных евразийцев.
В свою очередь восточноевропейские охотники-собиратели моделируются как смесь 58% популяций, связанных с обитателями стоянки Мальта в Прибайкалье, 28% популяций, связанных с Кавказом верхнего палеолита, как женщина из Котиас Клде (но в разные периоды, сначала 24%, а потом 4%) и 14% представителей Западной Азии верхнего палеолита. Эта смесь также проходила в несколько этапов. Тем самым новые данные подтверждают генетическое разделение охотников и собирателей в Восточной и Западной Европе после максимума последнего оледенения, когда люди снова расселялись по Евразии, при этом эти различия сохранялись на протяжении всего мезолита.
В анализе примесей, эти сложносоставные генофонды представлены отдельными компонентами. А различия между ними, а именно между западноевропейскими и восточноевропейскими охотниками-собирателями мезолита максимально выражены у представителей Италии, возрастом от 15 до 9 тыс. лет и Северо-Запада современной России возрастом от 11 до 8 тыс. лет.
Обратите внимание, что генетический состав одинаков даже у людей с разными гаплогруппами и наоборот, по всему геному люди с одинаковыми линиями гаплогрупп отличаются.
Кстати, в представлении сложных процессов, не стоит забывать о временных промежутках, о которых идёт речь.
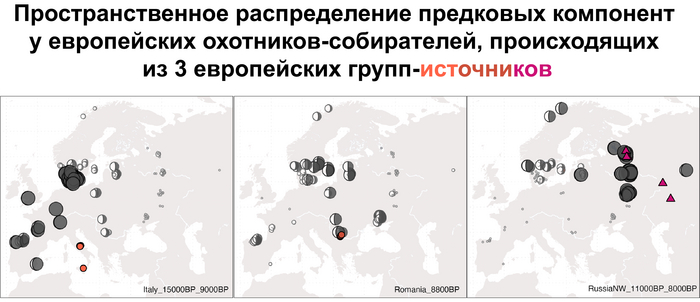
Что касается североевропейских охотников и собирателей, то они отличались по пропорциям смеси компонент как у западноевропейских и восточноевропейских охотников-собирателей, за исключением Дании, где популяции продемонстрировали южноевропейское происхождение и были генетически схожи с охотниками-собирателями из Италии, возрастом от 15 до 9 тыс. лет. При чём эта родословная сохранялась в регионе на протяжении 5 тыс. лет.
В целом результаты указывают на то, что существовало по меньшей мере три отчетливые волны миграций охотников и собирателей в Скандинавию:
1. Первая волна связана с миграцией людей южноевропейского происхождения на территорию Дании и прибрежной части Юго-Западной Швеции.
2. Вторая волна в Прибалтику и Юго-восточную Швецию связана с популяциями из Юго-Восточной Европы.
И 3. Третья волна миграций на крайний север Скандинавии, а затем на юг вдоль атлантического побережья Норвегии связана с охотниками и собирателями с северо-запада современной России.
Эти миграции, вероятно, представляют собой послеледниковую экспансию из рефугиумов или районов-убежищ, общих как для людей, так и для многих видов растений и животных.
Стоит отметить, что генофонд европейских охотников-собирателей верхнего палеолита, живших до максимума последнего оледенения был в значительной степени заменён генофондом, происходящим из рефугиума в Южной Италии. На Пиренейском полуострове самые древние образцы из этого исследования, в том числе охотника-собирателя, возрастом около 9500 лет (NEO694) из бухты Санта-Майра на востоке Испании, показали преимущественно южноевропейское происхождение с незначительным вкладом охотников и собирателей верхнего палеолита, который, вероятно, отражает генофонд, связанный с мадленским периодом до максимума последнего оледенения.
А более поздние жители севера Пиренейского полуострова, были ближе к охотникам и собирателям Юго-Восточной Европы, демонстрируя около 40–30% вклада балканских охотников-собирателей.
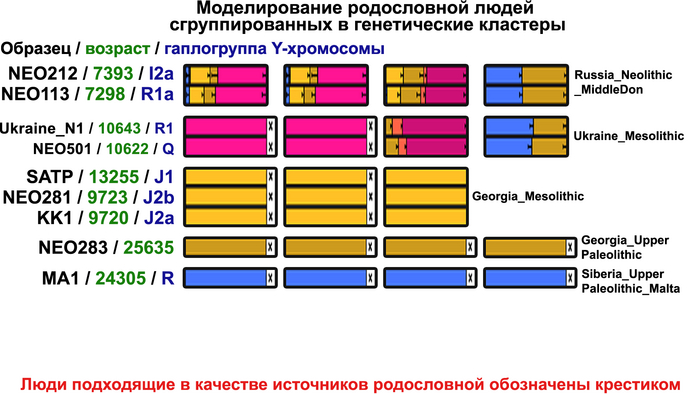
Поток генов, связанных с охотниками-собирателям Юго-Восточной Европы был направлен и в Восточную Европу после мезолита, на территорию современной Украины и далее. А у двух человек из могильника мариупольского типа раннего энеолита (NEO212, NEO113 ) близ хутора Голубая Криница на юге Воронежской области, возрастом около 7300 лет, генетический профиль состоял уже из смеси более ранних охотников-собирателей с территории Украины с вкладом от 18 до 24% из источника, связанного с охотниками-собирателями Кавказа.
1/2
Поэтому на графике анализа главных компонент они отклонены в сторону Ирана и Кавказа. По расчётам эта примесь произошла примерно 8300 лет назад. Результаты указывают на более ранний генетический контакт между популяциями Кавказа и степи ещё до появления культур кочевников в Причерноморско-Каспийской степи. Помимо этого, этот контакт задокументирован западнее, чем предполагалось ранее.
Основные генетические изменения в Европе
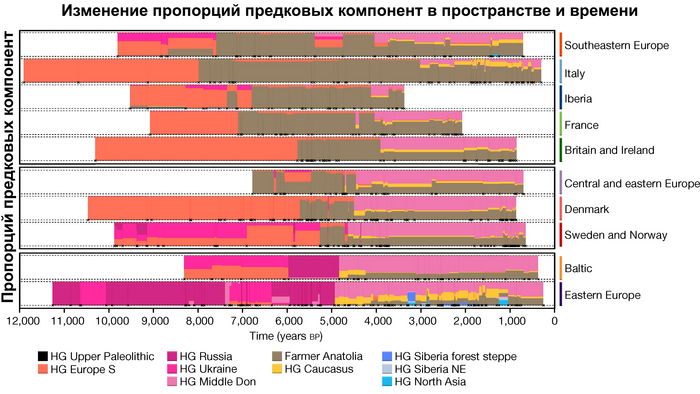
Предыдущие исследования древней геномики задокументировали несколько эпизодов крупномасштабной смены населения в Европе за последние 10 тыс. лет, но 317 новых геномов из данной работы, заполняют важные пробелы в наших знаниях.
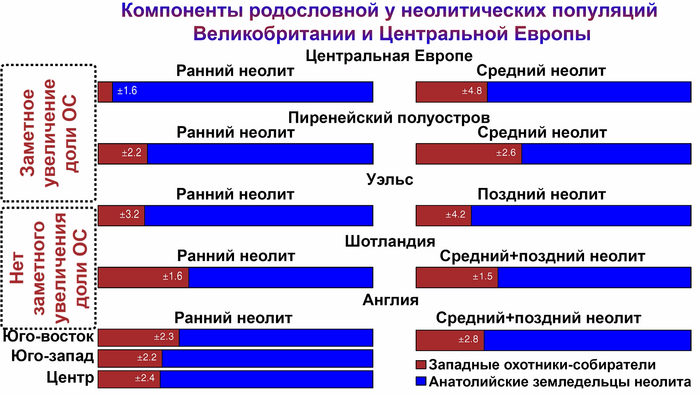
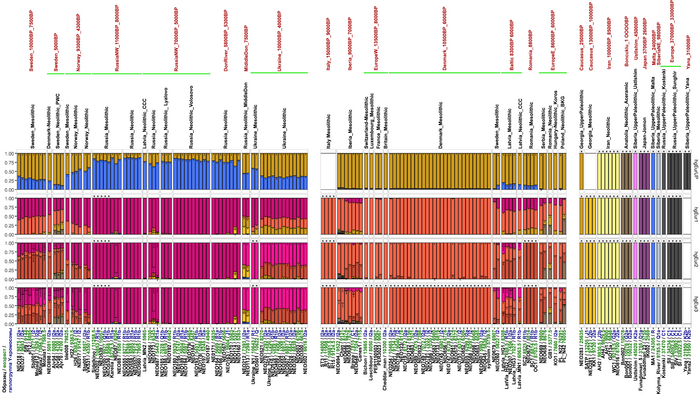
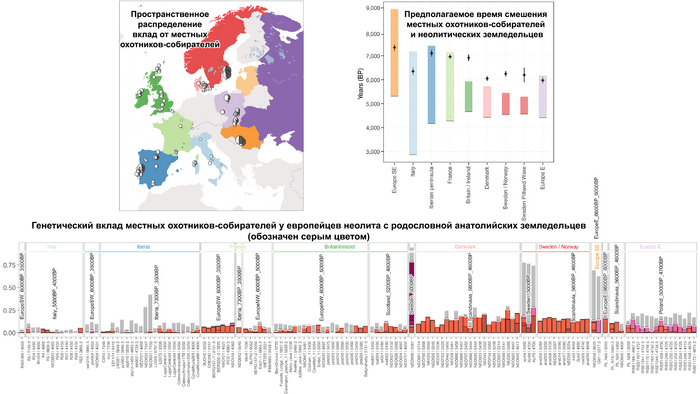
Анализ выявил глубокие различия в пространственно-временной динамике неолитизации между восточной и западной частями Европы вдоль пограничной зоны, простирающейся от Черного моря до Балтийского. К западу от этой пограничной зоны, переход к неолиту сопровождался крупномасштабными сдвигами в генетической родословной, от местных охотников-собирателей к земледельцам анатолийского происхождения (Boncuklu_10000BP). Этот переход в зависимости от региона охватывал обширный период времени, более 3 тыс. лет, от самых ранних свидетельств на Балканах около 8700 лет назад до 5900 лет назад в Дании.
В последующие периоды смешение между ранними европейскими земледельцами и местными охотниками-собирателями привело к частичному увеличению родословной охотников и собирателей во многих регионах Европы, но этот вклад редко превышал 10%, помимо заметных исключений, наблюдаемых у людей из района Железных ворот на границе современных Сербии и Румынии, а также у носителей культур ямочной керамики из Швеции и кардиумной керамики раннего неолита из Португалии, которые, по оценкам, содержат от 27 до 43% вклада от охотников и собирателей, как представители Пиренейского полуострова, возрастом от 9 до 7 тыс. лет (Iberia_9000BP_7000BP).
Обитатели эпохи неолита с территории современной Дании продемонстрировали до 25% генофонда охотников-собирателей (EuropeW_13500BP_8000BP), но преимущественно не местных.
Также обнаружены региональные различия в происхождении последующих неолитических групп. Так последующие неолитические популяции Западной Европы происходят от ранних земледельцев Южной Европы. А в последующие неолитические группы Восточной Европы и Скандинавии внесли свой вклад преимущественно ранние земледельцы Центральной Европы.
А вот с восточной стороны упомянутой пограничной зоны, на территории Прибалтики, Украины и западной части России, не наблюдается существенных генетических изменений, а предковые линии местных охотников и собирателей, без заметного вклада от анатолийских земледельцев,сохраняются примерно до 5 тыс. лет назад. Эта генетическая преемственность согласуется с археологическими данными, которые свидетельствуют о сохранении в этом обширном регионе групп охотников, рыболовов и собирателей, которые использовали керамику, но не практиковали земледелие и животноводство ещё на несколько тысяч лет.
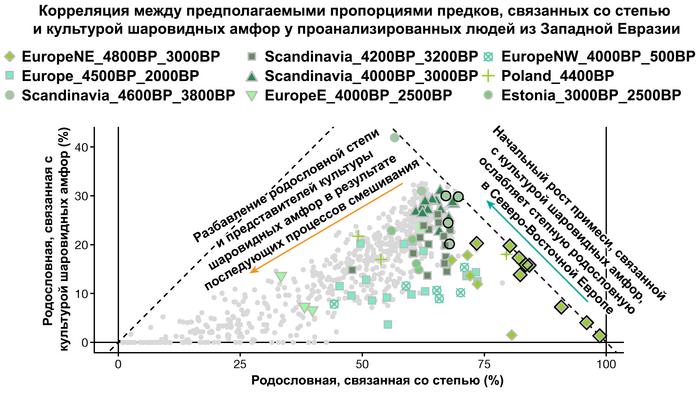
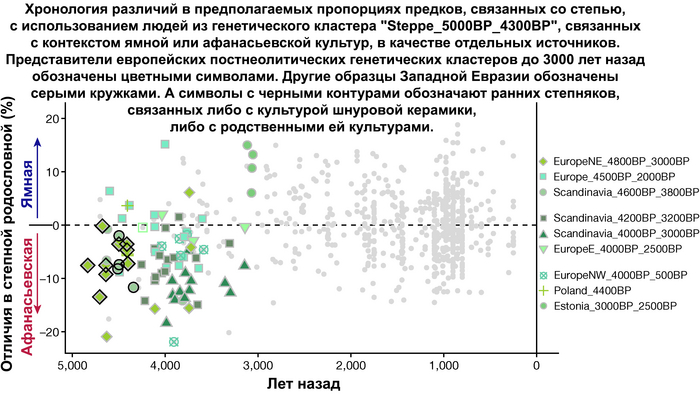
Но несмотря на то, что влияние степной родословной на популяции Евразии хорошо известно, вопросы о её происхождении оставались не решенными. В данной работе авторы показывают, что состав степной родословной (Steppe_5000BP_4300BP) может быть смоделирован как смесь примерно 65% компонент, связанных с охотниками-собирателями, использующими керамику, из района Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет (MiddleDon_7500BP) и 35% компонент, связанных с охотниками и собирателями Кавказа, возрастом от 13 до 10 тыс. лет (Caucasus_13000BP_10000BP).
Таким образом, охотники-собиратели Среднего Дона, которые уже имели родословную, связанную с кавказскими охотниками-собирателями, являются неизвестным до сих пор ближайшим так называемым северным источником большей части родословной у людей, связанных с ямной культурой. При этом генетический состав людей из могильника Голубая Криница, отличается от такового у других древних людей из могильников мариупольского типа Приднепровья. Однако распространение степной родословной в Европе связно с культурой шнуровой керамики, носители которой представляют собой смесь предковых линий, связанных со степью и неолитическими земледельцами, происхождение которых может быть связано исключительно с культурой шаровидных амфор.
Это наблюдение имеет большое значение для понимания возникновения культуры шнуровой керамики. Хотя уже давно высказывались предположения о стилистической связи между керамикой культур шаровидных амфор и шнуровой керамики, включая использование сосудов в форме амфор и узоры, выполненные шнуром. При этом незадолго до появления самых ранних групп культуры шнуровой керамики восточные группы культуры шаровидных амфор и западные группы ямной культуры обменялись культурными элементами в лесостепной переходной зоне к северо-западу от Черного моря, где керамические амфоры и кремневые топоры культуры шаровидных амфор были включены в погребения ямной, а типичное для ямной использование охры встречалось в погребениях культуры шаровидных амфор, что указывает на тесное взаимодействие между этими группами. Предыдущие генетические анализы нескольких людей из зоны контакта предполагали, что эти контакты были ограничены культурным влиянием, а не примесью. Однако, в свете новых генетических данных, похоже, что эта зона — и, возможно, другие подобные зоны контакта между культурой шаровидных амфор и группами из степи (как из ямной культуры) — были ключевыми в формировании культуры шнуровой керамики, благодаря которой генетические компоненты, связанные со степью, и культурой шаровидных амфор, распространились далеко на запад и север, что привело к регионально разнообразным ситуациям взаимодействия и смешения.
При этом распространение культуры шнуровой керамики происходило преимущественно благодаря связям, которые были выстроены культурой шаровидных амфор в предшествующий период. Но несмотря на сходство по аутосомным данным, различные линии гаплогрупп Y-хромосомы у представителей культур шнуровой керамики и ямной предполагают, что известные на данный момент образцы представителей ямной культуры не подходят в качестве непосредственного источника степной родословной у носителей культуры шнуровой керамики.
Это предположение было подтверждено моделированием предковых компонент у европейцев со степным происхождением возрастом от 3 тыс. лет назад с участием представителей родственных ямной и афанасьевской культур, потому как модели показывают, что представители афанасьевской культуры, хоть и незначительно, но оказались более лучшим источником степной родословной у европейцев, несмотря на разное географическое распространение.
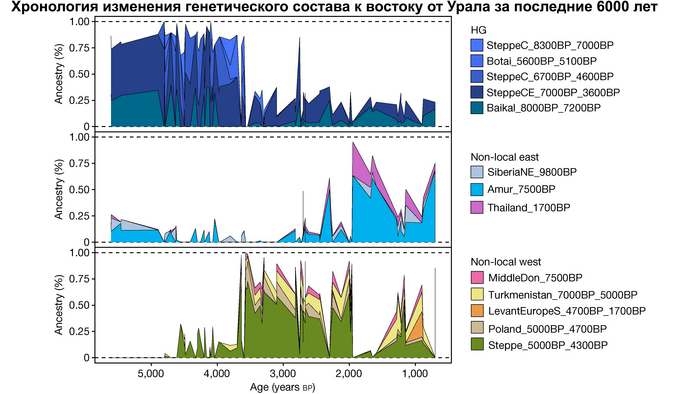
Стабильность популяций охотников-собирателей к востоку от Урала
В отличие от значительного числа геномов древних охотников-собирателей из Западной Евразии, которые были изучены до настоящего времени, геномные данные охотников-собирателей к востоку от Урала были скудны. Эти регионы характеризуются ранним появлением керамики из районов, расположенных дальше к востоку, они были населены сложными обществами охотников-собирателей с постоянными, а иногда и укрепленными поселениями. В данной работе сообщается о 38 новых образцах, 28 из которых от охотников-собирателей, использующих керамику 8300-5000 лет назад. Большинство этих геномов попадает на неолитическую степную клину, которая охватывает сибирские лесостепные зоны бассейнов рек Иртыш, Ишим, Обь, Енисей, а также озера Байкал.
Эти охотники и собиратели могут быть смоделированы как смесь компонент из трёх генетических кластеров:
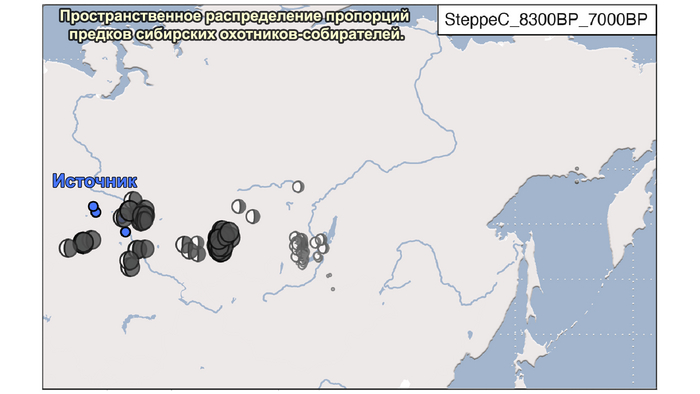
1. Компонента западносибирских охотников-собирателей возрастом от 8300 до 7000 лет (SteppeC_8300BP_7000BP), доминировала в западной части лесостепной зоны.
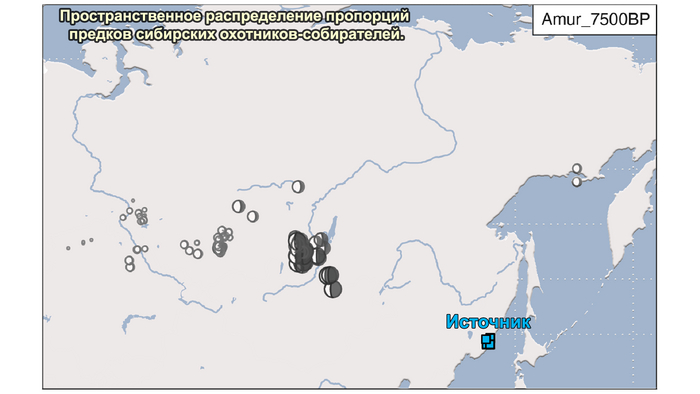
2. Компонента охотников-собирателей Северо-Восточной Азии возрастом около 7500 лет (Amur_7500BP) или древних северо-восточных азиатов из пещеры Чёртовы ворота в Приморье, была максимально выражена у популяций Байкала.
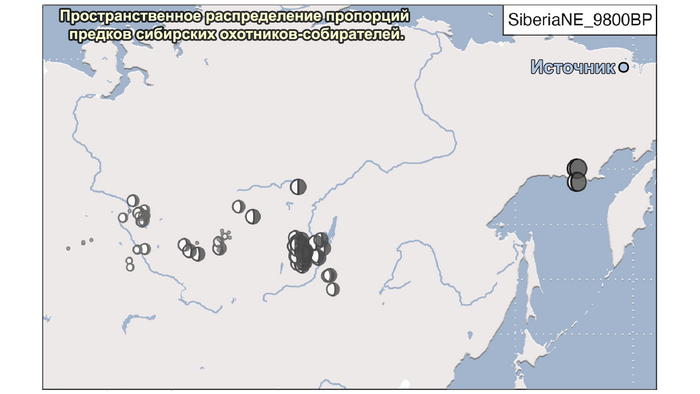
И 3. Компонента представлена палеосибирской родословной возрастом 9800 лет (Siberian_9800BP) как у обитателя Дуванного Яра в низовьях реки Колымы, она снижались по мере удаления от озера Байкал на запад.
1/5
Авторы использовали кластеры также и для исследования пространственно-временной динамики генетического состава в степи и районе озера Байкал после неолита. Результаты подтверждают увеличение компоненты охотников-собирателей лесостепи возрастом от 7000 до 3600 лет (SteppeCE_7000BP_3600BP) у жителей Байкала позднего неолит-ранней бронзы (кластеры Baikal_5600BP_5400BP и Baikal_4800BP_4200BP). Однако родословная, относящаяся к этому кластеру, уже наблюдалась у вновь описанных неолитических охотников-собирателей как у озера Байкал (NEO199 и NEO200), так и вдоль Ангары на севере (NEO843) около 7000 лет назад. У двух мужчин Байкала была определена гаплогруппа Y-хромосомы Q1b1, характерная для последующих групп позднего неолита-ранней бронзы из того же региона. Что совместно с расчётной датой примеси около 7300 лет назад, позволяет предположить, что поток генов между охотниками-собирателями Байкала и лесостепей Южной Сибири уже был во времена раннего неолита, что согласуется и с археологическими данными.
Новые образцы также проливают свет на генетическое происхождение носителей окуневской культуры раннего бронзового века в Минусинской котловине на юге Сибири. Её представителей можно смоделировать как смесь сибирских охотников-собирателей лесостепной зоны из кластеров возрастом 6700-4600 и 7000-3600 лет (Steppe_6700BP_4600BP и SteppeCE_7000BP_3600BP) со степными скотоводами возрастом от 5000 до 4300 лет (Steppe_5000BP_4300BP), связанными исключительно с афанасьевской культурой, время этой примеси датировано около 4600 лет назад.
1/2
А примерно 3700 лет назад жители в степном регионе и у озера Байкал демонстрируют уже заметно отличающиеся генетические профили. Ранние стадии этого перехода характеризуются притоком предков, связанных со степью, количество которых снижается со временем, от 70% на начальном этапе.
Аналогично динамике в Западной Евразии, происхождение, связанное со степью, здесь соотносится с таковым как у представителей культуры шаровидных амфор с территории современной Польши, возрастом 5000-4700 лет, подтверждая ранее задокументированный поток генов от групп из культуры шаровидных амфор в соседние группы степи и лесостепи, а также поток этой уже смешанной с европейскими земледельцами родословной дальше на восток уже с синташтинской и андроновской культурами в бронзовом веке. Однако у людей из окуневской культуры отсутствует вклад от людей культуры шаровидных амфор, а люди со степным происхождением начиная от 3700 лет назад демонстрируют чуть большую близость к ямной, чем к афанасьевской культуре, что указывает на две волны миграций западных степных скотоводов на восток. Первая волна в раннем бронзовом веке была связана с ямной культурой, а вторая, позже, была связана с синташтинской и андроновской культурами. Более поздние этапы переходного периода характеризуются увеличением родословной, связанной с иранскими земледельцами Центральной Азией (Turkmenistan_7000BP_5000BP) и древними северо-восточными азиатами Приморья (Amur_7500BP).
Суммарно, эти результаты показывают, что на востоке Евразийской степи родословная охотников и собирателей со сложной генетической структурой сохранялась и преобладала значительно дольше, чем в западной Евразии. Пока менее чем 4 тыс. лет назад по степи не прокатились волны миграций, которые привнесли в регион одомашненные породы лошадей с последующим появлением нового снаряжения для верховой езды и колесницы со спицами. Также эти миграции способствовали распространению проса в качестве сельскохозяйственной культуры.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
Такую задачу поставил Little.Bit пикабушникам. И на его призыв откликнулись PILOTMISHA, MorGott и Lei Radna. Поэтому теперь вы знаете, как сделать игру, скрафтить косплей, написать историю и посадить самолет. А если еще не знаете, то смотрите и учитесь.
одна столовая ложка вещества весит порядка 100 миллионов тонн.
Не могу на это смотреть без боли. Для такой массы радиус Шварцшильда (горизонта событий) будет составлять 1.485×10^-16 метров. Какая столовая ложка? С такой плотностью данный магнетар не мог бы существовать, а сколлапсировал бы давным-давно в черную дыру.