Всем привет! Очередная порция науки и фантастики. Про фантастику в этот раз немного, только про Дюну, ну и про японскую мангу.
А, вот про науку чуть побольше. Роскосмос показал снимки с нового спутника. По какому-то стечению обстоятельств для демонстрации выбрал территорию США. А еще одна научная новость так вообще на грани фантастики — 3D-модель вселенной
Как, согласно китайским легендам, появилась Поднебесная? Кто её основал? Кто такие Три Властителя? Как появились люди? И почему, по мнению древних китайцев, жить можно было только в Поднебесной?
Об этом рассказывает Сергей Дмитриев, китаевед, кандидат исторических наук, ведущий научный сотрудник отдела Китая Института Востоковедения РАН, заведующий сектором древней и средневековой истории Китая, доцент УНЦ «Философия Востока» Философского факультета РГГУ.
Примечание: текста много, тема сложная и состоит из двух постов. Разделы – "Социокультурные изменения" и "Итоги" (Появление и сокращение генетического разрыва), во второй части.
Вступление
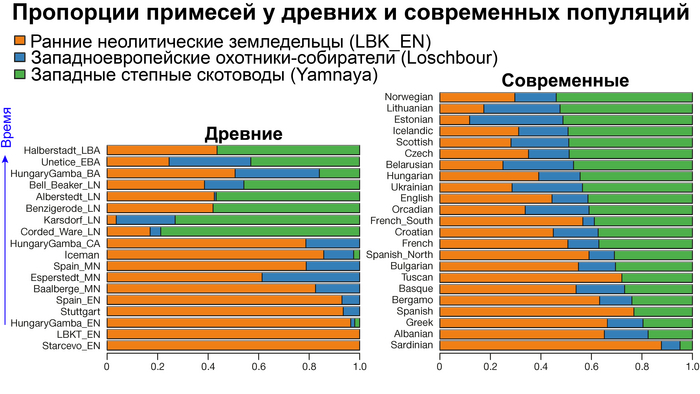
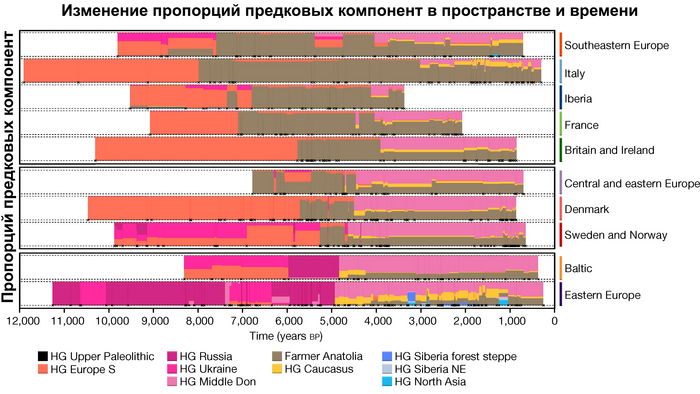
Генетическое разнообразие человеческих популяций Западной Евразии в значительной степени сформировалось в результате трех основных доисторических миграций:
1. Миграции охотников-собирателей, а именно анатомически современных людей, находящихся на ранней стадии социально-экономического развития, которые занимали территорию, примерно, с 45 тыс. лет назад. При этом вклад в последующие популяции внесли выжившие их линии после последнего ледникового максимума около 25-19 тыс. лет назад.
2. Далее, гораздо ещё более существенный вклад, внесли ранние неолитические земледельцы, пришедшие с Ближнего Востока примерно с 11 тыс. лет назад.
3. Степные скотоводы, распространившиеся из Причерноморско-Каспийской степи около 5000 лет назад, привнесли третью основную генетическую компоненту в популяции Западной Евразии.
Аутосомный генофонд практически всех европейских популяций по большей части состоит из различных пропорций этих предковых компонент, при этом независимо какие у них гаплогруппы.
Европейский континент заселялся людьми современной анатомии в несколько волн, при этом не все линии этих людей были предками современных. Первые «сапиенсы» Европы (более 40 тыс. лет назад) пересекались и скрещивались с неандертальцами, пока те совсем не вымерли. Об этой примеси свидетельствуют геномы людей из пещеры Бачо Киро в Болгарии, связанных с археологической культурой начального верхнего палеолита, и со стоянки Пештера-ку-Оасе в Румынии. Другие люди того периода, такие как женщина из пещеры Златый кунь Чехии и человек из Усть-Ишима России, не несут в своих геномах значительного количества неандертальского вклада, чем другие неафриканские группы. При этом люди, жившие в Европе более 40 тыс. лет назад, не оставили существенных следов в генетическом составе современных евразийских популяций, что говорит о том, что они в значительной степени вымерли или были ассимилированы последующими группами людей современной анатомии.
В те времена, на демографию древних людей влияли различные факторы, в том числе и климатические. Пик похолодания во время последнего ледникового максимума (LGM) был около 21 тыс. лет назад, когда Северная Европа была в значительной степени покрыта льдом, а южная граница ледниковых щитов проходила через Германию и Польшу. Многие участки Европы стали холодными пустынями с тундростепями и лесостепями на побережье Средиземного моря. А люди и животные переместились в южные широты, о чем свидетельствует, например, солютрейская культура на Пиренейском полуострове и на юге Франции. Другими предложенными рефугиумами или климатическими убежищами в тот период были Апеннинский полуостров, Балканы и юго-восточная часть Европейской равнины.
А в результате послеледниковой колонизации, палеогенетический анализ выявил разделение популяций охотников-собирателей на западных и восточных. Источники родословной западных охотников-собирателей (WHG) связаны с носителями эпиграветтской, азилийской и других эпипалеолитических культур генетического кластера Виллабруна, появление которого обусловлено миграциями в конце ледникового периода и появлением новой генетической компоненты, связанной с Ближним Востоком того времени, начиная с бёллинг-аллерёдского потепления около 14 тыс. лет назад, что археологически коррелирует с культурными переходами.
А родословная восточных охотников-собирателей (EHG) демонстрирует дальнейшую примесь с верхнепалеолитическими популяциями Сибири - древними северными евразийцами (ANE).
Предковый состав западных охотников-собирателей (WHG) был регионально изменчивым в мезолитических популяциях. Если охотники и собиратели из Британии и северо-западной континентальной Европы продемонстрировали однородный состав от палеолита и мезолита, а также вплоть до неолитической экспансии около 6000 лет назад, то их коллеги из Испании, представляли собой смесь людей из двух рефугиумов, когда к генетическому профилю кластера Виллабруна, добавляется примесь от более ранних местных пиренейских охотников-собирателей, связанных с солютрейской и мадленской культурами верхнего палеолита.
Этот генетический профиль сохранился во время последнего ледникового максимума в Юго-Западной Европе около 25-19 тыс. лет назад.
А время появления примеси сформировавшей восточных охотников-собирателей (EHG), оценивается в 15–13 тыс. лет назад, однако её пропорции в популяциях, по-видимому, соответствуют клине, которая в целом коррелирует с географией, так охотники-собиратели Прибалтики и Украины демонстрируют большее сходство с предками кластера Виллабруна, чем охотники и собиратели западной части современной России, у которых было больше вклада от древних северных евразийцев (ANE).
В свою очередь скандинавские охотники и собиратели продемонстрировали различные пропорции смеси западных и восточных охотников и собирателей (WHG и EHG) в позднем мезолите.
Стоит отметить, что охотники и собиратели мезолита и тем более палеолита, были очень разнообразны и неоднородны, но из-за ограниченного количества данных, наши знания о мезолитической структуре населения и процессах демографического смешения долгое время были скудны и содержали существенные хронологические и географические пробелы в информации. Отчасти это объясняется малым количеством хорошо сохранившихся мезолитических человеческих останков возрастом более 8000 лет, а отчасти и потому, что большинство исследований древней ДНК, посвященных периодам мезолита и неолита, были ограничены преимущественно находками из Европы.
1/3
Хотя давно напрашивается более детальная классификация охотников и собирателей по их характерным отличиям. Так, археологические данные указывают на условную границу от восточной Балтики до Черного моря, к востоку от которой сообщества охотников и собирателей существовали гораздо дольше, чем в Западной Европе, несмотря на аналогичное расстояние до центра распространения раннего земледелия на Ближнем Востоке.
Компоненты восточного и западного происхождения у охотников-собирателей в этом пограничном регионе сильно различаются, но более точные представления о демографических процессах, связанных с разделением на восток и запад и о динамике численности населения к востоку от Европы, включая Северную и Центральную Азию за тот же период времени, весьма ограничены. В этих регионах термин «неолит» характеризуется культурными, экономическими и социальными изменениями, а также использованием керамики. К примеру, неолитические культуры степей Центральной Азии и таежного пояса России уже изготавливали керамику, однако продолжали вести прежний образ жизни с присваивающей экономикой, как у предшествующих мезолитических культур охотников-собирателей, с различными инновациями в зависимости от условий среды обитания. Однако пока недостаточно данных для понимания того, как неолитизация отличалась по своим срокам, механизмам и последствиям в разных уголках Евразии.
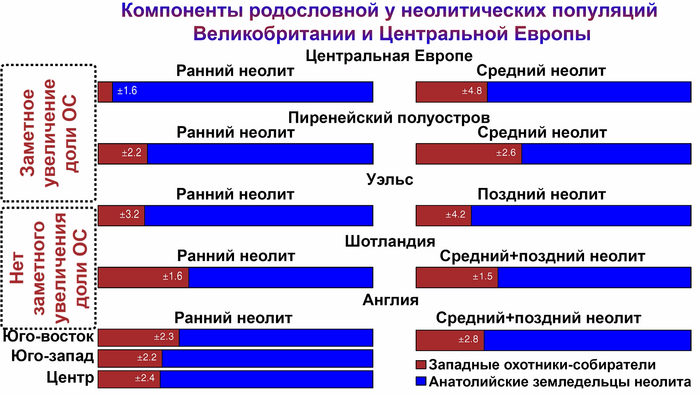
В Европе, кроме восточной её части, переход от охоты и собирательства к земледелию сопровождался прибытием мигрантов ближневосточного происхождения, практикующим производящее хозяйство на основе одомашненных растений и животных. С последующим значительным ростом населения. В Юго-Восточной, Центральной и Западной Европе анатолийские земледельцы впоследствии в некоторой степени смешались с местными охотниками и собирателями, а на территории Британии, прибывшие с континентальной Европы ранние европейские земледельцы полностью заменили местных охотников и собирателей.
А вот в восточно-балтийском регионе наблюдается заметно иная модель неолитизации, с появлением одомашненных животных только при появлении культуры шнуровой керамики (CWC) около 4800 лет назад. Аналогичным образом в Северном Причерноморье люди с генетическим профилем мезолитических охотников и собирателей тысячелетиями сосуществовали с земледельческими группами, расположенными дальше на запад.
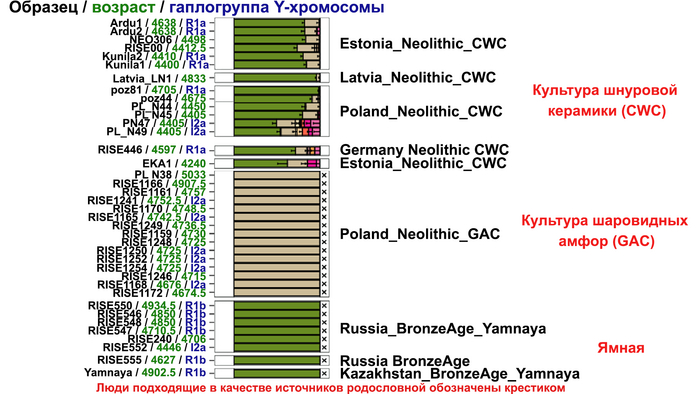
И третий крупнейший генетический пласт в истории европейских популяций связан со степными скотоводами раннего бронзового века, как представители ямной культуры около 5 тыс. лет назад. Этот генетический профиль, быстро распространился по Европе посредством экспансии как носителей культуры шнуровой керамики (CWC), так и родственных ей культур.
Хотя предыдущие исследования выявили эти крупномасштабные миграции в Европу и Центральную Азию, основные аспекты, касающиеся демографических процессов, не решены. Ямная или степная родословная в общих чертах охарактеризована как смесь восточноевропейских (EHG) и кавказских охотников-собирателей (CHG), образовавшаяся в результате гипотетической смеси «северного» степного и «южного» кавказского источников. Однако точное происхождение этих предковых источников не было установлено. Более того, за некоторым исключением, известные гаплогруппы Y-хромосомы представителей ямной культуры не соответствуют гаплогруппам, обнаруженным у европейцев после 5000 тыс. лет назад и происхождение этой отцовской линии также не установлено. К тому же "степная" родословная в Европе до сих пор была выявлена только в смешанной форме, а детали и механизмы, с помощью которых эта родословная впоследствии распространилась вместе культурой шнуровой керамики (CWC), оставались неясными.
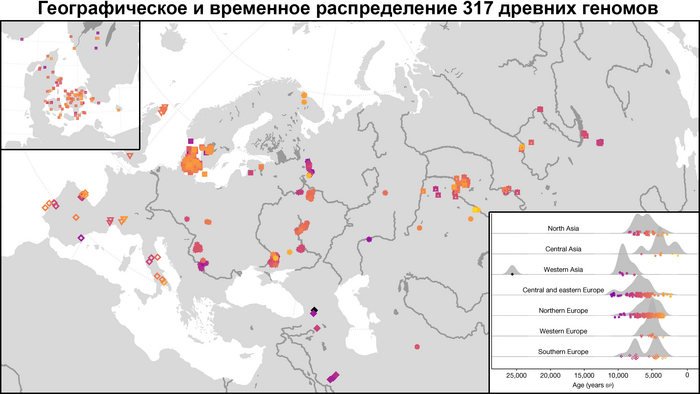
В новой работе учёные под руководством Эске Виллерслева из образцов, датированных радиоуглеродным методом при помощи ускорительной масс-спектрометрии, секвенировали геномы 317 человек, 309 из них датируются периодом от 11 до 3 тысяч лет назад.
А совместно с ранее опубликованными данными получилась выборка из 1600 древних геномов, секвенированных с помощью метода дробовика.
Географически 317 отобранных образцов охватывают обширную территорию Евразии, по долготе от Атлантического океана до озера Байкал, а по широте от Скандинавии до Ближнего Востока, и происходят из контекстов, которые включают курганы, пещеры, болота и дно водоёмов.
Географически образцы можно разделить на три региона:
1. Центральная, Западная и Северная Европа.
2. Восточная Европа, включая западную часть России, Беларусь и Украину.
3. Урал и Западная Сибирь.
Образцы включают представителей многих ключевых культур Западной Евразии мезолита, неолита и последующих периодов, такие как маглемозе, эртебёлле, воронковидных кубков (TRB) и шнуровой керамики или одиночных погребений в Скандинавии; импрессо или кардиумной керамики в Средиземноморье; кёрёш и линейно-ленточной керамики (LBK) в Юго-Восточной и Центральной Европе; и многие археологические культуры Украины, западной части России и Зауралья (как веретьё, льяловская, волосовская, китойская и др.). Выборка была особенно плотной в Дании, где сотня геномов, охватывала период от раннего мезолита до бронзового века. Обширная выборка была также получена из Украины, Западной России и Зауралья, охватывая период от раннего мезолита до неолита, примерно до 5000 лет назад.
Результаты
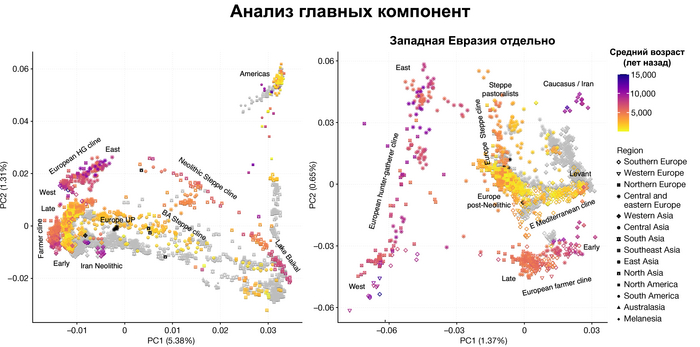
Полученный набор генетических данных позволил при анализе главных компонент использовать за основу древние геномы, и уже на них спроецировать положение современных популяций, а не наоборот, как это было во многих предыдущих исследованиях.
И хотя результаты согласуются с тем, что древние люди сильнее друг от друга генетически отличались, чем современные, такой подход позволил выявить ещё более значительные генетические различия между древними людьми, которые ранее не наблюдались, что отражает меньший эффективный размер их популяции и большую генетическую изоляцию.
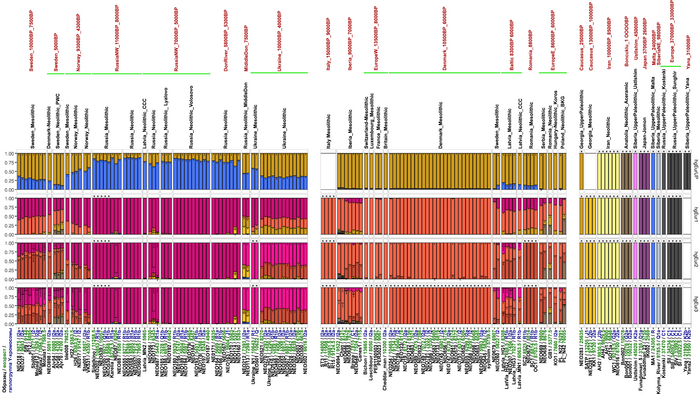
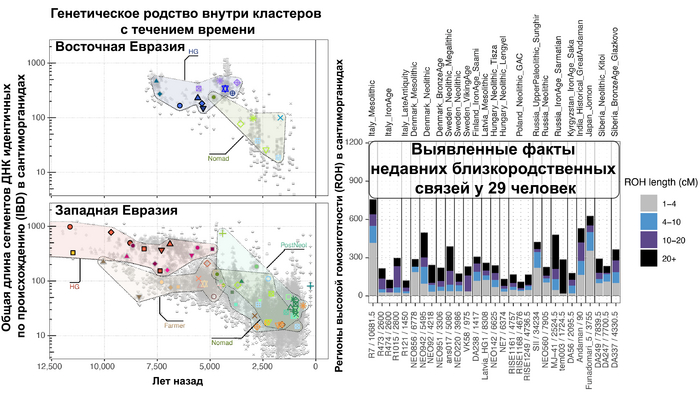
Чтобы получить более точную генетическую характеристику в пространстве и времени, авторы использовали попарную идентификацию сегментов ДНК идентичных по происхождению, чтобы сгруппировать анализируемых людей в связанные кластеры сходного генетического происхождения, а не по культурным или территориальным признакам.
В более крупном масштабе они отражали ранее описанные группы западных и восточных охотников-собирателей, а вот в более мелком масштабе они сгруппировались не только в пределах ограниченных пространственно-временных диапазонов и/или археологических контекстов, но также продемонстрировали ранее неизвестные связи в более широких географических диапазонах.
Полученные кластеры далее использовались для моделирования состава предков последующих популяций.
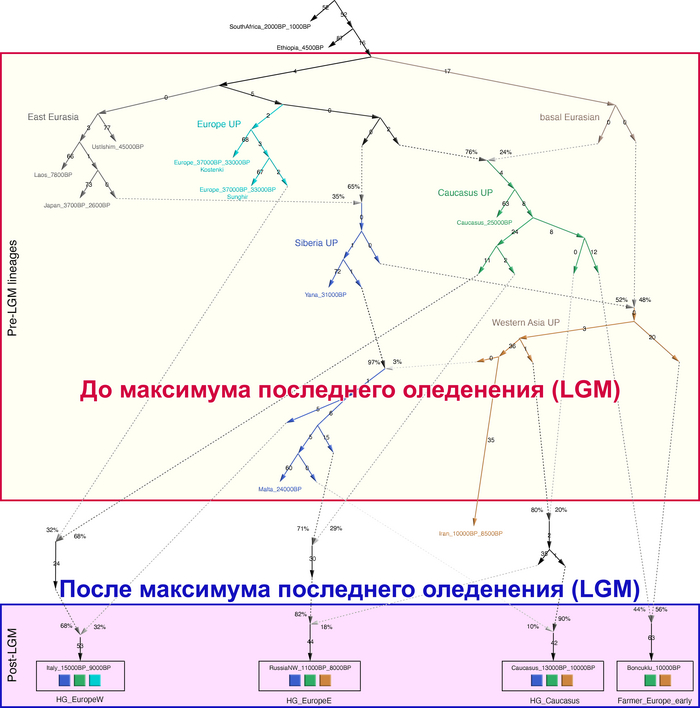
Популяционная структура охотников и собирателей после максимума последнего оледенения (LGM)
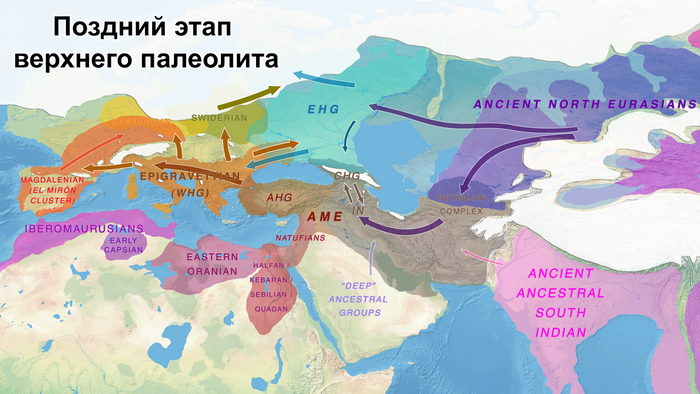
До последнего ледникового максимума, популяции Евразии довольно сильно отличались и в разной степени смешивались с неандертальцами. Некоторые популяции пока остаются гипотетическими, как к примеру, так называемые, «базальные евразийцы» с незначительной или нулевой примесью от неандертальцев, их прямых образцов, подтвержденных генетикой, на данный момент нет, но генетические следы этой популяции были обнаружены у охотников-собирателей Кавказа и Западной Азии верхнего палеолита, а также у более поздних западноевропейских охотников и собирателей.
В данной работе авторы сообщают о генетических данных полученных из зуба женщины верхнего палеолита, жившей около 26-25 тыс. лет назад в окрестностях карстового грота Котиас Клде в современной Грузии, не путать с более молодыми образцами кавказских охотников-собирателей из этого археологического комплекса, у которых было менее 20% этой кавказской родословной верхнего палеолита. А сам геном этой представительницы Кавказа верхнего палеолита представляет собой смесь 76% западноевразийских охотников-собирателей верхнего палеолита (т.е. до максимума последнего оледенения) и 24% «базальных евразийцев». Кстати популяции Ближнего Востока верхнего палеолита, почти наполовину произошли от «базальных евразийцев», которые, практически не имели примеси неандертальцев и отделились от других неафриканских линий ещё до их разделения между собой. При этом западноевропейские охотники-собиратели, жившие после последнего ледникового максимума, моделируются как итоговая, за несколько событий, смесь 46% популяций, связанных с Кавказом верхнего палеолита, 22% европейцев, живших около 33-37 тыс. лет назад, связанных со стоянками Сунгирь и Костёнки и 32% древних популяций, связанных с Сибирью верхнего палеолита, как представитель стоянки Мальта, возрастом 24 тыс. лет, т.е. древних северных евразийцев.
В свою очередь восточноевропейские охотники-собиратели моделируются как смесь 58% популяций, связанных с обитателями стоянки Мальта в Прибайкалье, 28% популяций, связанных с Кавказом верхнего палеолита, как женщина из Котиас Клде (но в разные периоды, сначала 24%, а потом 4%) и 14% представителей Западной Азии верхнего палеолита. Эта смесь также проходила в несколько этапов. Тем самым новые данные подтверждают генетическое разделение охотников и собирателей в Восточной и Западной Европе после максимума последнего оледенения, когда люди снова расселялись по Евразии, при этом эти различия сохранялись на протяжении всего мезолита.
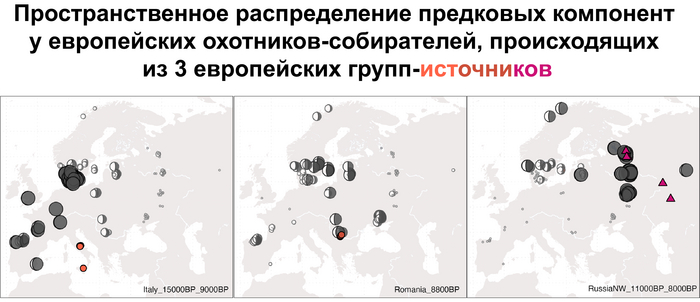
В анализе примесей, эти сложносоставные генофонды представлены отдельными компонентами. А различия между ними, а именно между западноевропейскими и восточноевропейскими охотниками-собирателями мезолита максимально выражены у представителей Италии, возрастом от 15 до 9 тыс. лет и Северо-Запада современной России возрастом от 11 до 8 тыс. лет.
Обратите внимание, что генетический состав одинаков даже у людей с разными гаплогруппами и наоборот, по всему геному люди с одинаковыми линиями гаплогрупп отличаются.
Кстати, в представлении сложных процессов, не стоит забывать о временных промежутках, о которых идёт речь.
Что касается североевропейских охотников и собирателей, то они отличались по пропорциям смеси компонент как у западноевропейских и восточноевропейских охотников-собирателей, за исключением Дании, где популяции продемонстрировали южноевропейское происхождение и были генетически схожи с охотниками-собирателями из Италии, возрастом от 15 до 9 тыс. лет. При чём эта родословная сохранялась в регионе на протяжении 5 тыс. лет.
В целом результаты указывают на то, что существовало по меньшей мере три отчетливые волны миграций охотников и собирателей в Скандинавию:
1. Первая волна связана с миграцией людей южноевропейского происхождения на территорию Дании и прибрежной части Юго-Западной Швеции.
2. Вторая волна в Прибалтику и Юго-восточную Швецию связана с популяциями из Юго-Восточной Европы.
И 3. Третья волна миграций на крайний север Скандинавии, а затем на юг вдоль атлантического побережья Норвегии связана с охотниками и собирателями с северо-запада современной России.
Эти миграции, вероятно, представляют собой послеледниковую экспансию из рефугиумов или районов-убежищ, общих как для людей, так и для многих видов растений и животных.
Стоит отметить, что генофонд европейских охотников-собирателей верхнего палеолита, живших до максимума последнего оледенения был в значительной степени заменён генофондом, происходящим из рефугиума в Южной Италии. На Пиренейском полуострове самые древние образцы из этого исследования, в том числе охотника-собирателя, возрастом около 9500 лет (NEO694) из бухты Санта-Майра на востоке Испании, показали преимущественно южноевропейское происхождение с незначительным вкладом охотников и собирателей верхнего палеолита, который, вероятно, отражает генофонд, связанный с мадленским периодом до максимума последнего оледенения.
А более поздние жители севера Пиренейского полуострова, были ближе к охотникам и собирателям Юго-Восточной Европы, демонстрируя около 40–30% вклада балканских охотников-собирателей.
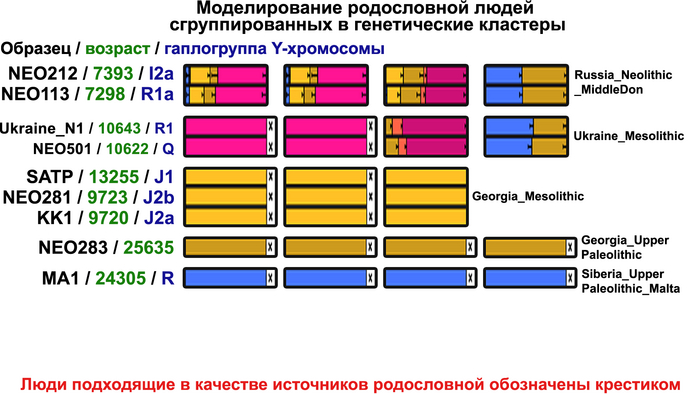
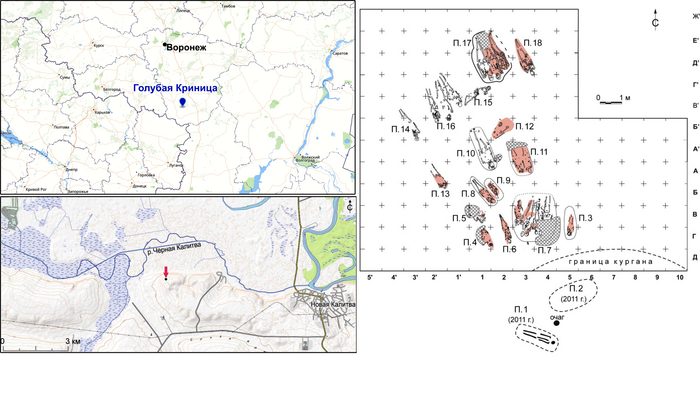
Поток генов, связанных с охотниками-собирателям Юго-Восточной Европы был направлен и в Восточную Европу после мезолита, на территорию современной Украины и далее. А у двух человек из могильника мариупольского типа раннего энеолита (NEO212, NEO113 ) близ хутора Голубая Криница на юге Воронежской области, возрастом около 7300 лет, генетический профиль состоял уже из смеси более ранних охотников-собирателей с территории Украины с вкладом от 18 до 24% из источника, связанного с охотниками-собирателями Кавказа.
1/2
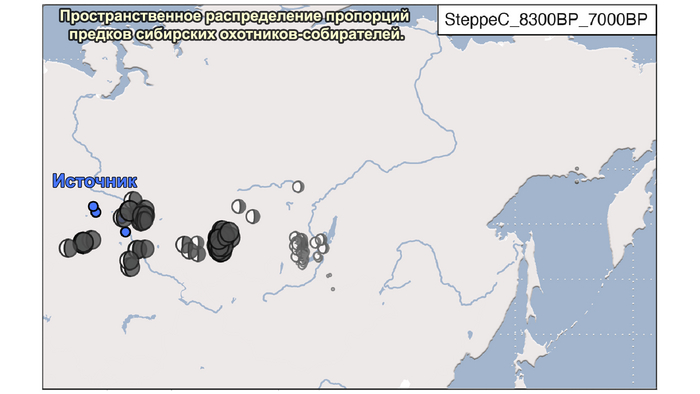
Поэтому на графике анализа главных компонент они отклонены в сторону Ирана и Кавказа. По расчётам эта примесь произошла примерно 8300 лет назад. Результаты указывают на более ранний генетический контакт между популяциями Кавказа и степи ещё до появления культур кочевников в Причерноморско-Каспийской степи. Помимо этого, этот контакт задокументирован западнее, чем предполагалось ранее.
Основные генетические изменения в Европе
Предыдущие исследования древней геномики задокументировали несколько эпизодов крупномасштабной смены населения в Европе за последние 10 тыс. лет, но 317 новых геномов из данной работы, заполняют важные пробелы в наших знаниях.
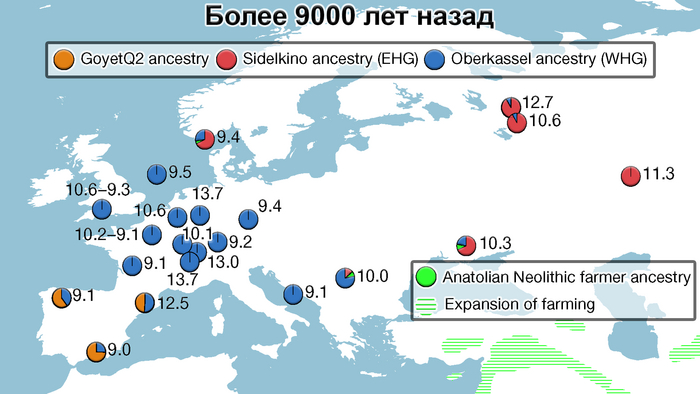
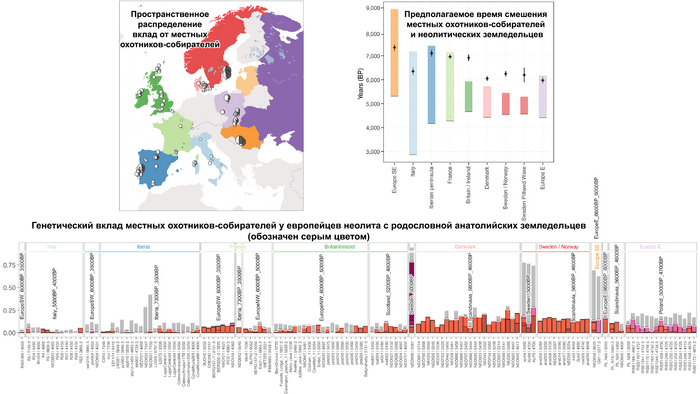
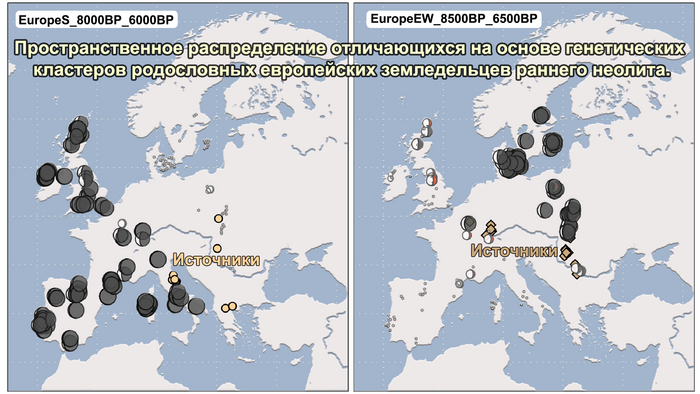
Анализ выявил глубокие различия в пространственно-временной динамике неолитизации между восточной и западной частями Европы вдоль пограничной зоны, простирающейся от Черного моря до Балтийского. К западу от этой пограничной зоны, переход к неолиту сопровождался крупномасштабными сдвигами в генетической родословной, от местных охотников-собирателей к земледельцам анатолийского происхождения (Boncuklu_10000BP). Этот переход в зависимости от региона охватывал обширный период времени, более 3 тыс. лет, от самых ранних свидетельств на Балканах около 8700 лет назад до 5900 лет назад в Дании.
В последующие периоды смешение между ранними европейскими земледельцами и местными охотниками-собирателями привело к частичному увеличению родословной охотников и собирателей во многих регионах Европы, но этот вклад редко превышал 10%, помимо заметных исключений, наблюдаемых у людей из района Железных ворот на границе современных Сербии и Румынии, а также у носителей культур ямочной керамики из Швеции и кардиумной керамики раннего неолита из Португалии, которые, по оценкам, содержат от 27 до 43% вклада от охотников и собирателей, как представители Пиренейского полуострова, возрастом от 9 до 7 тыс. лет (Iberia_9000BP_7000BP).
Обитатели эпохи неолита с территории современной Дании продемонстрировали до 25% генофонда охотников-собирателей (EuropeW_13500BP_8000BP), но преимущественно не местных.
Также обнаружены региональные различия в происхождении последующих неолитических групп. Так последующие неолитические популяции Западной Европы происходят от ранних земледельцев Южной Европы. А в последующие неолитические группы Восточной Европы и Скандинавии внесли свой вклад преимущественно ранние земледельцы Центральной Европы.
А вот с восточной стороны упомянутой пограничной зоны, на территории Прибалтики, Украины и западной части России, не наблюдается существенных генетических изменений, а предковые линии местных охотников и собирателей, без заметного вклада от анатолийских земледельцев,сохраняются примерно до 5 тыс. лет назад. Эта генетическая преемственность согласуется с археологическими данными, которые свидетельствуют о сохранении в этом обширном регионе групп охотников, рыболовов и собирателей, которые использовали керамику, но не практиковали земледелие и животноводство ещё на несколько тысяч лет.
Но несмотря на то, что влияние степной родословной на популяции Евразии хорошо известно, вопросы о её происхождении оставались не решенными. В данной работе авторы показывают, что состав степной родословной (Steppe_5000BP_4300BP) может быть смоделирован как смесь примерно 65% компонент, связанных с охотниками-собирателями, использующими керамику, из района Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет (MiddleDon_7500BP) и 35% компонент, связанных с охотниками и собирателями Кавказа, возрастом от 13 до 10 тыс. лет (Caucasus_13000BP_10000BP).
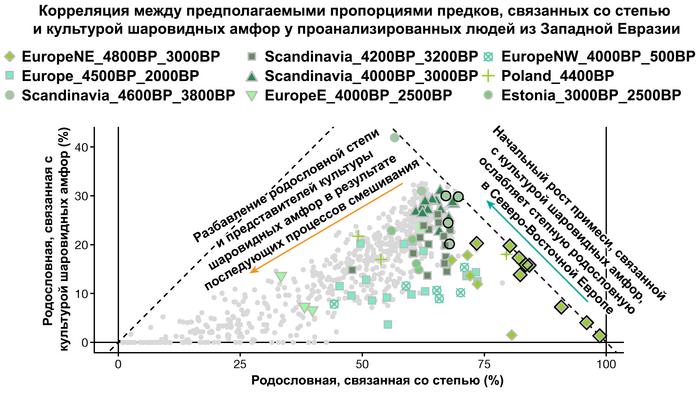
Таким образом, охотники-собиратели Среднего Дона, которые уже имели родословную, связанную с кавказскими охотниками-собирателями, являются неизвестным до сих пор ближайшим так называемым северным источником большей части родословной у людей, связанных с ямной культурой. При этом генетический состав людей из могильника Голубая Криница, отличается от такового у других древних людей из могильников мариупольского типа Приднепровья. Однако распространение степной родословной в Европе связно с культурой шнуровой керамики, носители которой представляют собой смесь предковых линий, связанных со степью и неолитическими земледельцами, происхождение которых может быть связано исключительно с культурой шаровидных амфор.
Это наблюдение имеет большое значение для понимания возникновения культуры шнуровой керамики. Хотя уже давно высказывались предположения о стилистической связи между керамикой культур шаровидных амфор и шнуровой керамики, включая использование сосудов в форме амфор и узоры, выполненные шнуром. При этом незадолго до появления самых ранних групп культуры шнуровой керамики восточные группы культуры шаровидных амфор и западные группы ямной культуры обменялись культурными элементами в лесостепной переходной зоне к северо-западу от Черного моря, где керамические амфоры и кремневые топоры культуры шаровидных амфор были включены в погребения ямной, а типичное для ямной использование охры встречалось в погребениях культуры шаровидных амфор, что указывает на тесное взаимодействие между этими группами. Предыдущие генетические анализы нескольких людей из зоны контакта предполагали, что эти контакты были ограничены культурным влиянием, а не примесью. Однако, в свете новых генетических данных, похоже, что эта зона — и, возможно, другие подобные зоны контакта между культурой шаровидных амфор и группами из степи (как из ямной культуры) — были ключевыми в формировании культуры шнуровой керамики, благодаря которой генетические компоненты, связанные со степью, и культурой шаровидных амфор, распространились далеко на запад и север, что привело к регионально разнообразным ситуациям взаимодействия и смешения.
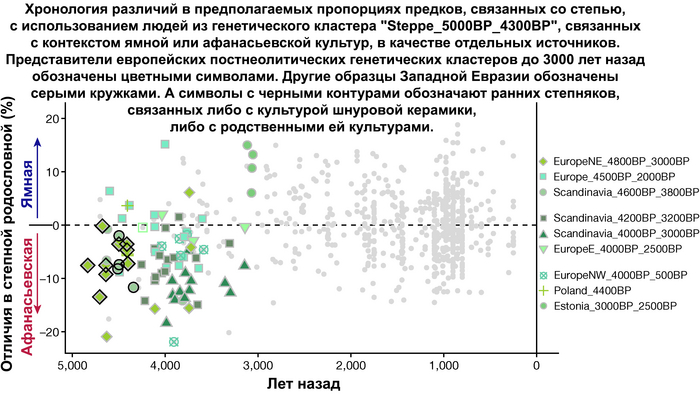
При этом распространение культуры шнуровой керамики происходило преимущественно благодаря связям, которые были выстроены культурой шаровидных амфор в предшествующий период. Но несмотря на сходство по аутосомным данным, различные линии гаплогрупп Y-хромосомы у представителей культур шнуровой керамики и ямной предполагают, что известные на данный момент образцы представителей ямной культуры не подходят в качестве непосредственного источника степной родословной у носителей культуры шнуровой керамики.
Это предположение было подтверждено моделированием предковых компонент у европейцев со степным происхождением возрастом от 3 тыс. лет назад с участием представителей родственных ямной и афанасьевской культур, потому как модели показывают, что представители афанасьевской культуры, хоть и незначительно, но оказались более лучшим источником степной родословной у европейцев, несмотря на разное географическое распространение.
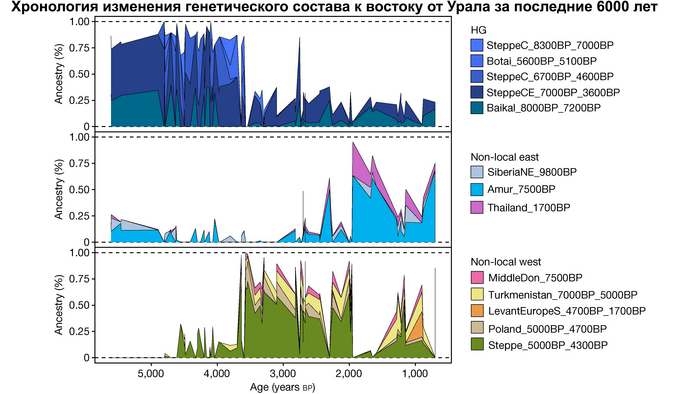
Стабильность популяций охотников-собирателей к востоку от Урала
В отличие от значительного числа геномов древних охотников-собирателей из Западной Евразии, которые были изучены до настоящего времени, геномные данные охотников-собирателей к востоку от Урала были скудны. Эти регионы характеризуются ранним появлением керамики из районов, расположенных дальше к востоку, они были населены сложными обществами охотников-собирателей с постоянными, а иногда и укрепленными поселениями. В данной работе сообщается о 38 новых образцах, 28 из которых от охотников-собирателей, использующих керамику 8300-5000 лет назад. Большинство этих геномов попадает на неолитическую степную клину, которая охватывает сибирские лесостепные зоны бассейнов рек Иртыш, Ишим, Обь, Енисей, а также озера Байкал.
Эти охотники и собиратели могут быть смоделированы как смесь компонент из трёх генетических кластеров:
1. Компонента западносибирских охотников-собирателей возрастом от 8300 до 7000 лет (SteppeC_8300BP_7000BP), доминировала в западной части лесостепной зоны.
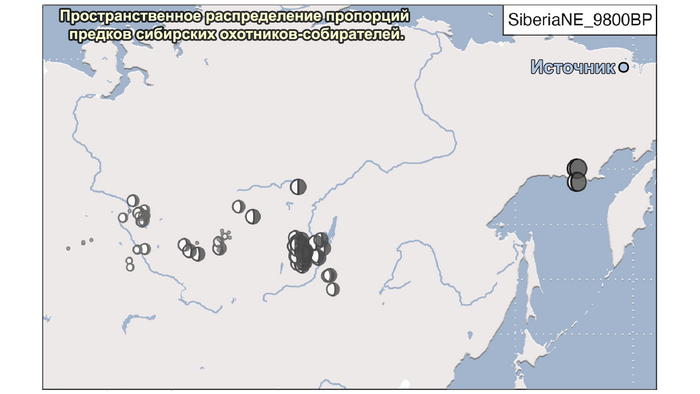
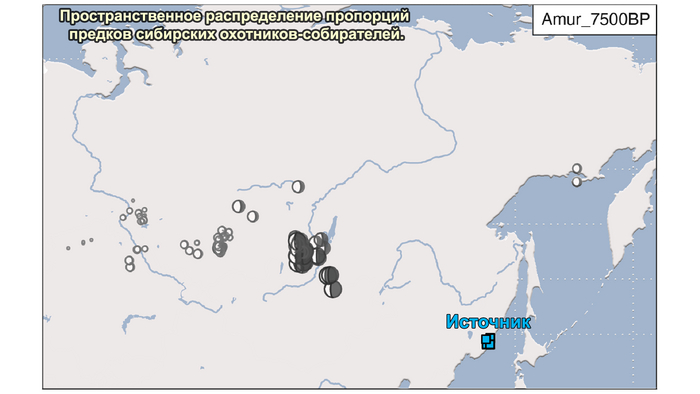
2. Компонента охотников-собирателей Северо-Восточной Азии возрастом около 7500 лет (Amur_7500BP) или древних северо-восточных азиатов из пещеры Чёртовы ворота в Приморье, была максимально выражена у популяций Байкала.
И 3. Компонента представлена палеосибирской родословной возрастом 9800 лет (Siberian_9800BP) как у обитателя Дуванного Яра в низовьях реки Колымы, она снижались по мере удаления от озера Байкал на запад.
1/5
Авторы использовали кластеры также и для исследования пространственно-временной динамики генетического состава в степи и районе озера Байкал после неолита. Результаты подтверждают увеличение компоненты охотников-собирателей лесостепи возрастом от 7000 до 3600 лет (SteppeCE_7000BP_3600BP) у жителей Байкала позднего неолит-ранней бронзы (кластеры Baikal_5600BP_5400BP и Baikal_4800BP_4200BP). Однако родословная, относящаяся к этому кластеру, уже наблюдалась у вновь описанных неолитических охотников-собирателей как у озера Байкал (NEO199 и NEO200), так и вдоль Ангары на севере (NEO843) около 7000 лет назад. У двух мужчин Байкала была определена гаплогруппа Y-хромосомы Q1b1, характерная для последующих групп позднего неолита-ранней бронзы из того же региона. Что совместно с расчётной датой примеси около 7300 лет назад, позволяет предположить, что поток генов между охотниками-собирателями Байкала и лесостепей Южной Сибири уже был во времена раннего неолита, что согласуется и с археологическими данными.
Новые образцы также проливают свет на генетическое происхождение носителей окуневской культуры раннего бронзового века в Минусинской котловине на юге Сибири. Её представителей можно смоделировать как смесь сибирских охотников-собирателей лесостепной зоны из кластеров возрастом 6700-4600 и 7000-3600 лет (Steppe_6700BP_4600BP и SteppeCE_7000BP_3600BP) со степными скотоводами возрастом от 5000 до 4300 лет (Steppe_5000BP_4300BP), связанными исключительно с афанасьевской культурой, время этой примеси датировано около 4600 лет назад.
1/2
А примерно 3700 лет назад жители в степном регионе и у озера Байкал демонстрируют уже заметно отличающиеся генетические профили. Ранние стадии этого перехода характеризуются притоком предков, связанных со степью, количество которых снижается со временем, от 70% на начальном этапе.
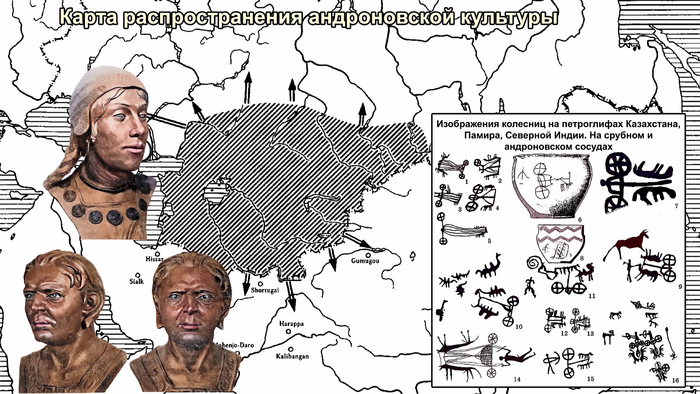
Аналогично динамике в Западной Евразии, происхождение, связанное со степью, здесь соотносится с таковым как у представителей культуры шаровидных амфор с территории современной Польши, возрастом 5000-4700 лет, подтверждая ранее задокументированный поток генов от групп из культуры шаровидных амфор в соседние группы степи и лесостепи, а также поток этой уже смешанной с европейскими земледельцами родословной дальше на восток уже с синташтинской и андроновской культурами в бронзовом веке. Однако у людей из окуневской культуры отсутствует вклад от людей культуры шаровидных амфор, а люди со степным происхождением начиная от 3700 лет назад демонстрируют чуть большую близость к ямной, чем к афанасьевской культуре, что указывает на две волны миграций западных степных скотоводов на восток. Первая волна в раннем бронзовом веке была связана с ямной культурой, а вторая, позже, была связана с синташтинской и андроновской культурами. Более поздние этапы переходного периода характеризуются увеличением родословной, связанной с иранскими земледельцами Центральной Азией (Turkmenistan_7000BP_5000BP) и древними северо-восточными азиатами Приморья (Amur_7500BP).
Суммарно, эти результаты показывают, что на востоке Евразийской степи родословная охотников и собирателей со сложной генетической структурой сохранялась и преобладала значительно дольше, чем в западной Евразии. Пока менее чем 4 тыс. лет назад по степи не прокатились волны миграций, которые привнесли в регион одомашненные породы лошадей с последующим появлением нового снаряжения для верховой езды и колесницы со спицами. Также эти миграции способствовали распространению проса в качестве сельскохозяйственной культуры.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
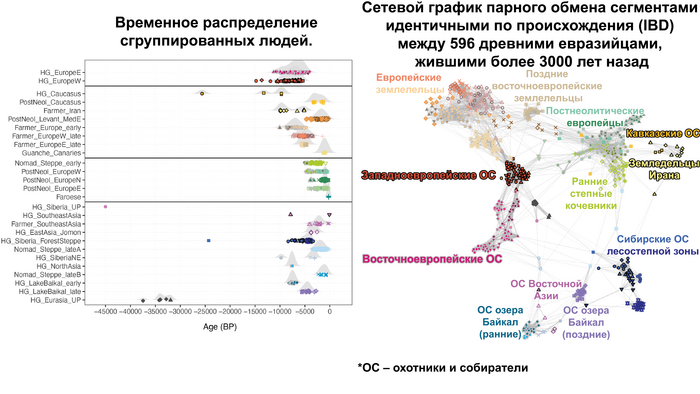
Закономерности распределения сегментов ДНК идентичных по происхождению между людьми, продемонстрировали снижение родства внутри кластеров с течением времени как в западной, так и в восточной частях Евразии. Что согласуется с увеличением эффективного размера популяции в течение этого времени. Однако наблюдаются заметные демографические различия между западной и восточной Евразией. Если на западе во время перехода к неолиту около 9000-6000 лет назад, люди с родословной анатолийских земледельцев демонстрируют снижение доли сегментов ДНК идентичных по происхождению, по сравнению с охотниками-собирателями, то на востоке генетическое родство оставалось высоким примерно до 4000 лет назад, что согласуется с гораздо более длительным существованием небольших локализованных групп охотников-собирателей.
Помимо этого, анализ сегментов гомозиготности (ROH) не выявил значительного числа близкородственных связей, только 29 из 1396. При этом каких-то пространственно-временных или культурных закономерностей в распространенности близкородственных связей обнаружено не было. Зато авторы задокументировали первый случай однородительской дисомии по древнему геному (когда человек получает две копии хромосомы или части хромосомы от одного родителя и ни одной копии от другого родителя). Эта особенность обнаружена у представителя из Темясовских курганов позднесарматской культуры возрастом около 1700 лет.
Также были выявлены люди с различной степенью родства.
Итоги
Появление и сокращение генетического разрыва
После максимума последнего оледенения около 25-19 тыс. лет назад во время расселения древних людей по Евразии, установилось чёткое генетическое разделение между востоком и западом по линии от Черного моря до Балтийского, что отражено и в археологических данных. Это разделение сохранялось на протяжении всего мезолита и неолита. В Восточной Европе распространение земледелия происходило с опозданием примерно на 3000 лет, и эта задержка может быть связана с особенностями окружающей среды, поскольку регионы к востоку от условной границы имеют более континентальный климат и более суровые зимы, возможно, менее подходящие для ближневосточных методов ведения земледелия. На этих территориях сохранились высокоразвитые сообщества охотников-собирателей со стабильными, сложными, а иногда и укрепленными поселениями, с обменом на большие расстояния и крупными кладбищами. Рацион, включающий пресноводную рыбу, зафиксирован как по изотопным данным из этого исследования, так и по анализам остатков липидов в керамике.
В северных лесных районах этой пограничной зоны сообщества охотников-собирателей сохранялись вплоть до появления культуры шнуровой керамики около 5 тыс. лет назад, тогда как в южных и восточных степных регионах охота и собирательство со временем были дополнены животноводством с крупным рогатым скотом и овцами и, возможно, коневодством в Центральной Азии, но этот факт ещё под вопросом. Некоторые из этих групп, такие как представители хвалынской культуры Северного Кавказа и Среднего Поволжья, стали свидетелями появления мужских сообществ, вовлеченных в широкомасштабные обменные связи на основе медных изделий с Восточной и Центральной Европой, а также Кавказом. Поселения в то время были приурочены в основном к равнинным поймам и долинам рек, тогда как степной пояс оставался в значительной степени неиспользованным.
Окончательное исчезновение этой генетической, экономической и социальной границы было вызвано событиями, развернувшимися в степном регионе. Здесь археологически можно наблюдать две временные фазы технологических инноваций: широкое распространение колесных повозок, запряженных волами, около 5500 лет назад и более позднее развитие колесниц со спицами и верховой езды.
Начиная ещё с первой фазы инноваций, в сочетании с возможными изменениями условий окружающей среды, степь стала выгодной экономической зоной, которая позволила группам по типу ямной культуры эксплуатировать степь в качестве кочевых степных скотоводов около 5000 лет назад. А поселения эпохи энеолита вдоль речных долин были заменены этой новой формой мобильной экономики на основе скотоводства, которая окончательно разрушила генетическую границу, существовавшую в предыдущие тысячелетия.
Далее, после второй фазы инноваций около 4 тыс. лет назад, изобретение боевых колесниц и использование проса в качестве продовольственной культуры позволили андроновцам и родственным им группам окончательно распространиться на восток в Среднюю Азию и за ее пределы, оставив глобальное наследие в виде распространения индоиранских языков.
Это исследование внесло свою лепту в наши знания о прошлом, особенно касательно степных миграций.
Во-первых, авторы выявили до сих пор неизвестный источник родословной, который способствовал появлению степных скотоводов, на его роль хорошо подходили охотники-собиратели Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет.
А во-вторых, было определено, что степная родословная, которая распространилась по Европе, благодаря представителям культуры шнуровой керамики, сформировалась в результате культурного и генетического слияния степных групп, родственных ямной культуре и культуре шаровидных амфор. При этом благодаря своему смешанному культурному и генетическому происхождению, носители культуры шнуровой керамики практиковали смешанную экономику, используя различные стратегии выживания в различных условиях. Такая гибкость в значительной степени способствовала их успеху в расселении и адаптации к очень разным экологическим и климатическим условиям за очень короткий период времени.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
Как появились самураи? На какие эпохи делится история самурайства? Чем занимались самураи в эпоху Токугава, когда не было войн и ситуация в стране была стабильна? Как появилось бусидо? Кто был последним самураем?
Об этом и многом другом рассказывает Василий Щепкин, японовед, кандидат исторических наук, старший научный сотрудник Института восточных рукописей РАН, доцент кафедры японоведения НИУ ВШЭ в Санкт-Петербурге.
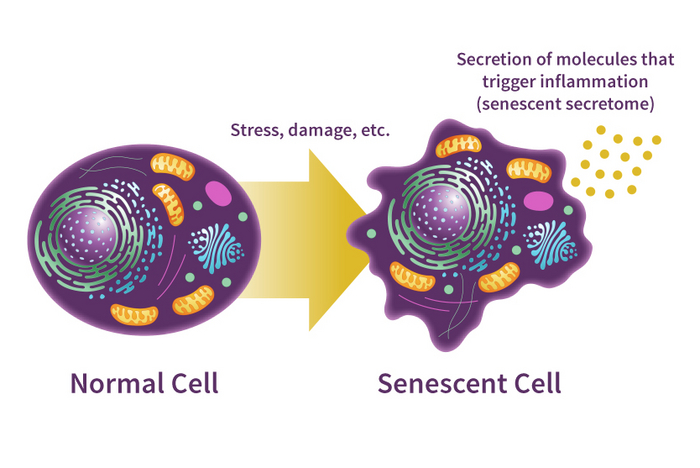
В вашем теле живут клетки-зомби. Они не совсем мёртвые. Такие клетки скорее живые, но они больше не делятся и могут свою «зомби-сущность» передавать соседям. И соседние клетки тоже превращаются в зомби! Зомби-клетки повышают риск рака и разных других заболеваний — например, артеросклероза. Сейчас учёные активно думают о том, как от таких клеток избавиться. Однако есть нюанс: наши внутренние «зомби» выполняют и полезные функции.
Учёные называют «зомби» сенесцентными клетками. И накопление этого типа клеток в нашем организме — очень важный элемент человеческого старения. Говоря простым языком, сенесцентные клетки — это клетки, которые ранее обладали способностью делиться, но из-за накопления разных повреждений делиться больше не могут. Обычно количество «зомби» с возрастом увеличивается. Из-за этого появляются разные старческие заболевания, возникает боль в суставах, нарушения когнитивных функций, системные воспаления и так далее.
Откуда в нашем теле берутся сенесцентные клетки? Это точно не старые клетки — некоторые «зомби» образуются ещё на стадии эмбрионального развития. Чаще всего сенесцентные клетки, вызывающие заболевания, появляются из-за ошибок, мутаций в ДНК и иных повреждений. Клетка, в которой накопилось много повреждений, выходит из клеточного цикла и как бы говорит: «Всё, я дальше делиться не буду». А ещё «зомби» сигнализируют о своём состоянии иммунной системе, выбрасывая провоспалительные факторы. На эти факторы «приходит» иммунная система и некоторые сенесцентные клетки утилизирует.
Ещё «зомби» могут образовываться из-за повышенной активности отдельных генов. Например, когда клетка подозревает, что может стать раковой, она превращается в сенесцентную. И это хорошо! А сейчас приведу пример, который всем понятен — родинки. Наверняка мои читатели слышали про меланому — рак кожи. Родинки — это не меланома, но в родинках клетки начинают очень активно делиться. Затем в них включается механизм сенесценции — и они делиться перестают. И родинка оказывается «зафиксированной» в своём размере и не превращается в раковую опухоль.
Важно сказать, что сенесцентные клетки могут быть очень разными как по происхождению, так и по свойствам. Поэтому некоторые учёные очень скептично относятся к термину «сенесцентные клетки» — ведь не всегда понятно, какие именно клетки имеются в виду. Интуитивно может показаться, что, раз клетка перестала делиться, значит, в ней что-то сломалось. Но на самом деле «зомби» — это клетки, в которых сработал специальный и целенаправленный механизм, который ограничивает клеточное деление. Так задумано — это не случайная поломка. Это, скорее всего, специальная «поломка» ради предотвращения онкологического заболевания.
Когда клетка сильно повреждается и не может эти повреждения исправить, у неё есть две опции. Первая — запрограммированная клеточная смерть, самоубийство клетки или апоптоз. Вторая — превращение в «зомби». Во втором случае клетка выживает, но перестаёт делиться.
Перед тем как выбрать одну из двух опций клетка пытается себя починить. Когда в клетке повреждается ДНК, включается огромное количество механизмов «репарации». Активируются разные белки, которые пытаются это дело исправить. Однако, если ошибку устранить не получается, клетке приходится выбирать одну из двух упомянутых опций.
Существует по меньшей мере четыре вида сенесценции:
Репликативное старение. У большинства клеток есть встроенный ограничитель делений — предел Хейфлика. При каждом делении кончики хромосом — теломеры — укорачиваются. И через 40-60 делений клетка перестаёт делиться — это и есть репликативное старение. Это старение особенно распространено у крупных видов животных — в частности, у человека;
Сенестенция, вызванная стрессом. Здесь речь идёт не о депрессии, а о том, что клетка может повреждаться из-за ультрафиолетового излучения, ионизирующей радиации, активных форм кислорода и так далее. В результате клетка перестаёт делиться;
Сенесценция, вызванная онкогенами. Онкогены — это гены, чья повышенная активность увеличивает риск рака. Если клетка «замечает» онкогены, то перестаёт делиться;
Сенесценция эмбрионального развития. У человека есть разные эмбриональные структуры, которые не должны чересчур разрастись в утробе. А некоторые структуры даже должны исчезнуть совсем — например, плавники и хвостик у зародыша. Благодаря сенесцентному механизму младенцы появляются на свет без хвоста!
В общем, сенесценция — это довольно нужно и важно. И всё же есть учёные, которые называют сенесцентные клетки «зомби» и считают, что от некоторых из них нужно избавляться — чтобы победить старение.
Итак, у сенесценции есть плюсы и минусы. Сначала расскажу о плюсах. Благодаря «зомби» организм предотвращает появление некоторых онкологических заболеваний. Учёные создавали ГМ-животных с отключенным геном p16. p16 — один из ключевых генов, запускающих сенесцентность. В результате подопытные массово страдали от рака. То есть полностью сенесценцию отключать нельзя! Кроме того, учёные выяснили, что «зомби» иногда помогают регенерировать ткани кожи и мышц. А сенесценция клеток поджелудочной железы может увеличивать выработку инсулина. Сенесценция же клеток печени предотвращает цирроз. В общем, «зомби» делают массу всего полезного. Кстати, такие «зомби» встречаются даже у сверхдолгожителей — голых землекопов.
При этом сенесцентные клетки обладают и рядом «недостатков». Так, с возрастом в нашем теле накапливается очень много «зомби». Особенно опасны клетки, ставшие «зомби» из-за повреждений в ДНК. Они начинают производить молекулы, которые могут вызывать хронические воспаления в тканях вокруг. Так, есть очень важная провоспалительная молекула — интерлейкин-6. Она нужна для коммуникации клеток между собой. Интерлейкин-6 активирует высокую температуру при заболеваниях, влияет на исход некоторых видов рака... Когда клетки перестают делиться из-за сенесценции, порой они начинают производить в 40 раз больше интерлейкина-6. От этого могут повреждаться соседние клетки — и у них тоже включается сенесценция. Происходит цепная реакция.
И ещё один момент: в нашем теле есть «спящие» эндогенные вирусы, которые изначально встроены в наш геном. Они попали туда в ходе эволюции и зашифрованы прямо в нашей ДНК. Так вот, в норме особые клеточные механизмы подавляют вирусы и не дают им сделать ничего плохого. Однако, как показало недавнее исследование, «зомби» не подавляют такие эндогенные вирусы. И в итоге клетка собирает из своего же генома вирусные частицы и заражает ими здоровые клетки. Заражённые клетки, в свою очередь, тоже становятся сенесцентными. Но всё не так плохо: авторы упомянутого исследования создали антитела против вирусных частиц — и остановили заражение.
Но всё же сенесцентные клетки опасны в первую очередь из-за хронического воспаления, которое они вызывают. Инфекции в организме нет, а воспаление есть. И иммунитет борется, по сути, с самим организмом. В итоге из-за хронического системного воспаления возникают артрит, атеросклероз и прочие неприятные «старческие» заболевания. Иногда из-за воспаления возникает рак, деменция, болезнь почек, депрессия. А ещё выраженность системного хронического воспаления очень неплохо предсказывает вероятности смерти человека.
Откуда мы знаем, что сенесцентные клетки существуют и работают? Надо признать: есть учёные, которые скептически относятся к концепции сенесцентных клеток. Как минимум потому, что «зомби» трудно дать точное определение — уж очень разными они бывают. Но одна команда исследователей провела эксперимент, показавший «работоспособность» сенесцентных клеток. Итак, учёные взяли клетки эпителия в предраковом состоянии — в них уже «назрели» мутации, вызывающие рак. Потом эти предраковые клетки исследователи смешали либо с сенесцентными клетками, либо с обычными, несенесцентными, которые могут делиться. А дальше ввели и первую, и вторую смесь подопытным грызунам. Оказалось, что при введении смеси из предраковых и сенесцентных клеток повышается и риск появления рака, и скорость роста опухоли. А при введении другой смеси — с нормальными клетками — таких страшных последствий не наблюдалось.
Считается, что рак — это результат мутаций в клетках. Но на самом деле на успешность и активность раковой клетки влияют ещё и соседние клетки. И если рядом с потенциально мутантной раковой клеткой находятся сенесцентные клетки, шанс развития рака повышается.
Другая команда учёных поставила ещё один опыт — взяла свиные клетки и превратила часть из них с помощью радиации в сенесцентные. Затем и нормальные клетки, и «зомби» исследователи перенесли в почки молодых мышей. Выяснилось, что из-за сенесцентных клеток у грызунов начали возникать фиброзы, воспаления и различные повреждения почек. Похожий опыт провела ещё одна группа учёных: они брали из жировой ткани мышек как сенесцентные, так и нормальные клетки, которые способны делиться. Далее специалисты понемногу пересаживали клетки другим грызунам. Выяснилось, что мышки, которым подсаживали «зомби-клетки», начали проявлять признаки старения — они хуже бегали, становились слабее. Чем больше грызун получал сенесцентных клеток, тем дряхлее он становился. В ходе эксперимента учёные выяснили, что 1 сенесцентная клетка на 10 000 нормальных примерно в 5 раз увеличивает риск смерти.
Что же делать — избавляться от «зомби» или нет? Видимо, надо оставить механизм сенесценции в покое — но при этом можно устранять уже получившиеся «зомби» по мере их накопления. Как это сделать? Наши гены должны активироваться в определённых условиях. Поэтому у каждого гена есть регуляторный участок, который «говорит», где именно и в каких условиях ген будет работать (пример такого участка — промотор, который находится перед началом гена и показывает, с какого места его «считывать»). Так мы можем «подсмотреть», какие гены активируются в сенесцентных клетках. В общем, учёные выяснили, что у существенной части сенесцентных клеток их сенесцентность запускалась уже упомянутым геном p16. У p16, как и у любого другого гена, есть промотор — вот его-то и можно «приделать» к другому гену. Тогда при «запуске» p16 параллельно запустится и тот второй ген. А теперь фокус: приклеиваем промотор p16 к гену, который запускает запрограммированную смерть клетки! Тогда в момент, когда клетка по какой-то причине «решит» стать сенесцентной, она тут же себя и убьёт.
А можно пойти ещё более хитрым путём. Например, сделать так, чтобы наш дополнительный ген, к которому мы приклеили промотор от p16, срабатывал бы только при наличии синтетического вещества, которого не бывает в организме человека. То есть, когда мышке или человеку вводят это секретное вещество, все клетки, где активен p16, убьются. Так исчезнут клетки с повреждённой ДНК, которые больше не делятся. Но будет ли такая «зачистка» полезна для здоровья?
Учёные выяснили, что да, «зачистка» скорее полезна. Исследователи в ходе своего эксперимента удаляли сенесцентные клетки у ГМ-мышей — и продолжительность жизни грызунов немного выросла! При этом животные выглядели моложе сверстников и были более активными, любознательными и бодрыми. Это круто — получается, удаление сенесцентных клеток поможет в борьбе со старением.
Но, увы, такая генная терапия должна происходить до рождения — а мы с вами взрослые люди, нас в эмбриональное состояние уже не вернуть. Что же нам делать? Как избавиться от вредных клеток? Хорошие новости: уже разработаны препараты — сенолитики — которые могут уничтожить некоторые типы сенесцентных клеток. По крайней мере, так показали опыты на животных. Так вот, самая известная комбинация сенолитиков — это дазатиниб и кверцетин. Дазатиниб — это вообще лекарство от рака и опухолей. А кверцетин — это природное вещество, которое достаточно часто встречается в продуктах питания. Исследования показали, что кверцетин усиливает действие дазатиниба.
Я уже описывал эксперимент, авторы которого пересаживали мышам сенесцентные клетки из жировой ткани. Так вот, в той работе учёные брали клетки у ГМ-доноров со светящимися клетками. Клетки доноров отличались от клеток реципиентов, их можно было легко увидеть под микроскопом. В ходе опыта реципиентам давали дазатиниб и кверцетин — и выяснилось, что сенолитики устраняли светящихся «зомби», но не устраняли нормальные светящиеся клетки. А ещё учёные поставили дополнительный эксперимент на старых мышах, чей возраст примерно эквивалентентен возрасту 80-летних стариков. Им давали те же дазатиниб и кверцетин. «С момента лечения продолжительность жизни мышей увеличилась на 36%», — резюмировали авторы статьи. Но на самом деле старые мышки просто чуть-чуть дольше протянули.
Другие учёные показали, что дазатиниб и кверцетин замедляют старение лёгких у мышей. А ещё одни исследователи поставили эксперимент на людях — правда, людей было всего 9, то есть выборка оказалась ничтожно мала. Тем не менее, учёные заявили, что сенолитики снизили число «зомби» в тканях. А вот как препараты повлияли на продолжительность жизни, непонятно — люди, в отличие от мышей, слишком долго живут, на них такие эксперименты ставить сложно.
Кроме микса из дазатиниба и кверцетина, существуют и другие сенолитики. Например, популярен экспериментальный препарат навитоклакс — тоже средство против рака. Вы, наверное, спросите: как противоопухолевое лекарство от рака помогает против сенесцентных клеток? Ведь раковые клетки суперактивно делятся, а «зомби» не делятся вовсе. Но у этих двух типов клеток есть нечто общее — у них постоянно включены «сигналы об ошибке», о повреждениях. Правда, «зомби» в ответ на сигнал перестают делиться, а раковые клетки сигнал игнорируют и очень активно делятся.
В одной работе учёные исследовали сенесцентные клетки в сердцах грызунов. Да-да, в сердце тоже могут появляться «зомби» — это приводит к фиброзу и гипертрофии сердца, что увеличивает риск внезапной смерти. В общем, учёные вызывали инфаркт у мышей. И выяснили, что старые 23-месячные грызуны, которым ранее давали навитоклакс, выздоравливали после инфаркта как 3-месячные мышки. Это как если бы 70-летний старик отошёл от инфаркта как подросток.
К сожалению, у сенолитиков есть побочные эффекты — в частности, малоприятный кашель с кровью. А теперь — к хорошим новостям:
Для борьбы с «зомби» нужны не такие высокие дозировки, как для борьбы с раком;
Сенолитики можно принимать не постоянно, а курсами: попил, а потом дальше живёшь и «копишь» своих «зомби» лет 10-20;
Есть такой сенолитик — квертицин — который идёт в комбо с дазатинибом и почти не имеет побочек. Сам по себе это флавоноид, которого много в разных фруктах и овощах, особенно в каперсах. Правда, есть овощи ради кверцитина бесполезно: чтобы получить нужное количество вещества, придётся есть по полкило каперсов в день. Что делает кверцитин? Он подавляет белок, который не даёт клетке убить себя, говорит ей «Живи!» Грубо говоря, этот белок как психотерапевт. А кверцитин приходит и убивает психотерапевта, а клетка такая: «Всё, не хочу больше жить».
Сейчас много хайпа вокруг ещё одного сенолитика — физетина. Он тоже встречается в разных фруктах и овощах — в яблоках, огурцах, луке, винограде. Биохакеры очень любят кушать физетин! Но есть нюанс. Существует научная программа тестирования интервенций — Interventions Testing Program. Она беспристрастно испытывает всякие средства против старения — три разных лаборатории в США независимо тестируют предлагаемые способы на мышах, самцах и самках. В общем, согласно Interventions Testing Program, физетин жизнь не продлевает.
Но вообще, даже если физетин или другой сенолитик работает на мышах, это не значит, что он будет работать и на нас с вами. Из 100 препаратов, которые работают на грызунах, лишь 10 оказываются эффективными на людях. Так что ждём исследований сенолитиков на людях — чтобы понять, какие из них работают — и работают без серьёзных побочных эффектов.
У нас есть часть тела, где клетки вообще не делятся — это наш мозг. Да-да, нейроны делиться не умеют, а ещё они с годами могут ухудшаться и дряхлеть. У дряхлых нейронов может, например, уменьшаться объём митохондрий. А ещё стареющие нейроны могут выделять интерлейкин-6. А значит, вызывать старческое хроническое воспаление в мозге. То же самое может происходить и с клетками, которые питают и берегут нейроны — например, с астроцитами. И даже у стволовых клеток может случаться сентенция!
В общем, кажется, дела плохи: в нашем мозге, скорее всего, с возрастом настанет старческое хроническое воспаление. Кстати, были опыты, в ходе которых в мозг мышам вводили интерлейкин-6 — и это приводило к ухудшению когнитивных функций грызунов. А ещё учёные знают, что у пациентов с Паркинсоном и Альцгеймером в спинномозговой жидкости находится много противовоспалительных маркёров.
Учёные исследовали влияние сенолитиков на мозг мышей. Оказалось, что сенолитики могут избирательно устранять «сенесцентноподобные» нейроны — повреждённые нейроны, которые вызывают воспаления. И в итоге у грызунов после устранения «проблемных» нейронов улучшились когнитивные способности. Вывод тут простой: лучше потерять немного повреждённых нейронов сегодня, но предотвратить воспаление и сохранить гораздо большее количество здоровых нейронов завтра.
Но всё же пока учёные не знают точно, стоит ли избавляться от «зомби» с помощью лекарств, предназначенных для лечения рака. А ещё устранение сенесцентных клеток — не панацея от старости, ведь «зомби» — лишь один из симптомов этого неприятного процесса. У старения больше дюжины причин!
Тем не менее, обращать внимание на сенесцентные клетки важно, потому что они приводят к системным воспалениям, а такие воспаления — сильный предсказатель смертности у людей. Недаром есть исследование: у людей, которые прожили больше 110 лет, гораздо ниже уровни маркеров воспаления — таких как интерлейкин-6. И это не совпадение. Так что продолжаем изучать сенолитики!
Напоследок расскажу про недавние эксперименты на мушках-дрозофилах. Авторы работы решили объединить два метода продления жизни:
Они удаляли «зомби»;
А ещё омолаживали другие клетки организма с помощью факторов Яманаки, превращая их в стволовые — чтобы они активнее делились и восполняли удалённые дряхлые клетки.
Учёные пришли к выводу, что вместе эти два способа дали результат лучше, чем использование лишь одного метода. Это в очередной раз говорит о том, что старение — это множество разных процессов. И чтобы его победить, нужно использовать одновременно разные методы — чинить митохондрии, увеличивать число стволовых клеток и уменьшать число «зомби», удлинять теломеразу и устранять мутации. Думаю, в будущем возникнет комбинированная терапия против старения — и люди будут жить значительно дольше, чем сейчас.