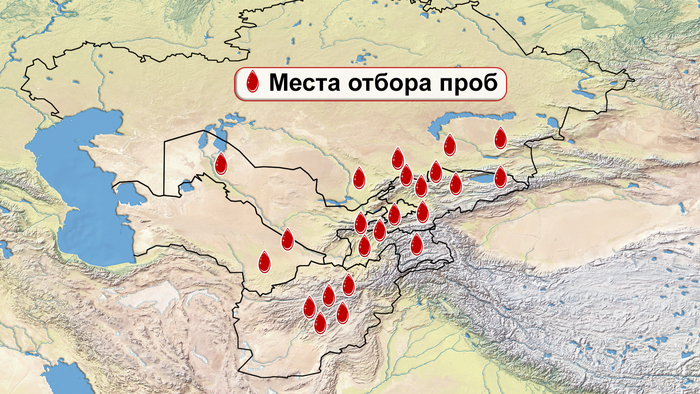
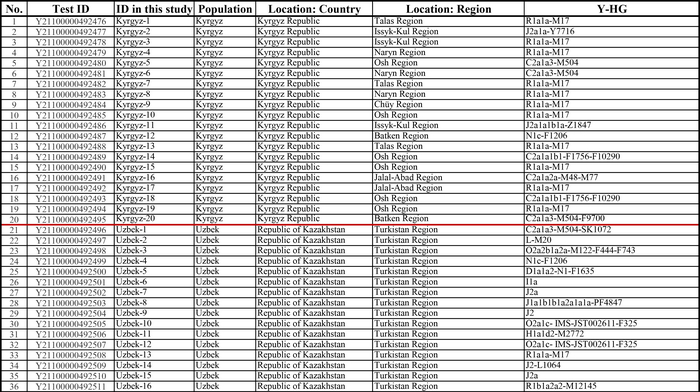
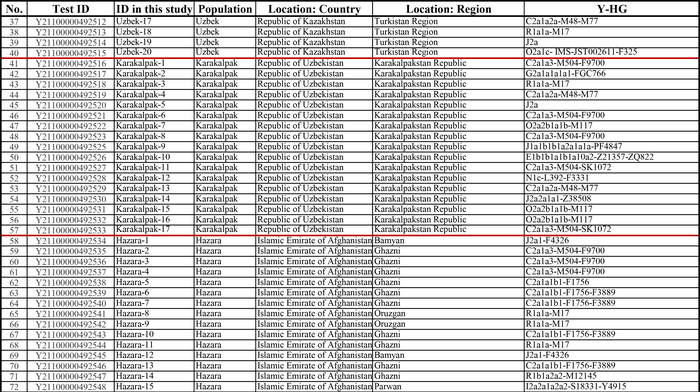
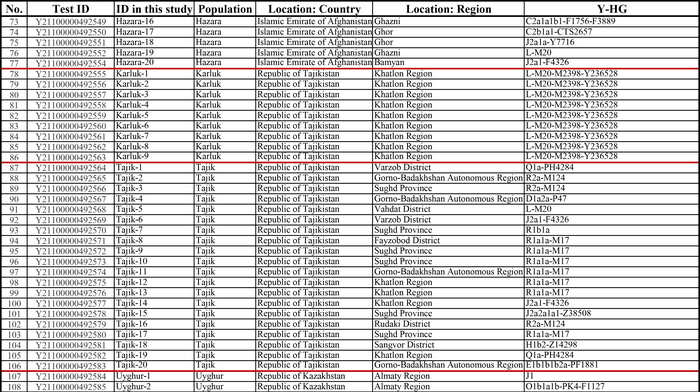
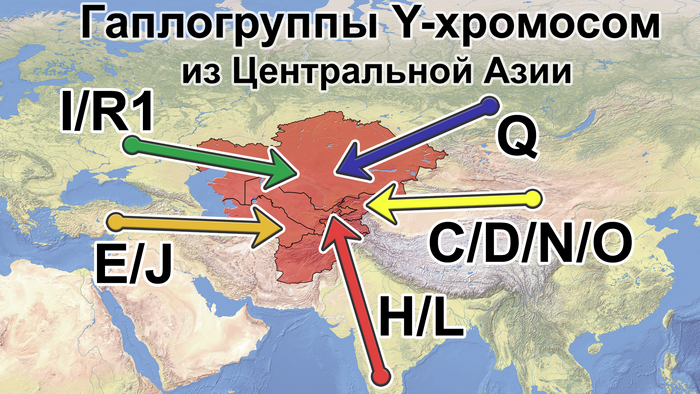
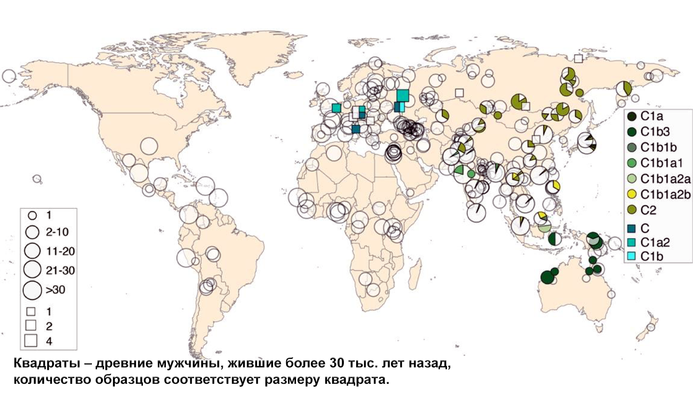
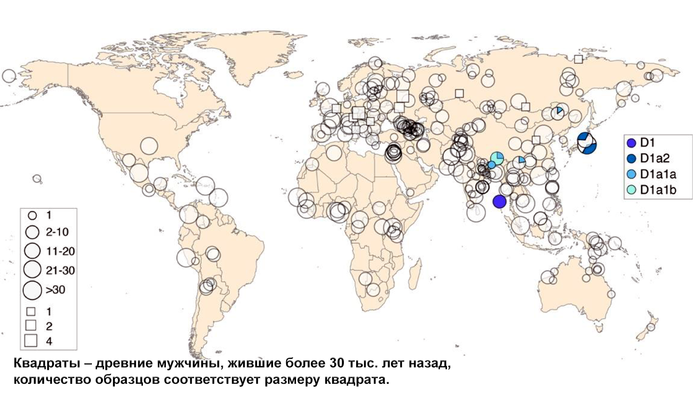
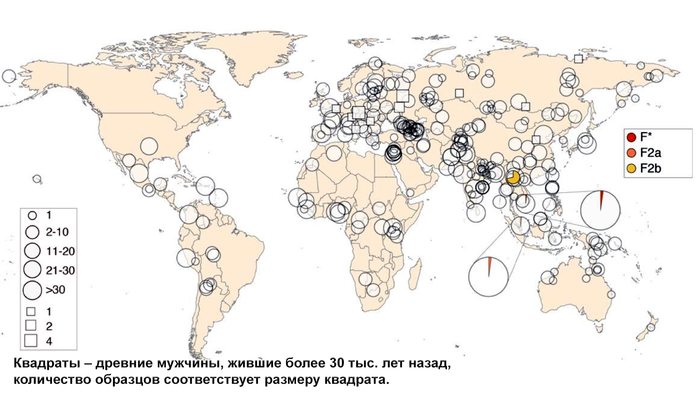
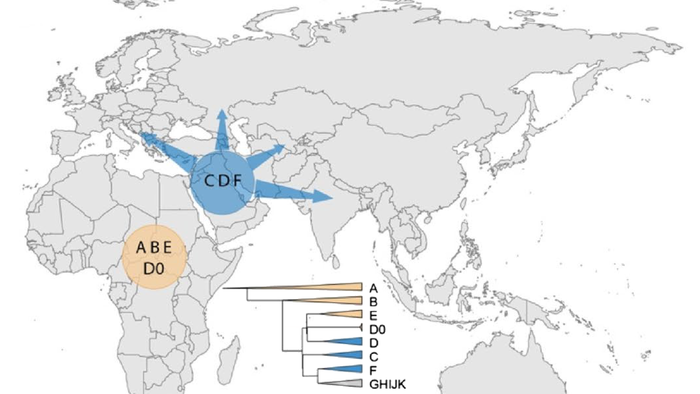
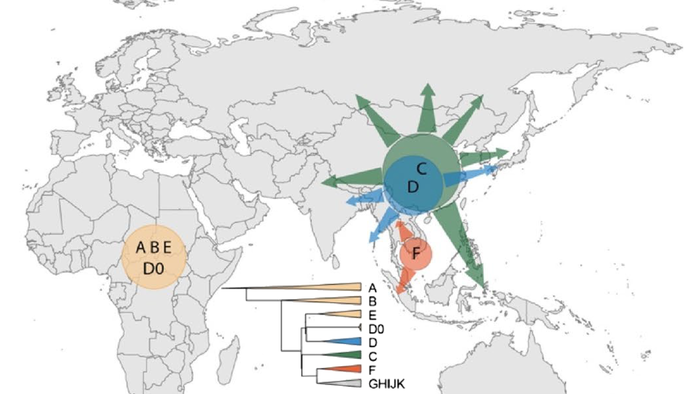
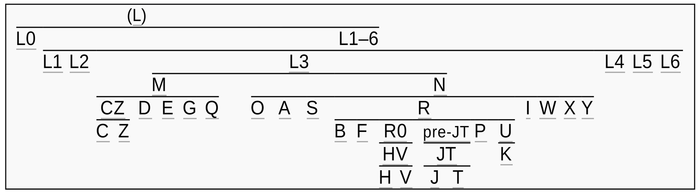
Население Центральной Азии, проживавшее вдоль Великого Шелкового пути, весьма разнообразно, оно подверглось генетическому и культурному влиянию тысячелетнего взаимодействия между Востоком и Западом.
Генетические исследования останков бронзового века (3100-1300 гг. до н.э.) из Центральной Азии показывают существенные временные изменения в генетическом составе популяций, указывающие на обширные миграции и экспансию с запада на восток оседлых скотоводов из западных степей, которые сформировали однородный генофонд к концу второго тысячелетия до н.э. В железном веке (1300-900 гг. до н.э.) кочевые скотоводы распространились по евразийской степи, рассеяв скифскую культуру. Анализ древней ДНК из погребений саков и сарматов, принадлежащих к скифской культуре, демонстрирует увеличение иранского и восточноевразийского генетического притока в южных и восточных образцах соответственно. В течение первого тысячелетия нашей эры на территории современного Казахстана образовались многочисленные конфедерации и империи, которые были связаны со значительным потоком генов. Например, ориентированная на мужчин экспансия кочевников хунну на запад из восточной степи привела к значительной примеси восточноевразийских линий к центральным сакам и вытеснению индоевропейских народов, известных как кангюи и усуни.
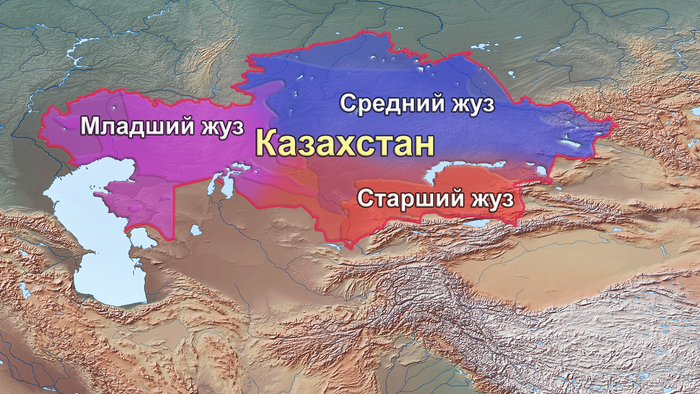
Впоследствии сформировались и смешались друг с другом разнообразные тюркские кочевые государства, что привело к потокам генов между разнородными популяциями бывшей Империи гуннов. После монгольского нашествия на территорию в 1211 году была создана Золотая Орда, которая в последующие века подверглась ряду раздроблений, что привело к созданию Казахского ханства (1465–1847). В течение этого времени кочевые племена разного происхождения проживали по всей территории современного Казахстана, и в конечном итоге они были организованы в три жуза или социально-территориальные объединения, основанные в преимущественно по географическому признаку: Старший жуз, Средний жуз и Младший жуз.
Кочевое общество казахской степи было организовано на основе иерархической патрилинейной клановой системы генеалогических линий. Люди одной и той же генеалогической линии утверждают, что имеют общего предка, и несколько генеалогических линий объединяются в рода, которые в совокупности образуют племена.
12 племен Старшего жуза в основном занимали Южный и Юго-Восточный Казахстан, семь племен Среднего жуза проживают в Восточном, Северном и Центральном Казахстане, в то время как три племени Младшего жуза традиционно проживали в Западном Казахстане. Некоторые степные рода не входили в состав жузов, особенно духовенство (кожа и сунак) и аристократия (торе́). Представители родов кожа и сунак связывают свою родословную с исламскими миссионерами, происходившими от родственников пророка Мухаммеда по отцовской линии. Люди Торе утверждают, что являются прямыми потомками Чингисхана. В отличие от оседлого земледельческого населения Центральной Азии, казахи практиковали экзогамные браки в пределах семи поколений, а партнер выбирался из другого рода, при этом женщины интегрировались в род своего мужа.
Несмотря на глобализацию последних веков и переход к оседлому образу жизни, родоплеменная структура казахов сохранилась, и многие современные казахи знают племенную принадлежность и историю своего рода. При этом учитывая, что родоплеменная принадлежность передаётся по мужской линии, анализ генетических маркеров Y-хромосомы может улучшить наше понимание мелкомасштабной демографической динамики популяции. Стоит отметить, что в случае с казахами, гаплогруппы используются для конкретных задач происхождения родов, а не для общей популяционной истории. Потому как по всему геному, как и по составу предков, казахи из всех жузов практически не отличаются ведь в основе происхождения казахов лежит сложный процесс смешения популяций Евразии, включая различные западные, южные и восточные группы, говорившие на разных языках.
В очередной работе, авторы представляют результаты исследования 1171 гаплотипа Y-хромосомы среди всех существующих казахских племен, включая торе и кожу, с целью оценки взаимосвязи племен и жузов среди казахского народа современного Казахстана.
Стоит отметить, что ожидаемые расширенные данные по среднему жузу авторы обещают опубликовать позже.
Результаты
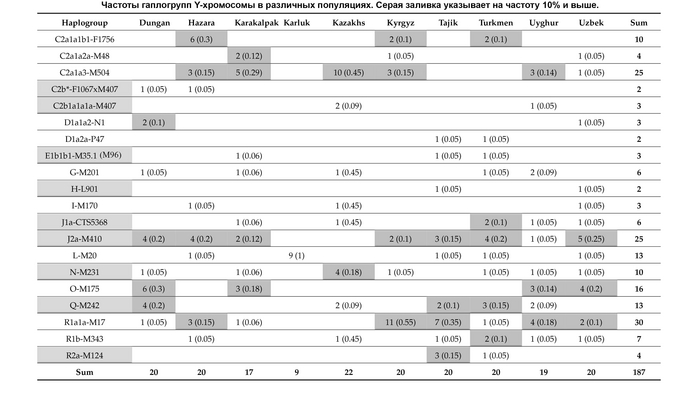
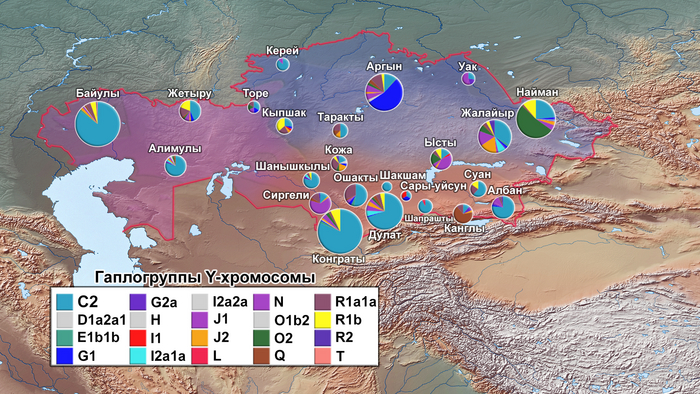
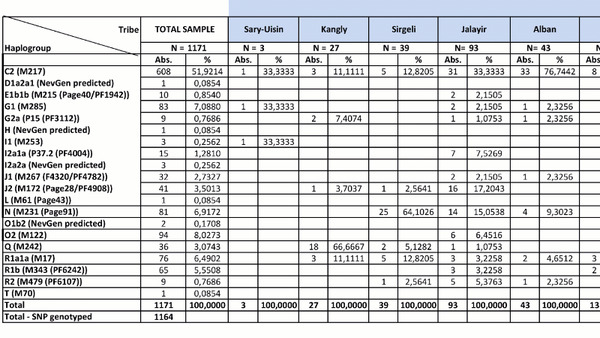
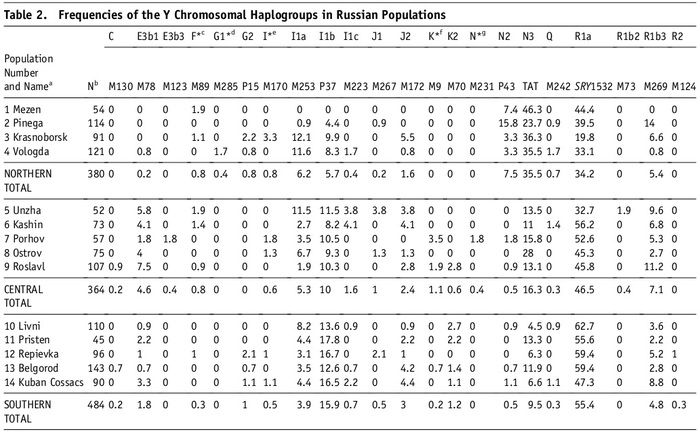
Наиболее частой гаплогруппой в казахском населении была C2-M217 (51,9%). Эта гаплогруппа присутствовала во всех исследованных племенах, и ее частота варьировалась от 11% у канглов и аргынов до 100% у шакшамов (но от них в анализе было всего 6 образцов). Стоит отметить, что количество информации по целому геному и по отдельным маркерам, значительно отличается.
Другой важный компонент казахского генофонда представлен гаплогруппой R (12,8%), которая имеет три субклада: R1a1a-M17 (6,5%), R1b-M343 (5,6%) и R2-M479 (0,8%). Гаплогруппа R1a1a-M17 наблюдалась в 18 из 24 племен и наиболее часто встречалась у представителей племени ошакты (31%), и так называемых людей белой кости из рода кожа (31,3%). Субклад R1b-M343 был обнаружен в 12 казахских племенах и имел самую высокую частоту у кыпшаков — 36,8%. Наконец, субклад R2-M479 наблюдался в пяти племенах с низкой частотой и был наиболее распространен у представителей рода кожа (6,3%) и племени жалайыров (5,4%).
Гаплогруппы O (представленные O2-M122 и O1b2, последняя предсказана проектом НевГен), G (G1-M285 и G2a-P15), N-M231, J (J1-M267 и J2-M172) и Q-M242 наблюдались с частотами <10% в разных племенах, но были обнаружены с более высокими частотами в одном или нескольких племенах. Например, гаплогруппа O2-M122 в целом в отобранных образцах встречалась с частотой 8,03%, но при этом была обнаружена у 52,3% найманов. Гаплогруппа G в общем встречалась с частотой 7,9%, причем большинство (7,1%) принадлежало к линии G1-M285, а 0,8% к G2a-P15, но значительная разница была между племенами, и ≈54% мужчин из племени аргын имели гаплогруппу G1-M285. Наибольшая частота гаплогруппы G2a-P15 наблюдалась в племени уак (18,2%). Гаплогруппа N-M231 имела глобальную частоту ≈6,9%, но была распространена в племенах сиргели (64,1%), уак (45,5%), племени торе (16,7%) и племени жалайыр (15,1%). Гаплогруппа J, представленная субкладами J1-M267 и J2-M172, имела глобальную частоту ≈6,2%, часто встречаясь в племени ысты (J1-M267 (39,4%) и J2-M172 (3%). Кроме того, J2-M172 также обнаружена у рода кожа (18,8%).
Наконец, гаплогруппа Q имела низкую общую частоту ≈3,1%, но была широко представлена у канглов (66,7%), в то время как ее частота составляла <5,5% во всех других племенах. Другие гаплогруппы показывают частоты ниже 2% в выборке.
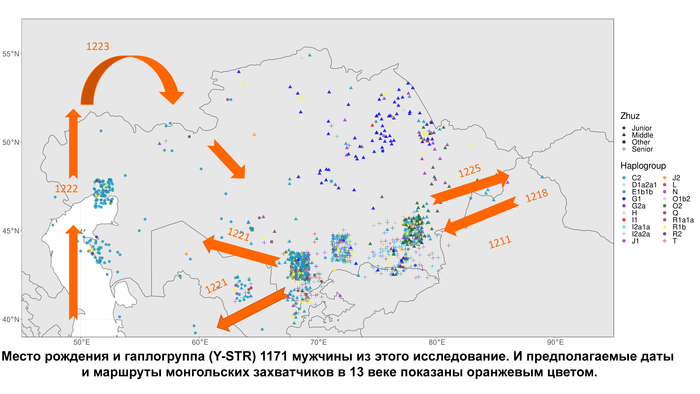
Y-гаплогруппы нанесённые на карту Казахстана отражают исторический след монгольских захватчиков, которые свирепствовали в Центральной Азии в 13 веке, поскольку гаплогруппа C2 чаще встречается в южной и западной частях страны, где прошли монголы.
Например, в южных и западных регионах Казахстана, племена албанов, конгратов, дулатов, байулов и алимулов, имеют частоты C2-M217 ≥ 70%, в то время как племена, расположенные в центре и на северо-востоке страны, такие как аргыны, уак, найманы, канглы, ысты и кыпшаки, имеют более низкие частоты (в большинстве случаев <30%).
Разнообразие гаплотипов
У 1171 человека было обнаружено 577 различных гаплотипов, из них 429 наблюдались однократно, а остальные 148 от 2 до 51 раза, при этом 15 гаплотипов наблюдались ≥10 раз.
В целом, это привело к значительному разнообразию гаплотипов Y-хромосомы – 0,9938 ± 0,0006, что отражает глубокие отцовские линии разного происхождения в выборке. Однако дифференцирующая или различающая способность выборок, основанная на распределении частот гаплотипов, составила 55,17%, что отражает высокую частоту нескольких гаплотипов. Семь из девяти наиболее часто встречающихся гаплотипов принадлежали к гаплогруппе C2. Из них два наиболее распространенных (Ht1, Ht2) наблюдались у 51 человека, в то время как третий гаплотип C2 (Ht3) наблюдался у 37 человек. Ht1 присутствовал у 38 байулов и шести алимулов, оба этих племени принадлежат к Младшему жузу, территориально расположенному в Западном Казахстане.
Ht2 был выявлен у 32 дулатов и десяти албанов, оба племени из Старшего жуза в юго-восточном регионе. Наконец, гаплотип C2 (Ht3) отмечен у 37 конгратов, Среднего жуза. Наиболее часто встречающийся гаплотип – Ht4, не относящийся к C2, принадлежал к гаплогруппе G1 и был обнаружен у 34 человек, 26 из которых принадлежали к племени аргынов Среднего жуза в Северном и Центральном Казахстане.
Структура популяции
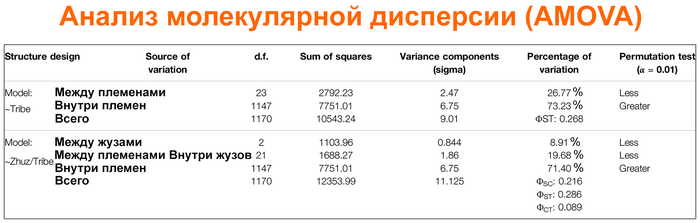
Анализ молекулярной дисперсии показал, что от ∼73% до ∼71% различий в разнообразии коротких тандемных повторов на Y-хромосоме (Y-STR) объясняется изменчивостью внутри племен.
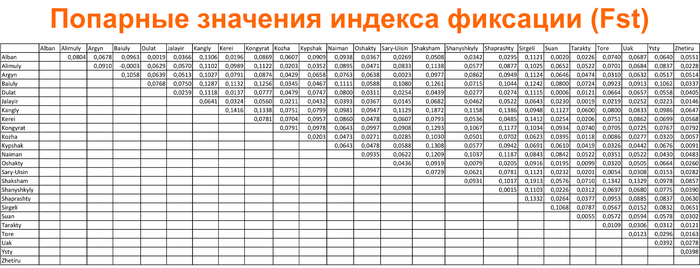
А по индексу фиксации (Fst) от 27,3 до 29,3% общего разнообразия обусловлено межплеменной дифференциацией. Попарные значения индекса фиксации между племенами варьировались от 0.0003 между байулами и алимулами до 0,19 между сиргели и шакшамами.
В таблице, для простоты восприятия, чем меньше значения, тем ближе представители этих племён друг к другу.
Помимо этого, несмотря на очевидную более высокую частоту гаплогруппы C2-M217 вдоль южных и западных границ Казахстана, отслеживающих монгольское вторжение, не было обнаружено существенной корреляции между генетическим и географическим расстоянием между людьми.
Структурный анализ
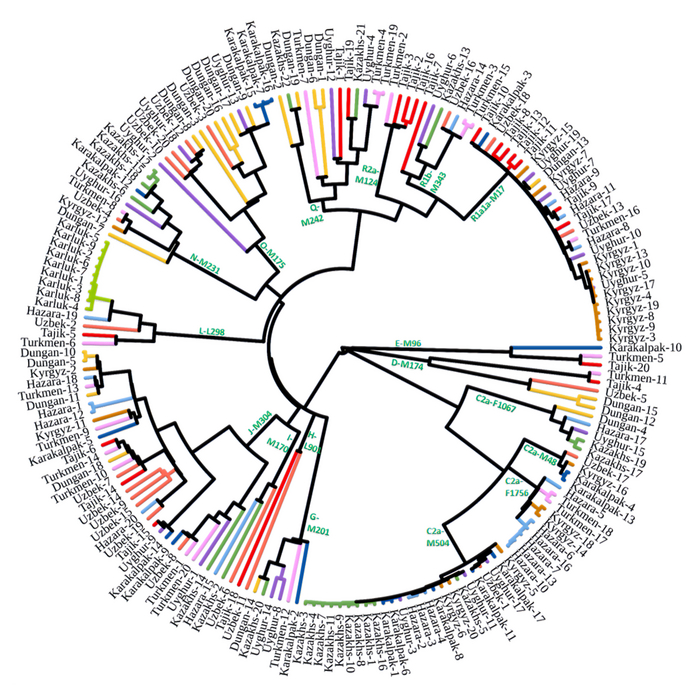
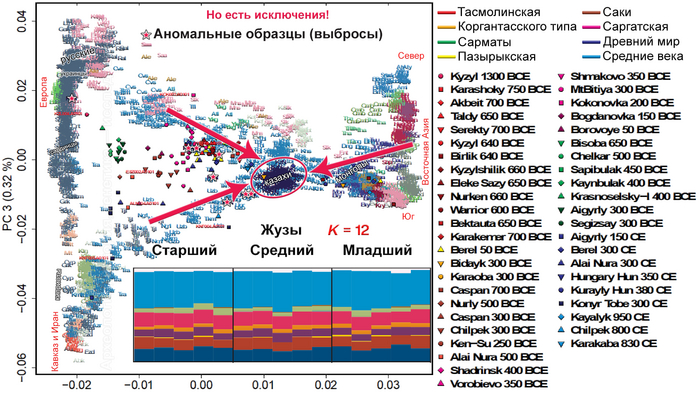
Визуализация структуры коротких тандемных повторов на Y-хромосоме по племенам и гаплогруппам подтверждает наличие по крайней мере трех подгрупп в пределах C2-M217: светло-зеленый (Старший жуз), бирюзово-голубой (Средний жуз) и розовый (Младший жуз).
Аналогичным образом, структурный анализ выявил наличие некоторых однородных гаплогрупп (например G1, O2 и R1a1a), которые тесно связаны с некоторыми племенами (например, G1с аргынами, а O2 с найманами).
Многомерное шкалирование
Этот анализ позволил более детально изучить взаимосвязь между племенами и гаплогруппами. Он показал, что, хотя многие из гаплогрупп попадают в середину координат с несколько отчетливой кластеризацией, гаплогруппы распределены по племенам рассеяно.
Однако, есть три отдельных кластера гаплогруппы C2-M217, которые в целом соответствуют трем подгруппам, обнаруженным в анализе структуры:
Первый встречается у многочисленных племен Младшего и Среднего жуза, в частности у байулов, аргынов, алимулов и кыпшаков, и соответствует гаплотипам розового цвета в анализе структуры.
Второй встречается среди различных представителей Старшего жуза, включая представителей албанов, шапраштов, ошактов, дулатов и суанов, все они проживают в юго-восточном Казахстане и соответствуют гаплотипам лимонно-зеленого цвета.
Наконец, третий кластер гаплогруппы C2-M217 встречается преимущественно среди представителей племени конгратов, соответствующего гаплотипу бирюзово-голубого цвета.
Другие отдельные кластеры в анализе многомерного шкалирования относятся к гаплотипу O2, который наиболее часто встречается у найманов с джалаирами и к гаплотипу G1, который встречается во многих племенах, но особенно у аргынов. Остальные гаплотипы на графике многомерного шкалирования имеют перекрывающиеся диапазоны. Это указывает на то, что между людьми с гаплогруппой C2-M217 существуют дополнительные различия в субкладах, которые требуют дальнейшего подтипирования. Таким образом, анализы демонстрируют, что существуют широкие различия между гаплогруппами среди трех жузов, в частности различия в гаплогруппе C2-M217 между жузами, а также большие различия в частотах некоторых Y- гаплогрупп среди племен.
Выводы
В этом исследовании разнообразие гаплотипов Y-STR у казахов достигло значения 0,9929, что отражает глубокие отцовские линии разного происхождения в выборке, в то время как разнообразие гаплотипов казахов из Синьцзяна оказалось ниже, возможно, из-за эффекта основателя, у мигрировавших в Китай казахов. Проще говоря, это когда на новую территорию переселилось малое количество людей и дало начало новой популяции.
Большая часть оценок генетического разнообразия, полученных для коротких тандемных повтор на Y-хромосоме, были одинаковыми для казахов из Казахстана и Китая, однако генетическое разнообразие локуса DYS448 было в два раза выше у казахов Синьцзяна.
Интересно, что наиболее частый гаплотип у казахов из Китая также является одним из наиболее распространенных гаплотипов среди казахов из Казахстана.
Гаплотип Ht8 связан с гаплогруппой O2, которая встречается с частотой 52,2% у представителей найманов, проживающих в Восточном Казахстане. Более того, казахские популяции в Алтайском крае, также характеризуются значительной долей носителей O2 (31-40%). Удивительно, но, несмотря на высокую частоту гаплотипа O2 в казахской популяции, у казахов из Северо-Западного Китая не было обнаружено мужчин с этой гаплогруппой.
В отличие от высокого разнообразия гаплотипов, дифференцирующая или различающая способность 15 локусов коротких тандемных повтор на Y-хромосоме составила всего 0,5517. Это говорит о том, что, несмотря на включение в исследование только неродственных мужчин, люди из одних и тех же или разных племен могут иметь идентичные гаплотипы, предположительно отражающие глубокое патрилинейное происхождение среди казахов. Например, различающая способность у китайцев из провинции Шаньси, Северный Китай, составила 0,9865, что указывает на высокий потенциал для генетического различия между мужчинами в этой популяции. Но у казахского населения из Синьцзяна, Северо-Западный Китай, различающая способность составляла только 0,5950.
Этот показатель в новой выборке из Казахстана также ниже по сравнению с некоторыми данными из европейских и азиатских популяций для того же набора из 15 локусов коротких тандемных повтор на Y-хромосоме.
Однако следует учесть, что показатели разнообразия варьируются в зависимости от используемых систем генотипирования коротких тандемных повтор. По мере увеличения количества наборов маркеров индексы разнообразия также увеличиваются.
Результаты анализа молекулярной дисперсии и теста Мантеля в новом исследовании подтвердили, что между жузами существует значительно меньше генетической изменчивости, чем ожидалось в рамках иерархической модели генетической структуры. Это говорит о том, что структура жуза не оказывает основного влияния на генетические отношения между казахскими племенами и что объединение в жузы было скорее условным, чем социально-территориальным, как предполагали другие авторы.
Тем не менее, примерно 10% генетической изменчивости среди казахов приходилось на вариации между жузами. Кроме того, анализ многомерного шкалирования показал некоторые различия в структуре гаплогрупп между племенами/жузами, особенно для гаплогруппы C2-M217
В анализе молекулярной дисперсии разделение генетической изменчивости внутри и между племенами показало, что ∼27% генетической изменчивости обнаруживается между племенами и ∼73% внутри племен.
Также авторы не нашли доказательств взаимосвязи между генетическими и географическими (по месту рождения) расстояниями между людьми, несмотря на очевидную более высокую частоту гаплогруппы C-M217 вдоль южных и западных границ Казахстана.
Анализ многомерного шкалирования и структуры показал, что в некоторых гаплогруппах существует значительное разнообразие. В этом плане особенно интересна гаплогруппа C-M217. Это наиболее распространенная гаплогруппа у современных казахов, но анализ, приведенный в описываемой работе, показывает, что у неё существует по крайней мере три различных субклада.
Один из них доминирует в племенах Младшего жуза (в основном у байулов с алимулами), другой у конгратов Среднего жуза и третий почти во всех племенах Старшего жуза.
В предыдущих работах в которых принимали участие люди из десяти племен Старшего жуза, пяти племен Среднего жуза и трех племен Младшего жуза, были выявлены три дочерние ветви гаплогруппы C-M217: C-M401, C-M86 и C-M407.
Авторы отмечают преобладание С-М401 в племенах Старшего жуза, С-М86 в племенах Младшем жузе, а С-М407 у конгратов Среднего жуза. Новые результаты подтверждают эти данные. Однако дальнейший углубленный анализ ряда однонуклеотидных маркеров гаплогруппы C-M217 позволит определить, какие субклады этой гаплогруппы точно присутствуют в популяции современных казахов.
В целом генетическая изменчивость между жузами отмечена на уровне ≈10% а между племенами внутри жуза ≈20%, что свидетельствует о наличии различий в структуре гаплогрупп среди казахских племен. И хотя авторы не нашли доказательств линейной зависимости между генетическим и географическим расстоянием, наблюдается более высокая частота гаплогрупп C2 вдоль южной и западной границ Казахстана, что соответствует как пути монгольского вторжения, так и приблизительному маршруту древнего Шелкового пути.
Для дальнейшего выяснения этой динамичной истории необходим более широкий пространственный и временной анализ разнообразия коротких тандемных повтор на Y-хромосоме среди казахских племен в контексте других групп Центральной Азии.
Источник:
Khussainova E, Kisselev I, Iksan O, Bekmanov B, Skvortsova L, Garshin A, Kuzovleva E, Zhaniyazov Z, Zhunussova G, Musralina L, Kahbatkyzy N, Amirgaliyeva A, Begmanova M, Seisenbayeva A, Bespalova K, Perfilyeva A, Abylkassymova G, Farkhatuly A, Good SV and Djansugurova L (2022) Genetic Relationship Among the Kazakh People Based on Y-STR Markers Reveals Evidence of Genetic Variation Among Tribes and Zhuz. Front. Genet. 12:801295. doi: 10.3389/fgene.2021.801295