Популяционная генетика послеледниковой Евразии.
2 поста
Сегодня речь пойдёт о поморах – представителях самых северных русских популяций Беломорья. Считается, что это потомки новгородцев, расселявшихся с XII в по берегу Белого моря от Онеги до Кеми, а также по берегу Баренцева моря. Поморы адаптировали свой быт и культуру к суровым природным условиям северных морей, занимаясь рыболовством, морской охотой, предпринимательством и мореходством на Белом море, в Северном Ледовитом океане и на его островах, конкурируя с норвежцами.

И хотя поморы отличались по быту от северных русских, они всё же сохранили черты древнерусской культуры, при этом взаимодействуя с населением Северной и Северо-Восточной Европы. Поэтому население этого региона крайне своеобразно, что и заинтересовало учёных наряду с нюансами эволюции популяций на самой периферии этнического ареала.

Заселение Беломорья началось около 8 тысяч лет назад ещё в мезолите. В неолите свои следы оставили представители «каргопольской» и «беломорской» культур.
А в бронзовом веке, около 3-4 тыс. лет назад, в регион прибыли мигранты, которых связывают с финно-угорскими популяциями (прежде всего, с группами, говорившими на языках, близких к саамским). Последней была славянская волна мигрантов. И первые упоминания о постоянных русских поселениях на беломорском побережье в конце XIII — начале XIV века связаны с Новгородской колонизацией северных земель. Где, согласно летописям, переселенцы встретили племена "заволочской чуди", как в летописях называли финноязычное население, часто ассоциирующееся с современными вепсами. Проживание на самом севере ареала русского народа, где в топонимике прочно сохранились следы дославянского населения, совместно с уникальной культурой и бытом, предполагает также и разнообразие генофонда поморов.
В предыдущих работах — по крайне узким панелям маркеров Y-хромосомы и мтДНК с небольшими выборками — поморы рассматривались в составе популяции «северных русских», а особенности их генофонда объяснены возможной ассимиляцией урало- или балтоязычного населения славянским. Данные об аутосомном геноме поморов Мезенского района Архангельской области также указывают на сходство с финноязычным населением, причем больше с финно-пермским, чем с финно-волжским.
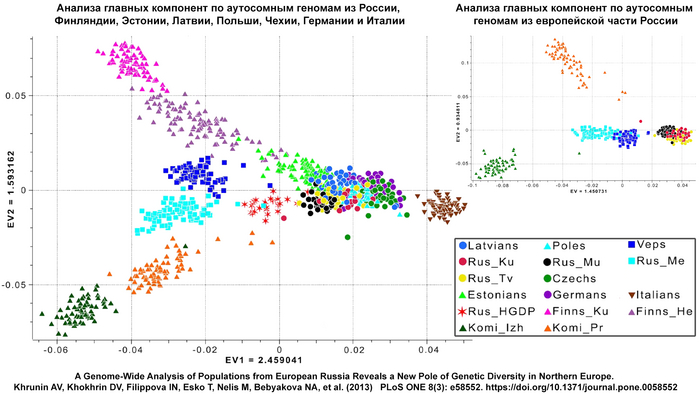
На рисунке: данные об аутосомном геноме поморов Мезенского района Архангельской области
Однако в исследованиях не поморов, а обширного круга популяций Русского Севера, выявлена их близость по мтДНК и аутосомным ДНК-маркерам к населению Северной Европы.
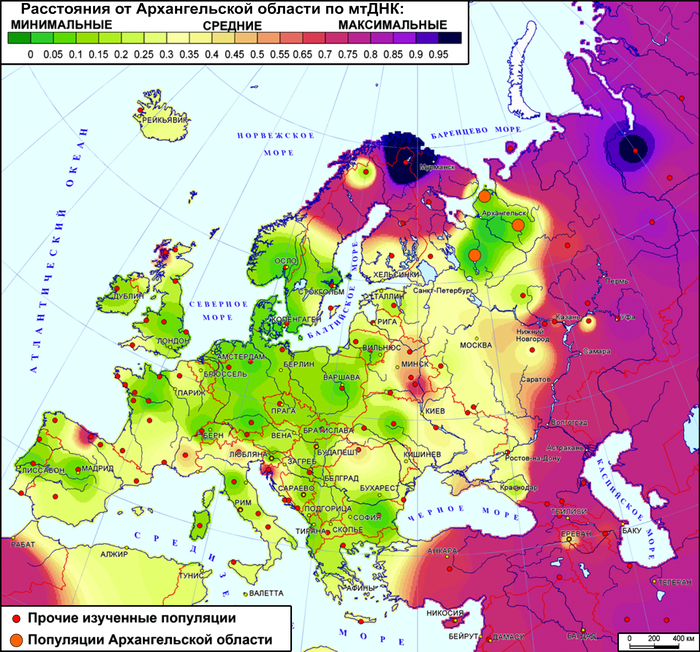
А разнообразие линий Y-хромосомы обнаружило сходство генофонда жителей Русского Севера с населением от Прибалтики до Печоры и объяснялось наследием древнейших палеоевропейских популяций.
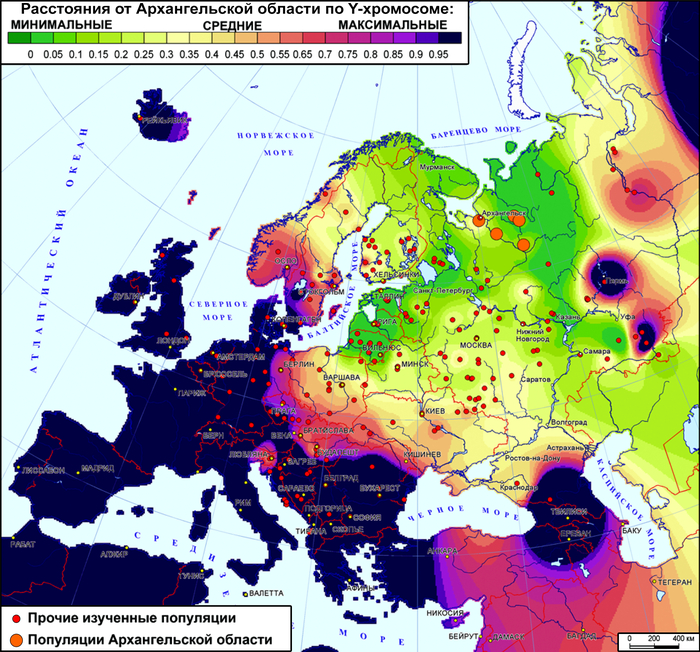
Интригу к вопросу о происхождении поморов добавляет и тот факт, что у жителей самого севера Архангельской области не обнаружена «новгородская» предковая компонента по широкогеномной панели аутосомного генома.
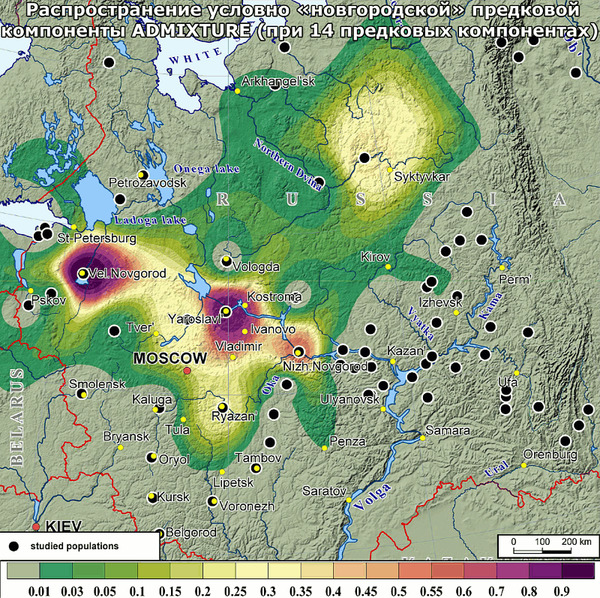
В новой работе детально изучены наиболее сохранившиеся и труднодоступные популяции поморов Белого моря. В ходе экспедиции 2021 г. обследованы практически все населенные пункты Онежского полуострова (Онежский и Летний берега), а также западного (Прионежского) фрагмента Зимнего берега. И только для краткости все три популяции названы «онежскими поморами».
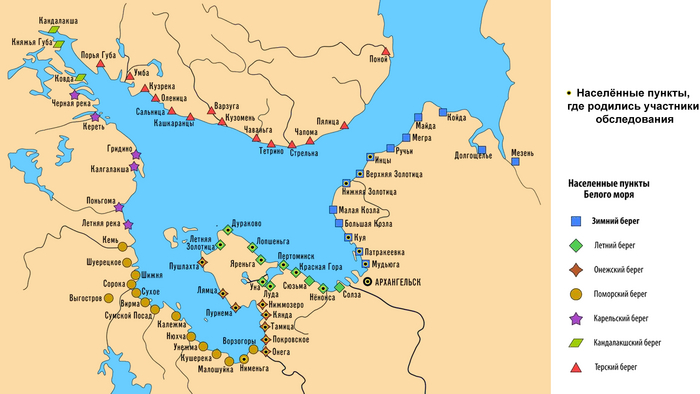
Авторы надеялись, что столь трудно добытые новые данные о генофонде поморов, изученном по современным панелям маркеров, смогут дать ключ к пониманию значительного генетического разнообразия и своеобразия популяций Русского Севера. Принимая во внимание традицию патрилокальности у поморов, авторы работы изучили маркеры Y-хромосомы для того, чтобы создать «генетические портреты» трёх впервые исследованных популяций поморов и попробовать найти в их генофонде генетическую память о новгородской колонизации.
Разнообразие гаплогрупп Y-хромосомы у поморов
У онежских поморов самыми частыми оказались три гаплогруппы: 26% R1a (три ветви); 25% I1 (одна ветвь), 24% N3a (две ветви). А всего у исследованных мужчин было выявлено 14 линий Y-хромосомы: I1-M253, N3a4-Z1936, R1a-CTS1211, R1a-PF6202, R1b-L51, I2-P37.2, R1a-Z92, N3a3-CST10760, E-M78 и T1a-M70, встречающиеся с частотами от 25 до 1% (перечислены по убыванию) и линии I2-M223, J2-M92, J2-M67, N2a-Y3205 с частотами менее 1%.
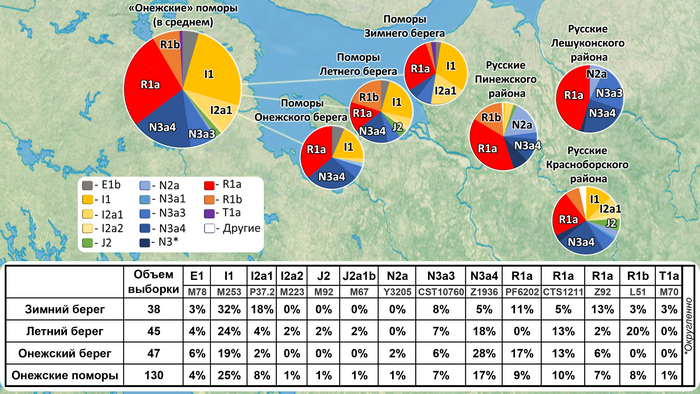
При этом, несмотря на небольшое расстояние между тремя популяциями поморов (от 80 до 170 км,) заметно отличаются частоты их гаплогрупп:
– у поморов Зимнего берега снижена частота гаплогруппы N3a4 и повышена частота I2;
– у жителей Онежского берега наоборот, снижена частота гаплогруппы I2 и повышена частота N3a4.
– а у находящихся между ними представителей Летнего берега не обнаружена гаплогруппа R1a-PF6202, характерная для двух других популяций, но повышена частота гаплогруппы R1b.
Примечательно, что у поморов Летнего берега встречается линия R1b-L51, более характерная для народов Северо-Западной Европы, в отличие от другой линий R1b, обнаруженной у русских из Пинежского района Архангельской области.
В зависимости от географического положения популяций поморов наблюдается снижение частот гаплогрупп N3a4 и E1 с юго-запада на северо-восток, а частоты гаплогрупп I1 и I2a наоборот возрастают в этом направлении. Однако этой тенденции нет у ветвей гаплогруппы R1a, частота которой высока в генофондах Зимнего (29%) и Онежского берегов (36%), но в два раза ниже на расположенном между ними Летнем берегу (причем только ветви R1a-CTS1211 (13%) и R1a-Z92 (2%)). Зато на Летнем берегу наблюдается аномально высокая частота R1b-L51 (20%), что может быть результатом миграций и дрейфа генов.
Сходство онежских поморов с другими русскими популяциями Архангельской области наблюдается по частотам гаплогрупп R1a и N3a4 (кроме Зимнего берега, где N3a4 редка). А различия наиболее выражены по частоте гаплогруппы I1: у онежских поморов она составляет в среднем четверть генофонда (25%), но для других северных русских не характерна (в Красноборском районе Архангельской области – 12%, в Пинежском – 1%, а в Лешуконском не обнаружена вовсе).
При оценке сходства разных генофондов суммарно по всем гаплогруппам наиболее примечательны значительные генетические расстояния между тремя популяциями онежских поморов, вопреки их географической близости.
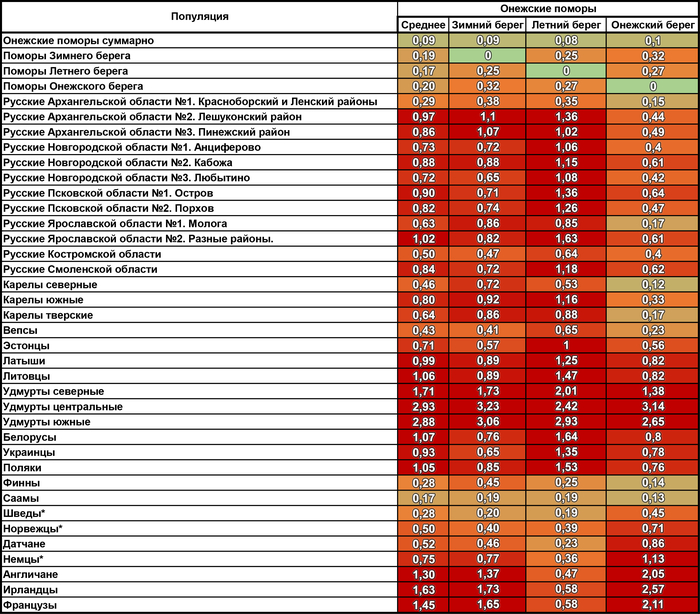
Но ещё более значительны расстояния от поморов до других русских популяций. Есть лишь два исключения. Во-первых, сходство поморов Онежского берега с географически далекими русскими Красноборского района Архангельской области. Во-вторых, их сходство с русскими из района Мологи Ярославской области. Причём ранее было показано, что среди ярославских популяций именно представители Мологи сохранили явный генетический след дославянского населения (предположительно мери). Однако поморы Онежского берега ещё ближе к северным карелам, а также финнам и саамам, чем к этим русским популяциям. Поморы Летнего берега также близки к саамам, но ещё они близки и к шведам с датчанами, причём до них генетические расстояния меньше, чем до коллег по быту. Похожая картина у поморов Зимнего берега, только они более близки к саамам и шведам по генетическим расстояниям.
В целом онежские поморы демонстрируют наибольшее генетическое сходство с народами Северной Европы.
Причём расстояние до генофондов финнов и саамов уменьшается по мере движения на юго-запад - от Зимнего берега к Онежскому в 4 раза, а генетическое расстояние до скандинавов уменьшается к Летнему берегу от Зимнего и Онежского берегов. При этом поморы Летнего берега помимо близости к датчанам, норвежцам и шведам, также близки и к немцам с англичанами, хоть и не настолько, как к скандинавам. Также представители Летнего берега генетически ближе к ирландцам и французам, чем к соседним русским популяциям.
Среди других народов России к онежским поморам наиболее генетически близки вепсы и северные карелы, причем в основном за счет поморов Онежского берега, которые сами ближе к финноязычным соседям, чем к другим популяциям поморов.
А наиболее генетически удалёнными от поморов оказались удмурты – представители восточного крыла финноязычных народов, что входит в противоречие с выводами предыдущих публикаций о сходстве геномов финно-пермских народов с поморами.
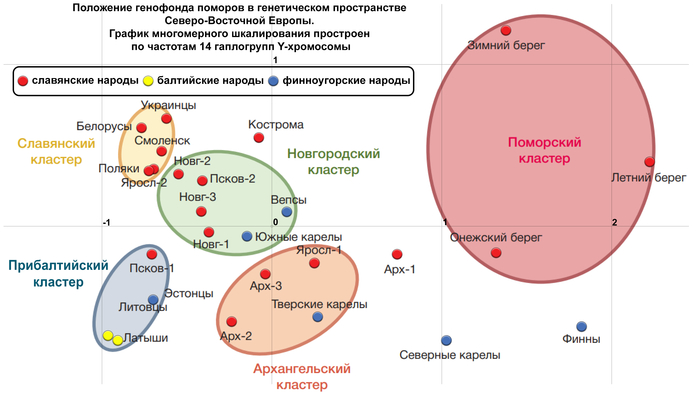
На графике многомерного шкалирования (построенном по 14 «поморским» гаплогруппам Y-хромосомы) выделяются 5 кластеров, с условными названиями:
1. «Славянский» кластер, в него вошли белорусы, украинцы, поляки, а также русские Смоленской и Ярославской областей.
2. «Новгородский» кластер, близкий к «Славянскому», объединил все три популяции Новгородской области и Порхов Псковской области, который входил в Новгородские земли. К ним присоединились также финноязычные популяции – вепсы и южные карелы.
3. «Прибалтийский» кластер, помимо латышей, литовцев и эстонцев, включил население Островского района Псковской области.
4. «Архангельский» кластер объединил популяции Пинежского и Лешуконского районов Архангельской области с Мологой Ярославской области и тверскими карелами.
5. «Поморский» кластер, из-за значительных расстояний между популяциями поморов, оказался лишь немногим меньше суммы расстояний предыдущих четырех кластеров, включивших финноязычные, балтоязычные и славянские популяции.
При этом авторы подчёркивают, что вопреки значительным различиям между поморскими популяциями, все вместе они занимают своё собственное «поморское» место среди популяций Северо-Восточной Европы.
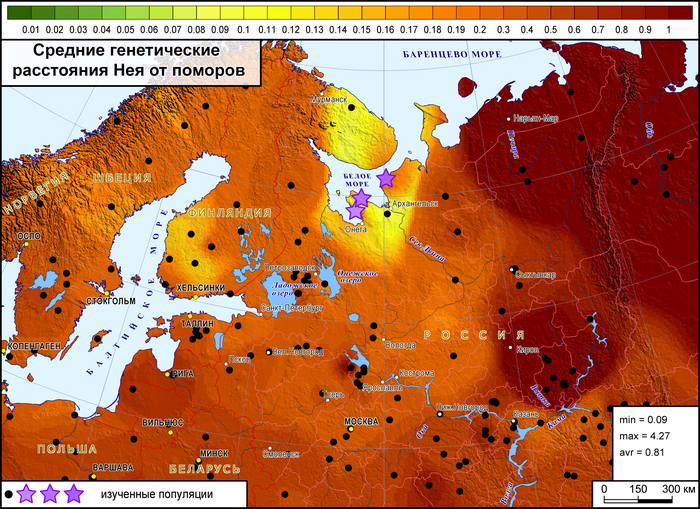
Более точно определить регионы, с которыми сходны генофонды поморов, и значительно расширить круг популяций сравнения помогают карты генетических расстояний по методу Масатоши Нея, рассчитанные по 26 гаплогруппам Y-хромосомы, характерным для всего региона.
Суммарный генофонд поморов помимо некоторых соседних популяций генетически более близок к южной части Финляндии, а не к северной, представленной саамами.
Генофонд поморов Онежского берега на карте генетических расстояний демонстрирует близость к популяциям на обширных географических участках, охватывая почти всю Финляндию на западе, кроме крайних северных и южных участков, а также от рек Сухона и Северная Двина на востоке России до Ярославской и Ленинградской областей на юге и юго-западе.
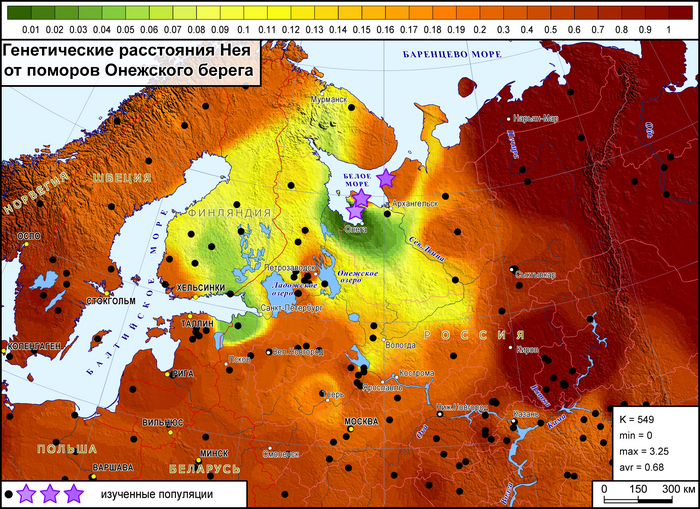
Эта область сходств охватывает финноязычные народы и те русские популяции, в генофонде которых прослеживается существенный вклад дославянского населения.
Генофонд поморов Летнего берега демонстрирует наибольшее сходство со шведами и норвежцами, а также с саамами, но слабее.
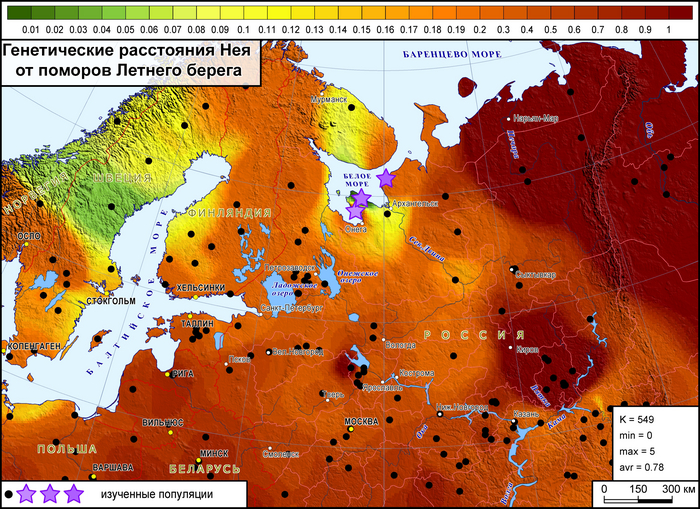
Генофонд поморов Зимнего берега, представленный «прионежской» его частью, показывает лишь некоторое сходство с отдельными популяциями финнов и шведов.
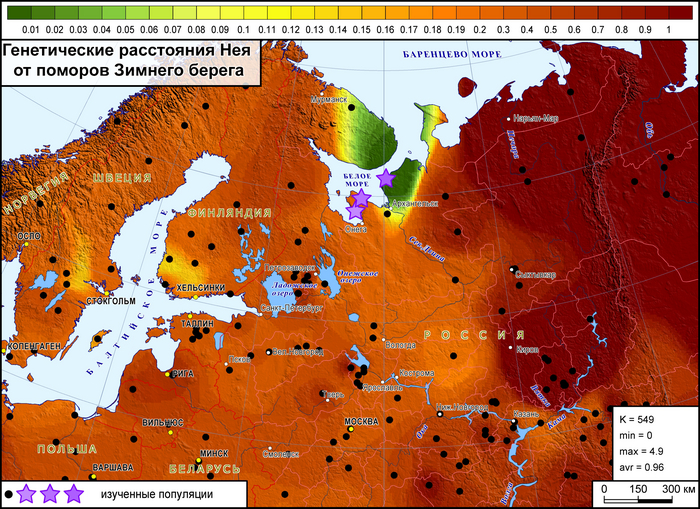
Это единственная популяция поморов, для которой можно допустить, что ее генетический портрет в значительной степени сформирован дрейфом генов, однако исследование популяций остальной части Зимнего берега ещё проводится.
Обсуждение и выводы
Учитывая схожий быт и культуру, а также географическую близость трёх исследованных популяций поморов разумно было бы предположить, что и их генофонды будут довольно схожи. Однако эти ожидания не подтвердились, и каждая из трех исследованных поморских популяций продемонстрировала свои уникальные отличия.
Поморы Онежского берега оказались близки к финноязычным народам России и Финляндии, а также к русским популяциям Архангельской области. Поморы Летнего берега продемонстрировали генетическое сходство только с населением Скандинавии. А у поморов Зимнего берега практически нет сходных генофондов за исключением некоторой близости к финнам и шведам.
Различия между популяциями поморов почти столь же велики, как между рассмотренными популяциями, говорящими на разных языках.
Связать такое уникальное своеобразие генофондов поморов с действием лишь дрейфа генов невозможно, хотя не оспаривается, что дрейф генов был важным фактором в генетической истории всех популяций поморов, численность которых в последнее время сокращалась.
Полногеномные исследования в будущем позволят пролить свет на происхождение поморов и на миграционные волны, оставившие след в их генофонде.
Кстати, обращаю Ваше внимание на необходимость дальнейших исследований, чтобы удержаться от поспешных выводов только по однородительским маркерам.
Но и результаты анализа отцовских линий, учитывая патрилокальность поморов прямо указывают, что причина генетического своеобразия популяций онежских поморов заключается в различном генетическом происхождении их отличающихся субстратов, с последующим дрейфом генов, который размывал генетическую память.
Ещё один важный вопрос заключался в поиске сходства генофондов поморов и новгородцев. Но среднее генетическое расстояние между этими популяциями оказалось довольно значительным, почти как между поморами и другими рассмотренными русскими популяциями. Примечательно, что в предыдущей работе авторы сообщали о различии аутосомных геномов новгородцев и русских севера Архангельской области. И в новой работе показаны различия между генофондами новгородцев и поморов по маркерам Y-хромосомы. Оба результата противоречат мнению о том, что генофонд Русского Севера был сформирован за счёт Новгородской колонизацией северных земель.
Здесь важно отметить, что генетические данные не перечёркивают связи поморов с новгородцами - просто культура и генетика порой отражают разные плоскости взаимодействий между разными группами населения. И генетическая память отражает лишь те события в истории народов, которые сопровождались демографическими изменениями.
Источник:
Окованцев, В. С., Пономарев, Г. Ю., Агджоян, А. Т., Агджоян, А. Т., Пылёв, В. Ю., Балановская, Е. В. Своеобразие поморов Онежского полуострова и Зимнего берега в генетическом контексте севера Европы // Вестник РГМУ. 2022. №5. С. –. DOI: 10.24075/vrgmu.2022.046
Дополнительные источники:
1. Генофонд Русского Севера: Славяне? Финны? Палеоевропейцы? Балановская, Д.В. Пежемский, А.Г. Романов, Е.Е. Баранова, М.В. Ромашкина, А.Т Агджоян, А.Г. Балаганский, И.В. Евсеева, Р. Виллемс, О.П. Балановский
2. Two Sources of the Russian Patrilineal Heritage in Their Eurasian Context Oleg Balanovsky, * Siiri Rootsi, Andrey Pshenichnov, Toomas Kivisild, Michail Churnosov, Irina Evseeva, Elvira Pocheshkhova, Margarita Boldyreva, Nikolay Yankovsky, Elena Balanovska, and Richard Villems DOI 10.1016/j.ajhg.2007.09.019. 2008 by The American Society of Human Genetics.
3. Своеобразие Новгородского генофонда в контексте народонаселения европейской части России. Е.В. Балановская, Д.К. Черневский, О.П. Балановский doi.org/10.34680/2076-8052.2021.3(124).51-57
4. The Eurasian Heartland: A continental perspective on Y-chromosome diversity R. Spencer Wells, Nadira Yuldasheva, Ruslan Ruzibakiev, Walter F. Bodmer et. all doi.org/10.1073/pnas.171305098
5. Khrunin AV, Khokhrin DV, Filippova IN, Esko T, Nelis M, Bebyakova NA, et al. (2013) A Genome-Wide Analysis of Populations from European Russia Reveals a New Pole of Genetic Diversity in Northern Europe. PLoS ONE 8(3): e58552. doi.org/10.1371/journal.pone.0058552
Введение
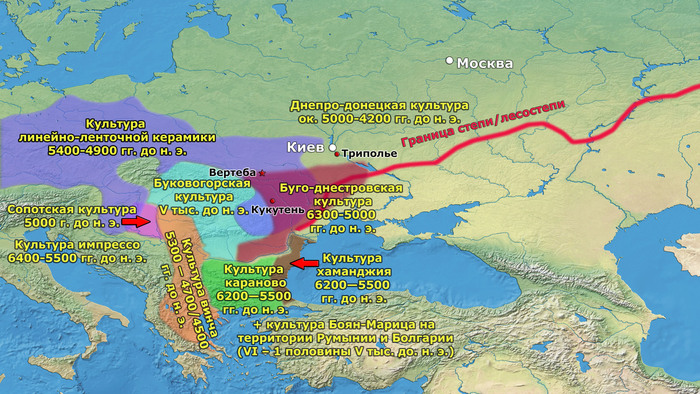
Процесс неолитизации в Европе привел к серьёзным технологическим и культурным сдвигам, которые включали в себя новые методы ведения хозяйства. Существуют две основные группы моделей, которые объясняют процесс неолитизации. Модели демической диффузии описывают неолитизацию как процесс колонизации земледельцами, который был движим экспоненциальным ростом населения, характерным для неолита. Вторая группа моделей – аккультурации – описывает процесс, в котором, по крайней мере, переход к земледелию мог практиковаться местными охотниками-собирателями после периода (той или иной продолжительности) взаимодействия с соседними, пришлыми земледельцами.
Анатолийские земледельцы достигли Балкан и других регионов Юго-Восточной Европы в седьмом тысячелетии до нашей эры и впоследствии распространились дальше через Средиземное море, а затем через Дунайский коридор, существенно вытеснив коренное мезолитическое европейское население.
В отличие от Центральной Европы, области Восточной Европы, включая Украину, Молдову, Западную Россию и Румынию, не занимались сельским хозяйством до позднего неолита (~ 4500 г. до н. э.), хотя различные оседлые и полуоседлые группы охотников-собирателей этих регионов начали использовать глиняную посуду еще в 8500 г. до н. э.
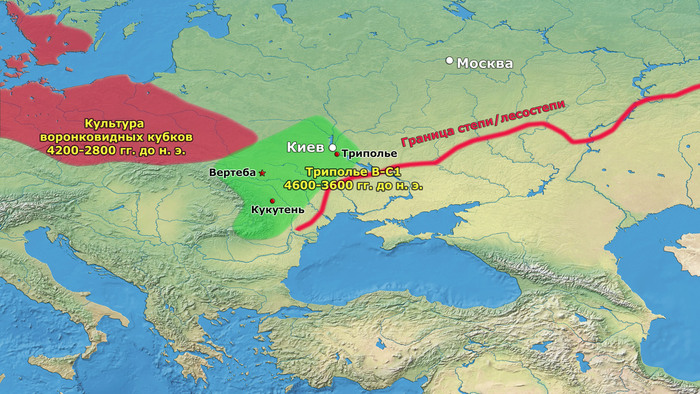
Археологическая общность Триполье-Кукутень представляет собой группу нескольких взаимосвязанных археологических культур среднего неолита/энеолита на территории ряда областей современных Украины, Молдовы и Румынии. Этот комплекс простирается от Южных Карпат до Днепра и назван в честь типовых памятников Кукутень в Яссы (Румыния) и Триполье (Обуховский район Киевской области).
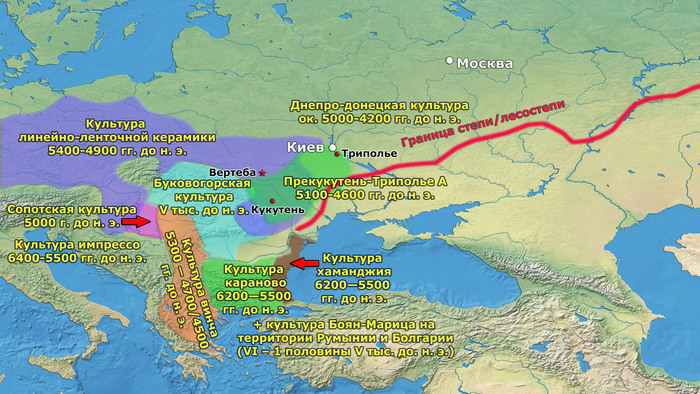
Культуры Кукутень и Триполье имеют общие корни в культуре Прекукутень; самые ранние стоянки археологической общности Триполье-Кукутень обнаружены в предгорьях Карпат, а самые ранние радиоуглеродные даты (от периода Прекукутень II) относятся примерно к 4800 г. до н.э. Археологическая общность Триполье-Кукутень возникла в результате взаимодействия нескольких дунайских неолитических групп, о чем свидетельствует сходство в строительстве домов, керамических стилях и производстве каменных артефактов.

Rassamakin, Y., & Menotti, F. (2011). Chronological Development of the Tripolye Culture Giant-Settlement of Talianki (Ukraine): 14C Dating vs. Pottery Typology. Radiocarbon, 53(4), 645-657. doi:10.1017/S0033822200039102
После возникновения этого культурного комплекса в Карпатском предгорье археологическая общность Триполье-Кукутень расширяла свой ареал и в конечном итоге заняла территорию, охватывающую большую часть современных территорий Украины, Молдовы и Румынии. Первые раннетрипольские (Триполье А) стоянки отошли от культуры Прекукутень ~ 4500 г. до н. э. в долине реки Днестр.
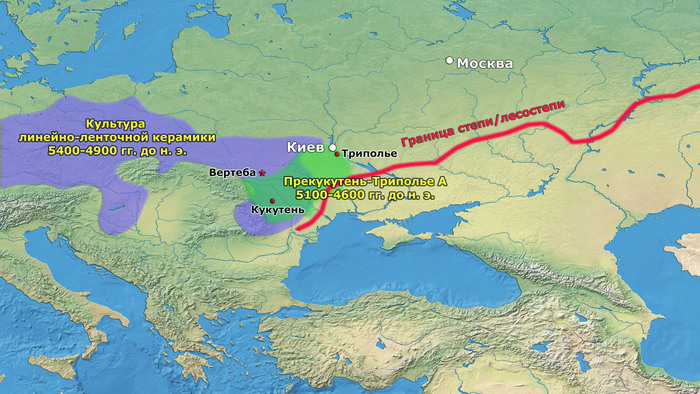
Более поздние перемещения населения, происходящие со среднего периода (Триполье BI) и далее, привели к тому, что трипольская культура распространилась на Волынь на западе и Днепр на востоке.
Считается, что эта территориальная экспансия была вызвана в первую очередь демографическим ростом, связанным с успешной стратегией земледелия и животноводства, с поиском новых пахотных земель для возделывания. Однако некоторый рост населения мог быть результатом включения в состав трипольцев местных групп охотников-собирателей, например, представителей буго-днестровской культуры.
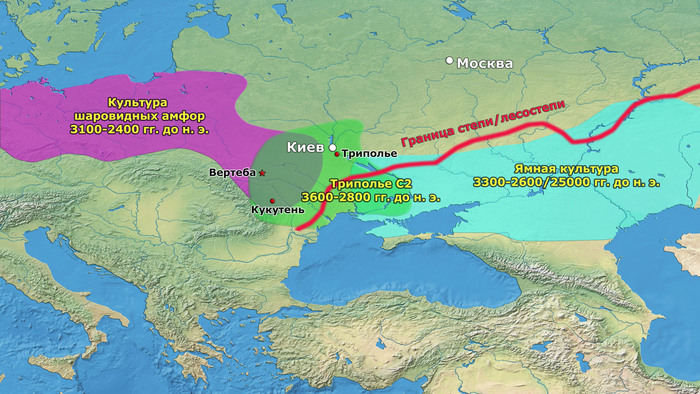
Другим каналом увеличения населения могла быть аккультурация переселенцев после краха неолита в Румынии, Венгрии и Болгарии. В средние и поздние периоды трипольской культуры (Триполье от BII до CI; 4100–3400 гг. до н. э.) некоторые группы трипольской культуры основали чрезвычайно крупные поселения в Центральной Украине, которые достигли размеров 100–320 га.

Быстрый демографический рост трипольцев на рубеже 4 тысячелетия до н. э. потребовал освоения новых территорий, ускорив миграцию в ранее периферийные районы.
Гипотезы возникновения мегапоселений разнообразны. Они могли быть защитным ответом на угрозы, исходящие от степных скотоводов или же от конкурирующих популяций внутри самой археологической общности Триполье-Кукутень. Они могли и просто быть временным эпизодом компактного скопления поселений из-за крупномасштабной миграции из Приднестровья.
Однако, несмотря на высокую плотность поселений в Западной и Центральной Украине, здесь обнаружено крайне мало захоронений. Лишь несколько кладбищ позднетрипольского периода были раскопаны в 1960-х и 1970-х годах, например, Чапаевка на территории Украины и Выхватинцы в Приднестровье. Хотя эти памятники дают некоторое представление о погребальных обрядах трипольцев, они ограничены по временному охвату и не подвергались современным лабораторным исследованиям.
Пример по могильнику в Выхватинцах:Чтобы лучше понять происхождение, связи и разнообразие населения археологической общности Триполье-Кукутень, авторы исследовали палеоантропологические образцы из трех камер на участке пещеры Вертеба в Тернопольской области Украины - одном из немногих мест, где они обнаружены.
Положение скелетов в могилах различно — наряду с преобладающими погребениями в скорченном положении на левом или на правом боку отмечено также несколько случаев погребений на спине, с согнутыми и поднятыми в коленях ногами. Ориентировка довольно устойчива — наибольшее число погребенных положено головой на северо-восток, с небольшим отклонением к востоку. В погребальном инвентаре численно преобладает керамика, представленная двумя типами: сосудами из розовой глиняной массы, украшенными черной и красной росписью; и нерасписными сосудами слабого обжига, орнаментированными веревочными оттисками.
Формы и стиль росписи керамики — типичные для позднего Триполья. Число сосудов в погребениях различно — от 1 до 8, чаще — 2-5. Помимо керамики, в мужских погребениях обнаружены роговые мотыги, кремневые вкладыши для серпов, костяные шила, а также каменный топор, кремневый нож, медное шило, глиняная и костяная антропоморфные статуэтки. Женские погребения сопровождались в основном сосудами, лишь в одном были найдены два пряслица и раковина. Выделяется своеобразием инвентарь детских погребений. Кроме сосудов, здесь были найдены антропоморфные глиняные погремушки; антропоморфные женские статуэтки были положены также главным образом в детские могилы. Помимо этого, обнаружены пряслица, бусы из раковин, кремневые отщепы, пластины, костяное шило.
М. С. Великанова «Палеоантропология Прутскоднестровского междуречья» (Издательство «Наука», 1975 г)
Результаты исследования методами масс-спектрометрии углерода-14 из останков людей и фауны указывают на присутствие трипольцев в пещере Вертеба в интервале между 3950 и 3520 гг. до н. э.
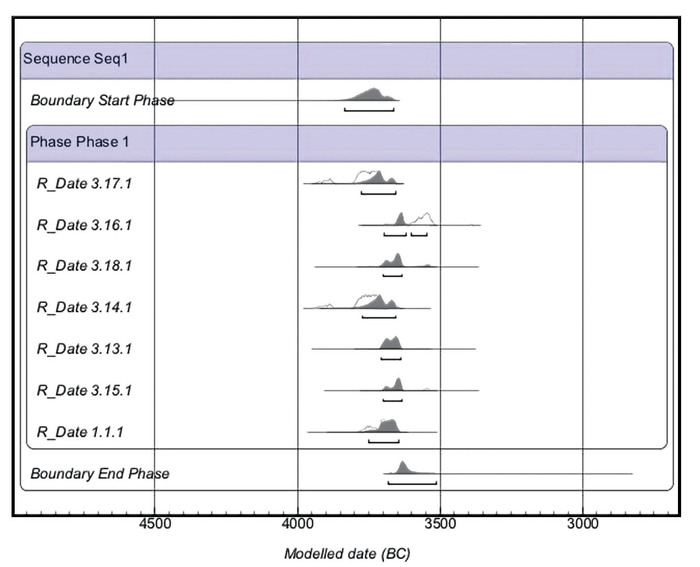
Lillie, M., Budd, C. E., Potekhina, I., Price, T. D., Sokhatsky, M., & Nikitin, A. G. (2017). First isotope analysis and new radiocarbon dating of Trypillia (Tripolye) farmers from Verteba Cave, Biche Zolote, Ukraine. Documenta Praehistorica, 44, 306–324. doi.org/10.4312p.44.18По данным керамических комплексов, присутствующих в пещере, и результатам жидкостно-сцинтилляционного метода анализа углерода-14 с более низким разрешением можно предположить, что заселение продолжалось в течение некоторого времени в период позднего Триполья (CII) и перехода к раннему бронзовому веку.
Помимо этого, масс-спектрометрия также выявила отложения в разных местах пещеры, относящиеся к мезолиту (7950–7490 гг. до н. э.), бронзовому веку, железному веку и средневековью.
Останки людей были извлечены из трех отдельных камер пещеры: участок 7, участок 17 и участок 20. Каждая из этих камер содержит материальную культуру, соответствующую археологической общности Триполье-Кукутень.

Однако захоронения в пещере носят вторичный характер, а нарушения, вызванные деятельностью человека в древности и процессом перемешивания останков животными или растениями, усложняют реконструкцию использования пещеры и хронологии. Большинство изученных образцов в этом исследовании происходят из участка 7, который был подробно задокументирован с помощью анализа керамики и радиоуглеродного датирования, причем пик использования этого участка древним населением датируется периодами CI и CII трипольской периодизации (~ 3900-3350 гг. до н. э.).

Фото: Педагог Светлана - CC BY-SA 4.0, commons.wikimedia.org/w/index.php?curid=92007799Интерпретации использования пещеры разнообразны: в качестве временного убежища, места проведения ритуалов или же погребения. Имеются дополнительные сведения, подтверждающие гипотезу о жертвах конфликтов или же человеческих жертвоприношений.
Палеогенетика трипольской популяции до сих пор включила анализ однородительских маркеров мтДНК и полногеномный анализ 8 представителей трипольской культуры.
Митохондриальные гаплогруппы, типичные для древних евразийских земледельческих групп (H, HV, T, K, J), выявлены у образцов из всех трех камер пещеры. В одной из камер обнаружена гаплогруппа W, характерная для степных популяций, связанных с культурами шнуровой керамики и унетицкой культуры Среднего Поволжья.
Полногеномный анализ представителей археологической общности Триполье-Кукутень показал, что предковые генетические компоненты преимущественно (60–80%) относятся к ранненеолитическим земледельческим группам, подтверждая, что ранние земледельцы, заселившие Западную и Центральную Украину, в значительной степени происходили из той же исходной популяции, что и земледельцы Анатолии и Западной Европы. Оставшиеся 40-20% предковых компонент определены с меньшей достоверностью.
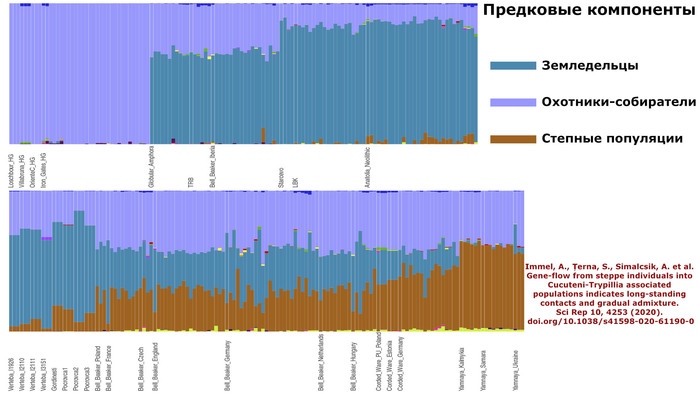
Immel, A., Țerna, S., Simalcsik, A. et al. Gene-flow from steppe individuals into Cucuteni-Trypillia associated populations indicates long-standing contacts and gradual admixture. Sci Rep 10, 4253 (2020). doi.org/10.1038/s41598-020-61190-0Авторы работы 2018 г. [Mathieson et al.] показали, что этот наследственный генетический компонент присутствовал у западных и восточных охотников-собирателей, населявших этот регион в эпоху неолита.
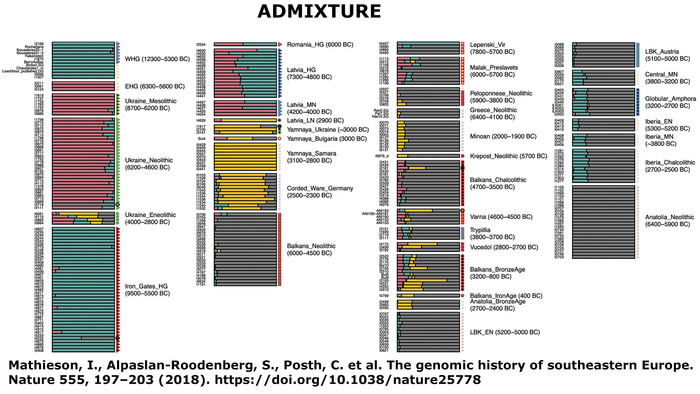
Mathieson, I., Alpaslan-Roodenberg, S., Posth, C. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018). doi.org/10.1038/nature25778Полногеномные данные четырех человек, происходящих из двух разных памятников на севере Молдовы (поздний трипольский период, 3500–3100 гг. до н. э., то есть на пять столетий моложе пещеры Вертеба), выявили у них значительный вклад степного населения (хотя и в разных пропорциях). Это может быть объяснено постепенной ассимиляцией местных мезолитических и неолитических групп охотников и собирателей трипольским населением, жившем на территории современной Молдовы и Приднестровья.
Ареал трипольцев расположен таким образом, что они контактировали как с центральноевропейскими, так и со степными популяциями. Археологические свидетельства взаимодействия с населением степей обнаруживаются в обожженной глиняной посуде. Некоторые из них выглядят почти идентично керамике, найденной в степях, в то время как другая керамика сочетает обжиг с декоративными мотивами трипольцев. Обнаружены и символические предметы, находящиеся под влиянием или непосредственно импортированные из степных сообществ. Например, каменные булавы обнаружены на некоторых стоянках среднего и позднего Триполья, а обмен глиняной посудой очевиден уже в Триполье BII.
Несомненно, существовала определенная степень взаимодействия между трипольским населением и днепро-донецкой культурой, в то время как взаимодействие трипольской культуры и последующей ямной, вероятно, было очень кратким.
Тем не менее, некоторые трипольские популяции, вероятно, находились в постоянном контакте со степными популяциями. После 3400 г. до н. э. трипольские мегаполисы были в значительной степени заброшены. Причина этого широко обсуждалась. Одна из гипотез - усиление конфликтов из-за расширения степных культур на запад. Такая гипотеза может найти подтверждение в частых свидетельствах насильственной смерти в пещере Вертеба.
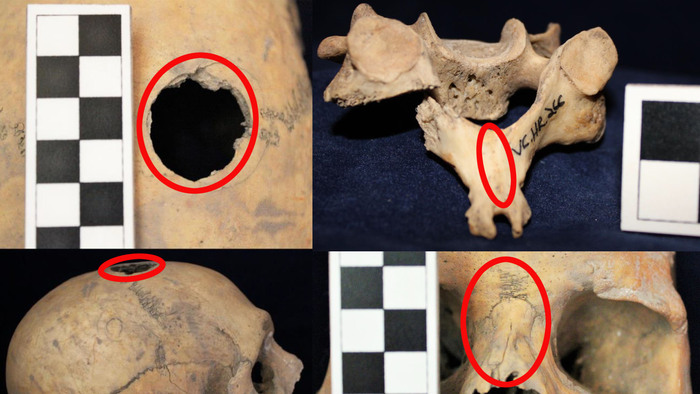
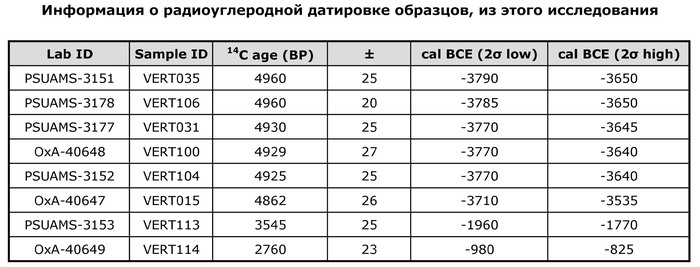
Примеры травм у захороненных в пещере Вертеба (Violence at Verteba Cave, Ukraine: New insights into the Late Neolithic intergroup conflict G.D. Madden, J.K. Karsten, S.H. Ledogar, R. Schmidt, M.P. Sokhatsky doi.org/10.1002/oa.2633)В данном исследовании авторы реконструировали последовательности генома 20 человек (8 женщин и 12 мужчин), захороненных в пещере Вертеба. Восемь образцов датированы масс-спектрометрией по углероду-14 в интервале от 3790 до 825 гг. до н. э, что соответствует трипольской культуре и позднее. Шесть человек (образцы ВЕРТ-035, ВЕРТ-106, ВЕРТ-031, ВЕРТ-100, ВЕРТ-104 и ВЕРТ-015) датируются поздним энеолитом (3790–3535 гг. до н.э.), один (VERT-113) из Зоны 7 датируется средним бронзовым веком (1960–1770 гг. до н.э.) и один из Зоны 17 (VERT-114) датируется поздним бронзовым веком (980–825 гг. до н.э.).
Используя полученные данные, авторы задались целью ответить на ряд вопросов:
1. Есть ли доказательства метисации с местными охотниками и собирателями, как было предположено в предыдущих работах?
2. Используя расширенный набор данных, можно ли уточнить неолитическую предковую составляющую трипольцев: сходны они с ранними земледельцами из Анатолии, представителями культуры линейно-ленточной керамики или иными?
3. Поскольку трипольцы жили в непосредственной близости от степных популяций, есть ли доказательства генетической близости к ямной или более ранним степным популяциям?
4. Имеет ли население более позднего бронзового века, заселившее регион, генетическую преемственность с представителями археологической общности Триполье-Кукутень из пещеры Вертеба?
Полученные результаты
Однородительские маркеры (мтДНК и Y-хромосомы)
Проанализированные образцы эпохи энеолита относятся к гаплогруппам митохондриальной ДНК T2b, H, HV, K1, N1, J1, U5 и T2c.
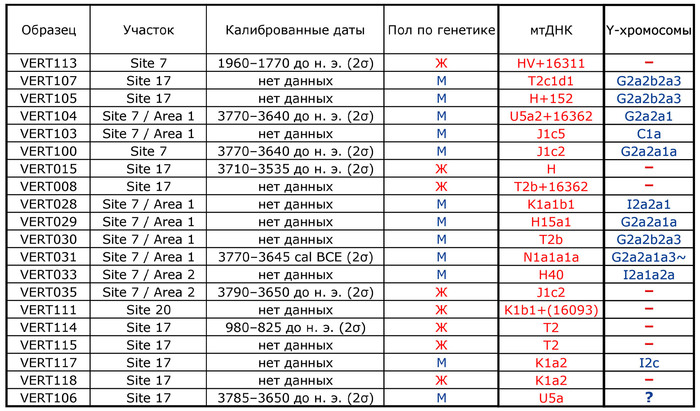
У представительницы среднего бронзового века (VERT-113) определена гаплогруппа HV, типичная для таких неолитических культур, как культура линейно-ленточной керамики к востоку от Дуная – Альфёльда, а также для жителей европейского бронзового века. Представительница позднего бронзового века (VERT-114) демонстрирует гаплогруппу T2, также связанную с многими культурами бронзового века. В целом эти гаплогруппы обычно встречаются в европейских популяциях эпохи неолита и бронзового века.
У мужчин выявлены гаплогруппы Y-хромосомы G2a, C1a и I2 (более подробно в таблице), которые также были выявлены в популяциях неолита и бронзового века Европы. При этом как гаплогруппы мтДНК, так и гаплогруппы Y-хромосомы всех образцов из Вертебы полностью согласуются с ранее опубликованными данными.
Полногеномные данные
На график анализа главных компонент вместе с 20 геномами из пещеры Вертеба авторы поместили 729 представителей современных популяций из Европы, а также 478 ранее опубликованных древних генома.

18 из 20 человек из Вертебы тесно связаны с европейскими популяциями эпохи неолита и энеолита: культура линейно-ленточной керамики, средний и поздний неолит Центральной Европы, а также трипольцы из Молдовы.
Анализ главных компонент также показал значительное сходство между 18 новыми образцами из Вертебы и четырьмя другими трипольцами из более ранней работы, также обнаруженных в пещере Вертеба. Поэтому их объединили в 22 образца, пометили на графике как Verteba_Trypillia и в дальнейшем проанализировали совместно.
А вот два генома бронзового века являются явными исключениями. Образец VERT-114 (позднего бронзового века) подпадает под разнообразие представителей культуры колоколовидных кубков и, по-видимому, занимает положение, близкое к чешской, венгерской и польской группам этой культуры. А образец, обозначенный как VERT-113 (среднего бронзового века), близок к европейским популяциям культур шнуровой керамики и срубной, демонстрируя значительное сходство со степным населением. Сразу стоит отметить, что статистический анализ указал на отсутствие доказательств связи со срубной культурой.
При этом все 22 трипольца из пещеры Вертеба содержат наследственный компонент, доминирующий у представителей анатолийского неолита (по методу ADMIXTURE).
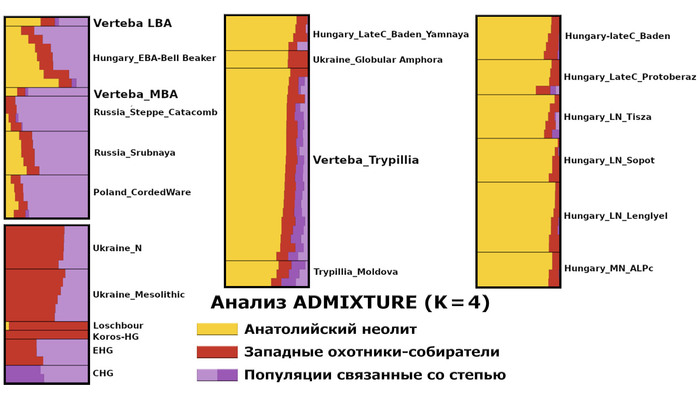
Это указывает на явную связь с европейскими неолитическими популяциями, как собственно и было показано в предыдущих исследованиях. Однако эти же образцы также демонстрируют присутствие компонентов восточных, кавказских и западных охотников-собирателей. Лишь один образец (I3151), по-видимому, не имеет предков восточных/кавказских охотников-собирателей.
В геноме образца эпохи поздней бронзы (VERT-114, укладывающимся в разнообразие представителей культуры колоколовидных кубков) преобладает компонент анатолийского неолита, но выявлен значительный компонент восточных охотников и собирателей. А геном среднего бронзового века (VERT-113) демонстрирует высокую степень сходства с геномами культуры шнуровой керамики и ямной культуры. При этом хотя моделирование геномов трипольцев свидетельствует об их связи со степными популяциями, статистически (f4) они не связаны с популяциями ямной культуры из ареалов современных России и Украины.
Моделируя величину вклада различных основных предков, которые могли бы внести свой вклад в генетический портрет трипольцев, показано, что 40% вклада связано с анатолийским неолитом, 20% - с западными охотниками-собирателями и 40% - с кавказскими охотниками-собирателями.
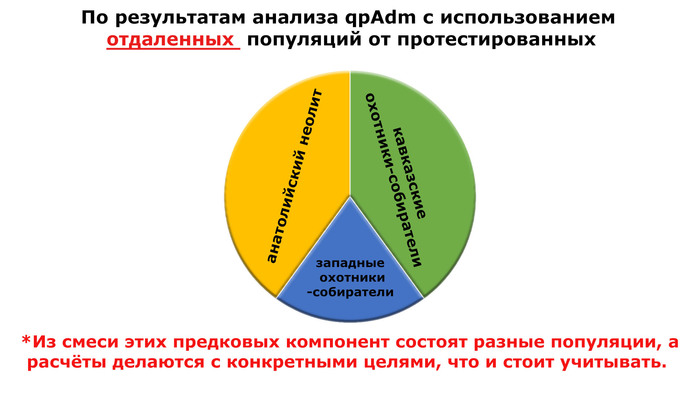
При этом охотники-собиратели юго-восточной Венгрии – Кёрёш (Koros_HG), являются источником западных охотников-собирателей у трипольцев из Вертебы. Также неолит Украины и западные охотники-собиратели являются двумя вероятными источниками генетического пласта, связанного с охотниками и собирателями, для трипольцев из Вертебы, в дополнение к смоделированному компоненту Венгрия-поздний энеолит-ранний бронзовый век-ямная (Hungary_LateC_EBA_Baden_Yamnaya).
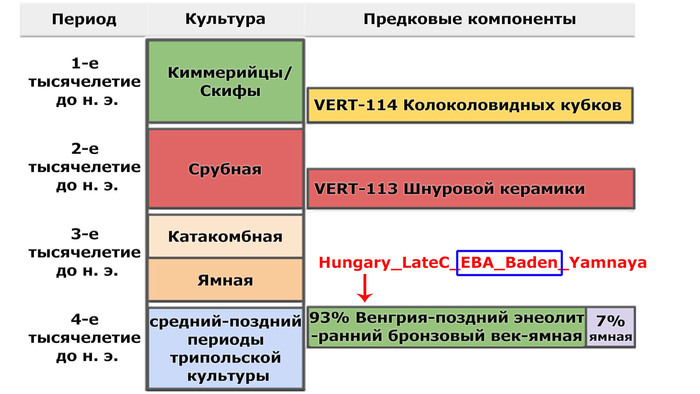
Это также указывает на очень незначительное присутствие среди предков трипольцев местных охотников-собирателей.
Кстати обратите внимание как происходит замена популяций на одной и той же территории и как сильно современные популяции отличаются от древних, вопреки распространяемым мифам.
Также стоит отметить, что большинство трипольцев из пещеры Вертеба не имеют статистически значимого сходства с молдавской популяцией археологической общности Триполье-Кукутень, но этих различий недостаточно, чтобы считать их разными популяциями.
Образец среднего бронзового века (VERT-113) является единственным, который демонстрирует четкий сигнал степного происхождения, причём наблюдается большая близость к ямной культуре с территории России, а не Украины
Кроме того, это единственный образец, который демонстрирует большую близость к представителям неолита на территории Украины, а не к западным охотникам-собирателям. В его геноме фиксируется до 33% вклад неолита Украины и 66% кавказских охотников-собирателей, что подтверждает большую долю степного происхождения.
Образец позднего бронзового века (VERT-114) генетически близок к геномам колоколовидных кубков на графике главных компонент и при моделировании «примесей» (ADMIXTURE). Однако, хотя модель с популяцией колоколовидных кубков в качестве единственного источника работает, есть и модели двух источников включающие 60% представителей культуры шаровидных амфор с территории Украины и 40% степных популяций.
Авторы также исследовали возможную связь генома поздней бронзы их Вертебы и киммерийцами, которые обосновались на территории современной Украины около 1000 г. до н. э. Однако геном из Вертебы оказался значительно ближе к носителям культуры колоколовидных кубков, чем к киммерийцам.
Анализ близкого родства в изученных геномах
Размеры гомозиготных участков ДНК указывают, что обитатели Вертебы жили в довольно многочисленных популяциях.
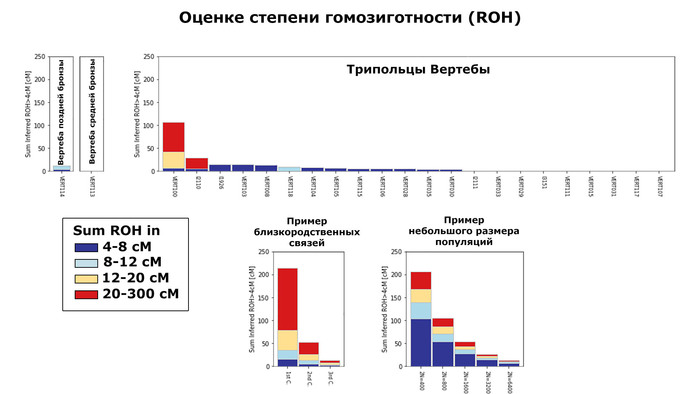
Исключением стал образец VERT-100, предки которого, вероятно, были родственниками.
Фенотипические и биохимические особенности
Ни один из проанализированных людей пещеры Вертеба не обладал толерантностью к лактозе.
Лишь два человека, видимо, были темноглазыми, но остальные обладали ДНК-маркером rs12913832, связанным с голубым цветом глаз.
Обсуждение
Генетический анализ 20 человек из пещеры Вертеба, которые датируются четвертым, вторым и первым тысячелетиями до нашей эры, выявил важные генетические изменения как в раннем бронзовом веке, так и в позднем. Эти датировки подтверждены радиоуглеродным датированием образцов 8 человек, поскольку пещера Вертеба многократно использовалась человеком, начиная с мезолита.
Предыдущие анализы митохондриальной ДНК представителей археологической общности Триполье-Кукутень показали их близкое родство по материнской линии с популяциями раннего неолита: гаплогруппы H, HV, T, V, J и K. И в новом анализе, за исключением двух человек с гаплогруппой U5a, все остальные 18 человек, имеют гаплогруппы, сходные с неолитическим населением Центральной Европы. Это разнообразие мтДНК резко контрастирует с населением более ранних неземледельческих неолитических памятников на территории Украины, которые имеют только гаплогруппу U, указывающую на преемственность с мезолитическими охотниками-собирателями.
Mathieson, I., Alpaslan-Roodenberg, S., Posth, C. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018). doi.org/10.1038/nature25778Разнообразие гаплогрупп мтДНК предполагает, что местное население в значительной степени было заменено населением, связанным с трипольской культурой. Большинство представителей Вертебы имеют Y-гаплогруппу G2a2, которая широко распространена среди европейцев эпохи неолита, связанных с Анатолией.
Другие выявленные гаплогруппы - C1 и I2 - также обнаружены среди европейских неолитических популяций.

Это указывает на миграцию и мужчин, и женщин, что контрастирует со степными миграциями во время бронзового века.
Данные популяционной генетики показывают, что люди, похороненные в пещере Вертеба в эпоху позднего энеолита (3790-3535 гг. до н. э.), генетически похожи на другие, ранее проанализированные, геномы из Молдовы. Эти наблюдения в целом предполагают, что люди энеолита археологической общности Триполье-Кукутень произошли от того же или близкородственного населения, которое распространило неолит по большей части Европы (в случае с трипольцами конкретно указывая на баденцев из Венгрии), и практически без признаков метисации с более ранним мезолитическим или неолитическим населением с территории современной Украины, связанным с охотниками-собирателями.
Фактически, большинство трипольцев могут моделироваться энеолитическими популяциями из Европы, имеющими степное происхождение, однако четверо из 20 человек могут быть смоделированы как трипольцы из Молдовы.
Результаты моделирования (qpAdm) предполагают, что существовали различия в составе предков трипольцев, которые могут быть связаны с долей охотников и собирателей, хотя, как отмечалось выше, эта изменчивость недостаточно существенна, чтобы разделить людей с разных участков на разные популяции.
При этом источником генетического пласта, унаследованного от охотников-собирателей у трипольцев, являются в основном западные охотники-собиратели. Значительная часть предков западных охотников-собирателей, обнаруженная у трипольцев (до 18%), может быть связана с увеличением доли этого компонента и в других популяциях среднего неолита Центральной Европы, вероятно, из-за смешения с популяциями на западе, которые уже имели более высокий компонент западных охотников-собирателей, полученный от неолитических групп, связанных с Анатолией до возникновения археологической общности Триполье-Кукутень.
Это также указывает на то, что неолитические популяции охотников-собирателей на территории Украины не внесли большого вклада в происхождение трипольцев. Кроме того, как и в Молдове, выявлен генетический вклад степного происхождения, но в Вертебе он ниже. Это может быть связано с их разными датировками, если предполагать непрерывный импульс движения с востока на запад, постепенно увеличивающий связь с ямной культурой в течение четвертого тысячелетия до н. э.
А вот геном из среднего бронзового века (VERT-113, 1960-1770 гг. до н. э.) значительно отличается от более ранних трипольцев: для него характерен больший вклад кавказских охотников-собирателей/ямной и восточных охотников-собирателей. Результаты (qpAdm) предполагают связь между геномом VERT-113 и представителями культуры шнуровой керамики из Польши. Кроме того, это единственный геном, который ближе к неолиту Украины, чем к западным охотникам и собирателям. Это позволяет предположить, что популяция, возникшая во втором тысячелетии до н. э., возможно, имела общие черты с популяциями неолита на территории Украины.
Интересно, что у генома поздней бронзы (VERT-114) нет значимой связи с геномом средней бронзы (VERT-113). Геном VERT-114 предполагает связь с культурой колоколовидных кубков, несмотря на то, что этот человек почти на 2000 лет моложе конца этого надкультурного феномена, и с датировкой ближе к киммерийцам или скифам. Это подтверждает данные, приведенные в более ранних работах, о притоке западного населения в степь в конце бронзового века.
Однако для понимания более полной картины необходимо получить ещё больше образцов разных периодов из этого региона.
Результаты новой работы имеют важное значение для понимания процесса неолитизации в Восточной Европе. Поскольку население археологической общности Триполье-Кукутень распространилось с современных территорий Румынии и Молдовы в лесостепные районы Западной и Центральной Украины, оно должно было вступить в контакт с населением, связанным с местной буго-днестровской культурой, чья система жизнеобеспечения была сосредоточена главным образом на охоте, рыбалке и собирательстве. Эта группа, вероятно, представляла собой потомков мезолитических охотников и собирателей. Анализ геномов населения пещеры Вертеба предполагает, что местные мезолитические охотники-собиратели не внесли существенного вклада в генофонд более поздних трипольцев. Иными словами, процесс неолитизации в Западной Украине был в большей степени результатом миграции (демической диффузии), а не принятия местными жителями практики земледелия (аккультурации).
Результаты также подтверждают гипотезу длительно существовавшей границы между оседлыми земледельцами лесостепной зоны и соседними кочевыми скотоводами из Причерноморской степи. Эта граница характеризуется резкими контрастами в материальной культуре и способах жизнеобеспечения и, вероятно, сохранялась благодаря этим факторам и языковым различиям. Отсутствие метисации на этой культурной границе является ключом к пониманию контекста, в рамках которого произошла миграция представителей ямной культуры.
В целом результаты показывают, что пещера Вертеба представляет собой важный погребальный комплекс, соединяющий Восток и Запад. Генетическая структура представителей археологической общности Триполье-Кукутень включает предковые компоненты, связанные как с более ранними западными охотниками-собирателями, так и с земледельцами Ближнего Востока, а также те, которые генетически отличаются от предков молдавских представителей этой общности.
Отсутствие местных предков, связанных с охотниками-собирателями эпохи неолита на территории современной Украины, предполагает, что эти земледельцы в основном заменили местных охотников-собирателей и не смешивались с соседними степными популяциями. Кроме того, в течение бронзового века пещера Вертеба использовалась последовательными волнами кочевников-скотоводов с востока, которые в конечном итоге принесли значительные генетические, а также культурные изменения в Европу и в конечном итоге смешались с местными потомками населения трипольской культуры.
Дополнительные генетические данные из этих и более поздних временных периодов позволят дополнить картину, а также ответить на вопросы хронологии и, возможно, указать, причины краха трипольской культуры.
Источник: Gelabert, P., Schmidt, R.W., Fernandes, D.M. et al. Genomes from Verteba cave suggest diversity within the Trypillians in Ukraine. Sci Rep 12, 7242 (2022). doi.org/10.1038/s41598-022-11117-8
Текстовый вариант не публикую, так как он доступен на русском языке в научном журнале Вестник РГМУ
Упоминание первых земледельцев встречается довольно часто в исследованиях, связанных с популяционной генетикой и не только, поэтому очень важно иметь представление о происхождении этих людей для дальнейшего понимания популяционной истории различных древних и современных народов.
Неолитическая революция в Западное Евразии и связанные с ней процессы привели не только к оседлости и производству продуктов питания, но также и к появлению в Европе новых людей, отличающихся от предыдущих, со своей материальной культурой и обычаями, примерно 8600 лет назад. Истоки этих процессов были отмечены в Юго-Западной Азии ≈11700 лет назад, где были впервые одомашнены растения и животные.
Среди учёных бытует устоявшееся мнение, что земледелие из этого региона распространилось в Европу по двум основным маршрутам: «средиземноморскому» и «дунайскому», последний кстати и находится в центре внимания нового исследования.
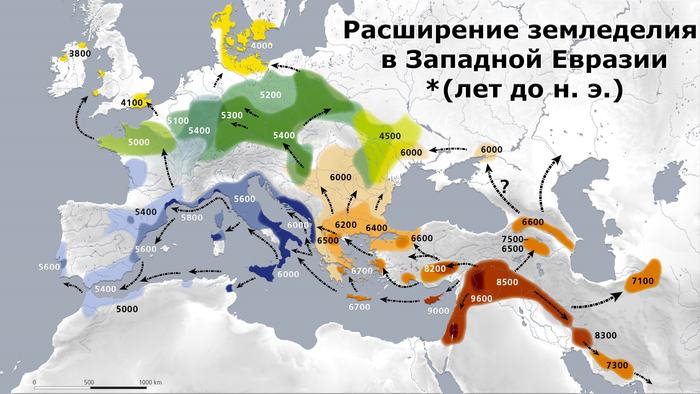
Но несмотря на это, генетическое происхождение первых земледельцев и пространственно-временные рамки, связанные с этими процессами, остаются неясными, в значительной степени из-за отсутствия высококачественных древних геномов, полученных от популяций, участвовавших непосредственно в начальном этапе распространения сельского хозяйства.
На данный момент палеогенетические исследования установили, что ранние европейские земледельцы генетически отличались от охотников-собирателей, населявших континент. А генетический обмен между ними, по-видимому, был ограничен на ранних этапах распространения земледельцев, но нарастал со временем.
Большинство земледельцев, проживавших в континентальной Европе около 9 тыс. лет назад, по-видимому, произошли от популяций, населявших районы Эгейского моря и восточную часть Мраморного, но их более точное генетическое и географическое происхождение до сих пор является предметом споров.
А ранненеолитические земледельцы Эгейского моря явно связаны с земледельцами Центральной Анатолии, но они также демонстрируют и сходство с земледельцами докерамического неолита Южного Леванта. Что предполагает общее происхождение всех этих популяций до распространения сельского хозяйства на запад, возможно, в районе Плодородного полумесяца – археологически значимом регионе, который включал в себя части современных Ирана, Ирака, Израиля, Палестины, Иордании, Ливана, Сирии и Турции. Однако исследования также показали, что эгейские земледельцы генетически отличаются от ранних популяций из восточной части Плодородного полумесяца, региона Загрос в Иране и северном Ираке, что может указывать на параллельное принятие методов земледелия генетически отдаленными группами охотников и собирателей по всей Юго-Западной Азии. Кроме того, есть некоторые свидетельства генетической преемственности между эпипалеолитическими и неолитическими популяциями Центральной Анатолии, что предполагает местный переход к сельскому хозяйству без значительного потока генов.
Помимо этого, картину усложняет тот факт, что некоторые ранние земледельцы Центральной Анатолии также демонстрируют генетическое сходство с кавказскими охотниками и собирателями, представленными геномом человека из карстового грота Котиас Клде в Западной Грузии, возрастом около 10 тыс. лет. При этом сами кавказские охотники и собиратели были тесно связаны с земледельцами раннего неолита Ирана.
Авторы новой работы задались целью реконструировать происхождение ранних земледельцев Юго-Западной Азии и Европы, а также процессы, которые способствовали их отличию от охотников и собирателей.
Для этого авторы получили 15 геномов с высоким разрешением ранних земледельцев и охотников-собирателей, распределенных по географическому и временному разрезу от Юго-Западной Азии до долины Рейна в Западной и Центральной Европе
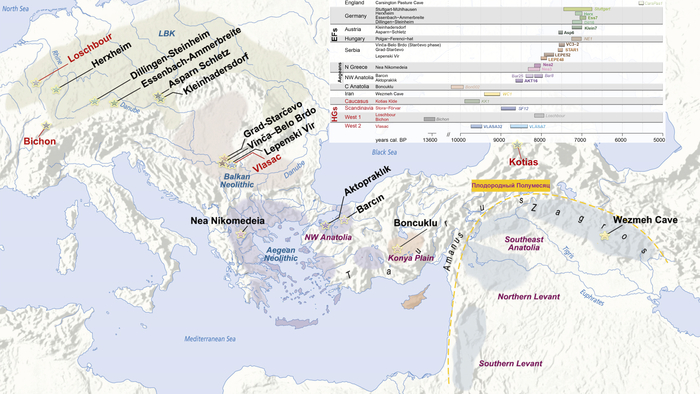
ДНК была извлечена из останков, найденных в пределах некоторых из наиболее важных археологических памятников раннего голоцена в Европе и Анатолии, включая первые поселения земледельцев в Эгейском бассейне (Барчин Хëюк, Актопраклик и Неа-Никомидия). Мезолитические и неолитические стоянки в районе Железных Ворот и других районах Центральных Балкан (Лепенски-Вир, Власац, Старчево и Винча-Бело-Брдо). А также старейшие кладбища или места массовых захоронений в Центральной Европе раннего неолита (Кляйнхадерсдорф, Аспарн-Шлётц, Эссенбах-Аммербрайт, Диллинген-Штайнхайм и Херксхайм).
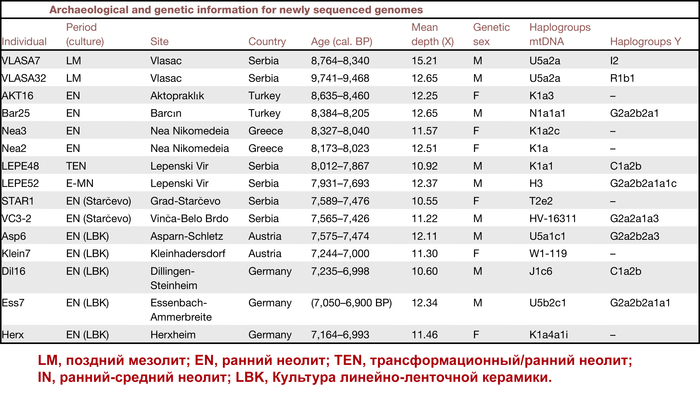
Результаты
Генетическая структура и сходство древних людей
Анализ методом многомерного шкалирования древних и современных людей разделяет образцы на три кластера:
1. Европейских охотников и собирателей.
2. Западных ранних земледельцев Европы и Анатолии.
3. В третий кластер вошли – представитель ранних земледельцев из Ирана и мезолитический охотник-собиратель Кавказа из грота Котиас Клде.
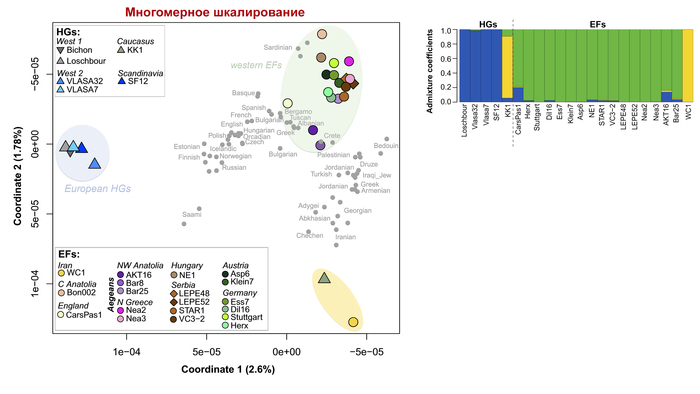
В соответствии с предыдущими анализами, основанными на установленных отличиях последовательности ДНК размером в один нуклеотид, западные ранние земледельцы демонстрируют самое сильное сходство с современными сардинцами, за исключением одного древнего жителя из Великобритании (из пещеры на пастбище Карсингтон CarsPas1) и двух представителей ранних земледельцев Северо-Западной Анатолии (Барчин8 и Актопраклик16), которые, как обнаружено, группируются с современными жителями других частей Южной Европы. А вот европейские охотники и собиратели палеолита и мезолита заметно генетически отличаются от всех современных европейцев.
Ранние иранские земледельцы и кавказские охотники-собиратели, по-видимому, генетически близки к современным популяциям из их района отбора проб, что соответствует некоторой долгосрочной генетической преемственности в этих регионах.
Демографическое моделирование с использованием генетической информации
На схеме демографического моделирования обозначены 3 метапопуляции:
1. Западная, состоящая из линии западноевропейских охотников-собирателей.
2. Центральная – включающая ранних земледельцев из Европы и Анатолии.
3. Восточная, в которую вошли ранние иранские земледельцы.
Благодаря такому моделированию авторы установили ряд важных особенностей.
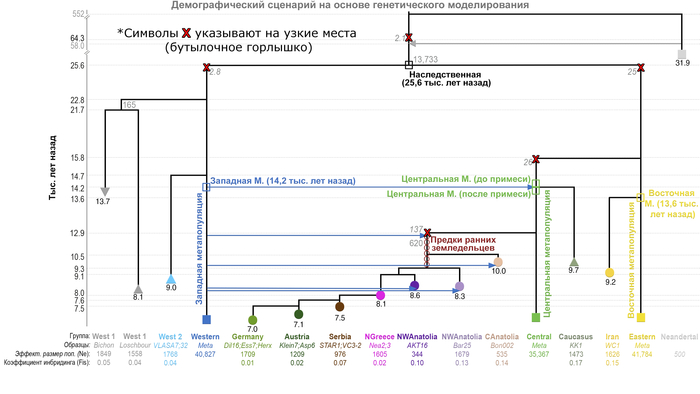
Все западные ранние земледельцы имеют отдаленных общих предков с кавказскими охотниками и собирателями
Этот вывод согласуется с недавним генетическим исследованием, которое показало, что человек из грота Котиас Клде был более тесно связан с ранними западными земледельцами, чем с западноевропейскими охотниками-собирателями. При этом эта предковая для ранних земледельцев из Европы и Анатолии метапопуляция получила около 14% своего генофонда от западноевропейских охотников-собирателей около 14,2 тыс. лет назад. Однако эта примесь не затронула предков популяций иранского неолита, потому как эта линия отделилась от предков ранних земледельцев из Европы и Анатолии ≈15,8 тыс. лет назад, т.е. примерно за 1600 лет до смешения последних с западноевропейскими охотниками и собирателями. Помимо этого, несмотря на то, что кавказские охотники и собиратели демонстрируют более тесное генетическое сходство с ранними земледельцами из Ирана, новые результаты показывают, что они также имеют общее происхождение и со всеми ранними земледельцами из Европы и Анатолии. А стало быть близки к линии примеси с западноевропейскими охотниками и собирателями уже после отделения от линии предковой иранским земледельцам.
Предки ранних анатолийских и европейских земледельцев дважды смешивались с западными охотниками и собирателями
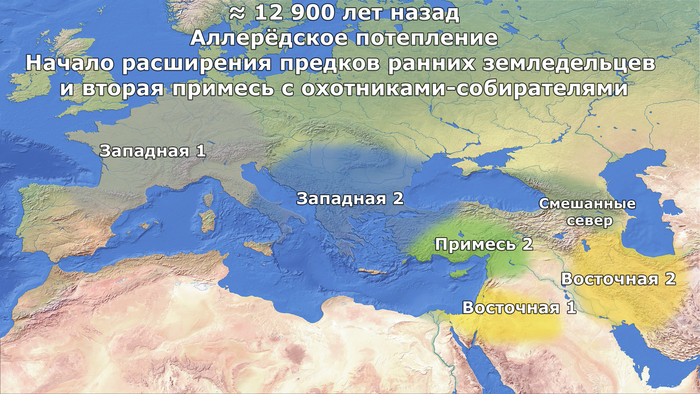
Данные указывают на то, что предки ранних земледельцев получили второй импульс потока генов, около 15% от западноевропейских охотников и собирателей ≈12900 лет назад, в отличие от кавказских охотников-собирателей.
Таким образом, предки анатолийских и европейских земледельцев являются продуктом повторяющихся потоков генов от западноевропейских охотников-собирателей начиная от 12900 лет назад и до 9100 лет назад. При этом их эффективный размер популяции сократился до 620 человек. В следствии чего эти популяции стали отличаться не только от своих предков, но и от кавказских и европейских охотников-собирателей, а также от иранских земледельцев.

Кстати не стоит путать эффективный размер популяции с общей численностью.
Разделение анатолийских и эгейских земледельцев
Популяции из Северо-Западной Анатолии (Актопраклик и Барчин) и Северной Греции (Неа-Никомидия), по–видимому, отделились друг от друга примерно в одно и то же время ∼9100-9300 лет назад, возможно, во время "освоения" ранними земледельцами более обширных территорий у берегов Эгейского моря.

А вот ранние земледельцы из Центральной Анатолии (представленные образцом генома из Бончуклу) выделились по крайней мере на 1000 лет раньше ≈10,5 тыс. лет назад.

При этом тот факт, что анатолийские и эгейские популяции демонстрируют различное количество недавнего для их периода потока генов от окружающих их западноевропейских охотников-собирателей, предполагает разные уровни взаимодействия с ними.
Действительно, геномы из Северной Греции демонстрируют более слабый поток генов от охотников и собирателей – около 3%, в отличие от 10% у жителей Центральной Анатолии из Бончуклу, а также от 12% до 17% у обитателей Северо-Западной Анатолии, участков Барчин и Актопраклик соответственно.
Поэтапная демическая диффузия неолитических земледельцев в Центральную Европу
Авторы считают, что простая модель со строгой поэтапной миграцией ранних земледельцев, происходящей из более широкого региона Эгейского моря (Северо-Западная Анатолия или Северная Греция) в Сербию через Балканы и вдоль так называемого "Дунайского коридора" (долин Дуная и Рейна), а затем в Австрию и, в конечном итоге, в Германию, поддерживается лучше, чем сценарий, допускающий дальнюю миграцию от Эгейского моря непосредственно в Австрию.
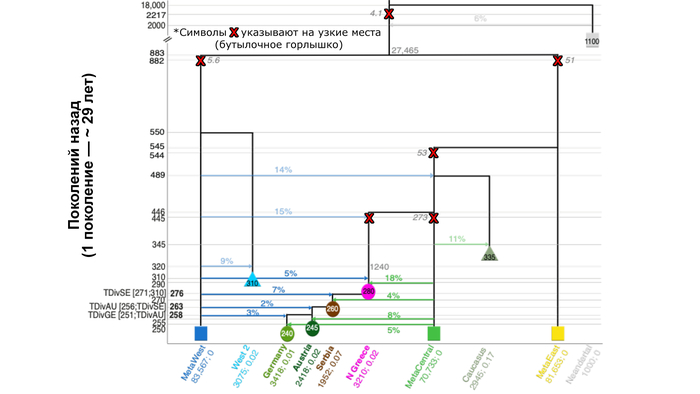
Данные также указывают на то, что сообщества ранних земледельцев включали в себя некоторое количество охотников-собирателей на каждом смоделированном этапе их расселения вдоль "Дунайского коридора" от 2% до 7%, что согласуется и с предыдущими оценками – от 3 до 9%.
Разделение между западноевропейскими охотниками-собирателями и предками ранних иранских земледельцев
Новая модель также дает важные сведения о глубоком разветвлении донеолитических популяций. Расхождение между западноевропейскими охотниками-собирателями и предками ранних иранских земледельцев оценивается примерно в 25600 лет назад. Это намного моложе, чем предполагалось ранее – не менее 46 тыс. лет назад.
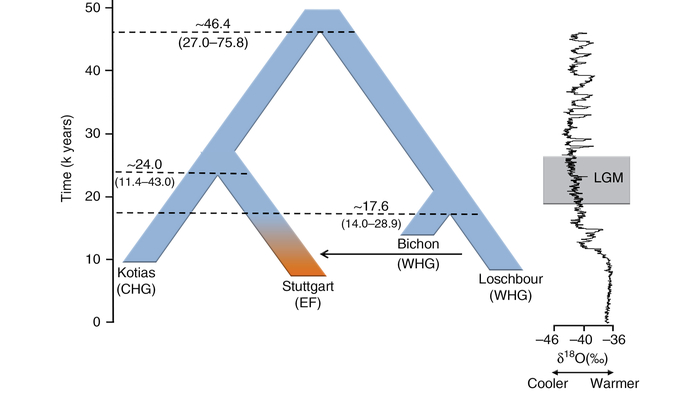
Однако эти предыдущие даты были получены с использованием относительно простых моделей без учёта узких мест и только с недавними примесями или вообще без примесей.
При этом сравнение двух западноевропейских охотников-собирателей из Бишона и Лошбура с недавно секвенированными жителями мезолита из Власаца также показывает, что европейские популяции охотников-собирателей уже разделились во время последнего ледникового максимума (LGM) ∼ 22800 лет назад, причем популяции из Бишона и Лошбура впоследствии отделились друг от друга примерно через 1000 лет.
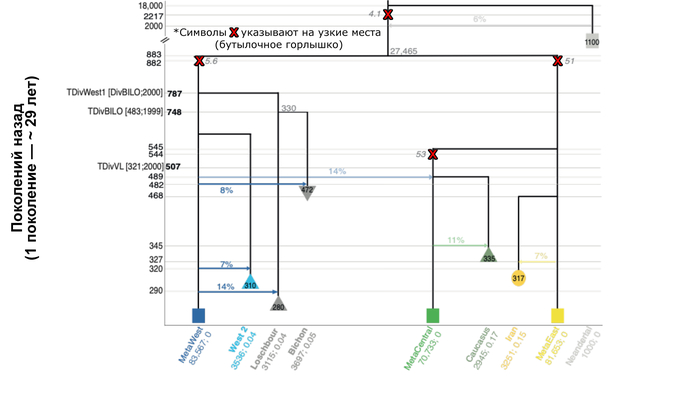
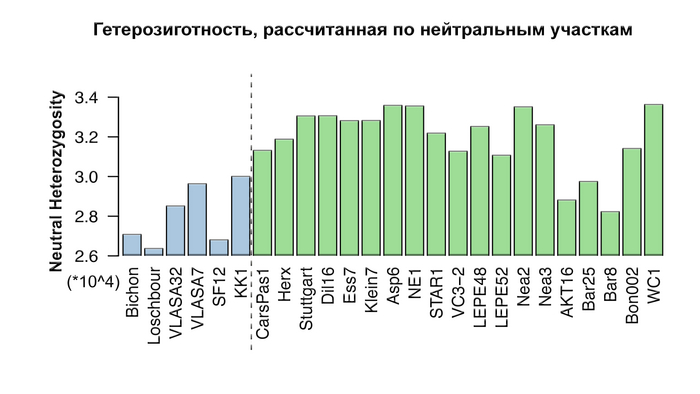
Сокращение разнообразия европейских охотников и собирателей
Генетическое разнообразие, количественно определяемое гетерозиготностью по нейтральным участкам, у охотников и собирателей намного ниже, чем у ранних земледельцев, за исключением их представителей из Северо-Западной Анатолии, что соответствует и предыдущим исследованиям.
Также охотники-собиратели по показателю гомозиготности (ROH) демонстрируют фоновое родство, которое обычно связывают с небольшим размером популяции.
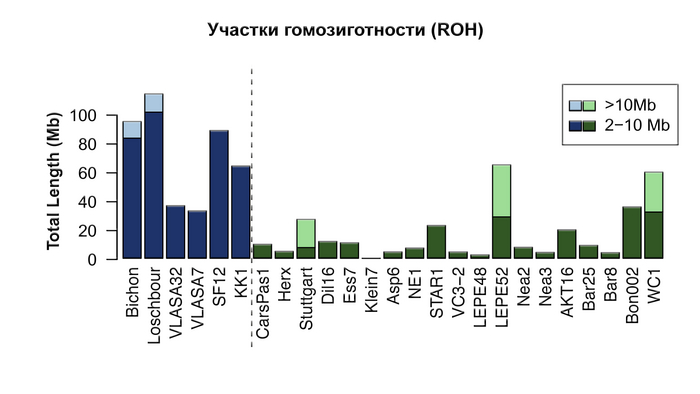
При этом авторы оценивают эффективный размер популяции охотников и собирателей выше, чем у большинства ранних земледельцев, особенно из Анатолии, где их эффективный размер популяции составлял всего несколько сотен человек, несмотря на их очевидно большую численность. Таким образом, меньшее разнообразие, наблюдаемое у западноевропейских охотников-собирателей, скорее всего больше связано с эффектом бутылочного горлышка, во времена максимума последнего оледенения, чем с жизнью в небольших изолированных группах.
Соответствие фактических данных с моделями
Авторы отмечают, что геномные данные, смоделированные в соответствии с наиболее полным демографическим сценарием, приводят к таким соотношениям между популяциями, которые очень похожи на наблюдаемые на графике многомерного шкалирования, по фактическим данным. Отчетливо видны три кластера, причем все европейские охотники и собиратели находятся близко друг к другу и заметно отличаются от ранних земледельцев, а вот кавказские охотники-собиратели наоборот, близки к иранским земледельцам.
Таким образом, хорошее соответствие между наблюдаемыми и смоделированными данными обеспечивает апостериорную проверку данных из исследования.
Итоги
Раскол европейских охотников и собирателей, вызванный максимумом последнего оледенения
Новая модель предполагает, что европейские охотники и собиратели уже разделились на две подгруппы ∼ 22800 лет назад, после того как во время максимума последнего оледенения столкнулись с эффектом бутылочного горлышка, который ответственен за их низкий уровень генетического разнообразия. Но в отличие от предыдущих исследований, было обнаружено, что охотники и собиратели, как правило, имели больший эффективный размер популяции, чем ранние земледельцы. Такие размеры могли привести к медленному различию между ними, что может объяснить, почему разные группы охотников и собирателей демонстрируют генетическое сходство, несмотря на длительное время расхождения и широкое географическое распространение. Большие эффективные размеры популяции охотников-собирателей могут быть обусловлены генетическим обменом между группами на большие расстояния. Напротив, предполагаемый низкий эффективный размер популяции ранних земледельцев (несмотря на очевидно большие размеры общей численности) мог быть вызван “оседлостью” или привязанностью к месту и ограниченным потоком генов между мелкими сообществами земледельцев, как на участках докерамического неолита Бончуклу и Ашиклы-Хююк.
Связь предков ранних земледельцев с кавказскими охотниками и собирателями
Результаты демонстрируют, что охотники и собиратели Кавказа имеют общего предка с ранними западными земледельцами, поскольку и те, и те демонстрируют следы наследственной примеси от западноевропейских охотников-собирателей и предков ранних иранских земледельцев. При этом несмотря на схожие модели примесей у кавказских охотников и собирателей с иранскими земледельцами, они всё же не так близки, как кажется.
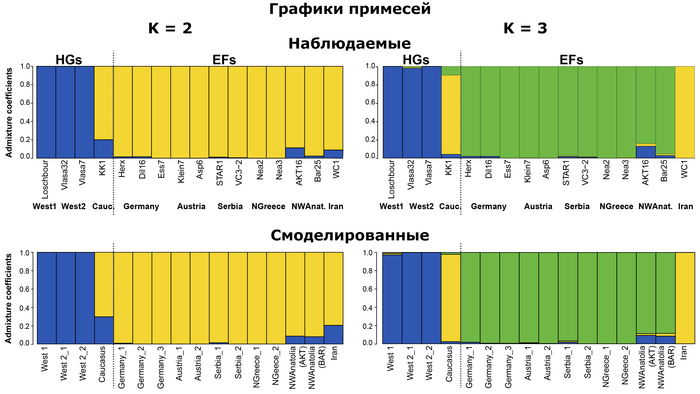
А это указывает на то, что наблюдаемых закономерностей генетического сходства недостаточно для восстановления фактических родственных связей.
Что привело к отличию ранних земледельцев от мезолитических охотников-собирателей и от их предковых популяций
Предки ранних земледельцев из Европы и Анатолии изначально были близки с предками иранских земледельцев, однако два последовательных события смешения с западноевропейскими охотниками и собирателями изменили их состав, после чего 2500 лет интенсивного генетического дрейфа сделали их особенно отличными от всех других групп.
Авторы предполагают, что этот быстрый процесс, который привёл к различию между популяциями, мог быть вызван повторяющимися эффектами основателя, возникшими во время их расселения по Анатолии. Это когда новые территории заселялись изначально малым количеством людей.
Хотя западные ранние земледельцы ранее были описаны как генетически промежуточные между другими группами Юго-Западной Азии или как смесь неолитических популяций Ирана и Южного Леванта с западными охотниками-собирателями, этот первоначальный сигнал о примеси оставался скрытым для классического анализа, поскольку он постепенно размывался генетическим дрейфом, который произошел во время миграции этих популяций через Анатолию.
В то время как популяции, смоделированные вскоре после двух основных событий смешения с западноевропейскими охотниками-собирателями, выглядят как смешанные, этот сигнал постепенно исчезает с течением времени с появлением ранних земледельцев и их дальнейшей миграцией, после чего последние демонстрируют уже совершенно неродственный генофонд.
Пространственная интерпретация разделения населения
Сроки и последовательность демографических событий, вытекающих из новой модели, предполагают сценарий разделения популяций с четким географическим и хронологическим разрешением от максимума последнего оледенения до раннего голоцена.
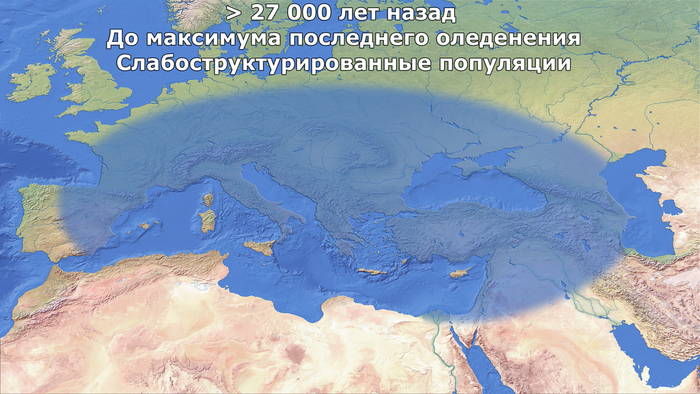
Структурирование и разделение охотников-собирателей, вызванное максимумом последнего оледенения
Авторы считают, что расхождение между западноевропейскими охотниками-собирателями и предками ранних иранских земледельцев, было инициировано максимумом последнего оледенения около 26 тыс. лет назад.
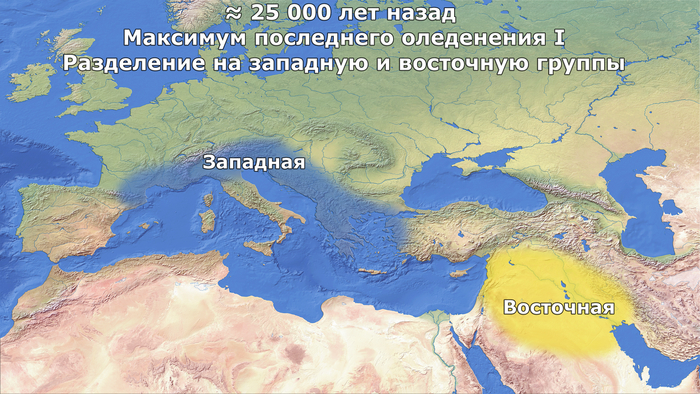
Вероятно, из-за ухудшения среды обитания и сокращения числа убежищ, потенциально расположенных в регионах с более мягким климатом – рефугиумах.

Археологические, экологические и климатические данные действительно свидетельствуют о том, что большие участки Евразии были пустынны в разгар максимума последнего оледенения, ≈26 тыс. лет назад, когда ледниковые щиты достигли своей максимальной протяженности, и популяции как людей, так и животных выжили в рефугиумах, расположенных в более южных регионах Европы. При этом если бы эффект бутылочного горлышка длился на протяжении 4000 лет (что соответствует 138 поколениям по 29 лет), то эффективный размер популяции сократился бы до 383 человек. Такое низкое число согласуется с археологическими данными, свидетельствующими о 60-процентном сокращении численности населения ближе к концу периода граветтской культуры около 29-25 тыс. лет назад, при общей численности популяции в Европе всего 700-1550 человек.
Анализ также показывает, что европейские охотники и собиратели сформировались в двух отдельных рефугиумах к концу ледникового максимума 21,7 тыс. лет назад, что, возможно, соответствует распространению солютрейской и эпиграветтской культур.
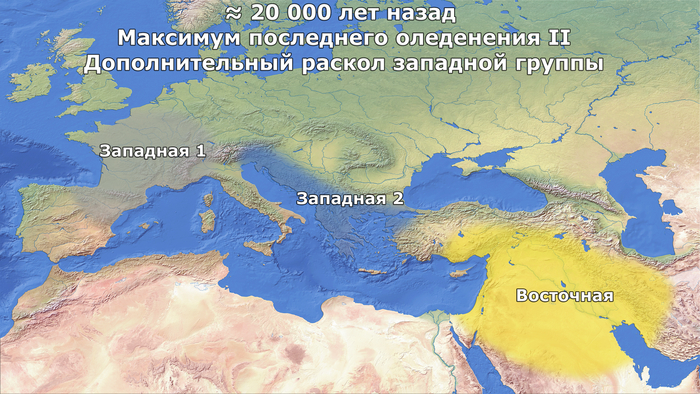
Увеличение разнообразия и примеси после максимума последнего оледенения
После периода расширения ареала обитания

в конце максимума последнего оледенения охотники и собиратели - предки ранних земледельцев из Европы и Анатолии, которые, вероятно, были потомками популяции из эпиграветтского рефугиума, ≈14,2 тыс. лет назад смешались с популяцией, предковой как кавказским охотникам и собирателям, так и ранним западным земледельцам (Восточные 1).

Учитывая предполагаемое географическое распределение предшествующих ледниковых рефугиумов, эта примесь времён Бёллингского потепления, вероятно, произошла в регионе, охватывающем Юго-Восточную Анатолию и Северный Левант, или даже в соседних регионах, таких как Центральная и Восточная Анатолия или Южное побережье современной Турции.
Разделение анатолийских и эгейских земледельцев
Демографические процессы, лежащие в основе различий между населением Центральной Анатолии и Эгейского моря, труднее точно определить и локализовать. Предполагаемая низкая численность популяции предков западных ранних земледельцев после их отделения от центральной метапопуляции во время среднего дриаса может быть вызвана расширением ареала на запад и связанными с этим повторяющимися эффектами основателя во времена аллерёдского потепления.
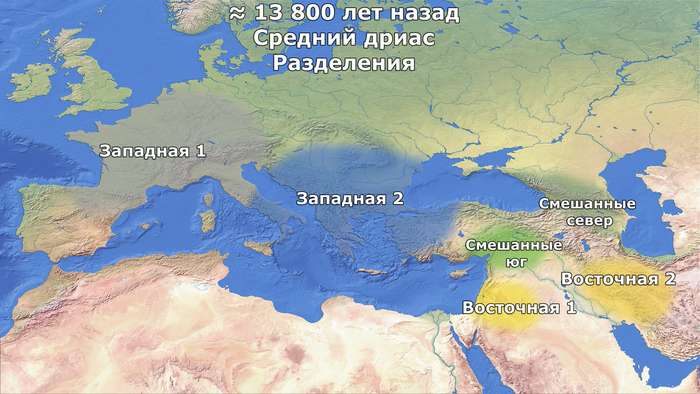
Этот период характеризовался относительно благоприятными климатическими условиями, которые, возможно, позволили предкам западных ранних земледельцев в дальнейшем смешаться с эпипалеолитическими охотниками и собирателями в Анатолии (≈12,9 тыс. лет назад).
Тот факт, что земледельцы Центральной Анатолии разделяют одно и то же событие смешения и дрейфа с эгейскими земледельцами, предполагает, что они были частью одной и той же волны миграции и что Центральная Анатолия была заселена земледельцами до того, как они достигли региона Эгейского моря, возможно, другим маршрутом.
Однако предыдущее исследование показало, что центрально-анатолийские земледельцы и эпипалеолитические охотники-собиратели были генетически схожи, а это указывает на то, что в Центральной Анатолии смешанные группы существовали ещё до перехода к неолиту. Хотя по альтернативному сценарию, смешанные популяции охотников и собирателей могли переселиться туда из региона Плодородного полумесяца, переняв полностью развитые методы ведения сельского хозяйства на более позднем этапе, что согласуется с наблюдением, что ранние участки докерамического неолита, такие как Бончуклу и Ашиклы на Анатолийском плоскогорье, демонстрируют эксперименты по выращиванию сельскохозяйственных культур и содержанию коз ∼9,7 тыс. лет назад.
Напротив, миграция в Северо-Западную Анатолию, вероятно, произошла во времена полностью развитого керамического неолита, характеризующегося установлением широко распространенного смешанного земледелия.
Неолитическая экспансия происходила в результате смеси культурной и демической диффузии
Несмотря на то, что первоначальное распространение неолита, должно быть, происходило путем культурной диффузии в Плодородном Полумесяце среди генетически хорошо различающихся групп, новые результаты показывают, что расширение в Северо-Западную Анатолию, Эгейский бассейн и через Дунайский коридор происходило главным образом за счет демической диффузии.

Первоначальное распространение населения за пределы Плодородного полумесяца, безусловно, было далеко не линейным и связано с многочисленными генетическими влияниями из Леванта, некоторые из которых не были связаны с возникновением полностью развитой неолитической экономики. А как только неолитический образ жизни достиг Европы, способ расселения в Центральную Европу стал более линейным и, по сути, может быть смоделирован как поэтапный.
Выводы
Популяционное моделирование, описанное в новой работе, позволило авторам извлечь ключевую, неожиданную, но дополняющую и гораздо более подробную информацию о сходстве населения, чем можно было бы сделать на основе сводной статистики и одного только многомерного анализа. Кроме того, эта модель обеспечивает временные рамки для разделения основных групп, населявших Юго-Западную Азию и Европу, от максимума последнего оледенения до внедрения сельского хозяйства и подчеркивает решающую роль климатических изменений как фактора разделения населения и событий смешения.
Хотя первые в мире земледельцы генетически сильно отличаются от европейских охотников и собирателей, моделирование, основанное на высококачественных геномах, показывает, что некоторые популяции Европы и Юго-Западной Азии на самом деле имели общую историю, отмеченную повторяющимися взаимодействиями с конца последней ледниковой эпохи. Сильный генетический дрейф во время их экспансии через Анатолию способствовал тому, что ранние западные земледельцы выглядели более непохожими, чем они были на самом деле, скрывая их смешанное происхождение. Подводя итог, можно сказать, что идея единого культурного и генетического происхождения всех земледельцев Плодородного полумесяца без значительного первоначального вклада европейцев, в качестве которых выступали охотники и собиратели, в ее нынешнем виде, больше не выдерживает критики.
Источник:
The genomic origins of the world’s first farmers
Nina Marchi, Laura Winkelbach, Ilektra Schulz, Maxime Brami, Zuzana Hofmanová, Jens Blöcher, Carlos S. Reyna-Blanco, Yoan Diekmann, Alexandre Thiéry et al https://doi.org/10.1016/j.cell.2022.04.008
Авары прибыли в Карпатский бассейн в 567–568 гг., а их империя, управляемая каганом, доминировала в восточной части Центральной Европы более 200 лет, пока не была завоевана франками около 800 гг. н. э. Византийские источники сходятся во мнении, что их движение на запад было спровоцировано становлением Тюркского каганата в 550-х годах с центром на территории современной Монголии, когда тюрки разрушили Жужаньский каганат. Однако в текстах нет единого мнения о том, кем были эти люди и откуда именно они пришли. При этом византийские источники, с подачи тюрков, называли их «псевдоаварами», которые якобы присвоили имя авар и высокий титул кагана, а на самом деле были огурами – тюркоязычным народом западной части Центральной Евразии. Хотя можно считать, что жужани, скорее всего, называли себя аварами, вопрос о том, в какой степени европейские авары произошли от них, является предметом споров.
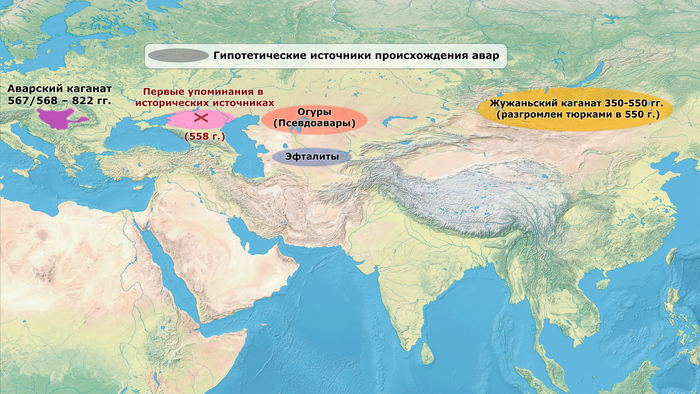
Также сразу хочу отметить, что не стоит путать авар, о которых пойдёт речь, с аварцами Кавказа. Это исторически и генетически разные народы.А в новой работе авторы представляют новые данные, которые в дополнение к предыдущим дают новую основу для реконструкции раннесредневековых перемещений степных народов на большие расстояния и дают возможность объединить генетические, исторические и археологические данные.

До прихода авар римляне занимали западную часть Карпатского бассейна, а сарматы – восточную (около 1-400 гг. н. э.). На смену римлянам пришла недолговечная империя гуннов (400–455 гг. н. э.), а также различные германоязычные группы: готы и лангобарды в Паннонии, а гепиды вдоль Тисы (400-568 гг. н. э.). В 567/68 году лангобарды разгромили гепидов и переселились в Италию, в то время как авары завоевали Карпатский бассейн и подчинили его местное население.
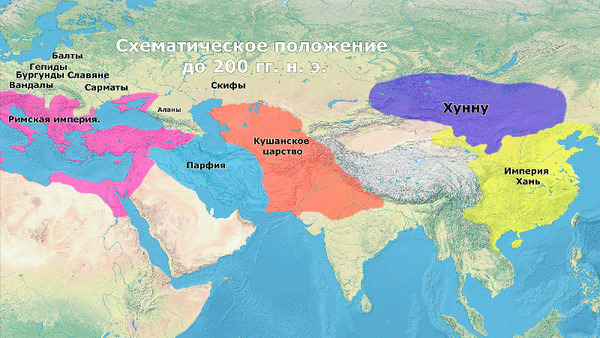
Данное исследование как раз и сосредоточено на этом важном изменении и его генетическом влиянии.
Предыдущие исследования с использованием однородительских маркеров предоставили наводящие на размышления генетические доказательства, кстати, обзор одного из таких исследований также есть на канале, но полногеномные данные для реконструкции происхождения населения аварского периода отсутствовали.
В новой работе авторы при помощи ядерной ДНК задались целью получить ответы на следующие вопросы:
1. Можно ли на основе их геномного профиля подтвердить происхождение основной группы авар из восточной части Центральной Азии?
2. Были ли элиты вновь прибывших степных воинов генетически однородными или имели смешанное происхождение?
и 3. Насколько элита авар отличается от предыдущего населения Карпатского бассейна?
Богатый археологический материал аварского периода в Карпатском бассейне (конец VI – начало IX вв. н. э.) включает около 600 поселений и около 100 тыс. раскопанных захоронений, которые демонстрируют большую неоднородность, особенно на ранних этапах.
Авторы новой работы получили полногеномные данные для 66 древних людей, охватывающих как доаварский (17 в сарматском и один в гуннском контексте, IV–V вв. н. э.), так и аварский периоды (VII–VIII вв. н. э.) из трех различных регионов:

25 человек из междуречья Дуная и Тисы, из них 8 были из высокостатусных мужских погребений (группа Бочи и Кунбабони), датируемых второй третью VII в. н. э., после неудавшейся осады Константинополя в 626 г. н. э., содержащих изысканное оружие и пояса, украшенные золотом и серебром, различные знаки отличия и другие предметы престижа с внутриазиатскими параллелями. Остальные 17 человек были взяты из контекстов с непосредственным контактом с этой элитной группой.
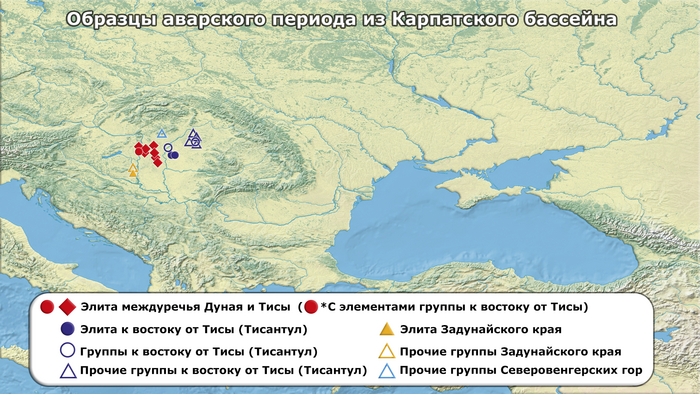
Авторы также обратили своё внимание на группу захоронений, найденных в основном к востоку от реки Тиса (Тисантуле), предполагаемого вторичного центра власти, которые имеют многочисленные культурные сходства с таковыми из восточноевропейских степей VI-VII вв. н. э. Для них характерны одиночные захоронения или небольшие могильники, захоронение частей или целых животных, а также могилы с уступами или концевыми нишами.
12 из 17 образцов региона принадлежат к этой “группе из Тисантулы”. Ещё 6 образцов аварского периода были из соседних регионов Карпатского бассейна и отражают неоднородность археологического материала и погребальных практик того времени, включая два богатых элитных захоронения из Задунайского края (Колькед), где археологические данные указывают на множество местных групп, демонстрирующих тесные связи со Средиземноморьем, а также с владениями Меровингов в Западной Европе.
Результаты
Геномная структура жителей доаварского периода
Анализ главных компонент демонстрирует заметные различия в генетических профилях между людьми доаварского и аварского периодов из Карпатского бассейна. За исключением единственных доступных двух геномов гуннского периода, все доаварские жители Карпатского бассейна попадают в рамки генетической изменчивости западноевразийцев.
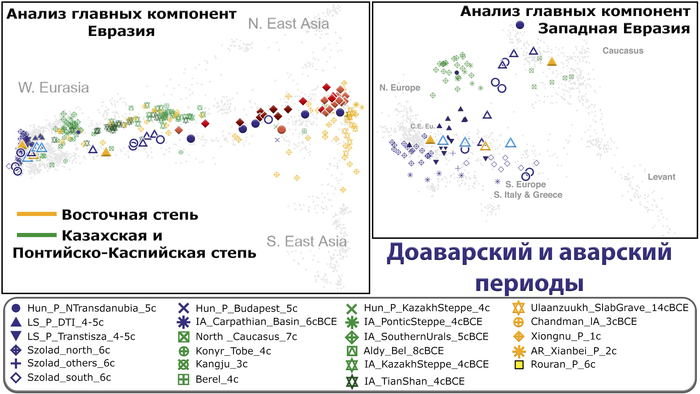
Они в основном пересекаются с современными жителями Центральной и Восточной Европы, хотя некоторые попадают в кластер к южным европейцам, особенно ранее описанная группа периода лангобардов, которая пересекается с современными южными итальянцами и греками.
Сравнение доступных древних популяций Карпатского бассейна показывает, что 16 недавно проанализированных людей позднесарматского/ гуннского периода IV-V веков н. э. близки, но всё же отличаются от обитателей этого же региона VIII-IV веков до н. э. и частично ближе к людям лангобардского периода VI века н. э.
Среди представителей позднесарматского периода люди из региона к востоку от Тисы (Тисантуле), пересекаются с отдельными представителями Солада VI века, а обитатели междуречья Дуная и Тисы пока по-прежнему близки к жителям позднесарматского периода из Тисантулы IV-V веков, но уже смещены в сторону представителя гуннского периода V века и групп железного века из степей. Что подтверждается и в моделях примесей.
Геномная структура жителей аварского периода
В отличие от предыдущих периодов, популяции аварского периода демонстрируют значительную генетическую изменчивость, поскольку отобранные образцы разбросаны по всей клине от популяций Западной Евразии до Северо-Восточной Азии.

Однако несмотря на эту общую неоднородность, существуют четкие закономерности генетической подструктуры, соответствующие как географии Карпатского бассейна, так и социально-археологическим категориям.
Все люди из элитных захоронений на участках междуречья Дуная и Тисы имеют профили предков из Северо-Восточной Азии, попадающие вдоль генетической линии современных популяций от Алтая до Монголии и бассейна Амура.
Они также в целом подпадают под изменчивость древних людей, связанных с некоторыми из основных археологических культурных горизонтов Восточной степи позднего железного века и раннего средневековья.
Среди них довольно крупная группа состоит из жителей периода хунну III века до н. э.-I века н. э. с Монгольского плато, которые в целом очень неоднородны и ранее были сгруппированы в три кластера на основе их генетических профилей. Другие образцы включают: жителей сяньбийского периода I-III вв. н. э. из Алтая и бассейна Амура; двух человек периода гуннов, одного представителя Карпатского бассейна начала V века, одного обитателя Казахской степи IV века и одного человека жужаньского периода VI века из современной Монголии.
Было определено, что все эти люди, хотя и в разной степени смешанные с другими популяциями, имели восточноевразийский компонент, который прослеживается до «древних северо-восточных азиатов», как неолитические охотники-собиратели пещеры Чёртовы ворота Приморья.
Генетический анализ не учитывает культурную принадлежность или хронологическую информацию, но, тем не менее, группирует представителей элиты аварского периода из междуречья Дуная и Тисы в соответствии с аварской хронологией (ранняя, средняя и поздняя). Все жители раннеаварского периода, за исключением одного младенца и человека, погребённого с типичными характеристиками группы к востоку от Тисы, образуют плотный кластер с высоким содержанием компонента «древних северо-восточных азиатов».
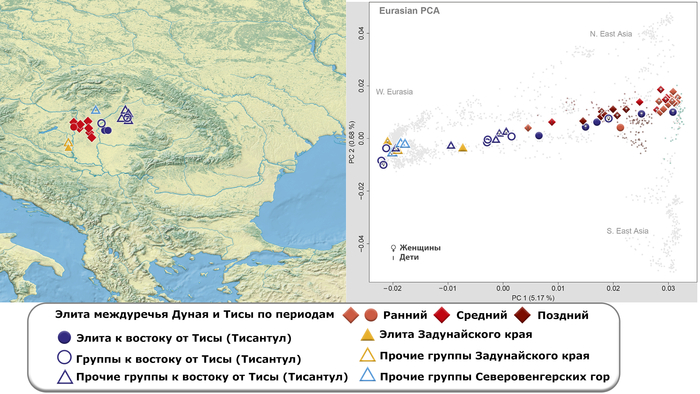
Они расположены между современными монголоязычными популяциями (например, бурятами с хамниганами) и нивхами с тунгусо-маньчжурскими народами (например, негидальцами, нанайцами, ульчами и, собственно, нивхами).

Помимо этого, представители раннеаварского периода близки к трём жителям сяньбийского времени из бассейна Амура. Трое из шести жителей среднеаварского периода междуречья Дуная и Тисы группируются с представителями раннеаварского периода из того же региона, двое других незначительно отклоняются от них в сторону Западной Евразии, а один из захороненных детей занимает промежуточное положение между западными и восточными популяциями Евразии. И наконец жители позднеаварского периода уже сильнее смещены от людей предыдущих периодов в сторону Западной Евразии.
Моделирование родословной из восточноазиатских степей у элит в основе Аварского каганата
Моделирование генетического происхождения, подтвердило, что представители аварской элиты междуречья Дуная и Тисы раннеаварского периода, совместно с тремя их коллегами среднеаварского периода, на 88–98% состоят из генетических компонентов как у древних северо-восточных азиатов.
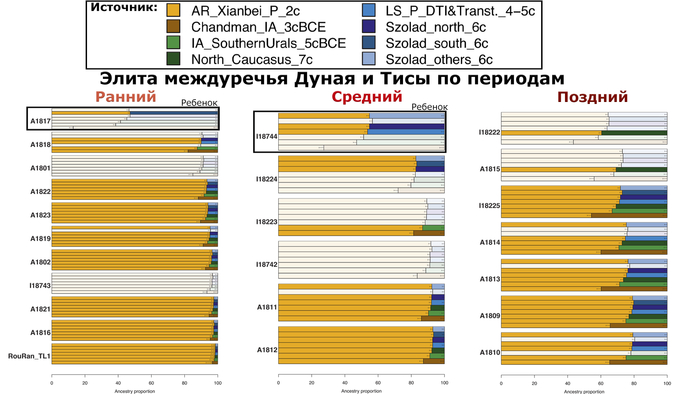
В то время как в позднеаварском периоде количество этой родословной сократилось уже до 70-80%. Остальная часть родословной происходит от источника, несущего более высокое западноевразийское происхождение. За исключением двух поздних обитателей междуречья Дуная и Тисы, носителями этого западноевразийского происхождения в равной степени могут быть как жители причерноморско-каспийских степей, так и популяции Карпатского бассейна IV-VI веков.
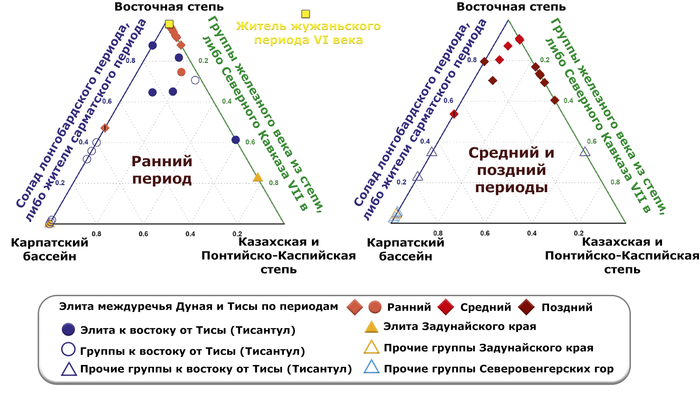
Однако противопоставляя попарные комбинации местных и неместных источников, авторы показали, что для большинства этих людей, неместный источник был предпочтительнее местного, для которого лучше всего подходили представители Северного Кавказа VII века или современные жители Северной Осетии.
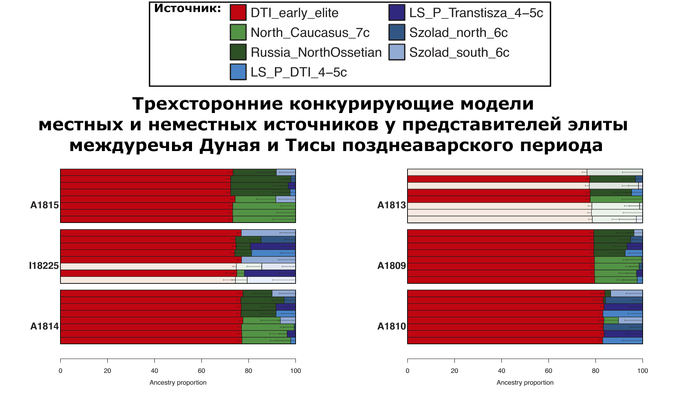
Предполагаемые даты примеси у элит междуречья Дуная и Тисы подтверждают различие между представителями ранне- и позднеаварского периодов: элиты раннего периода, а также три представителя среднего периода представляют более давние даты примеси, совместимые с датой, оцененной для жителя жужаньского периода VI века из современной Монголии, в то время как элиты позднеаварского периода и два человека среднего периода показывают значительно менее древние даты.
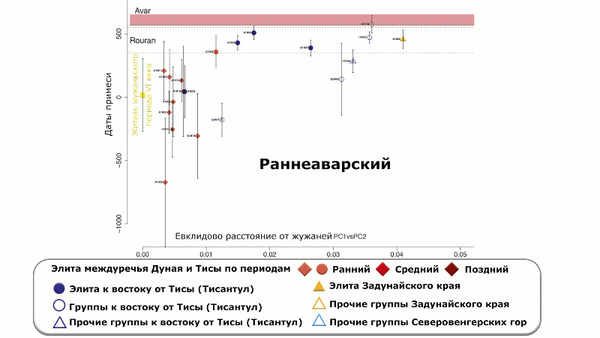
В целом даты примеси для отдельных представителей элиты раннего и среднего периодов варьируются от IV века до н. э. до III века н. э. А представители элиты позднеаварского периода, демонстрируют более поздние даты примеси, которые попадают в периоды жужань или ранних европейских авар, а точнее ближе к концу жужаньского периода в групповом анализе.
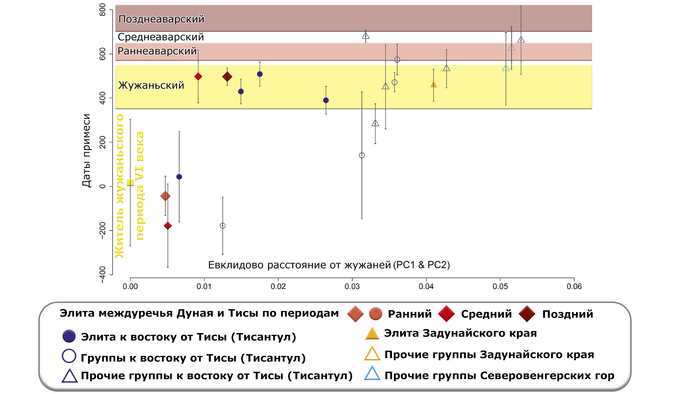
Стоит отметить, что представитель жужаньского периода VI века из современной Монголии моделируется в рамках тех же пропорций, как и представители раннеаварского периода и часть жителей среднеаварского периода.
Помимо этого, невзирая на значительное количество родственников в пределах одних и тех же участков среди аварской элиты междуречья Дуная и Тисы как в раннем, так и позднем периодах, у них не обнаружено следов близкородственных связей (т.е. инбридинга) в их недавнем прошлом, за исключением одного случая.

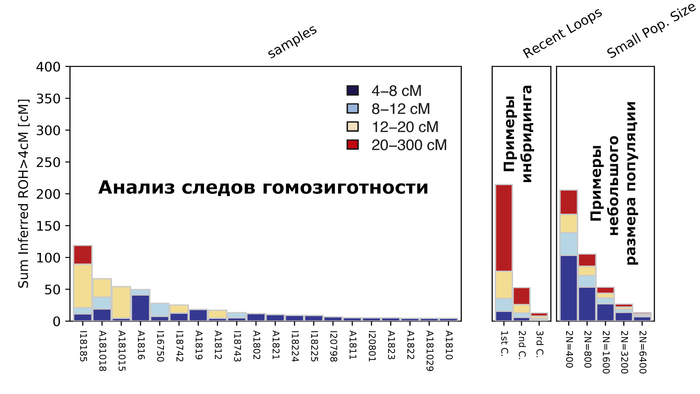
Тем не менее, восточноцентральноазиатская отцовская линия N-F4205, которая была обнаружена ранее как типичная для мужской части элит, захороненных в этом регионе, демонстрирует преемственность на протяжении всего среднего и позднего аварского периодов. Все 12 мужчин аварского периода из междуречья Дуная и Тисы несли линию N1a1a1a1a (N-F4218), и все, кроме одного, могли быть отнесены к подветви N-F4205, типичной для современных монгольских и забайкальских популяций. А вот разнообразие мтДНК значительно выше. Это, в сочетании с отсутствием геномных признаков инбридинга или довольно низким размером популяции, могло свидетельствовать о практике патрилокальности/патрилинейности, а также о более высокой женской экзогамии, которая помешала бы близкородственным связям.
Неоднородное происхождение в регионах, окружающих центр Аварского каганата
Остальные 23 человека, проанализированные в новом исследовании, происходят из соседних регионов, окружающих междуречье Дуная и Тисы, и по сравнению с элитами основного центра власти Аварского каганата, они генетически довольно разнообразны и на графике анализа главных компонент простираются от жителей предыдущих сарматского и лангобардского периодов до элит междуречья Дуная и Тисы раннеаварского периода с максимальным восточноазиатским происхождением.
Тем не менее, только некоторые люди, из захоронений к востоку от Тисы, несут более 50% предков, связанных с «древними северо-восточными азиатами» (7 из 13), несмотря на то, что все образцы, кроме одного, были отобраны на двух участках с захоронениями аварской элиты по археологическому контексту.
А вот у остальных представителей группы к востоку от Тисы примесь, связанная с «древними северо-восточными азиатами», слабая или практически не обнаруживается.
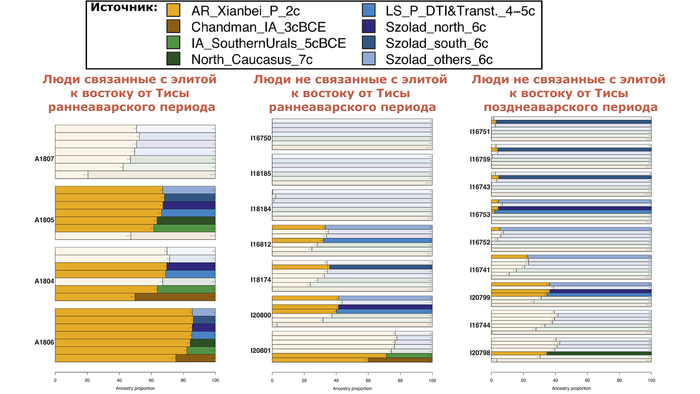
При этом двое из них, у которых почти полностью отсутствовали эти восточные компоненты, демонстрируют более высокую близость к средиземноморским популяциям, сгруппированным с современными сицилийцами и мальтийцами, и находятся в самом конце генетического кластера «юг Солада VI века» (Szolad_south_6c). Генетическая неоднородность представителей группы к востоку от Тисы примечательна тем, что эта популяция в основном приписывается мигрантам из причерноморских степей. Однако их неоднородность, вероятно, связана с разной степенью и временем примеси. Так, два человека с самой высокой долей «древних северо-восточных азиатов», неотличимой от элит раннеаварского периода междуречья Дуная и Тисы, имеют одинаково более древние даты примеси. В то время как остальные жители, с более западноевразийским происхождением, имеют значительно более недавние и более изменчивые даты примеси, вплоть до нескольких поколений ранее. А это говорит о том, что, несмотря на общепринятую культурную практику, на кладбищах такого типа хоронили людей с очень разным генетическим происхождением.
Интересным примером могут послужить мужчина и женщина раннеаварского периода из погребения на юге Задунайского края (Колькед), которые демонстрируют два совершенно разных генетических профиля. На графике анализа главных компонент женщина находится выше предшествующей условно местной популяции Карпатского бассейна и в моделях примеси её генофонд на 100% может быть смоделирован с их участием.
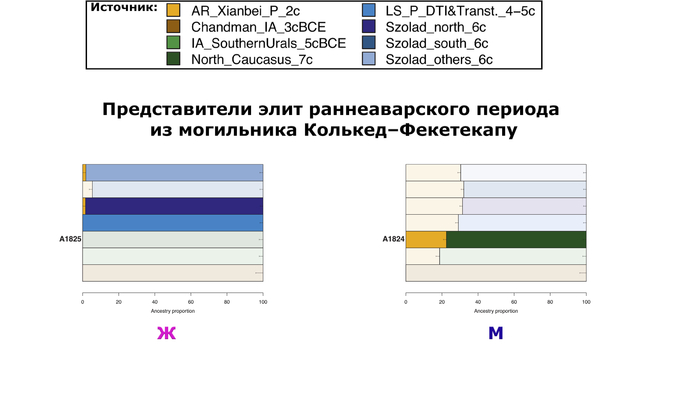
А вот мужчина имеет уникальную генетическую родословную по отношению ко всем другим людям, описанным на данный момент, демонстрируя как сдвиг в сторону Восточной Евразии, так и в сторону популяций, связанных с Ираном или Кавказом. Этот человек может быть смоделирован как смесь ~20% компонентов как у «древних северо-восточных азиатов» и ~80% как у представителей Северного Кавказа VII века или как у современных жителей Северной Осетии. Дата примеси около 5-го века н.э., полученная для этого человека, подтверждает его недавнее, неместное, смешанное происхождение.
Стоит отметить, что в этот диапазон генетической изменчивости также попадают люди, захороненные в могильнике Коныртобе IV века Отрарского района Туркестанской области Казахстана и представители государства Кангюй III века. А 9 человек позднеаварского периода демонстрируют уже от менее 40% до 5% примеси с людьми, связанными с «древними северо-восточными азиатами», в то время как основной компонент происхождения большинства из них схож с таковым у предшествующих древних популяций Карпатского бассейна, которые условно названы «местными».
Даты примесей подтверждают общую закономерность более поздних примесей, происходящих в основном на относительно ранних этапах существования Аварского каганата, хотя некоторые ещё более ранние даты, в V веке н. э., могут указывать на события примеси с восточной родословной ещё в гуннский период.
Также стоит отметить, что за исключением ранее описанных случаев с двумя детьми, поток генов от аварской элиты к людям местного происхождения не наделял их более высоким статусом и большинство людей получивших восточноазиатскую примесь в своём недавнем прошлом, были захоронены на неэлитных участках.
Итоги
Новые результаты подтверждают восточноазиатское происхождения ядра элиты Аварского каганата из районов междуречья Дуная и Тисы, указывая на наличие преобладающего количества родословной «древних северо-восточных азиатов» с середины трети VII в. н. э. до начала VIII в. н. э., в ранний и средний аварские периоды. Данные также указывают на поразительное генетическое сходство аварской элиты с представителем жужаньского периода, а также с древними жителями восточноазиатской степи периода хунну и особенно с сяньбийцами из бассейна Амура.
А вот представители ядра аварской элиты позднеаварского периода уже демонстрируют сдвиг в сторону более позднего, смешанного с западноевразийским источником, происхождения, примерно, на 20-30% их родословной, причём связанной не с местными жителями доаварского периода Карпатского бассейна, а с генофондом как в степях к северу от Кавказа VII века или как у современных жителей Северной Осетии. Хотя авторы отмечают, что скудность данных из этого региона первого тысячелетия н. э. не позволяет точно определить возможные источник этой примеси. Однако эта неместная примесь в сочетании с сохраняющимся, относительно высоким количеством восточного происхождения, связанного с «древними северо-восточными азиатами» и отсутствием близкородственных связей может указывать на продолжающуюся миграцию из степей после первоначального прибытия авар в Карпатский бассейн. Но в качестве альтернативы, эта примесь могла произойти и с группами более низкого статуса, не включенных в это исследование, которые, как утверждается, прибыли вместе с аварской элитой.
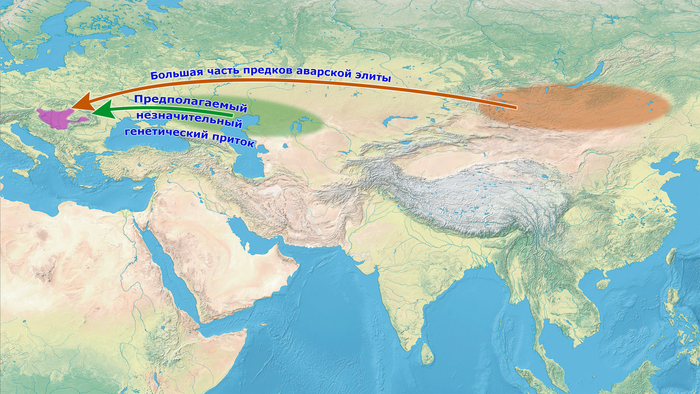
Проще говоря, по ходу миграций с востока, к аварам в районе Северного Кавказа могли прибиться попутчики, добровольно или нет, не известно... А потом по мере сокращения потенциальных пар, потомки попутчиков по статусу расценивались выше чем местное население Карпатского бассейна.
Высокий уровень восточного происхождения среди элит, сохранявшийся столетие и более после первоначальной миграции, может указывать на значительное количество прибывших, которые заключали браки между собой. Однако это также может указывать и на более поздний приток людей с востока и продолжающиеся контакты с Центральной Азией. А тот факт, что восточным профилем обладали как мужчины, так и женщины свидетельствует о том, что и мужчины, и женщины прибыли из Восточной Азии.
В отличие от общих однородных генетических профилей с высокой долей «древних северо-восточных азиатов» у элит междуречья Дуная и Тисы, люди из элитных захоронений в соседних регионах, гораздо более разнородны. И хотя элиты к востоку от Тисы, также демонстрируют самую высокую долю предков, связанных с «древними северо-восточными азиатами», более сложные модели примесей могут быть результатом различных временных этапов этих связей с восточной степью, возможно, восходящих к гуннскому периоду или более поздним перемещениям из степей в Карпатский бассейн. Особенно примечательны двое представителей элит из Задунайского края (Колькед), с очень разным генетическим происхождением.
Женщина имела несмешанный генетический профиль доаварского периода и похоронена с погребальными принадлежностями, связанными с позднеантичными, меровингскими и византийскими традициями. А генетическое происхождение мужчины указывает на степи к северу от Кавказа или другие регионы с преобладанием иранского генофонда. Эти данные указывают на появление генетически неоднородного слоя местной элиты под властью неместных аварских элит. При этом среди простых жителей раннего и позднего аварского периодов преобладал более западный генетический профиль, демонстрирующий сильную связь с населением Карпатского бассейна доаварского периода. При этом потомки этих людей после их связи с мигрантами, особенно в позднеаварский период, не получали какого-либо статуса. Поэтому авторы делают вывод о преимущественно однонаправленном потоке генов от элиты к местному населению.
Аварский каганат в своё время очень сильно повлиял на демографию и политику в Карпатском бассейне. При этом свидетельства подобных перемещений населения во время миграций гуннов (V века), лангобардов (VI века) или мадьяр (IX-X веков) гораздо менее убедительны. А по этим периодам и популяциям ещё остаются вопросы. И в отличие от этих пока ещё плохо генетически задокументированных периодов, новые результаты обеспечивают надежную поддержку происхождения элиты авар раннего и среднего периодов, в Северо-Восточной Азии.
Также новые результаты уверенно указывают на то, что в раннесредневековом Карпатском бассейне существовала выдающаяся генетическая изменчивость, охватывающая почти всю изменчивость современной Евразии и предоставляют четкие доказательства трансъевразийских миграций на большие расстояния.
Источник:
Ancient genomes reveal origin and rapid trans-Eurasian migration of 7th century Avar elites
Guido Alberto Gnecchi-Ruscone, Anna Szécsényi-Nagy, István Koncz, Zuzana Hofmanová, Choongwon Jeong, Johannes Krause et all doi.org/10.1016/j.cell.2022.03.007
Тибетское нагорье характеризуется низким атмосферным давлением, пересеченной местностью, а также относительно низкой биологической продуктивностью и температурой. Несмотря на эти ограничения, этнические тибетцы успешно адаптировались к этой среде и жили на плато в течение тысячелетий. Понимание их генетической и культурной адаптации к этой сложной среде, со сниженным содержанием кислорода, представляет большой интерес с точки зрения археологии, антропологии, генетики и физиологии. Но чтобы это понять, необходимо ответить на многие фундаментальные вопросы, касающиеся происхождения современных тибетцев, включая место и сроки формирования предкового современным тибетцам генофонда и первоначальные маршруты миграций людей на Тибетское нагорье.
Хотя археологические данные, относящиеся к ранним миграциям на Тибетское нагорье, скудны, карстовая пещера Байшия (3280 м над уровнем моря) на его крайнем северо-восточном краю, предполагает присутствие древних людей, связанных с денисовцами, между 160 и 60 тысячами лет назад.

Датировка на археологическом участке Нвия Деву в центральной части нагорья (4600 м над уровнем моря) предполагает присутствие современного человека между 30 и 40 тыс. л. н. Однако это признаки присутствия, а не постоянного проживания, к которым некоторые авторы относят следы охотников и собирателей в Чусанге (4270 м над уровнем моря) 12700 - 7400 лет назад. И то не все учёные с этим согласны, некоторые из них утверждали, что постоянное заселение центральной части нагорья было невозможным до появления земледелия на основе ячменя около 3,6 тыс. лет назад. Последняя модель предполагает, что сельское хозяйство было привнесено на Тибетское нагорье мигрантами с более низких участков (<2500 м над уровнем моря) вдоль северо-восточных окраин нагорья, которые также внесли существенный вклад в генофонд современных тибетцев. Однако существуют доказательства более сложного, множественного происхождения современных тибетцев, которые подтверждаются и генетическими данными.
Плотно отобранные однородительские маркеры Y-хромосомы и митохондриальной ДНК, можно проследить по большей части до линий, присутствующих в северной части Восточной Азии с раннего голоцена, но более старые гаплогруппы, такие как митохондриальная M16 и Y-хромосомная D-M174, происходящие из глубокой евразийской линии, также уникально присутствуют среди современных тибетцев.
Идея древнего палеолитического вклада в тибетский генофонд также была предложена на основе данных о последовательности всего генома. Исследование, сравнивающее современные тибетские геномы с геномами древних сибиряков и архаичных представителей рода людей, выявило вклад смеси древних предков - архаичных и неархаичных — среди предполагаемых ранних обитателей Тибетского нагорья. Это предложение согласуется с наличием у тибетцев специфических аллелей гена EPAS1, которые достались им от денисовцев.
Суммарно, современные генетические данные свидетельствуют о нескольких стадиях заселении нагорья. За популяциями эпохи плейстоцена, с некоторым уровнем архаичной примеси, последовали миграции эпохи голоцена с северо-восточных окраин Тибетского нагорья.

И хотя идентичность и происхождение населения эпохи плейстоцена остаются неизвестными, недавний анализ выявил четкую клину генетической изменчивости с востока на запад в пределах современных географически рассредоточенных тибетских популяций. Эта клина может отражать неолитические миграции людей, занимавшихся выращиванием ячменя.
До заселения Тибетского нагорья выращивание ячменя практиковали популяции позднего неолита и раннего бронзового века из районов современных провинций Ганьсу и Цинхай, как к примеру, представители культуры Цицзя (около 2300-1800 гг. до н.э.).
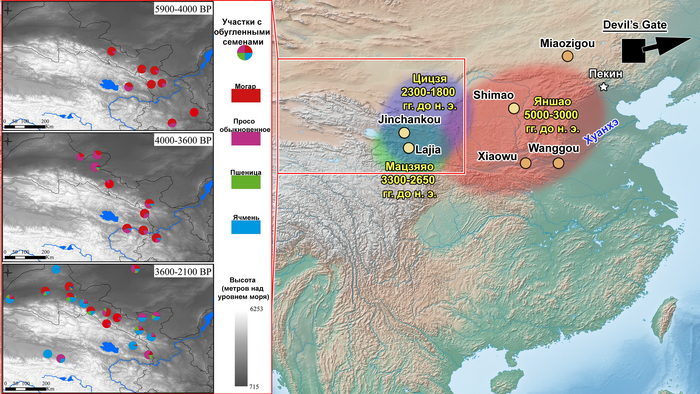
Однако клина также могла быть сформирована или усилена более поздними историческими событиями, такими как расширение Тибетской империи с VII века н. э., или в результате длительного процесса потока генов между соседними популяциями с изоляцией расстоянием, не связанного с дальними миграциями.
Предыдущие исследования древней ДНК людей из трех высокогорных гималайских поселений в непальском районе Мустанг датируемых 800 г. до н. э. – 650 г. н. э., показали, что эти места были заселены популяциями с явным восточноазиатским происхождением, которые, вероятно, мигрировали с Тибетского нагорья. Авторы новой работы дополняют эти данные, увеличивая временной охват более чем на 600 лет, примерно с 1420 г. до н. э. по 650 г. н. э., и предоставляя самые ранние генетические данные на сегодняшний день для популяций нагорья.
Работа демонстрирует, что эти древние гималайские популяции генетически группируются с современными тибетцами и что они представляют собой раннюю ветвь тибетской линии, что делает их особенно информативными для определения истории тибетского генофонда, его происхождения и его нынешнего распределения среди современных тибетцев, и их соседей.
А теперь обо всём более подробно:
Результаты
Древние образцы из Гималаев
В новом исследовании авторы проанализировали общегеномные данные 38 древних людей из семи участков непальских районов Мананг и Мустанг, а именно: Suila (n = 1; 1494-1317 гг. до н.э.), Lubrak (n = 2; 1269-1123 гг. до н. э.), Chokhopani (n = 3; 801-770 гг. до н. э.), Rhirhi (n = 4; 805-767 гг. до н. э.), Kyang (n = 7; 695-206 гг. до н. э.), Mebrak (n = 9; 500 г. до н. э. –1 г. н. э.) и Samdzong (n = 12; 450-650 гг. н. э.). Новые образцы, анализировались совместно с ранее опубликованными древними геномами, а также с геномами современных шерпов и тибетцев Непала.
Генетическая структура жителей высокогорья Восточной Азии и их соседей
На графике анализа главных компонент древние жители районов Мананг и Мустанг Непала группируются с другими популяциями из Восточной Азии.
Как видно ниже на графике, современные популяции образуют структуру с тремя отростками.
Первый состоит из жителей Южного Китая и Юго-Восточной Азии, второй из представителей Северо-Восточной Азии и третий, куда собственно и попадают древние образцы, включает тибето-бирманские популяции и соответствует генетической линии современных тибетцев с востока на запад. А также они разделяют уникальные наследственные компоненты с современными популяциями из высокогорных районов и средней высоты.

Древние жители Непала, в первую очередь, генетически схожи друг с другом, а после с шерпами и тибетцами, за которыми следуют носители тибето-бирманских языков с более низких высот, такие как наси, и или йи, а также нага из Нагаленда в Индии. Однородительские маркеры, в виде гаплогрупп, подтверждают их тесное генетическое родство с современными шерпами/тибетцами.
13 из 14 мужчин имели производные маркеры Y-гаплогруппы O-M117, а 12 мужчин несли производные маркеры ее субклада Oα1c1b-CTS5308.
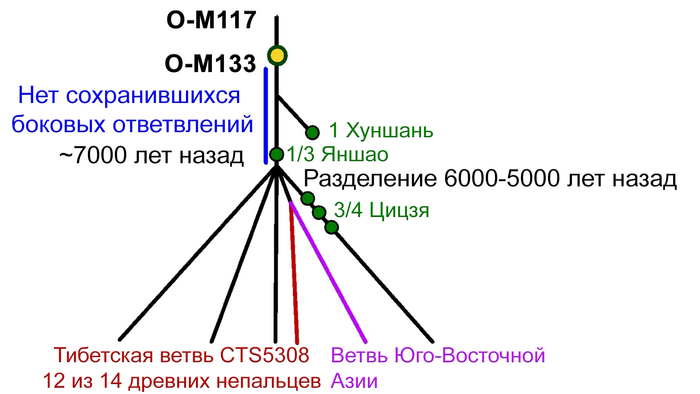
Среди современных популяций этот субклад встречается в основном среди тибетцев и шерпов Тибетского нагорья, в отличие от его родственного субклада Oα1c1b-Z25929, который сегодня встречается в основном в Южном Китае и Северо-Восточной Индии.
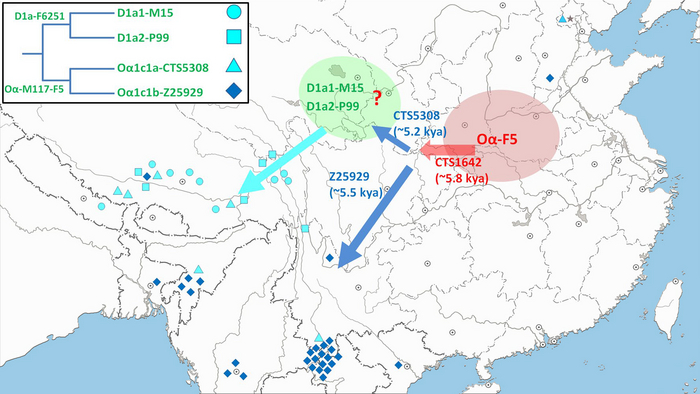
Быстрое распространение всех существующих линий O-M117, по оценкам, произошло 7-5 тыс. лет назад и было интерпретировано как отражение распространения сино-тибетских языков, вероятно происходящих из Северного Китая. Примечательно, что Y-гаплогруппа O-M117 была также обнаружена и у древних людей из неолитической культуры Яншао верхнего течения Хуанхэ и поздненеолитической Цицзя, а это свидетельствует о том, что большинство мужских линий древних жителей районов Мананг и Мустанг современного Непала восходит к региону распространения вышеупомянутых культур. Также у одного из древних непальцев была определена гаплогруппа - D1a, которая на сегодня является еще одной распространенной гаплогруппой на Тибетском нагорье.
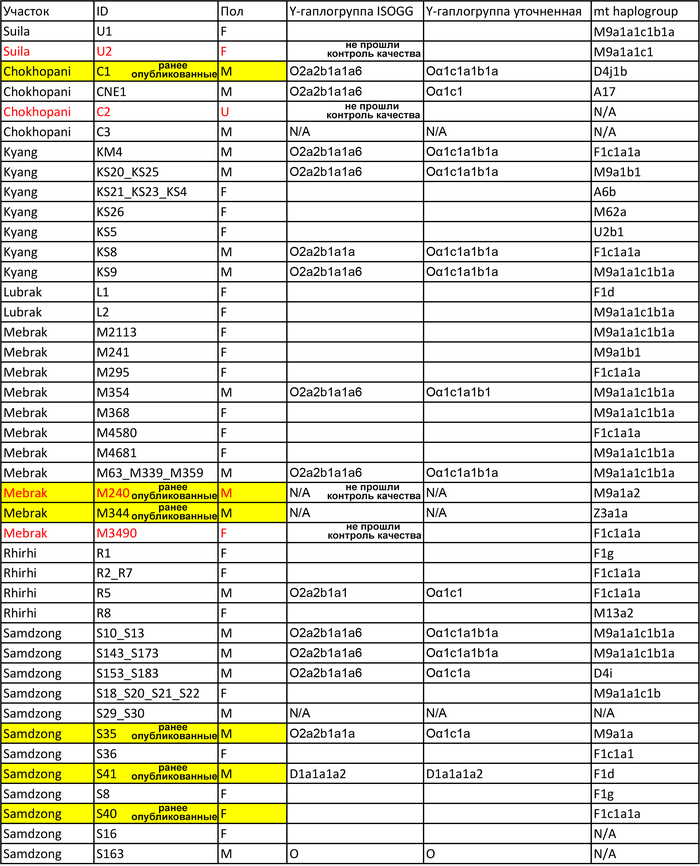
Митохондриальные линии исследованных древних людей, также распространены среди современных тибетцев, хотя они и более разнообразны.
Генетическая связь между древними и современными жителями Тибетского нагорья
Современные и древние группы районов Мананг и Мустанг, а также близлежащих районов Горкха и Солукхумбу вместе с тибетцами из более отдаленных мест, генетически ближе всего к представителям Lubrak, а затем друг к другу среди древних и современных жителей Восточной Азии. А самые древние представители из Suila, а также более поздние группы близки к современным шерпам и тибетцам в отличие от представителей Chokhopani, предки которых смешались с жителями более низких высот, вероятно ещё до 1500 г. до н. э. Таким образом, авторы делают вывод, что люди из участков Lubrak и Suila на сегодняшний день являются самыми ранними известными носителями генофонда, наиболее распространённого у высокогорных популяций Тибетского нагорья и Гималаев и называют его “тибетской” линией в этом исследовании.
Двойное генетическое происхождение жителей высокогорных районов Восточной Азии
Археологические данные свидетельствуют о том, что неолитическое население в верхнем и среднем течении реки Хуанхэ оказало значительное культурное влияние на распространение земледелия на Тибетское нагорье. Этот регион также был предложен в качестве вероятной родины сино-тибетской языковой семьи.
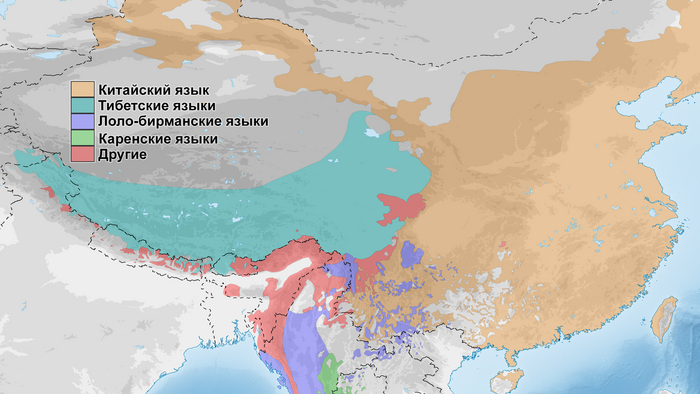
Распределение более крупных ветвей сино-тибетской языковой семьиИнтересно, что среди древних жителей низменностей Восточной Азии, группы среднего/позднего неолита из верхнего течения реки Хуанхэ и ее периферии демонстрируют наиболее близкое генетическое родство с группами из районов Мананг и Мустанг. Эти группы включают представителей поздненеолитических стоянок Цзиньшанькоу и Лацзя в верховьях реки Хуанхэ, принадлежащих к культуре Цицзя (около 2300-1800 гг. до н.э.; Upper_YR_LN), жителей стоянки Симао в Шенгедаляне провинции Шэньси (около 2250-1950 гг. до н.э.; Shimao_LN), а также представителей стоянок среднего неолита Мяодигоу во Внутренней Монголии (около 3550-3050 гг. до н.э.; Miaozigou_MN). Эти три группы имеют схожий генетический профиль, происходящий примерно на 80% от представителей среднего неолита культуры яншао участков Вангоу и Сяоу на Центральной равнине (около 4000-3000 гг. до н.э.; YR_MN) и на ~ 20% от древних северовосточных азиатов (ANA), как охотники-собиратели эпохи неолита пещеры Чёртовы ворота Приморья (“DevilsCave_EN”). Также авторы отмечают, что генетический обмен с жителями низин, вероятно, произошел до распространения ячменя.
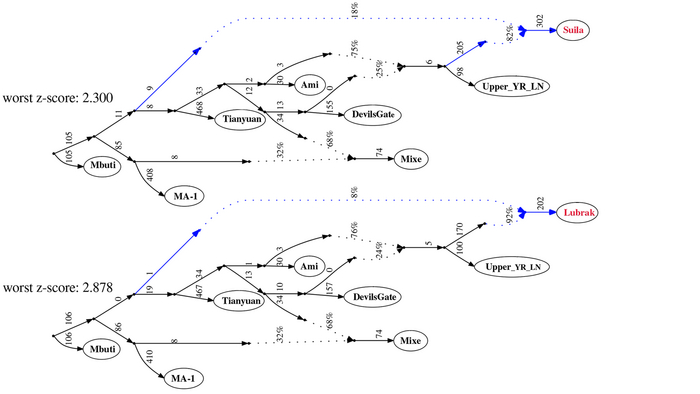
Несмотря на то, что 82-92% предков древних жителей из районов Мананг и Мустанг происходят от линии, связанной с земледельцами поздненеолитических стоянок в верховьях реки Хуанхэ, принадлежащих к культуре Цицзя, они всё же моделируются как сестринская к ним группа, как собственно и современные шерпы/тибетцы, благодаря своим уникальным генетическим компонентам, включающим аллель EPAS1 от людей в своё время связанных с денисовцами, который помогает адаптироваться к высокогорной среде.
Скорее всего, оставшиеся от 8 до 18% их предков происходят из древней популяции ещё до разделения между ветвями Западной и Восточной Евразии. При этом этот поток генов, не происходит от архаичных людей, таких как неандертальцы или денисовцы, которые вносят <0,5% родословной.
Также новые результаты отвергают и другие ранее предложенные источники потока генов в тибетскую родословную, включая глубоко разветвленные восточноевразийские линии, такие как южносибирский усть-ишимский человек, возрастом 45 тыс. лет, 40- тысячелетний древний житель Северного Китая – Тяньюань и родственные линии Хоа-Бинь/Онге в Юго-Восточной Азии.
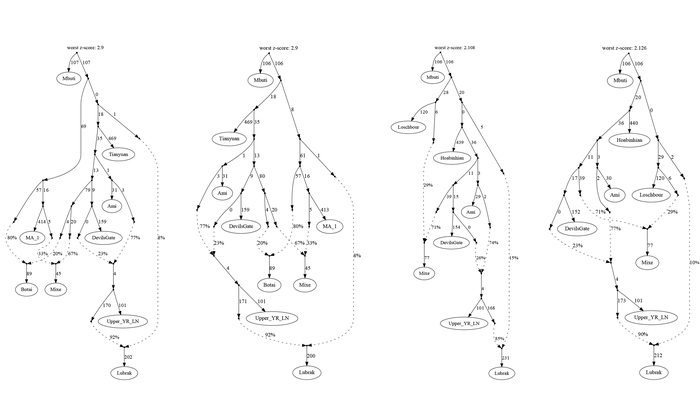
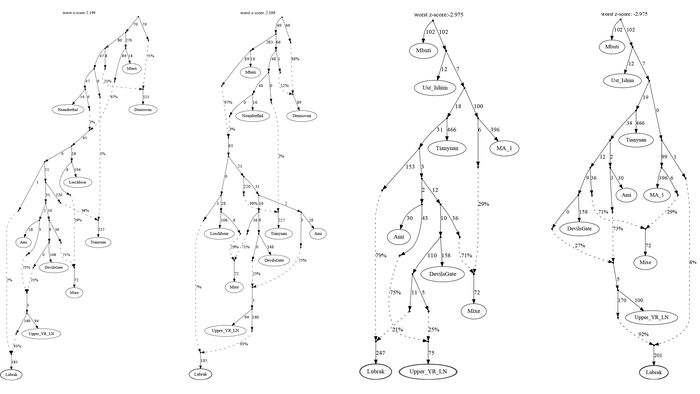
Предполагая, что эта глубокая евразийская родословная, вероятно, представляет собой ещё неисследованный палеолитический генетический субстрат от древних жителей Тибетского нагорья.
Два пути в Гималаи для носителей тибето-бирманских языков
На южных склонах Гималаев обитает множество этнолингвистических групп, которые демонстрируют поразительную структуру расслоения по высотам: индоираноязычное население Южной Азии занимает низменности, шерпы/тибетцы занимают высокогорья, а различные нетибетские народы, такие как таманги и гурунги, говорящие на тибето-бирманском языке, занимают диапазон средних высот.
В то время как шерпы/тибетцы Непала, вероятно, прибыли в Гималаи с нагорья (т.е. северным путём), предыдущее генетическое исследование предложило отдельный южный маршрут для миграции групп тибето-бирманцев средних высот.

Однако, как нетибетские группы, говорящие на тибето-бирманских языках, связаны друг с другом и с тибетской родословной, оставалось неясным. В данной работе авторы указывают на две генетические клины: тибетскую и нетибетскую тибето-бирманскую.
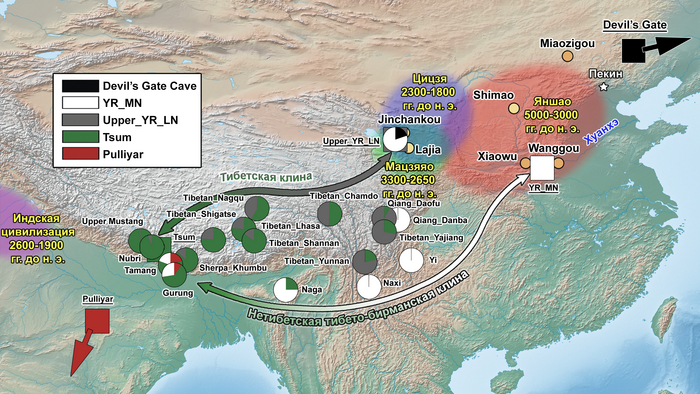
Тибетская клина соединяет древних и современных непальских тибетцев (которые в моделях представлены жителями Цумской долины) с обитателями поздненеолитических стоянок культуры Цицзя в верховьях реки Хуанхэ (Upper_YR_LN). А нетибетская тибето-бирманская клина отражает примесь вдоль маршрута вокруг Тибетского нагорья и соединяет популяции средних высот, такие как наси, йи, нага, таманги и гурунги с представителями среднего неолита культуры Яншао на Центральной равнине (YR_MN). Помимо этого, таманги и гурунги, имея более высокий вклад от тибетской линии 60-63%, дополнительно демонстрируют генетическую примесь из Южной Азии от 9 до 19%, как и популяции Бутана из предыдущих исследований.
Примечательно, что для населения Непала со значительным южноазиатским вкладом (например, барамцев, чантьяльцев, гурунгов и представителей народа чепанг), южноиндийские группы, такие как пулаяр, лучше подходят для моделирования их южноазиатского происхождения, чем соседние группы Северной Индии. Эти результаты подчеркивают сложную и многослойную историю смешения тибето-бирманских популяций в Гималаях.
Длительный положительный отбор на участках генов EPAS1 и EGLN1 у тибетцев
В своём предыдущем исследовании авторы сообщали, что производные аллели для положительно отбора в гене EPAS1 наблюдались только в менее древних образцах участка Самдзонг, в отличие от более древних жителей Чохопани и Мебрака. Проанализировав новые геномы древних жителей из районов Мананг и Мустанг, авторы по-прежнему не обнаружили производных аллелей в блоке гаплотипов EPAS1 в образцах из Чохопани и Суила, однако они наблюдались с промежуточной частотой на 5 других участках (от 25 до 58%).
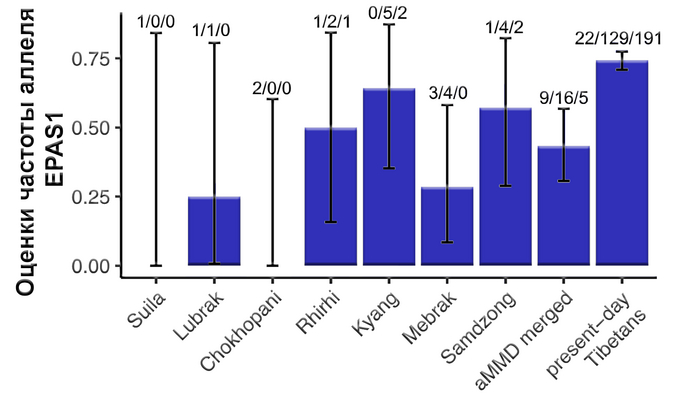
Интересно, что частота производных аллелей в древних образцах в целом ниже, чем у современных тибетцев (75%), а это указывает на то, что отбор все еще воздействовал на эти аллели в недавнем прошлом.
Итоги
В новой работе генетические данные 38 древних гималайцев указывают на то, что родословная, обнаруженная у современных тибетцев и шерпов, уже присутствовала у жителей высокогорья к 1494-1317 годам до нашей эры, отчётливо отличая их от жителей низменностей. Тем самым делая старше свидетельства существования тибетского генофонда по крайней мере на 500 лет по сравнению с предыдущими данными из Чохопани.
При этом исследование демонстрирует, что тибетский генофонд хорошо моделируется как смесь двух источников генетической родословной. Один из источников на 82-92% состоит из генетических компонентов связанных с жителями низменностей, которые проживали на северо-восточной окраине Тибетского нагорья во времена позднего неолита. А другой источник представляет собой древний и ранее неизученный палеолитический субстрат на долю которого приходится до 18%. Палеолитический субстрат, по-видимому, внес свой вклад исключительно в тибетский генофонд среди современных популяций, изученных на сегодняшний день.
Моделирование современных тибетских популяций и их нетибетских соседей, носителей тибето-бирманских языков, демонстрирует две генетические линии, которые, по-видимому, отражают два различных маршрута миграций – северный и южный, как видно на карте. Тибетские популяции смешаны вдоль северной клины, а генетическое, культурное и языковое разнообразие современных носителей тибето-бирманских языков вдоль южного склона Гималаев отражает слияние древних популяций, прибывших по этим двум маршрутам после их разделения со времен позднего неолита.
В отличие от предыдущих, весьма противоречивых работ, основанных на анализе ДНК современных тибетцев и ханьцев, результаты новой работы, благодаря древним геномам, показывают, что предки современных тибетцев присутствовали в Гималаях, по крайней мере, с 1420 года до н.э., во времена, когда на участках Суила и Лубрак появляются самые ранние прямые свидетельства постоянного присутствия человека. Помимо этого, исследование подтверждает тесную связь между ранними гималайскими популяциями и группами позднего неолита, жившими вдоль северо-восточной окраины Тибетского нагорья около 2300-1800 гг. до н.э. (Upper_YR_LN).
Эти неолитические группы с территорий современных провинций Ганьсу и Цинхай, вероятно, включали в себя популяции, которые позже заселили нагорье, однако точные даты этого события ещё не ясны.
Долгое время считалось, что выращивание ячменя, который лучше, чем просо подходит для более прохладного и сухого климата нагорья, способствовало неолитической экспансии в регион. Хотя новые результаты могут предположительно соответствовать давней гипотезе о распространении ячменя по нагорью около 1650 г. до н.э., такая массовая демографическая диффузия из районов Ганьсу и Цинхай всего за 200 лет вряд ли будет единственным объяснением древней генетической связи между Тибетским нагорьем и этими районами. Поэтому авторы предлагают альтернативный сценарий, в котором генетическая связь между популяциями нагорья и низменностей могла сформироваться намного раньше и, следовательно, не могла быть связана с внедрением ячменя или других одомашненных растений или животных западноевразийского происхождения.
Стоянка Karou в восточном Тибете (около 5000-3000 лет назад) и стоянка культуры Qugong близ Лхасы (около 3800-3000 лет назад) демонстрируют местную археологическую традицию и имеют композицию сборки и керамические мотивы, отличные от таковых в культуре Цицзя.

Karou, Qugong, ZongriКроме того, свидетельства с участка Zongri (около 2600-2000 гг. до н. э.) предполагает, что охотники-собиратели Тибетского нагорья обменивались просом с жителями равнин намного раньше предполагаемого появления ячменя.
А отсутствие следов отбора EGLN1 в верховьях реки Хуанхэ, у людей, принадлежащих к культуре Цицзя (Upper_YR_LN) в сочетании с предполагаемой селективной разверткой EGLN1, примерно 8000 назад, предполагает, что две популяции, возможно, уже разделились задолго до прибытия ячменя в район Ганьсу и Цинхай.
Ячмень выращивался как второстепенная культура в этих районах еще ок. 2000 г. до н.э., оставляя открытой возможность более ранней экспансии людей, выращивающих ячмень, на Тибетское нагорье ещё до 1650 г. до н.э., но археологических свидетельств, подтверждающих такой сценарий, не хватает.
Авторы отмечают, что их нынешние данные не могут полностью отвергнуть гипотезу, основанную на распространении ячменя, поэтому они призывают к поиску более древних геномов с Тибетского нагорья, а именно старше 1650 года до н. э., чтобы непосредственно проверить гипотезу.
Наконец, новое исследование показывает длительное воздействие естественного отбора на формирование генофонда высокогорных жителей Восточной Азии. Следует отметить, что увеличение частоты аллеля EPAS1 за период времени, от древних жителей из районов Мананг и Мустанг до современных тибетцев, подчеркивает медленное, но устойчивое действие положительного отбора на этот генетический вариант денисовского происхождения.
Будущие исследования позволят лучше понять популяционную историю этого региона, хотя и не только этого!
Источник:
Liu, CC., Witonsky, D., Gosling, A. et al. Ancient genomes from the Himalayas illuminate the genetic history of Tibetans and their Tibeto-Burman speaking neighbors. Nat Commun 13, 1203 (2022). doi.org/10.1038/s41467-022-28827-2
Вспомогательные источники: Gnecchi-Ruscone, G.A., Jeong, C., De Fanti, S. et al. The genomic landscape of Nepalese Tibeto-Burmans reveals new insights into the recent peopling of Southern Himalayas. Sci Rep 7, 15512 (2017). doi.org/10.1038/s41598-017-15862-z
Ren L, Yang Y, Wang Q, et al. The transformation of cropping patterns from Late Neolithic to Early Iron Age (5900–2100 BP) in the Gansu–Qinghai region of northwest China. The Holocene. 2021;31(2):183-193. doi.org/10.1177/0959683620941137
Jeong C, Peter BM, Basnyat B, Neupane M, Beall CM, Childs G, et al. (2017) A longitudinal cline characterizes the genetic structure of human populations in the Tibetan plateau. PLoS ONE 12(4): e0175885. doi.org/10.1371/journal.pone.0175885
Ancestral Origins and Genetic History of Tibetan Highlanders Dongsheng Lu, Haiyi Lou, Kai Yuan, Bing Su, Longli Kang, Shuhua Xu doi.org/10.1016/j.ajhg.2016.07.002
Genetic Evidence of Paleolithic Colonization and Neolithic Expansion of Modern Humans on the Tibetan Plateau, Molecular Biology and Evolution, Volume 30, Issue 8, August 2013, Pages 1761–1778, Xuebin Qi, Chaoying Cui, Yi Peng, Xiaoming Zhang, Zhaohui Yang, Hua Zhong, Hui Zhang, et. all doi.org/10.1093/molbev/mst093
Население Центральной Азии, проживавшее вдоль Великого Шелкового пути, весьма разнообразно, оно подверглось генетическому и культурному влиянию тысячелетнего взаимодействия между Востоком и Западом.
Генетические исследования останков бронзового века (3100-1300 гг. до н.э.) из Центральной Азии показывают существенные временные изменения в генетическом составе популяций, указывающие на обширные миграции и экспансию с запада на восток оседлых скотоводов из западных степей, которые сформировали однородный генофонд к концу второго тысячелетия до н.э. В железном веке (1300-900 гг. до н.э.) кочевые скотоводы распространились по евразийской степи, рассеяв скифскую культуру. Анализ древней ДНК из погребений саков и сарматов, принадлежащих к скифской культуре, демонстрирует увеличение иранского и восточноевразийского генетического притока в южных и восточных образцах соответственно. В течение первого тысячелетия нашей эры на территории современного Казахстана образовались многочисленные конфедерации и империи, которые были связаны со значительным потоком генов. Например, ориентированная на мужчин экспансия кочевников хунну на запад из восточной степи привела к значительной примеси восточноевразийских линий к центральным сакам и вытеснению индоевропейских народов, известных как кангюи и усуни.
Впоследствии сформировались и смешались друг с другом разнообразные тюркские кочевые государства, что привело к потокам генов между разнородными популяциями бывшей Империи гуннов. После монгольского нашествия на территорию в 1211 году была создана Золотая Орда, которая в последующие века подверглась ряду раздроблений, что привело к созданию Казахского ханства (1465–1847). В течение этого времени кочевые племена разного происхождения проживали по всей территории современного Казахстана, и в конечном итоге они были организованы в три жуза или социально-территориальные объединения, основанные в преимущественно по географическому признаку: Старший жуз, Средний жуз и Младший жуз.
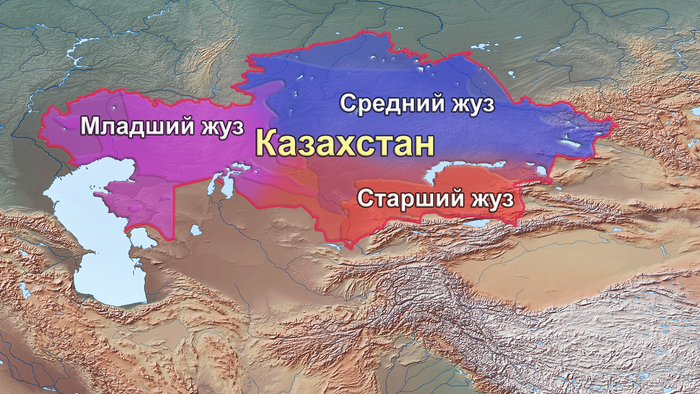
Кочевое общество казахской степи было организовано на основе иерархической патрилинейной клановой системы генеалогических линий. Люди одной и той же генеалогической линии утверждают, что имеют общего предка, и несколько генеалогических линий объединяются в рода, которые в совокупности образуют племена.
12 племен Старшего жуза в основном занимали Южный и Юго-Восточный Казахстан, семь племен Среднего жуза проживают в Восточном, Северном и Центральном Казахстане, в то время как три племени Младшего жуза традиционно проживали в Западном Казахстане. Некоторые степные рода не входили в состав жузов, особенно духовенство (кожа и сунак) и аристократия (торе́). Представители родов кожа и сунак связывают свою родословную с исламскими миссионерами, происходившими от родственников пророка Мухаммеда по отцовской линии. Люди Торе утверждают, что являются прямыми потомками Чингисхана. В отличие от оседлого земледельческого населения Центральной Азии, казахи практиковали экзогамные браки в пределах семи поколений, а партнер выбирался из другого рода, при этом женщины интегрировались в род своего мужа.
Несмотря на глобализацию последних веков и переход к оседлому образу жизни, родоплеменная структура казахов сохранилась, и многие современные казахи знают племенную принадлежность и историю своего рода. При этом учитывая, что родоплеменная принадлежность передаётся по мужской линии, анализ генетических маркеров Y-хромосомы может улучшить наше понимание мелкомасштабной демографической динамики популяции. Стоит отметить, что в случае с казахами, гаплогруппы используются для конкретных задач происхождения родов, а не для общей популяционной истории. Потому как по всему геному, как и по составу предков, казахи из всех жузов практически не отличаются ведь в основе происхождения казахов лежит сложный процесс смешения популяций Евразии, включая различные западные, южные и восточные группы, говорившие на разных языках.
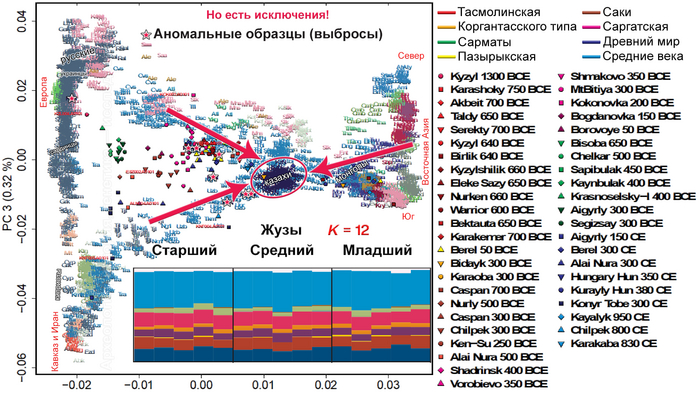
В очередной работе, авторы представляют результаты исследования 1171 гаплотипа Y-хромосомы среди всех существующих казахских племен, включая торе и кожу, с целью оценки взаимосвязи племен и жузов среди казахского народа современного Казахстана.
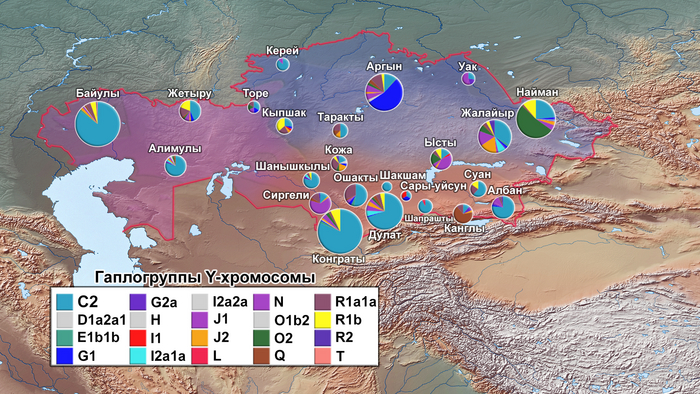
Стоит отметить, что ожидаемые расширенные данные по среднему жузу авторы обещают опубликовать позже.
Результаты
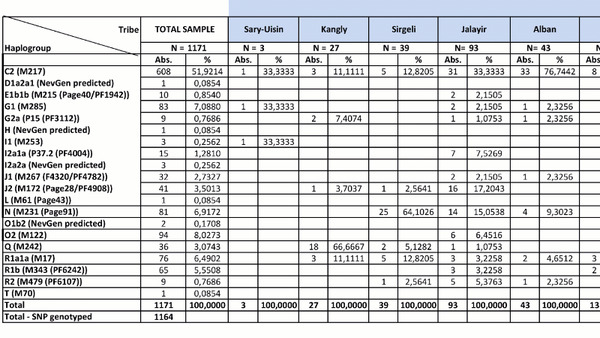
Наиболее частой гаплогруппой в казахском населении была C2-M217 (51,9%). Эта гаплогруппа присутствовала во всех исследованных племенах, и ее частота варьировалась от 11% у канглов и аргынов до 100% у шакшамов (но от них в анализе было всего 6 образцов). Стоит отметить, что количество информации по целому геному и по отдельным маркерам, значительно отличается.

Другой важный компонент казахского генофонда представлен гаплогруппой R (12,8%), которая имеет три субклада: R1a1a-M17 (6,5%), R1b-M343 (5,6%) и R2-M479 (0,8%). Гаплогруппа R1a1a-M17 наблюдалась в 18 из 24 племен и наиболее часто встречалась у представителей племени ошакты (31%), и так называемых людей белой кости из рода кожа (31,3%). Субклад R1b-M343 был обнаружен в 12 казахских племенах и имел самую высокую частоту у кыпшаков — 36,8%. Наконец, субклад R2-M479 наблюдался в пяти племенах с низкой частотой и был наиболее распространен у представителей рода кожа (6,3%) и племени жалайыров (5,4%).
Гаплогруппы O (представленные O2-M122 и O1b2, последняя предсказана проектом НевГен), G (G1-M285 и G2a-P15), N-M231, J (J1-M267 и J2-M172) и Q-M242 наблюдались с частотами <10% в разных племенах, но были обнаружены с более высокими частотами в одном или нескольких племенах. Например, гаплогруппа O2-M122 в целом в отобранных образцах встречалась с частотой 8,03%, но при этом была обнаружена у 52,3% найманов. Гаплогруппа G в общем встречалась с частотой 7,9%, причем большинство (7,1%) принадлежало к линии G1-M285, а 0,8% к G2a-P15, но значительная разница была между племенами, и ≈54% мужчин из племени аргын имели гаплогруппу G1-M285. Наибольшая частота гаплогруппы G2a-P15 наблюдалась в племени уак (18,2%). Гаплогруппа N-M231 имела глобальную частоту ≈6,9%, но была распространена в племенах сиргели (64,1%), уак (45,5%), племени торе (16,7%) и племени жалайыр (15,1%). Гаплогруппа J, представленная субкладами J1-M267 и J2-M172, имела глобальную частоту ≈6,2%, часто встречаясь в племени ысты (J1-M267 (39,4%) и J2-M172 (3%). Кроме того, J2-M172 также обнаружена у рода кожа (18,8%).
Наконец, гаплогруппа Q имела низкую общую частоту ≈3,1%, но была широко представлена у канглов (66,7%), в то время как ее частота составляла <5,5% во всех других племенах. Другие гаплогруппы показывают частоты ниже 2% в выборке.
Y-гаплогруппы нанесённые на карту Казахстана отражают исторический след монгольских захватчиков, которые свирепствовали в Центральной Азии в 13 веке, поскольку гаплогруппа C2 чаще встречается в южной и западной частях страны, где прошли монголы.
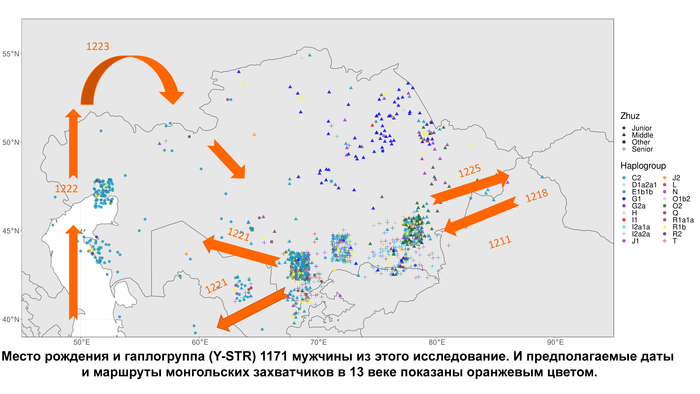
Например, в южных и западных регионах Казахстана, племена албанов, конгратов, дулатов, байулов и алимулов, имеют частоты C2-M217 ≥ 70%, в то время как племена, расположенные в центре и на северо-востоке страны, такие как аргыны, уак, найманы, канглы, ысты и кыпшаки, имеют более низкие частоты (в большинстве случаев <30%).
Разнообразие гаплотипов
У 1171 человека было обнаружено 577 различных гаплотипов, из них 429 наблюдались однократно, а остальные 148 от 2 до 51 раза, при этом 15 гаплотипов наблюдались ≥10 раз.
В целом, это привело к значительному разнообразию гаплотипов Y-хромосомы – 0,9938 ± 0,0006, что отражает глубокие отцовские линии разного происхождения в выборке. Однако дифференцирующая или различающая способность выборок, основанная на распределении частот гаплотипов, составила 55,17%, что отражает высокую частоту нескольких гаплотипов. Семь из девяти наиболее часто встречающихся гаплотипов принадлежали к гаплогруппе C2. Из них два наиболее распространенных (Ht1, Ht2) наблюдались у 51 человека, в то время как третий гаплотип C2 (Ht3) наблюдался у 37 человек. Ht1 присутствовал у 38 байулов и шести алимулов, оба этих племени принадлежат к Младшему жузу, территориально расположенному в Западном Казахстане.
Ht2 был выявлен у 32 дулатов и десяти албанов, оба племени из Старшего жуза в юго-восточном регионе. Наконец, гаплотип C2 (Ht3) отмечен у 37 конгратов, Среднего жуза. Наиболее часто встречающийся гаплотип – Ht4, не относящийся к C2, принадлежал к гаплогруппе G1 и был обнаружен у 34 человек, 26 из которых принадлежали к племени аргынов Среднего жуза в Северном и Центральном Казахстане.
Структура популяции
Анализ молекулярной дисперсии показал, что от ∼73% до ∼71% различий в разнообразии коротких тандемных повторов на Y-хромосоме (Y-STR) объясняется изменчивостью внутри племен.
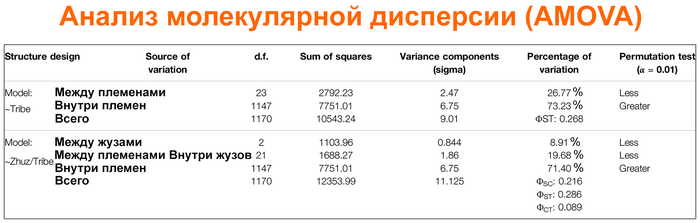
А по индексу фиксации (Fst) от 27,3 до 29,3% общего разнообразия обусловлено межплеменной дифференциацией. Попарные значения индекса фиксации между племенами варьировались от 0.0003 между байулами и алимулами до 0,19 между сиргели и шакшамами.
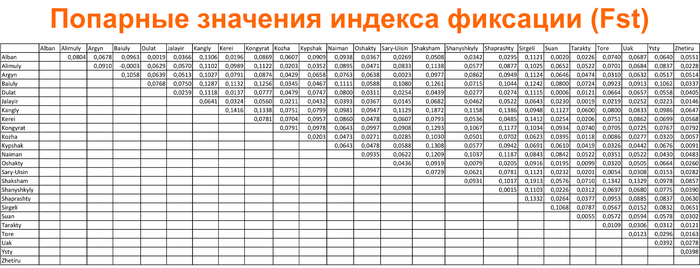
В таблице, для простоты восприятия, чем меньше значения, тем ближе представители этих племён друг к другу.
Помимо этого, несмотря на очевидную более высокую частоту гаплогруппы C2-M217 вдоль южных и западных границ Казахстана, отслеживающих монгольское вторжение, не было обнаружено существенной корреляции между генетическим и географическим расстоянием между людьми.
Структурный анализ
Визуализация структуры коротких тандемных повторов на Y-хромосоме по племенам и гаплогруппам подтверждает наличие по крайней мере трех подгрупп в пределах C2-M217: светло-зеленый (Старший жуз), бирюзово-голубой (Средний жуз) и розовый (Младший жуз).

Аналогичным образом, структурный анализ выявил наличие некоторых однородных гаплогрупп (например G1, O2 и R1a1a), которые тесно связаны с некоторыми племенами (например, G1с аргынами, а O2 с найманами).
Многомерное шкалирование
Этот анализ позволил более детально изучить взаимосвязь между племенами и гаплогруппами. Он показал, что, хотя многие из гаплогрупп попадают в середину координат с несколько отчетливой кластеризацией, гаплогруппы распределены по племенам рассеяно.
Однако, есть три отдельных кластера гаплогруппы C2-M217, которые в целом соответствуют трем подгруппам, обнаруженным в анализе структуры:
Первый встречается у многочисленных племен Младшего и Среднего жуза, в частности у байулов, аргынов, алимулов и кыпшаков, и соответствует гаплотипам розового цвета в анализе структуры.
Второй встречается среди различных представителей Старшего жуза, включая представителей албанов, шапраштов, ошактов, дулатов и суанов, все они проживают в юго-восточном Казахстане и соответствуют гаплотипам лимонно-зеленого цвета.
Наконец, третий кластер гаплогруппы C2-M217 встречается преимущественно среди представителей племени конгратов, соответствующего гаплотипу бирюзово-голубого цвета.
Другие отдельные кластеры в анализе многомерного шкалирования относятся к гаплотипу O2, который наиболее часто встречается у найманов с джалаирами и к гаплотипу G1, который встречается во многих племенах, но особенно у аргынов. Остальные гаплотипы на графике многомерного шкалирования имеют перекрывающиеся диапазоны. Это указывает на то, что между людьми с гаплогруппой C2-M217 существуют дополнительные различия в субкладах, которые требуют дальнейшего подтипирования. Таким образом, анализы демонстрируют, что существуют широкие различия между гаплогруппами среди трех жузов, в частности различия в гаплогруппе C2-M217 между жузами, а также большие различия в частотах некоторых Y- гаплогрупп среди племен.
Выводы
В этом исследовании разнообразие гаплотипов Y-STR у казахов достигло значения 0,9929, что отражает глубокие отцовские линии разного происхождения в выборке, в то время как разнообразие гаплотипов казахов из Синьцзяна оказалось ниже, возможно, из-за эффекта основателя, у мигрировавших в Китай казахов. Проще говоря, это когда на новую территорию переселилось малое количество людей и дало начало новой популяции.
Большая часть оценок генетического разнообразия, полученных для коротких тандемных повтор на Y-хромосоме, были одинаковыми для казахов из Казахстана и Китая, однако генетическое разнообразие локуса DYS448 было в два раза выше у казахов Синьцзяна.
Интересно, что наиболее частый гаплотип у казахов из Китая также является одним из наиболее распространенных гаплотипов среди казахов из Казахстана.
Гаплотип Ht8 связан с гаплогруппой O2, которая встречается с частотой 52,2% у представителей найманов, проживающих в Восточном Казахстане. Более того, казахские популяции в Алтайском крае, также характеризуются значительной долей носителей O2 (31-40%). Удивительно, но, несмотря на высокую частоту гаплотипа O2 в казахской популяции, у казахов из Северо-Западного Китая не было обнаружено мужчин с этой гаплогруппой.
В отличие от высокого разнообразия гаплотипов, дифференцирующая или различающая способность 15 локусов коротких тандемных повтор на Y-хромосоме составила всего 0,5517. Это говорит о том, что, несмотря на включение в исследование только неродственных мужчин, люди из одних и тех же или разных племен могут иметь идентичные гаплотипы, предположительно отражающие глубокое патрилинейное происхождение среди казахов. Например, различающая способность у китайцев из провинции Шаньси, Северный Китай, составила 0,9865, что указывает на высокий потенциал для генетического различия между мужчинами в этой популяции. Но у казахского населения из Синьцзяна, Северо-Западный Китай, различающая способность составляла только 0,5950.
Этот показатель в новой выборке из Казахстана также ниже по сравнению с некоторыми данными из европейских и азиатских популяций для того же набора из 15 локусов коротких тандемных повтор на Y-хромосоме.
Однако следует учесть, что показатели разнообразия варьируются в зависимости от используемых систем генотипирования коротких тандемных повтор. По мере увеличения количества наборов маркеров индексы разнообразия также увеличиваются.
Результаты анализа молекулярной дисперсии и теста Мантеля в новом исследовании подтвердили, что между жузами существует значительно меньше генетической изменчивости, чем ожидалось в рамках иерархической модели генетической структуры. Это говорит о том, что структура жуза не оказывает основного влияния на генетические отношения между казахскими племенами и что объединение в жузы было скорее условным, чем социально-территориальным, как предполагали другие авторы.
Тем не менее, примерно 10% генетической изменчивости среди казахов приходилось на вариации между жузами. Кроме того, анализ многомерного шкалирования показал некоторые различия в структуре гаплогрупп между племенами/жузами, особенно для гаплогруппы C2-M217
В анализе молекулярной дисперсии разделение генетической изменчивости внутри и между племенами показало, что ∼27% генетической изменчивости обнаруживается между племенами и ∼73% внутри племен.
Также авторы не нашли доказательств взаимосвязи между генетическими и географическими (по месту рождения) расстояниями между людьми, несмотря на очевидную более высокую частоту гаплогруппы C-M217 вдоль южных и западных границ Казахстана.
Анализ многомерного шкалирования и структуры показал, что в некоторых гаплогруппах существует значительное разнообразие. В этом плане особенно интересна гаплогруппа C-M217. Это наиболее распространенная гаплогруппа у современных казахов, но анализ, приведенный в описываемой работе, показывает, что у неё существует по крайней мере три различных субклада.
Один из них доминирует в племенах Младшего жуза (в основном у байулов с алимулами), другой у конгратов Среднего жуза и третий почти во всех племенах Старшего жуза.
В предыдущих работах в которых принимали участие люди из десяти племен Старшего жуза, пяти племен Среднего жуза и трех племен Младшего жуза, были выявлены три дочерние ветви гаплогруппы C-M217: C-M401, C-M86 и C-M407.
Авторы отмечают преобладание С-М401 в племенах Старшего жуза, С-М86 в племенах Младшем жузе, а С-М407 у конгратов Среднего жуза. Новые результаты подтверждают эти данные. Однако дальнейший углубленный анализ ряда однонуклеотидных маркеров гаплогруппы C-M217 позволит определить, какие субклады этой гаплогруппы точно присутствуют в популяции современных казахов.
В целом генетическая изменчивость между жузами отмечена на уровне ≈10% а между племенами внутри жуза ≈20%, что свидетельствует о наличии различий в структуре гаплогрупп среди казахских племен. И хотя авторы не нашли доказательств линейной зависимости между генетическим и географическим расстоянием, наблюдается более высокая частота гаплогрупп C2 вдоль южной и западной границ Казахстана, что соответствует как пути монгольского вторжения, так и приблизительному маршруту древнего Шелкового пути.
Для дальнейшего выяснения этой динамичной истории необходим более широкий пространственный и временной анализ разнообразия коротких тандемных повтор на Y-хромосоме среди казахских племен в контексте других групп Центральной Азии.
Источник:
Khussainova E, Kisselev I, Iksan O, Bekmanov B, Skvortsova L, Garshin A, Kuzovleva E, Zhaniyazov Z, Zhunussova G, Musralina L, Kahbatkyzy N, Amirgaliyeva A, Begmanova M, Seisenbayeva A, Bespalova K, Perfilyeva A, Abylkassymova G, Farkhatuly A, Good SV and Djansugurova L (2022) Genetic Relationship Among the Kazakh People Based on Y-STR Markers Reveals Evidence of Genetic Variation Among Tribes and Zhuz. Front. Genet. 12:801295. doi: 10.3389/fgene.2021.801295
Центральная Азия - это крупный регион, простирающийся от Каспийского моря на западе до озера Байкал на востоке, охватывающий Таджикистан, Казахстан, Туркмению, Узбекистан, Кыргызстан и север Афганистана. Этот регион оказался на перекрестке миграционных путей ещё с тех пор, как современные люди покинули Африку, что привело к долгосрочному присутствию людей, богатой истории и высокому культурному разнообразию. Например, Земледельческо-скотоводческие общины, существовавшие со времен джейтунской культуры 6000 лет до н. э., сменились земледельческими в эпоху энеолита (4800–3000 гг. до н.э.), с жителями деревень, ведущими хозяйство, связанное с орошаемым земледелием.
В эпоху средней бронзы на юге Центральной Азии процветала цивилизация Бактрийско-Маргианского археологического комплекса с характерными протогородскими поселениями, мощными ирригационными технологиями и выраженной социальной иерархией. Около 3000 г. до н.э., в северной части Центральной Азии распространился кочевой образ жизни, который приобрел важное значение в этом регионе в эпоху поздней бронзы (2400–2000 гг. до н.э.). Ранее было показано, что популяции Бактрийско-Маргианского археологического комплекса представляют собой в основном смесь иранских (~60–65%) и анатолийских (~20–25%) земледельцев. Однако некоторые бактрийцы имели повышенную долю степного компонента, похожего на таковой у ямной культуры, а это позволяет предположить, что степное происхождение начало появляться в Центральной Азии примерно к 2100 г. до н. э. Ранее сообщалось, что на трёх участках Бактрийско-Маргианского археологического комплекса 2100 -1700 гг. до н. э. были выявлены люди с родословной, связанной со скотоводами западной степи среднего и позднего бронзового века, которые уже имели на треть примесь от европейских земледельцев. Что говорит об обратном потоке генов с запада на восток и далее на юг.
В конце бронзового века, примерно с 1800 г. до н.э., цивилизация Окса на своем заключительном этапе претерпела важные преобразования:
⦁ оставаясь в той же традиции, материальная культура обеднела, исчезли некоторые керамические формы и артефакты;
⦁ отдельные места обитания были заброшены, монументальная архитектура исчезла, уровень технологического развития, казалось, снизился;
⦁ внешняя торговля, которая процветала во время предыдущей пиковой фазы, значительно замедлилась или даже прекратилась, за исключением контактов со степями северной части Центральной Азии;
⦁ погребальные обычаи изменились с появлением новых способов погребения, до полного исчезновения захоронений в раннем железном веке, что может быть связано с идеологической эволюцией.
В период между 1800 и 1500 годами до н. э. преобладала андроновская культура, вплоть до возникновения язской культуры раннего железного века, которая примечательна сдвигом в материальной культуре и погребальных практиках.
А позже Центральная Азия была ареной завоеваний Ахеменидов, греко-бактрийцев, парфо-сасанидов и арабов, продвигавшихся на восток, а также продвигающихся на запад различных азиатских народов, таких как гунны, хунну и монголы, прежде чем стать торговым центром вдоль Великого Шелкового пути, особенно во времена империи Сасанидов и после исламского завоевания.
В настоящее время непростая демографическая история Центральной Азии привела к сложному генетическому разнообразию, при этом современное население Центральной Азии разделено на две культурно разные группы:
⦁ Первая группа состоит из тюркоязычного и монголоязычного населения (далее именуемого тюрко-монгольским), которые являются полукочевыми скотоводами и демонстрируют генетическое сходство с населением Восточной Азии и Сибири.
⦁ А вторая группа, образована таджиками и ягнобцами, которые живут в южной части Центральной Азии, говорят на индоиранских языках, занимаются сельским хозяйством, ведут оседлый образ жизни и генетически более похожи на современные популяции Западной Евразии и иранцев.
Более того, известно, что ягнобцы долгое время находились в генетической изоляции без каких-либо признаков недавней примеси.

Современные исследования ДНК показали, что индоиранские популяции присутствовали в Центральной Азии до тюрко-монгольских групп, возможно, еще во времена неолита. А тюрко-монгольская группа возникла позже в результате смешения местных индоиранцев с южно-сибирскими или монгольскими группами с высокой долей восточноазиатского происхождения (около 60%). При этом туркмены, генетически выделяются из тюрко-монгольской группы, занимая промежуточное положение с индоиранской группой, что предполагает недавний сдвиг языка и культуры, возможно, в результате языковой замены, в основном благодаря доминированию элит.
Кстати, стоит учесть, что некоторые современные тюркоязычные популяции гораздо ближе к древним индоиранским популяциям, чем к прототюркам. Но это не делает эти древние индоиранские популяции тюрками, это по определению невозможно.
Палеогенетические исследования подтвердили, что за последние 10 тыс. лет в Евразии произошли многочисленные миграционные волны и события смешения, в которых степные популяции сыграли важную роль. При этом процессы в Европе, были лучше изучены чем в Центральной Азии, особенно в южной её части.
В северной части Центральной Азии (Казахстан, Юг России) генетические исследования указали на перемещения населения на восток и запад с позднего неолита, что привело к градиенту генетического происхождения западной степи.
В южной части Центральной Азии, где большинство древних геномов датируется поздним неолитом и бронзовым веком, было показано, что жители Бактрийско-Маргианского археологического комплекса были тесно связаны с древними популяциями южной части современного Ирана, причем некоторые их представители демонстрировали дополнительное степное происхождение.
Однако связи между современным ираноязычным населением и древними жителями южной части Центральной Азии остаются неясными.
Учёные поднимают ряд вопросов:
⦁ Каковы генетические истоки современных носителей индоиранских языков?
⦁ Можно ли проследить их происхождение до железного или бронзового века?
⦁ Существует ли одна или несколько различных популяционных историй среди данной языковой группы населения?
⦁ И какова роль туркмен в этой истории?
Палеогенетические исследования предоставили дополнительные инструменты для поиска ответов на эти вопросы. В новой работе авторы проанализировали полногеномные данные 16 современных популяций из Узбекистана, Кыргызстана, Таджикистана, Западной Монголии и Южной Сибири, совместно с древними и современными геномами.
Результаты
Генетическое сходство современных и древних индоиранцев
Чтобы исследовать связь современных жителей Центральной Азии с разнообразными древними и современными евразийскими популяциями, авторы в анализе главных компонент спроецировали 3102 древних генома на 1915 современных.
На графике анализа главных компонент современные популяции по первой главной компоненте разделяются на восточные и западные, по второй на европейские и южноазиатские, а третья компонента различает байкальские популяции из восточноазиатского кластера.
Современные индоиранцы из Центральной Азии группируются вместе на первых трех компонентах, в то время как тюрко-монгольские представители образуют градиент от индоиранского кластера к древним байкальским образцам на 3-й компоненте, в соответствии с культурной кластеризацией вместо географии.
В индоиранской группе наблюдается субструктура, состоящая их ягнобцев (TJY), тесно связанных с западным кластером, в то время как популяции таджиков (TJA, TJE, TAB) тянутся к байкальскому кластеру, что указывает на некоторую незначительную дополнительную их близость к охотникам-собирателям из Восточной Азии или Байкала (BHG).
Древние представители степи бронзового и железного веков, а также исторического периода попадают на клину простирающуюся от европейских до восточноазиатских групп.
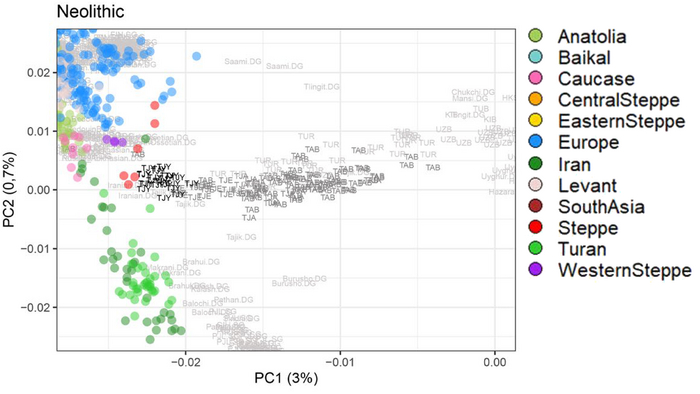
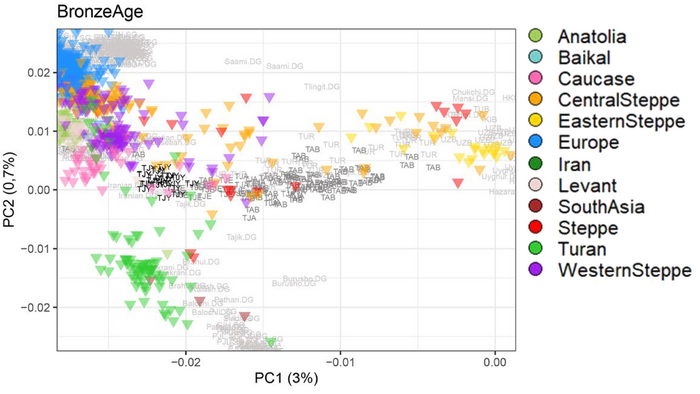

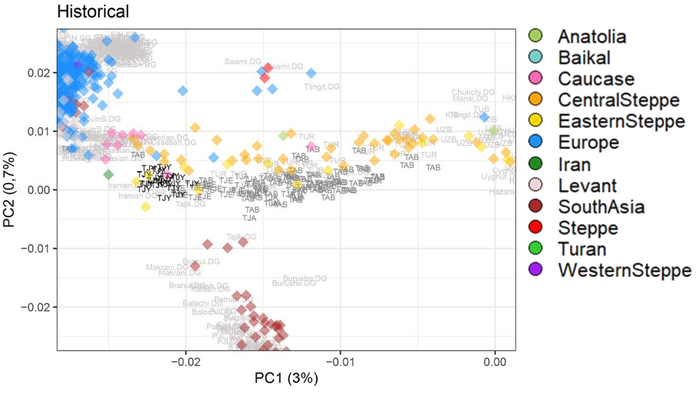
А древние жители южной части Центральной Азии (неолита, бронзового и железного веков) расположены на линии, простирающейся от иранских земледельцев неолита до современных иранцев и ягнобцев.
В целом древние и современные индоиранские популяции из Центральной Азии вместе образуют линию между неолитическими иранскими земледельцами и кластером центральной степи бронзового века с явным сдвигом в сторону степного происхождения между бронзовым и железным веками и незначительным сдвигом в сторону восточноазиатского происхождения между железным веком и настоящим временем.
При чём этот сдвиг более заметен у таджиков, чем у ягнобцев.
Анализ примесей, с тем же набором данных, подтвердили у всех современных индоиранцев наличие генетического компонента, максимизированного у иранских неолитических земледельцев (темно-зеленый; среднее значение для ягнобцев: 37%; а для таджиков 25%), другой компонент индоиранцев (бледно-зеленый; среднее значение для ягнобцев: 13%; а для таджиков: 10%), максимизирован у охотников-собирателей Восточной Европы, а также у западных и скандинавских охотников-собирателей.
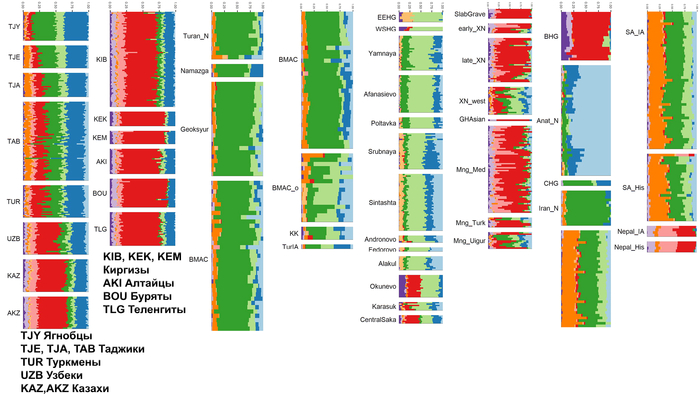
Третий компонент (темно-синий; среднее значение для ягнобцев: 36%; для таджиков: 29%), полностью не максимизирован не в одной из проанализированных популяций, но встречается у современных европейцев и у анатолийских неолитических земледельцев. А четвертый компонент, максимизированный у байкальских охотников-собирателей и в значительной степени присутствующий во всех современных тюрко-монгольских популяциях (красный; 50% в среднем), также присутствует, но в меньшей степени, и у современных индоиранских популяций, причем его доля у ягнобцев значительно меньше, чем у таджиков, а именно 7% и 14% соответственно. При этом таджики разделяют около 4% предков с современными жителями Восточной Азии, этот компонент, обозначенный на графике розовым цветом, максимизирован у китайцев, а также присутствует во всех тюрко-монгольских популяциях из Центральной Азии (среднее значение 10%) совместно с 8% компонента который максимизирован в современных популяциях Южной Азии (оранжевый). При этом оба этих компонента отсутствуют у ягнобцев.
Этот анализ также согласуется с анализом главных компонент и для древних групп. Жители юга Центральной Азии железного века демонстрируют профиль, удивительно похожий на профиль ягнобцев без вклада охотников и собирателей Байкала.
Скотоводы Центральной степи бронзового века демонстрируют аналогичный профиль, за исключением значительного увеличения иранского происхождения, а скотоводы Западной степи имеют компонент, максимально выраженный у западноевропейских охотников-собирателей (бежевый цвет WEHG), который отсутствует у современных индоиранцев.
Таким образом, современные ираноязычные популяции находятся по середине между популяциями бронзового века Центральной Степи и южной части Центральной Азии, аналогично людям железного века Туркмении, с ограниченным вкладом от групп Восточной и Южной Азии.
Итоги
Новое исследование дает представление об истории индоиранцев на юге Центральной Азии, используя фактические данные для отслеживания популяционной истории вплоть до железного века. Как было предложено предыдущими генетическими исследованиями, а также подтверждено историческими и археологическими данными, носители индоиранского языка появились в Центральной Азии задолго до носителей тюркских и монгольских.
Главное событие в основе индоиранской родословной на юге Центральной Азии произошло на рубеже бронзового и железного веков, в результате смешения местных групп Бактрийско-Маргианского археологического комплекса и населения, генетически связанного с андроновской культурой, возможно, с финальной стадией цивилизации Окса. При этом степная группа, смешавшаяся с бактирйцами, не имела восточноазиатского происхождения, что согласуется как с археологическими, так и с генетическими данными о том, что люди восточноазиатского происхождения, начали появляется в Центральной степи только в конце железного века.
Поскольку кочевники конца бронзового и железного веков генетически очень неоднородны, авторы считают, что источник происхождения западных степей, найденный в южной части Центральной Азии железного века, возможно, еще не определен. Интересно отметить, что поток генов между Степью и южной частью Центральной Азии шел в двух направлениях. Недавнее исследование показало, что поток генов от людей из Бактрии и Маргианы, способствовал генетическому формированию скифов. Новые результаты в сочетании с предыдущими исследованиями убедительно подтверждают гипотезу, основанную на археологических свидетельствах, о том, что цивилизации южной Центральной Азии, начиная с Бактрийско-Маргианского археологического комплекса, и культур западной степи имели прочную культурную связь.
Стоит упомянуть, что помимо данной работы в другом исследовании, конца прошлого года авторы, используя геномные данные южной части Узбекистана от бронзового века до железного, сообщают об увеличении примеси, связанной со степью, в железном веке по сравнению с популяциями бронзового века в регионе Бактрии и Маргианы.
Источник степного происхождения, был определён как степные популяции позднего бронзового века, которые смешались с местными популяциями, связанными с более ранними обитателями Бактрийско-Маргианского археологического комплекса, преобладающими в этом регионе. Однако миграция и смешение со степными источниками не заменили существовавшую ранее иранскую и анатолийскую родословную, связанную с земледельцами.
Популяции Узбекистана железного века моделируются как смесь иранских земледельцев (31-39%), анатолийских земледельцев (30-34%) и представителей степи (15-17%) а также с добавлением незначительного вклада от западных охотников собирателей. Популяции железного века Узбекистана также демонстрируют незначительную генетическую связь с популяциями Южной и Восточной Азии, предполагая миграции. А в целом население Узбекистана железного века было ближе к современным европейцам, чем к современным узбекам. Но тут стоит отметить, что эта схожесть обусловлена не происхождением от европейцев, а общими компонентами в генофонде, как анатолийские земледельцы и степные кочевники. Но позже в Узбекистан был поток генов из Восточной Азии и Сибири, который и привёл к формированию современного генофонда в этом регионе.
Однако в отличие от Узбекистана результаты работы по индоиранским популяциям Центральной Азии, демонстрируют замечательный пример генетической преемственности со времен железного века, несмотря на очень интенсивную миграцию населения в этом районе со времен бронзового века.
Помимо этого, как и предыдущие работы, настоящее исследование не показывает влияния арабской культурной экспансии в Центральной Азии на генетическое разнообразие носителей индоиранского языка, несмотря на то, что эта экспансия привела к изменению языка таджиков.
Помимо этого, также не выявлено потока генов из Ирана, несмотря на персидскую культурную экспансию, которая привела к языковому переходу от восточноиранского языка к западноиранскому у таджиков - в то время как ягнобцы сохранили свой язык, относящийся к северо-восточной подгруппе восточной группы иранских языков. При этом ягнобцы характеризуются генетической стабильностью с течением времени, что может быть связано с их долгосрочной изоляцией из-за труднодоступности долины реки Ягноб. Имеющиеся данные свидетельствуют о том, что разделение между ягнобцами и таджиками произошло по меньшей мере 1000 лет назад, что объясняет высокую генетическую дифференциацию, наблюдаемую у индоиранцев в предыдущих исследованиях. Это подразумевает, что ягнобцы могли бы послужить хорошим примером генофонда, присутствовавшего в Центральной Азии до миграционных волн, которые привели к нынешнему генетическому разнообразию, несмотря на произошедший сильный дрейф. Хотя количество восточноазиатских предков из-за смешения с современными тюрко-монгольскими группами остается низким, даже у таджиков, что согласуется и с предыдущими выводами, в которых учёные отметили лишь незначительное влияние двигавшихся на запад гуннов и монголов на индоиранские группы Центральной Азии. Авторы работы обнаружили у ягнобцев, таджиков и туркмен незначительный поток генов, связанный с охотниками-собирателями Байкала, около тысячи лет назад, что свидетельствует о недавней волне миграции на запад со стороны Алтая после железного века. Эта недавняя волна может быть связана с появлением тюрко-монгольских языков в Центральной Азии, что было продемонстрировано и в других работах за последние 10 лет.
Кроме того, данные подтверждают генетическую однородность среди ягнобцев, таджиков и туркмен, несмотря на их культурные и, особенно, языковые отличия, хотя и с некоторыми генетическими различиями, возникшими в результате различных моделей потоков генов у таджиков и туркмен.
Примечательно, что в новой работе авторы засвидетельствовали ранее неизвестное событие примеси из Южной Азии у таджиков, несмотря на свидетельства такой примеси у иранских туркмен.
Согласно предыдущим археологическим исследованиям, известно, что разнонаправленный культурный обмен с Южной Азией имел место быть, еще в период энеолита: в частности, от культуры Сиалк и других иранских культур в направлении Белуджистана или от культуры Гёксюра в Туркмении до южного Афганистана. В противоположном направлении, с юга на север, керамика типа Мундигак III в провинции Кандагар находит параллели вплоть до Бадахшана на северо-востоке Афганистана, а материалы из Белуджистана и раковины, используемые в ожерельях и браслетах из Аравийского моря, найдены на участке Саразм в Таджикистане, что свидетельствует о торговом обмене на большие расстояния.
Интересно, что генетическая близость между группами южной части Центральной Азии и Южной Азии уже была предложена для образцов Бактрийско-Маргианского археологического комплекса и поднимает вопрос о сроках этого потока генов. Дата смешения с южноазиатскими компонентами, полученная исследователями, совпадает с переходом с восточного на западноиранский язык у таджиков и связывается с персидской экспансией 1500 лет назад.
Наконец, случай с туркменами является ярким примером того, как население меняет язык и культуру без существенных генетических изменений. Действительно, тюркоязычные народы, населяющие всю Евразию, являются результатом нескольких кочевых миграций, которые охватывают территорию от Сибири до Восточной Европы и Ближнего Востока, через Центральную Азию и происходили в течение длительного периода, с V по XVI век.
Несколько предыдущих исследований показали, что в регионах, за пределами Центральной Азии, тюркоязычные народы генетически похожи на своих географических соседей без чётких генетических различий.
А это указывает на замену языка в этих регионах, на тюркские, благодаря доминированию элит, а не замене населения. Туркмены вписываются в эту глобальную модель, но являются исключением в своем регионе. Потому как, другие тюркоязычные популяции Центральной Азии, такие как киргизы или казахи, демонстрируют другой генетический профиль с явным доминированием восточноазиатских и байкальских компонентов, что свидетельствует о более значительной примеси кочевников из Южной Сибири и Монголии, которые датируются примерно X-XIV веками. А небольшое количество восточноазиатских предков у туркмен было связано с примесью, датируемой примерно XV веком, то есть немного позже первой примеси в Центральной Азии, и может происходить от этих тюрко-монгольских групп. А генетическая близость туркмен к современным западноевразийским популяциям, отмеченная в более ранних исследованиях, обусловлена общим степным происхождением. При этом в целом случай с туркменами представляет собой еще один пример, когда язык и генетика не совпадают, ставя под сомнение идею о распространении языков только по средствам миграций.
Источник: Guarino-Vignon, P., Marchi, N., Bendezu-Sarmiento, J. et al. Genetic continuity of Indo-Iranian speakers since the Iron Age in southern Central Asia. Sci Rep 12, 733 (2022). doi.org/10.1038/s41598-021-04144-4