


Гаплогруппы Y-ДНК из Центральной Азии. Филогения Y-хромосом в свете новых данных
Центральная Азия расположена на пути расселения людей современной анатомии с Ближнего Востока в Северную Евразию, что отражено в геномах древних людей, таких как усть-ишимский человек, обитатели Янской стоянки на севере Якутии и стоянки Мальта в Иркутская область. При этом климатические изменения, связанные с последним ледниковым максимумом (LGM) существенно изменили распределение популяций древних людей в Центральной Азии и прилегающих регионах. Древние носители ботайской культуры благодаря предковым линиям древних северных евразийцев генетически связаны с представителями стоянок Мальта в районе озера Байкал и Афонтова гора у современного Красноярска. Последующие миграции земледельцев с Ближнего Востока и скотоводов из степи, дополнили разнообразие в регионе. Распространение на восток в Центральную Азию и восточно-евразийские степи индоевропейских популяций привело к возникновению энеолита и бронзового века. Известны довольно примечательные Бактрийско-Маргианский археологический комплекс и андроновская культурно-историческая общность, предковая, как считается, ираноязычным популяциям.

Реконструкции: www.ancestralwhispers.org/reconstructions
Карасукская культура возникла в Южной Сибири и распространилась по обширным территориям, дав начало ряду преемников. Позднее подъём скифов вызвал широкомасштабные миграции и примесь через евразийские степи, включая Центральную Азию.

Реконструкции: www.ancestralwhispers.org/reconstructions
В более поздний исторический период тенденция к расселению людей с запада на восток изменилась, юэчжи, усуни и эфталиты мигрировали с востока на запад и создали ряд государств в Центральной Азии. А хунну/гунны массово мигрировали на запад и значительно изменили этническую структуру в Центральной Азии и Восточной Европе. Позже древние тюркские и монгольские племена распространились на запад и рассеялись по всей Центральной Азии, Ближнему Востоку и Восточной Европе. При этом потомки вышеупомянутых и не упомянутых, в сильно сокращенной предыстории, древних людей в течение длительного исторического периода смешивались и формировали современные популяции в Центральной Азии.
Предыдущие генетические исследования современных людей пролили свет на происхождение и генетическую структуру популяций из Центральной Азии. А анализ древней ДНК останков различных исторических периодов выявил закономерности смешения и генетическую структуру древних людей, а также их вклад в генофонды современных популяций.
В целом отцовский, материнский и аутосомный генофонды большинства популяций этого региона характеризуются высоким разнообразием и примесью с популяциями Ближнего Востока, Европы, а также Южной, Восточной и Северной Азии. Также у популяций этого региона выявлена сильная племенная структура, которая привела к значительным различиям в доминирующих отцовских линиях между племенами. Однако внутри племён разнообразие коротких тандемных повторов на отцовских Y-хромосомах (Y-STR), как правило, невелико.
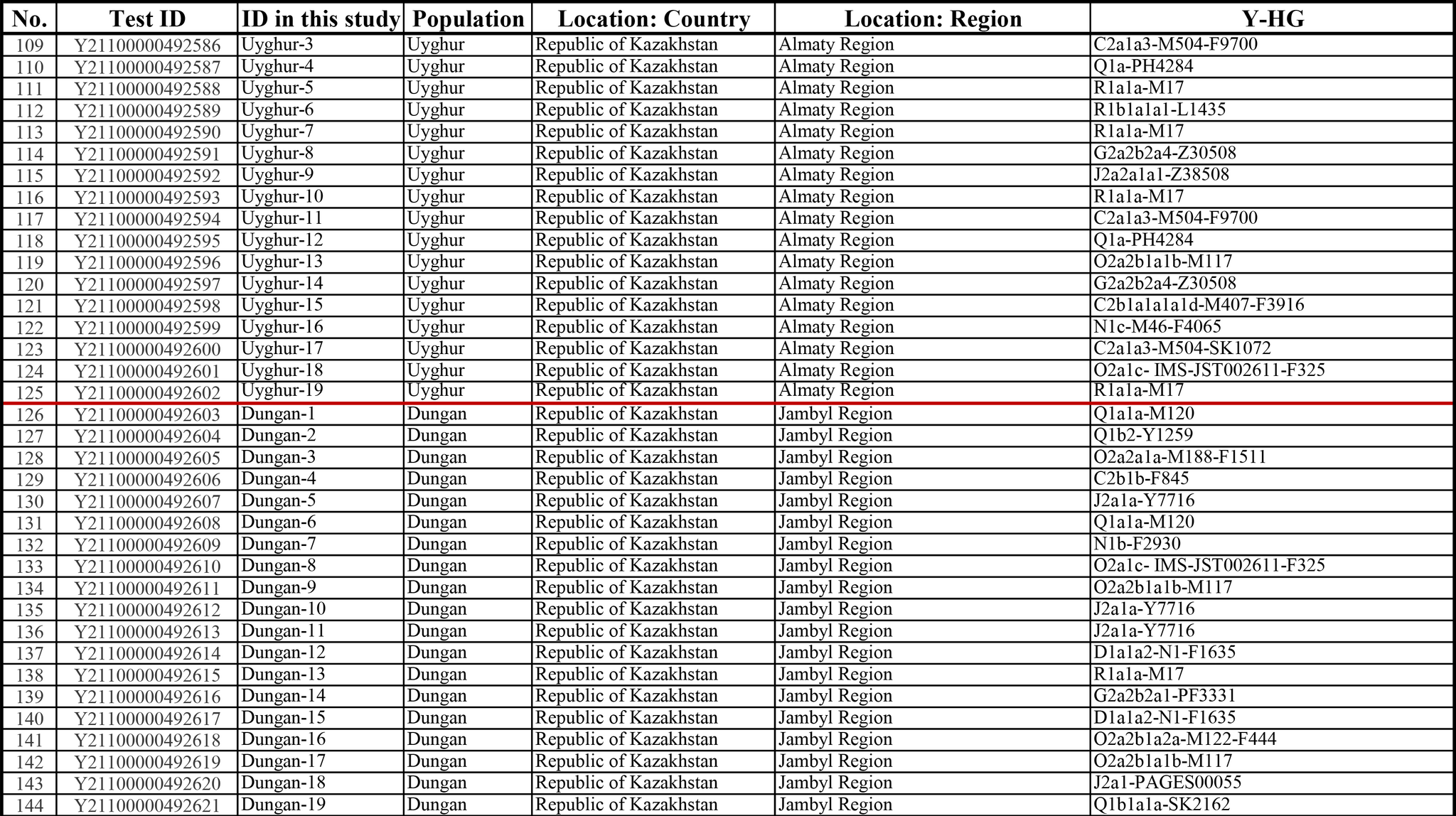
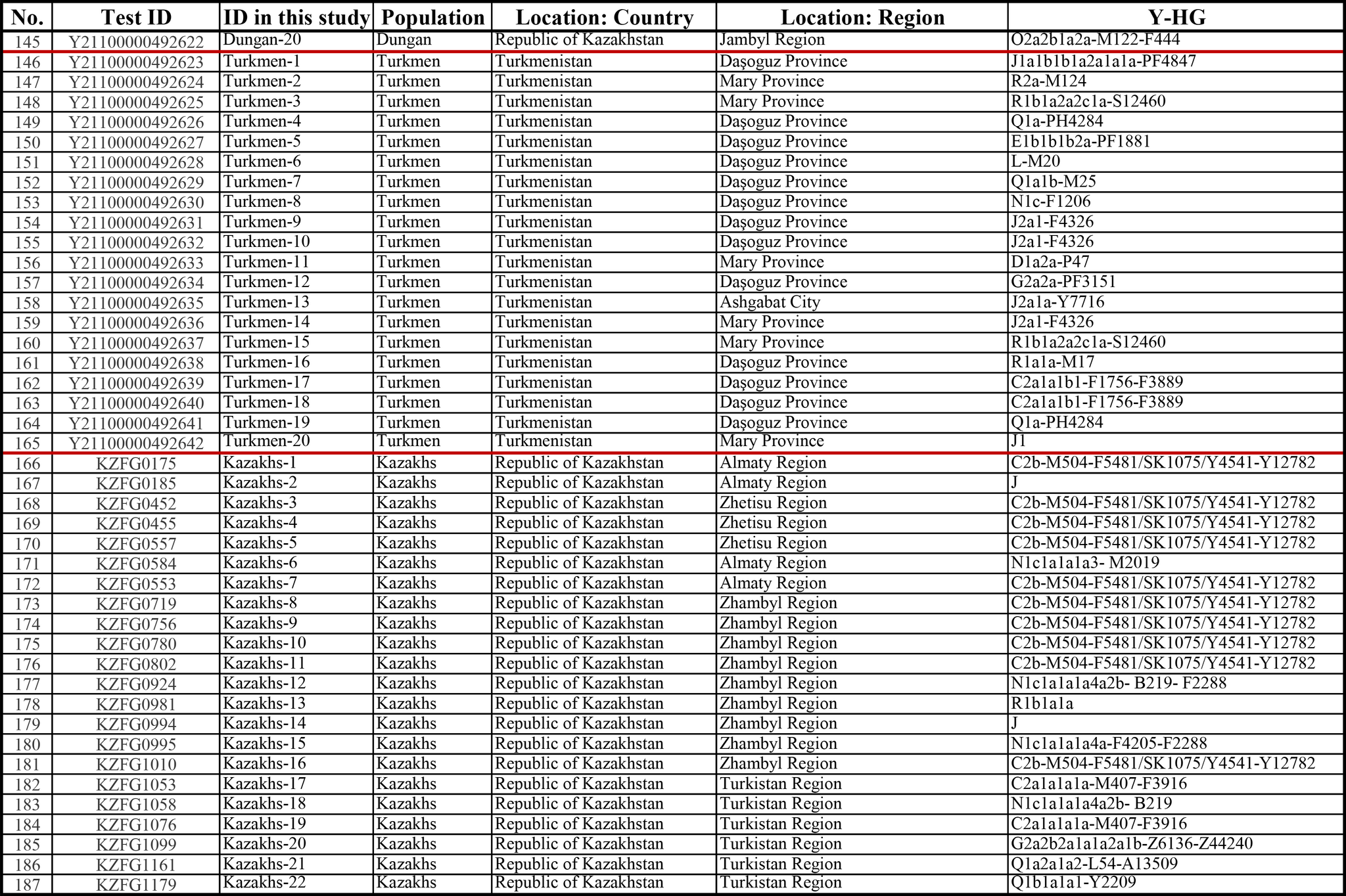
И хотя поток генов является обычным явлением, генетические взаимоотношения между популяциями по-прежнему тесно связаны с языком. Например, таджики генетически родственны другим индоевропейским популяциям, в то время как у кыргызов обнаружено много генетических компонентов из Сибири. Кроме того, были оценены последовательности Y-хромосом конкретных племен или семейных кланов для изучения формирования родственных популяций. Тем не менее, демографическая история населения Центральной Азии сложна, поэтому любые новые данные дополняют и уточняют наши знания об истории региона. Так и в новой работе авторы дополняют данные полноразмерными последовательностями Y-хромосом 187 человек. В выборку вошли казахи, туркмены, узбеки, таджики, кыргызы, каракалпаки, хазарейцы, карлуки, уйгуры и дунгане.
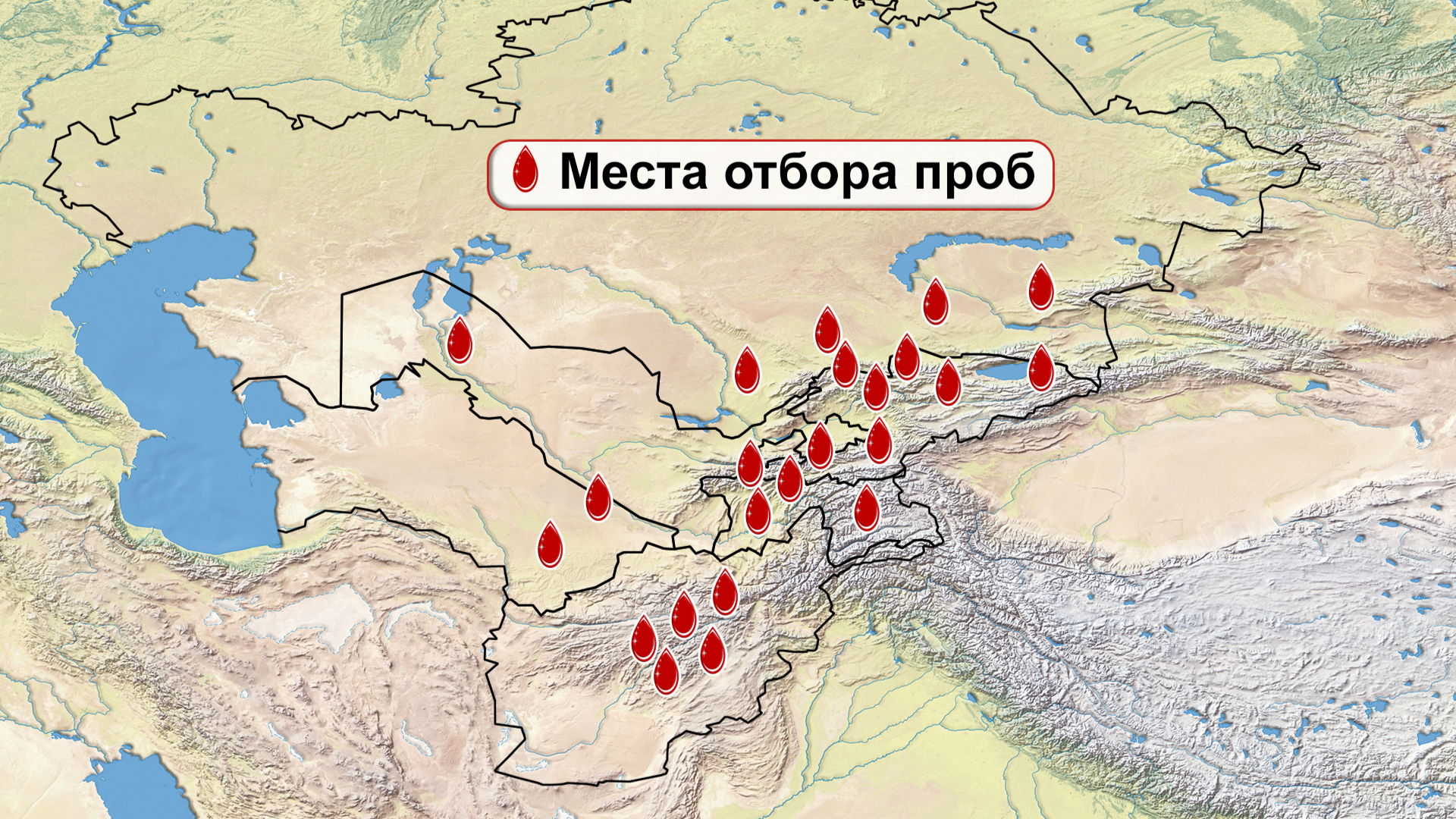
Основной целью авторов работы было выявление на основе полноразмерных последовательностей Y-хромосом общих закономерностей в отцовских генофондах. После чего идентифицировать отцовские линии, которые почти уникальны для какой-то конкретной популяции, и проанализировать их демографическую историю. В частности, исследователи оценили различные периоды генетического обмена и расхождения среди древних популяций в Центральной Азии.
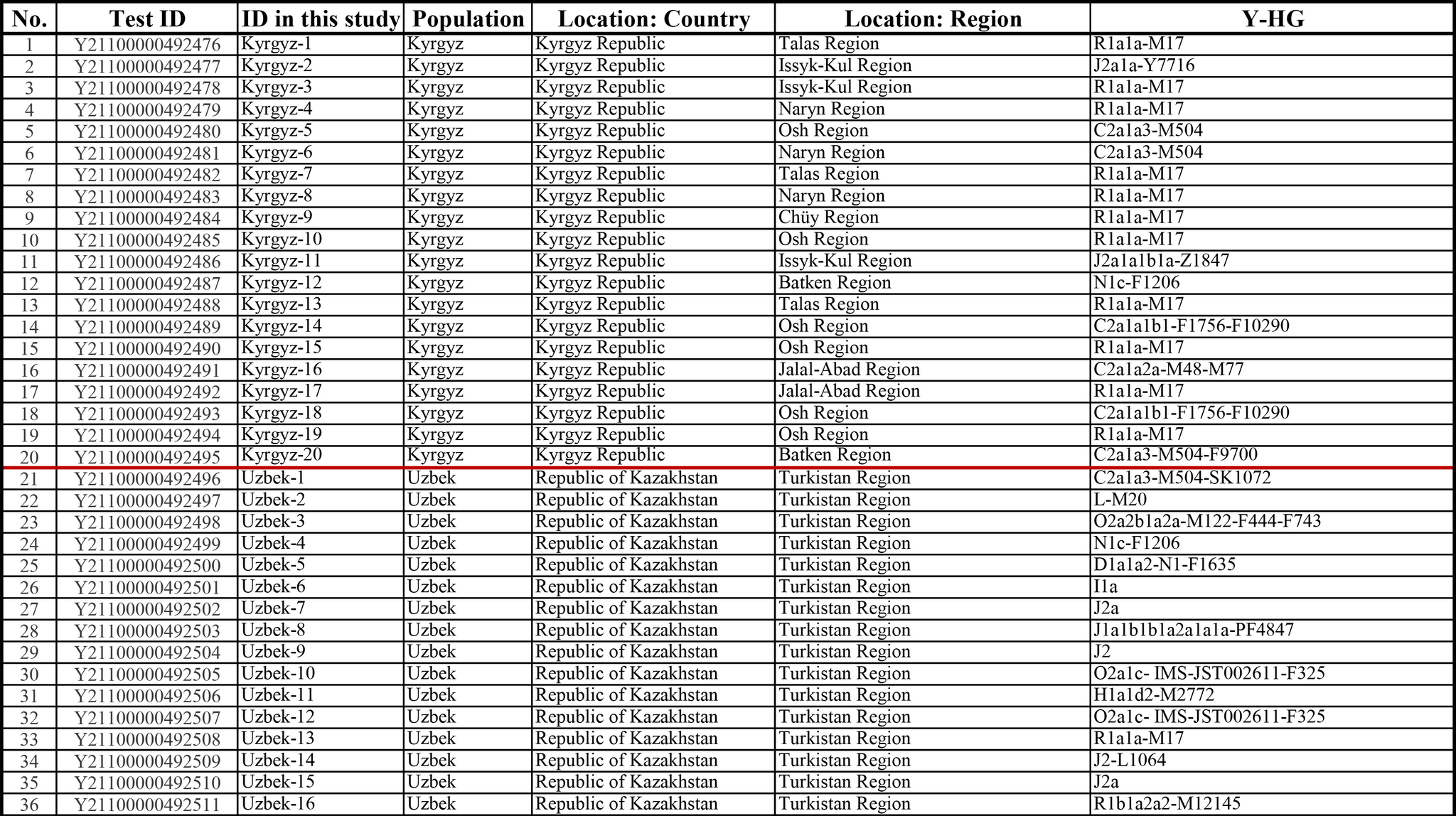
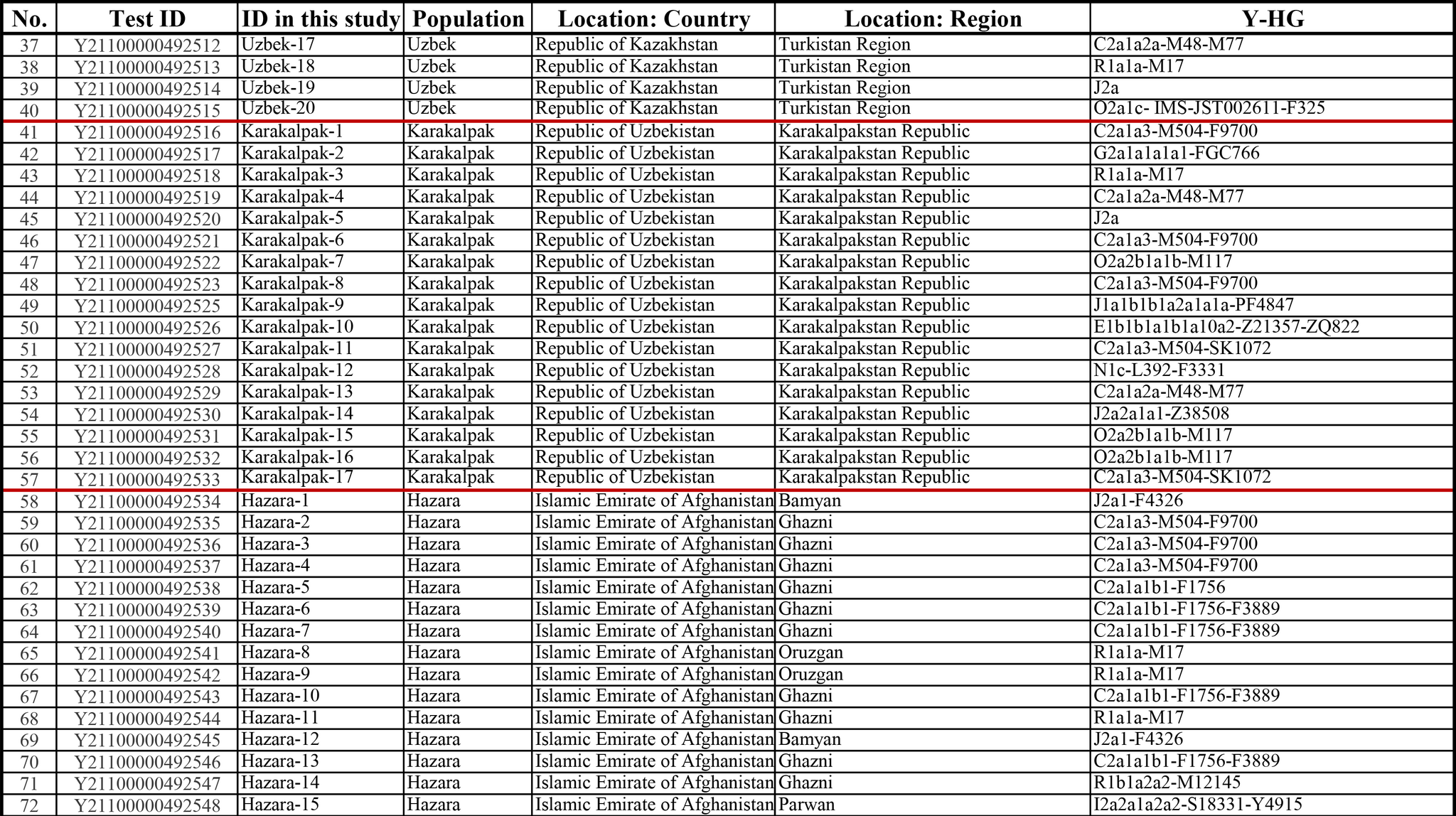
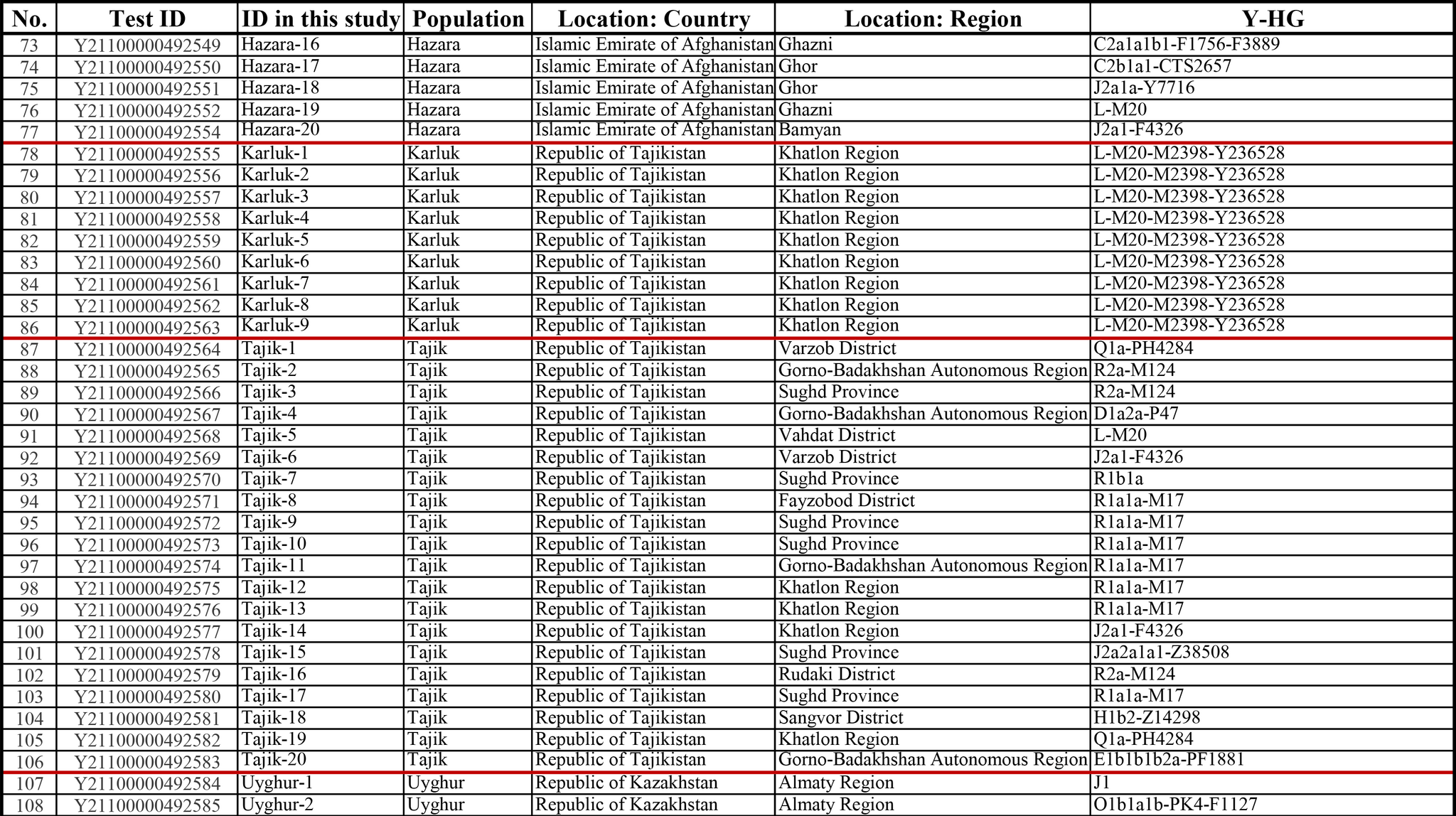
Результаты
Пересмотренное филогенетическое древо, основанное на Y-хромосомах популяций Центральной Азии
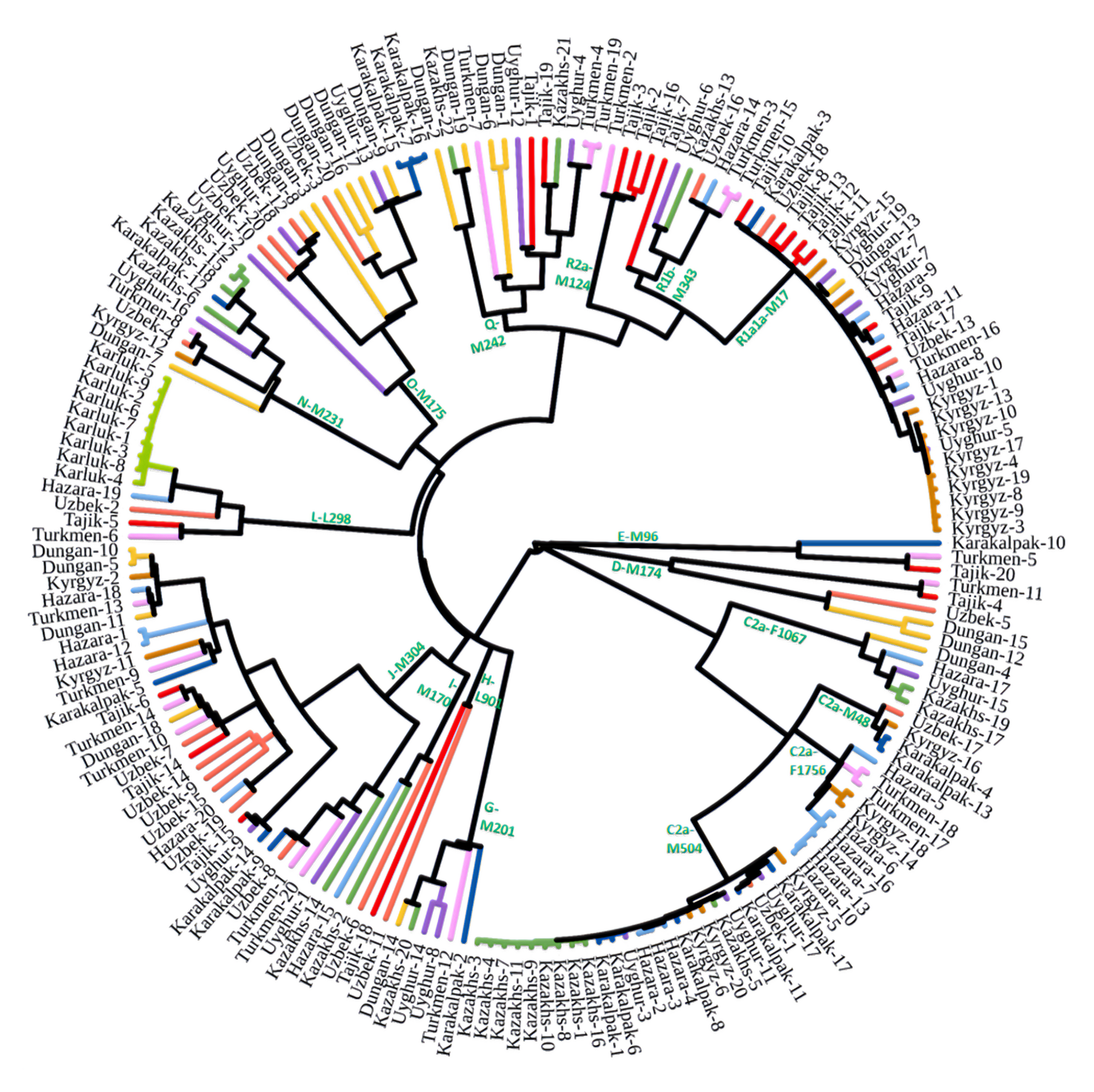
Гаплогруппа C2a1a1b1-F1756 была преобладающей отцовской линией древних племен дунху и сяньби, а появление этой линии у хазарейцев, кыргызов и туркмен указывает на то, что их предки по отцовской линии из Восточной Евразии. Гаплогруппа C2a1a2a-M48-F6379 имела высокую частоту в Младшем жузе казахов и ойратов — западных монголоязычных народов. Появление этой линии у каракалпаков свидетельствовало о близком родстве этого населения с казахами и древними монголами. Считается, что экспансия в Центральной Евразии С2а1а3- M504 напрямую связана с деятельностью древних монгольских племен в прошлом тысячелетии. В исследовании отмечена высокая частота этой линии в популяциях хазарейцев, каракалпаков, казахов, кыргызов и уйгуров, что согласуется с предполагаемым монгольским происхождением этих популяций из предыдущих исследований разных авторов. Линии N-M231 у казахов связаны с древнетюркскими племенами. А линии D-M174 и O-M175, обнаруженные в популяциях Центральной Азии, появились в результате смешения за последние 2000 лет с популяциями из Восточной Азии.
Как уже известно, гаплогруппы E-M96, G-M201, H-L901, I-M170 и L-M20 были широко распространены с низкой частотой в популяциях из Центральной Азии. Эти линии представляют собой смесь из Южной Азии, Ближнего Востока и Европы. Тем не менее, исследователи обнаружили особую сублинию L-M20 у 9 карлуков, а именно L1a2a-M2398-Y236528, которая около 1500 лет назад отделилась от линий, обнаруженных в Индии.
Предыдущие исследования показали, что Южная Сибирь, вероятно, является центром распространения гаплогруппы Q-M242, начавшейся около 30 тысяч лет назад. Кроме того, многие второстепенные сублинии Q-M242 были обнаружены в популяциях из Внутренней Евразии, включая монголо- и тюркоязычные популяции. В новой работе авторы также обнаружили различные сублинии Q-M242.
Также известно, что R1a1a-M17 и R1b-M343 являются двумя основными отцовскими линиями индоевропейского населения и большинство современных популяций в Центральной Азии по-прежнему демонстрируют высокую частоту R1a1a-M17. Пересмотренное филогенетическое дерево высокого разрешения с оценкой возраста дает подробный обзор вклада по отцовским линиям в популяциях Центральной Азии. Высокое разнообразие линий макрогаплогрупп предполагает интенсивную примесь из разных периферийных областей Евразийского континента, так линии C/D/N/O из Восточной Евразии, E/J с Ближнего Востока, H/L из Южной Азии, I/R1 из Западной Евразии и Q из Южной Сибири.
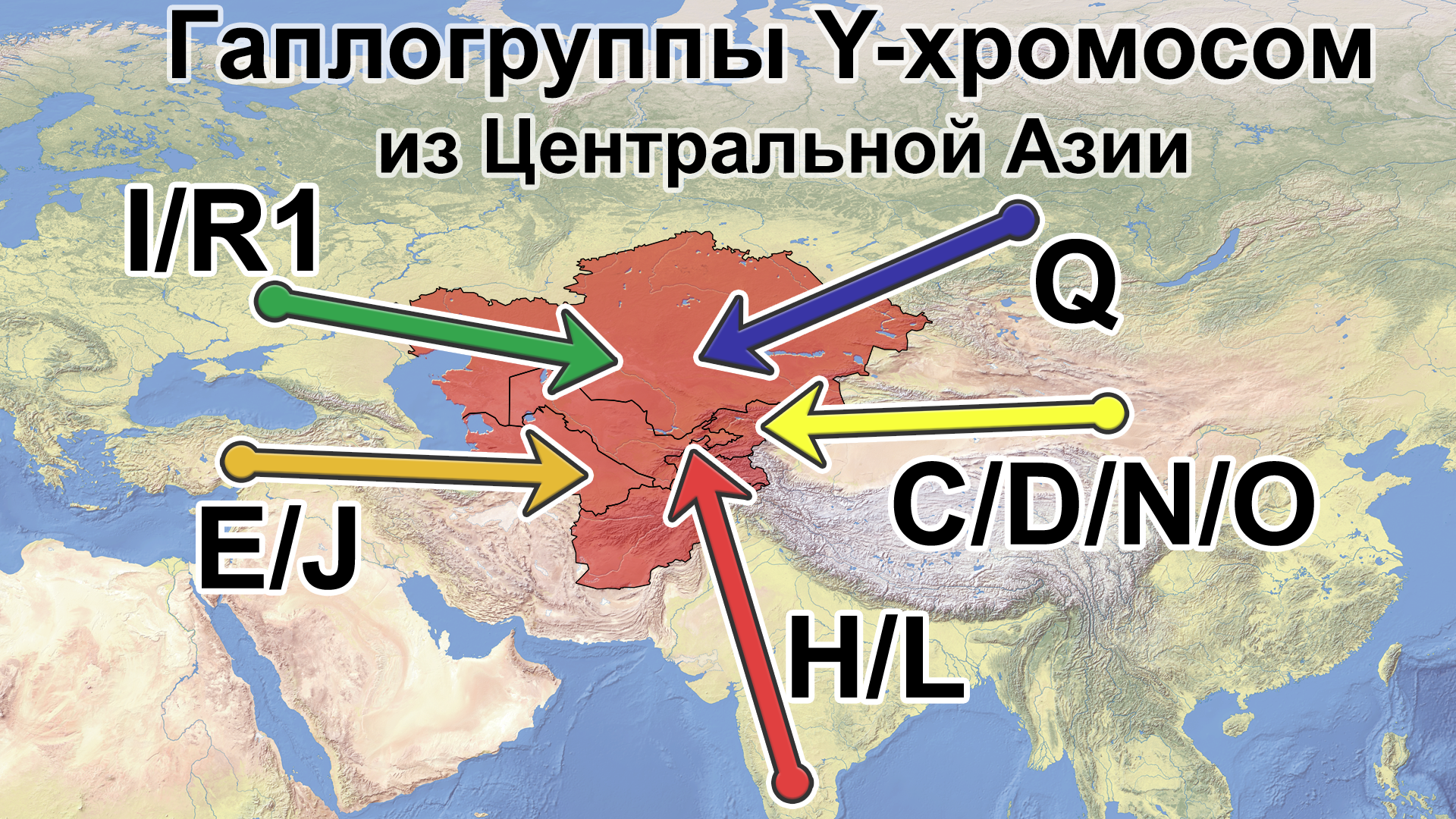
Однако наиболее важным открытием этой работы является идентификация уникальных редких, так называемых, минорных линий, которые в Центральной Азии распространились недавно. В том числе 33 такие линии моложе 2000 лет.
Выводы
Множественные события миграций и различные линии гаплогрупп
Центральная Азия не является регионом зарождения земледелия эпохи неолита или технологических систем эпохи бронзы. Поэтому роста населения, вызванного этими технологическими революциями, в Центральной Азии не было. Однако генетические данные показывают, что источниками популяций для Центральной Азии были различные регионы Евразии, включая Южную Азию, Ближний Восток, Европу, Северную и Восточную Азию. Этот общий сценарий согласуется с географическим положением Центральной Азии как центра Евразии. Причём эти миграции происходили постепенно в разные периоды. Так, неолитическая революция на Ближнем Востоке привела к глобальному распространению гаплогруппы J2-M172 и её дочерних линий, параллельно с появлением первых неолитических поселений в южной части Центральной Азии.
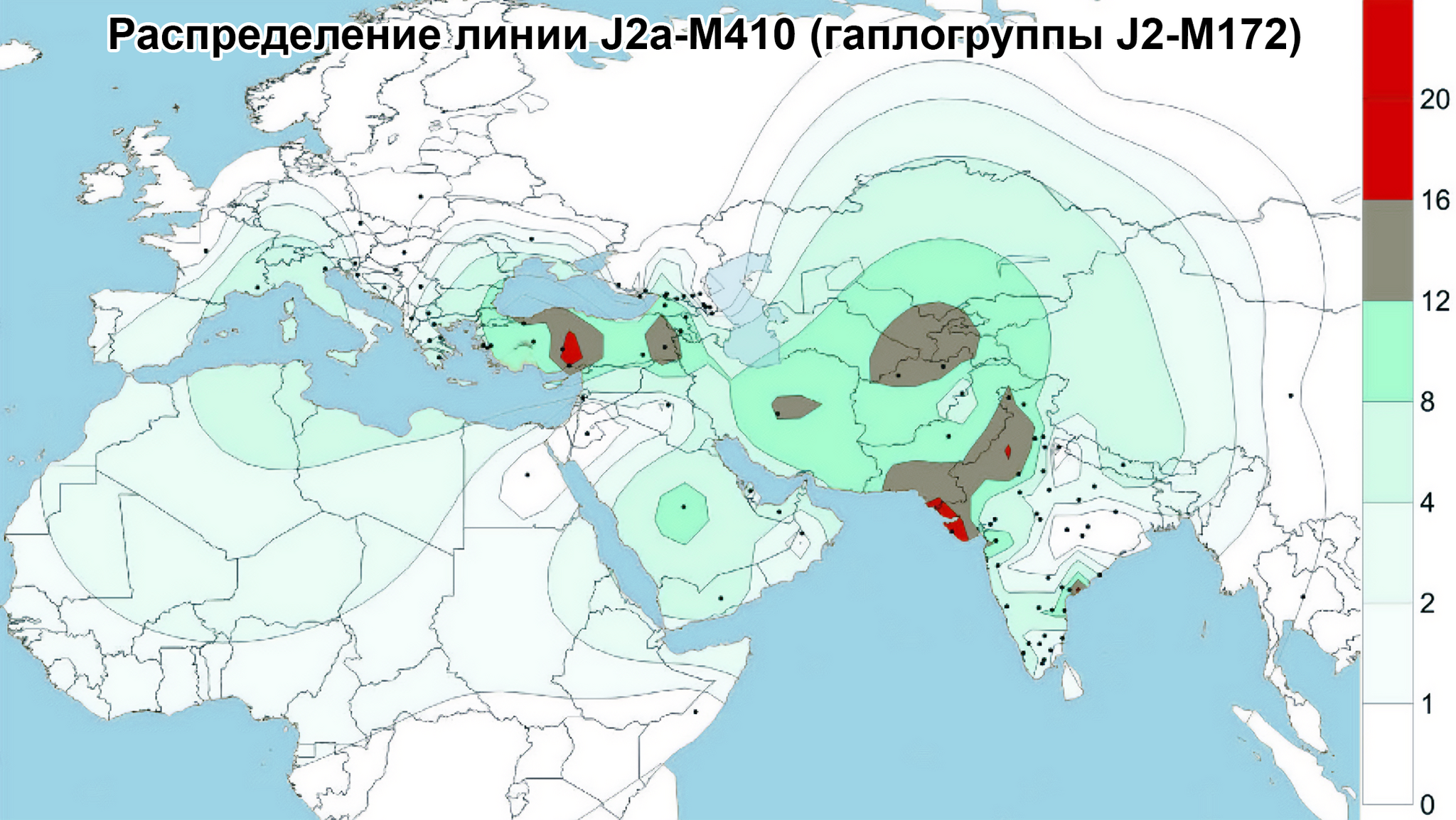
А позже непрерывная миграция с Ближнего Востока привела к появлению дополнительных сублиний J2-M172.
Многие исследования древней ДНК показали, что распространение отцовских линий R1a1a-M17 и R1b-M269 согласуется с экспансией индоевропейских популяций, и это может быть связано с разнообразными сублиниями R1a1a-M17 в Центральной Азии. Кроме того, гаплогруппа G-M201, вероятно, также происходит от индоевропейцев. Однако основываясь на ранее опубликованных генетических данных древних и современных популяций, авторы предполагают, что наличие в Центральной Азии некоторых линий Q-M242, N1a1-M46, R1a1a-M17 и C2a1b1-F1756 может быть связано с миграцией тюркских народов. А монголоязычные популяции привнесли в Центральную Азию линии C2a1a3-M504, отдельные C2a1b1-F1756 и сублинии C2a1a2a-M48. Во время миграции с Востока на Запад, на протяжении сотен лет, многие отцовские гаплогруппы распространились из Восточной Азии в Центральную Азию, включая несколько сублиний D-M174, C2a-F1096 и O-M175. В целом из-за высокой скорости мутаций Y-хромосома легко образует специфические для популяции сублинии или субклады, как кому удобнее.
Однако в комментариях люди часто неверно трактуют предковые гаплогруппы опираясь только на те популяции, у которых встречаются их сублинии. Не говоря про тех, которые им приписывают какие-то невероятные свойства.
Недавние миграции и формирование современного населения
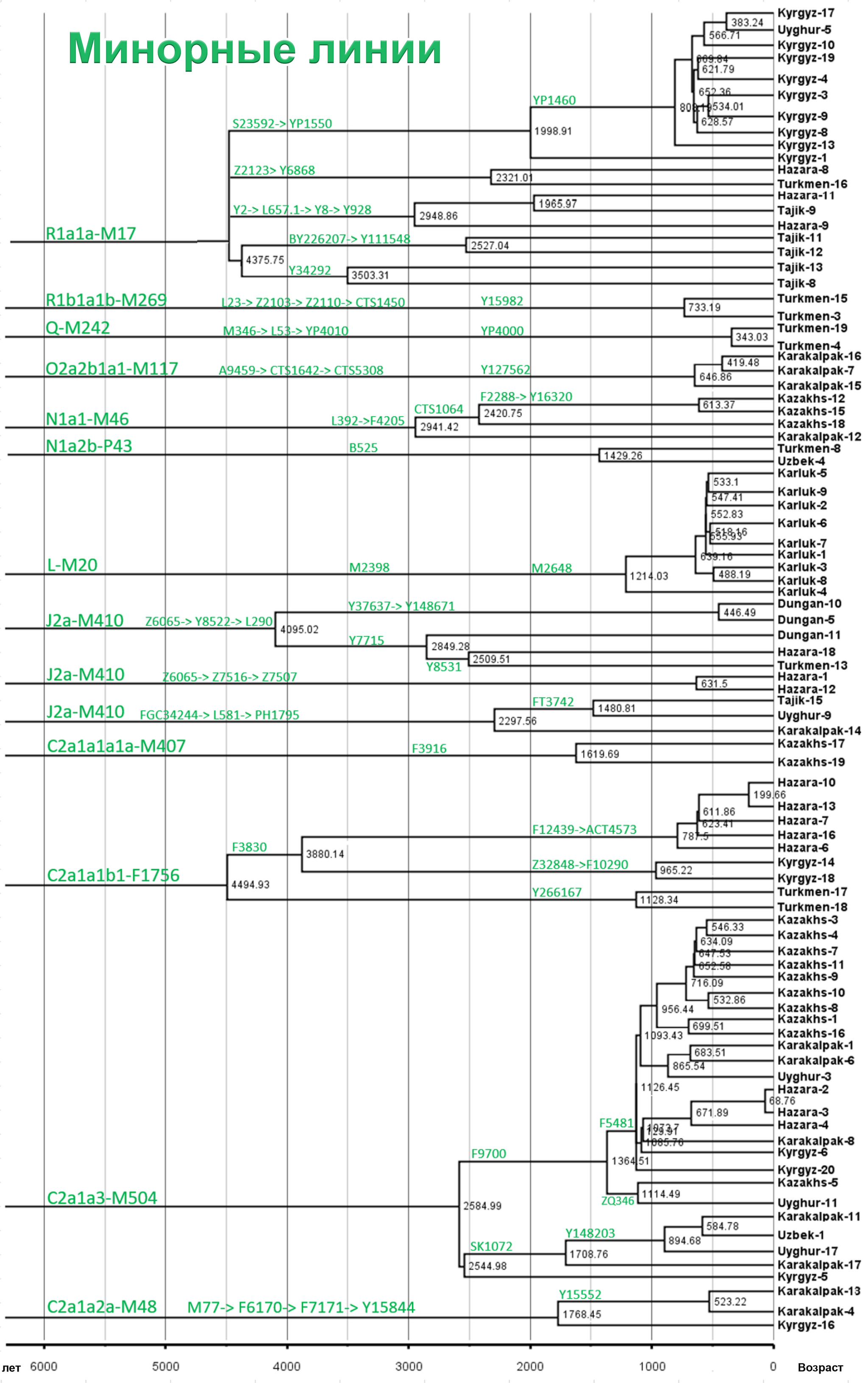
Авторы работы предполагают, что недавнее появление редких минорных предковых линий напрямую способствовало формированию современных популяций Центральной Азии.
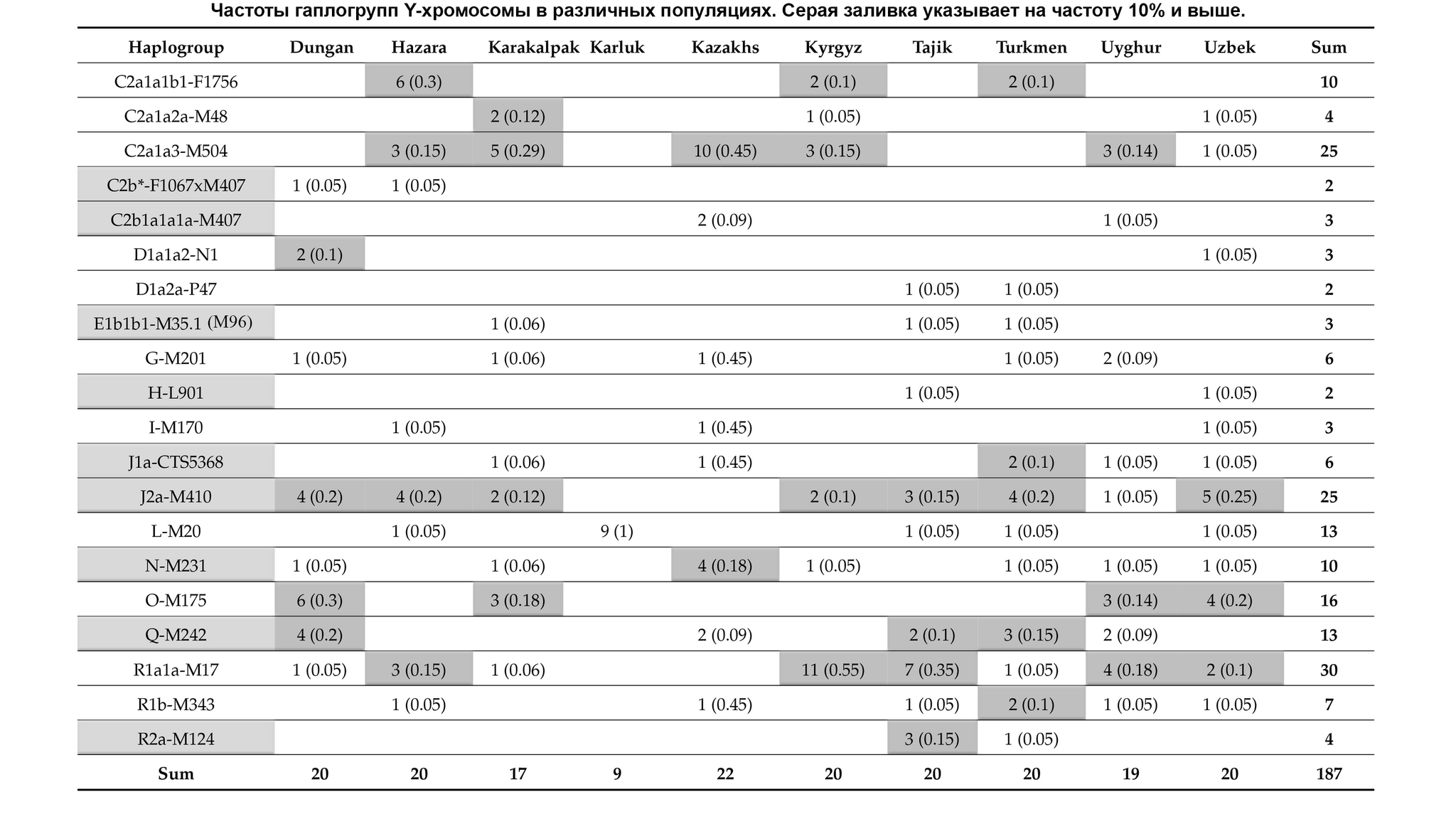
Основные отцовские гаплогруппы популяций в разных регионах Евразии включают C-M217, D-M174, E-M96, G-M201, H-L901, I-M170, J-M304, L-L298, T-M184, N-M231, O-M175, Q-M242, и R-M207. Эти макрогаплогруппы распространены и в популяциях Центральной Азии, однако их частоты существенно различаются. На уровне сублиний население Центральной Азии имеет определенные доминирующие линии.
Но как показано на графике, среди населения Центральной Азии было и много мелких минорных предковых линий характерных для некоторых этнических групп, как к примеру, R1a-M17-Y34292 и R1a-M17-Y111548 у таджиков. Наблюдается множество минорных предковых линий, которые расширились за последние 1000 лет, таких как C2a1a3-M504-F5481, L-M20-M2648 и R1a1a-M17-YP1460. Многие такие линии от R1a1a-M17 у таджиков связаны с их индоевропейскими предками. А в целом минорные линии наблюдались у казахов, кыргызов, карлуков, каракалпаков и хазарейцев.
Авторы считают, что расширение этих специфических линий напрямую связано с формированием современных этнических групп в Центральной Азии.
Таким образом, построено филогенетическое дерево линий Y-хромосом с высоким разрешением для популяций из Центральной Азии. Где значительное разнообразие отцовских линий, указывает на примесь из различных регионов Евразии в разные периоды времени. А множество минорных предковых линий указывает на недавние события примеси, которые могут соответствовать формированию современных популяций Центральной Азии за последнюю тысячу лет.
Источники: Maxat Zhabagin, Lan-HaiWei, Zhaxylyk Sabitov, Peng-Cheng Ma , Jin Sun, Zhanargul Dyussenova, Elena Balanovska, Hui Li and Yerlan Ramankulov. Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosome from Central Asia // Genes. 2022, 13, 1776. doi.org/10.3390/genes13101776
Лига историков
16.7K постов53K подписчиков
Правила сообщества
Для авторов
Приветствуются:
- уважение к читателю и открытость
- регулярность и качество публикаций
- умение учить и учиться
Не рекомендуются:
- бездумный конвейер копипасты
- публикации на неисторическую тему / недостоверной исторической информации
- чрезмерная политизированность
- простановка тега [моё] на компиляционных постах
- неполные посты со ссылками на сторонний ресурс / рекламные посты
- видео без текстового сопровождения/конспекта (кроме лекций от профессионалов)
Для читателей
Приветствуются:
- дискуссии на тему постов
- уважение к труду автора
- конструктивная критика
Не рекомендуются:
- личные оскорбления и провокации
- неподкрепленные фактами утверждения