


Зауроподы были гораздо успешнее, чем считалось раньше

Триумф титанов: как процветали зауроподы
Динозавры с длинной шеей, известные как зауроподы, когда-то считавшиеся символами вымирания, миллионы лет процветали по всему миру.
Долгое время считалось, что зауроподы пришли в упадок в конце юрского периода, около 145 миллионов лет назад, и были оттеснены на обочину эволюции новыми и улучшенными травоядными динозаврами.
Однако недавние открытия опровергли этот сценарий, показав, что зауроподы процветали еще 80 миллионов лет, на протяжении всего мелового периода — вплоть до вымирания, положившего конец эпохе динозавров. Эти более поздние зауроподы боролись с пришельцами, включая утконосых и рогатых динозавров, когда дело дошло до борьбы за важный новый источник пищи, дебютировавший в меловой период: цветковые растения.

(фото в большом разрешении)
-
Секрет их успеха, по-видимому, заключался в сочетании черт млекопитающих и рептилий в сочетании со способностью адаптироваться к меняющемуся миру.
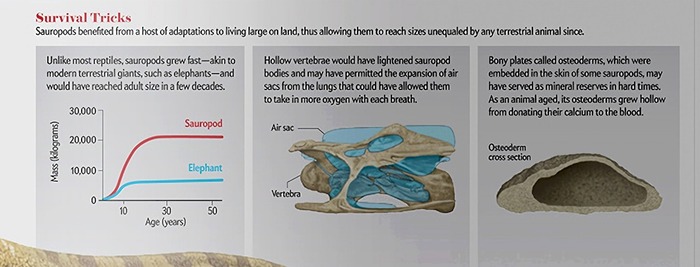
01 Зауроподы быстро росли (сравнение со слонами)
02 Полые позвонки (меньший вес и больше кислорода)
03 Остеодермы как запасы минералов в трудные времена

Показать полностью
3


Генетическая история тибетцев и их соседей, говорящих на тибето-бирманских языках (ДНК из Гималаев)
Тибетское нагорье характеризуется низким атмосферным давлением, пересеченной местностью, а также относительно низкой биологической продуктивностью и температурой. Несмотря на эти ограничения, этнические тибетцы успешно адаптировались к этой среде и жили на плато в течение тысячелетий. Понимание их генетической и культурной адаптации к этой сложной среде, со сниженным содержанием кислорода, представляет большой интерес с точки зрения археологии, антропологии, генетики и физиологии. Но чтобы это понять, необходимо ответить на многие фундаментальные вопросы, касающиеся происхождения современных тибетцев, включая место и сроки формирования предкового современным тибетцам генофонда и первоначальные маршруты миграций людей на Тибетское нагорье.
Хотя археологические данные, относящиеся к ранним миграциям на Тибетское нагорье, скудны, карстовая пещера Байшия (3280 м над уровнем моря) на его крайнем северо-восточном краю, предполагает присутствие древних людей, связанных с денисовцами, между 160 и 60 тысячами лет назад.

Датировка на археологическом участке Нвия Деву в центральной части нагорья (4600 м над уровнем моря) предполагает присутствие современного человека между 30 и 40 тыс. л. н. Однако это признаки присутствия, а не постоянного проживания, к которым некоторые авторы относят следы охотников и собирателей в Чусанге (4270 м над уровнем моря) 12700 - 7400 лет назад. И то не все учёные с этим согласны, некоторые из них утверждали, что постоянное заселение центральной части нагорья было невозможным до появления земледелия на основе ячменя около 3,6 тыс. лет назад. Последняя модель предполагает, что сельское хозяйство было привнесено на Тибетское нагорье мигрантами с более низких участков (<2500 м над уровнем моря) вдоль северо-восточных окраин нагорья, которые также внесли существенный вклад в генофонд современных тибетцев. Однако существуют доказательства более сложного, множественного происхождения современных тибетцев, которые подтверждаются и генетическими данными.
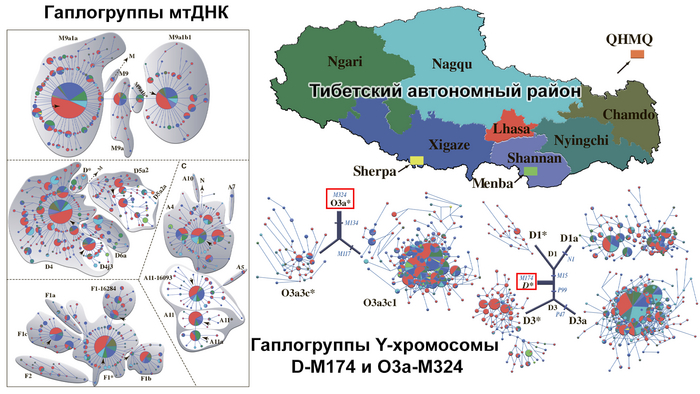
Плотно отобранные однородительские маркеры Y-хромосомы и митохондриальной ДНК, можно проследить по большей части до линий, присутствующих в северной части Восточной Азии с раннего голоцена, но более старые гаплогруппы, такие как митохондриальная M16 и Y-хромосомная D-M174, происходящие из глубокой евразийской линии, также уникально присутствуют среди современных тибетцев.
Идея древнего палеолитического вклада в тибетский генофонд также была предложена на основе данных о последовательности всего генома. Исследование, сравнивающее современные тибетские геномы с геномами древних сибиряков и архаичных представителей рода людей, выявило вклад смеси древних предков - архаичных и неархаичных — среди предполагаемых ранних обитателей Тибетского нагорья. Это предложение согласуется с наличием у тибетцев специфических аллелей гена EPAS1, которые достались им от денисовцев.
Суммарно, современные генетические данные свидетельствуют о нескольких стадиях заселении нагорья. За популяциями эпохи плейстоцена, с некоторым уровнем архаичной примеси, последовали миграции эпохи голоцена с северо-восточных окраин Тибетского нагорья.

И хотя идентичность и происхождение населения эпохи плейстоцена остаются неизвестными, недавний анализ выявил четкую клину генетической изменчивости с востока на запад в пределах современных географически рассредоточенных тибетских популяций. Эта клина может отражать неолитические миграции людей, занимавшихся выращиванием ячменя.
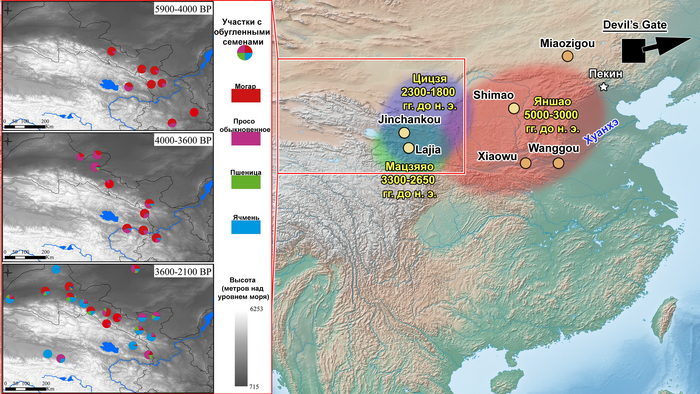
До заселения Тибетского нагорья выращивание ячменя практиковали популяции позднего неолита и раннего бронзового века из районов современных провинций Ганьсу и Цинхай, как к примеру, представители культуры Цицзя (около 2300-1800 гг. до н.э.).
Однако клина также могла быть сформирована или усилена более поздними историческими событиями, такими как расширение Тибетской империи с VII века н. э., или в результате длительного процесса потока генов между соседними популяциями с изоляцией расстоянием, не связанного с дальними миграциями.
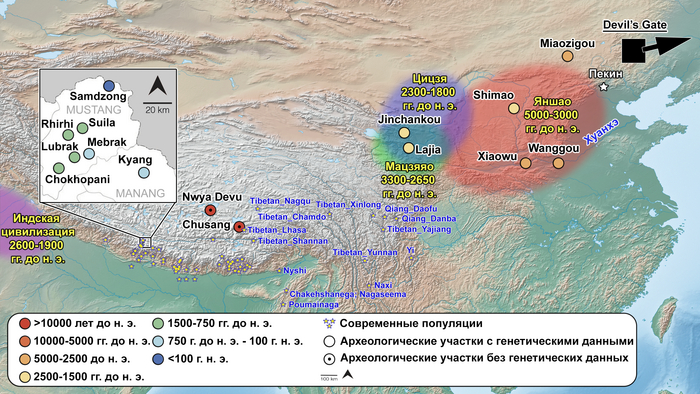
Предыдущие исследования древней ДНК людей из трех высокогорных гималайских поселений в непальском районе Мустанг датируемых 800 г. до н. э. – 650 г. н. э., показали, что эти места были заселены популяциями с явным восточноазиатским происхождением, которые, вероятно, мигрировали с Тибетского нагорья. Авторы новой работы дополняют эти данные, увеличивая временной охват более чем на 600 лет, примерно с 1420 г. до н. э. по 650 г. н. э., и предоставляя самые ранние генетические данные на сегодняшний день для популяций нагорья.
Работа демонстрирует, что эти древние гималайские популяции генетически группируются с современными тибетцами и что они представляют собой раннюю ветвь тибетской линии, что делает их особенно информативными для определения истории тибетского генофонда, его происхождения и его нынешнего распределения среди современных тибетцев, и их соседей.
А теперь обо всём более подробно:
Результаты
Древние образцы из Гималаев
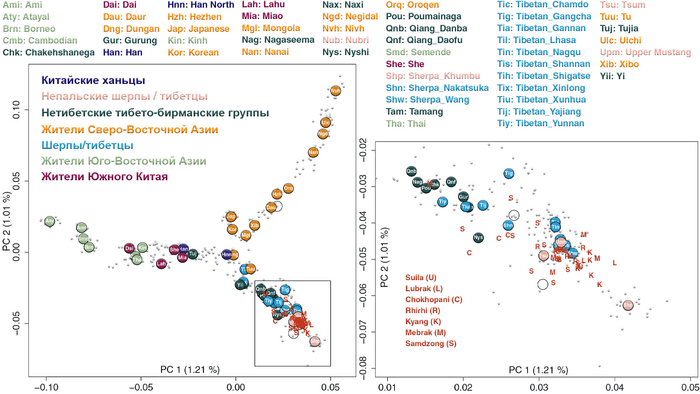
В новом исследовании авторы проанализировали общегеномные данные 38 древних людей из семи участков непальских районов Мананг и Мустанг, а именно: Suila (n = 1; 1494-1317 гг. до н.э.), Lubrak (n = 2; 1269-1123 гг. до н. э.), Chokhopani (n = 3; 801-770 гг. до н. э.), Rhirhi (n = 4; 805-767 гг. до н. э.), Kyang (n = 7; 695-206 гг. до н. э.), Mebrak (n = 9; 500 г. до н. э. –1 г. н. э.) и Samdzong (n = 12; 450-650 гг. н. э.). Новые образцы, анализировались совместно с ранее опубликованными древними геномами, а также с геномами современных шерпов и тибетцев Непала.
Генетическая структура жителей высокогорья Восточной Азии и их соседей
На графике анализа главных компонент древние жители районов Мананг и Мустанг Непала группируются с другими популяциями из Восточной Азии.
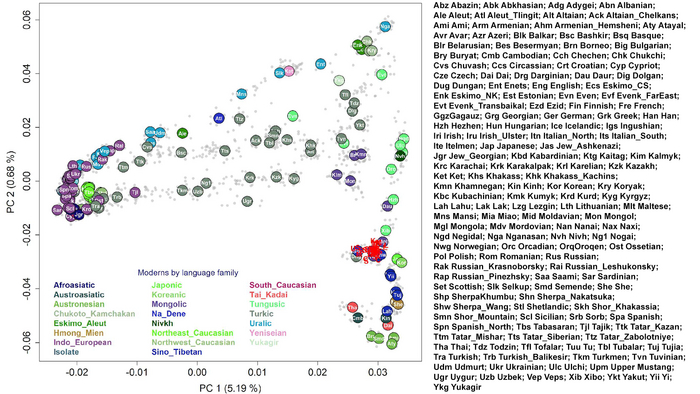
Как видно ниже на графике, современные популяции образуют структуру с тремя отростками.
Первый состоит из жителей Южного Китая и Юго-Восточной Азии, второй из представителей Северо-Восточной Азии и третий, куда собственно и попадают древние образцы, включает тибето-бирманские популяции и соответствует генетической линии современных тибетцев с востока на запад. А также они разделяют уникальные наследственные компоненты с современными популяциями из высокогорных районов и средней высоты.

Древние жители Непала, в первую очередь, генетически схожи друг с другом, а после с шерпами и тибетцами, за которыми следуют носители тибето-бирманских языков с более низких высот, такие как наси, и или йи, а также нага из Нагаленда в Индии. Однородительские маркеры, в виде гаплогрупп, подтверждают их тесное генетическое родство с современными шерпами/тибетцами.
13 из 14 мужчин имели производные маркеры Y-гаплогруппы O-M117, а 12 мужчин несли производные маркеры ее субклада Oα1c1b-CTS5308.
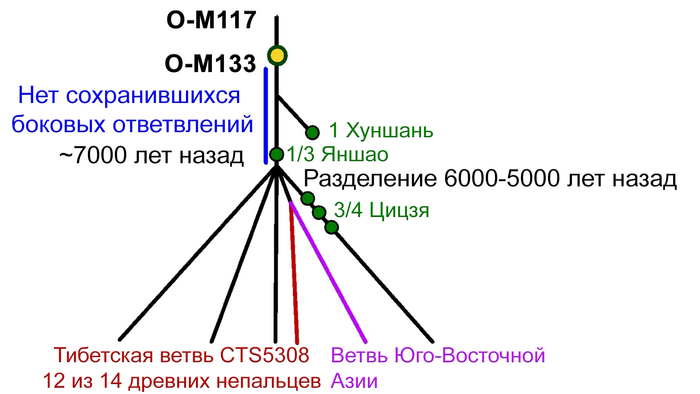
Среди современных популяций этот субклад встречается в основном среди тибетцев и шерпов Тибетского нагорья, в отличие от его родственного субклада Oα1c1b-Z25929, который сегодня встречается в основном в Южном Китае и Северо-Восточной Индии.
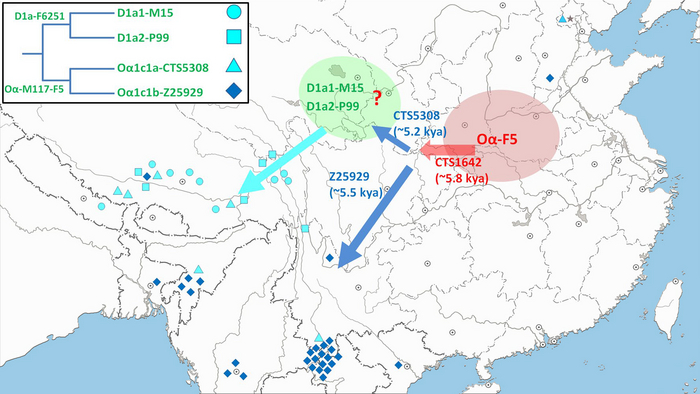
Быстрое распространение всех существующих линий O-M117, по оценкам, произошло 7-5 тыс. лет назад и было интерпретировано как отражение распространения сино-тибетских языков, вероятно происходящих из Северного Китая. Примечательно, что Y-гаплогруппа O-M117 была также обнаружена и у древних людей из неолитической культуры Яншао верхнего течения Хуанхэ и поздненеолитической Цицзя, а это свидетельствует о том, что большинство мужских линий древних жителей районов Мананг и Мустанг современного Непала восходит к региону распространения вышеупомянутых культур. Также у одного из древних непальцев была определена гаплогруппа - D1a, которая на сегодня является еще одной распространенной гаплогруппой на Тибетском нагорье.
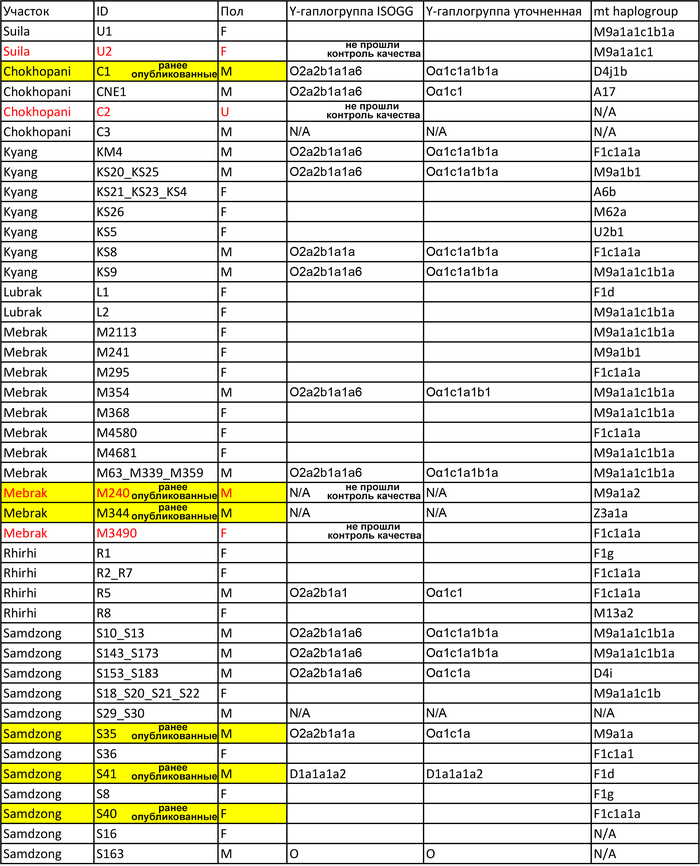
Митохондриальные линии исследованных древних людей, также распространены среди современных тибетцев, хотя они и более разнообразны.
Генетическая связь между древними и современными жителями Тибетского нагорья
Современные и древние группы районов Мананг и Мустанг, а также близлежащих районов Горкха и Солукхумбу вместе с тибетцами из более отдаленных мест, генетически ближе всего к представителям Lubrak, а затем друг к другу среди древних и современных жителей Восточной Азии. А самые древние представители из Suila, а также более поздние группы близки к современным шерпам и тибетцам в отличие от представителей Chokhopani, предки которых смешались с жителями более низких высот, вероятно ещё до 1500 г. до н. э. Таким образом, авторы делают вывод, что люди из участков Lubrak и Suila на сегодняшний день являются самыми ранними известными носителями генофонда, наиболее распространённого у высокогорных популяций Тибетского нагорья и Гималаев и называют его “тибетской” линией в этом исследовании.
Двойное генетическое происхождение жителей высокогорных районов Восточной Азии
Археологические данные свидетельствуют о том, что неолитическое население в верхнем и среднем течении реки Хуанхэ оказало значительное культурное влияние на распространение земледелия на Тибетское нагорье. Этот регион также был предложен в качестве вероятной родины сино-тибетской языковой семьи.
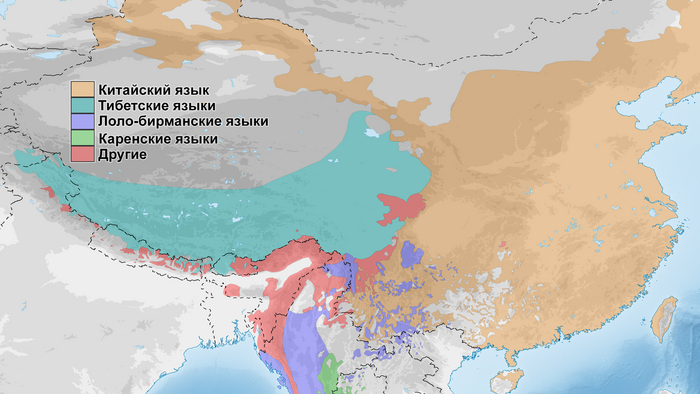
Распределение более крупных ветвей сино-тибетской языковой семьиИнтересно, что среди древних жителей низменностей Восточной Азии, группы среднего/позднего неолита из верхнего течения реки Хуанхэ и ее периферии демонстрируют наиболее близкое генетическое родство с группами из районов Мананг и Мустанг. Эти группы включают представителей поздненеолитических стоянок Цзиньшанькоу и Лацзя в верховьях реки Хуанхэ, принадлежащих к культуре Цицзя (около 2300-1800 гг. до н.э.; Upper_YR_LN), жителей стоянки Симао в Шенгедаляне провинции Шэньси (около 2250-1950 гг. до н.э.; Shimao_LN), а также представителей стоянок среднего неолита Мяодигоу во Внутренней Монголии (около 3550-3050 гг. до н.э.; Miaozigou_MN). Эти три группы имеют схожий генетический профиль, происходящий примерно на 80% от представителей среднего неолита культуры яншао участков Вангоу и Сяоу на Центральной равнине (около 4000-3000 гг. до н.э.; YR_MN) и на ~ 20% от древних северовосточных азиатов (ANA), как охотники-собиратели эпохи неолита пещеры Чёртовы ворота Приморья (“DevilsCave_EN”). Также авторы отмечают, что генетический обмен с жителями низин, вероятно, произошел до распространения ячменя.
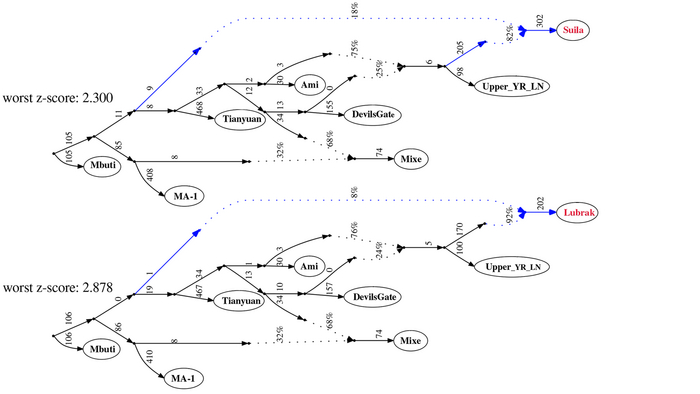
Несмотря на то, что 82-92% предков древних жителей из районов Мананг и Мустанг происходят от линии, связанной с земледельцами поздненеолитических стоянок в верховьях реки Хуанхэ, принадлежащих к культуре Цицзя, они всё же моделируются как сестринская к ним группа, как собственно и современные шерпы/тибетцы, благодаря своим уникальным генетическим компонентам, включающим аллель EPAS1 от людей в своё время связанных с денисовцами, который помогает адаптироваться к высокогорной среде.
Скорее всего, оставшиеся от 8 до 18% их предков происходят из древней популяции ещё до разделения между ветвями Западной и Восточной Евразии. При этом этот поток генов, не происходит от архаичных людей, таких как неандертальцы или денисовцы, которые вносят <0,5% родословной.
Также новые результаты отвергают и другие ранее предложенные источники потока генов в тибетскую родословную, включая глубоко разветвленные восточноевразийские линии, такие как южносибирский усть-ишимский человек, возрастом 45 тыс. лет, 40- тысячелетний древний житель Северного Китая – Тяньюань и родственные линии Хоа-Бинь/Онге в Юго-Восточной Азии.
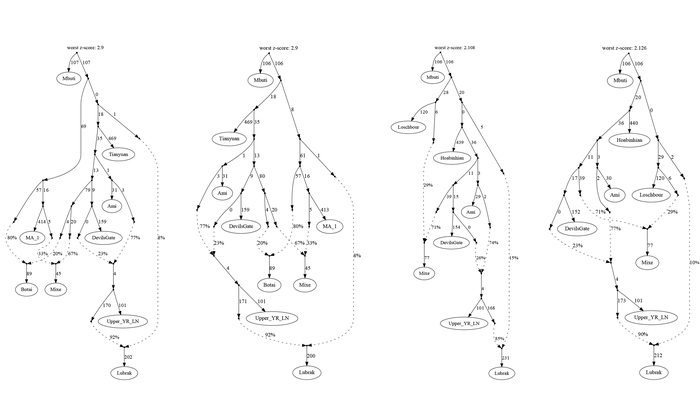
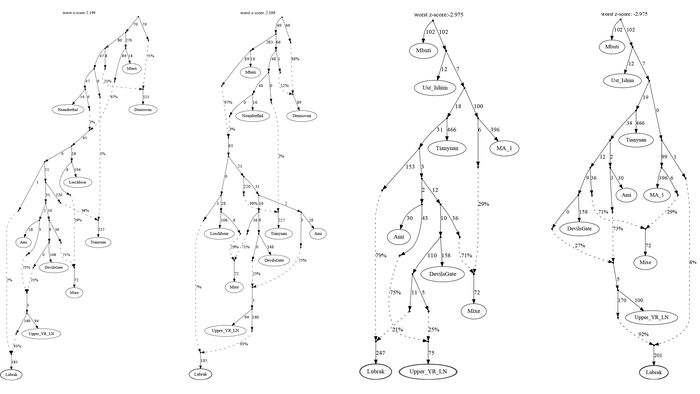
Предполагая, что эта глубокая евразийская родословная, вероятно, представляет собой ещё неисследованный палеолитический генетический субстрат от древних жителей Тибетского нагорья.
Два пути в Гималаи для носителей тибето-бирманских языков
На южных склонах Гималаев обитает множество этнолингвистических групп, которые демонстрируют поразительную структуру расслоения по высотам: индоираноязычное население Южной Азии занимает низменности, шерпы/тибетцы занимают высокогорья, а различные нетибетские народы, такие как таманги и гурунги, говорящие на тибето-бирманском языке, занимают диапазон средних высот.
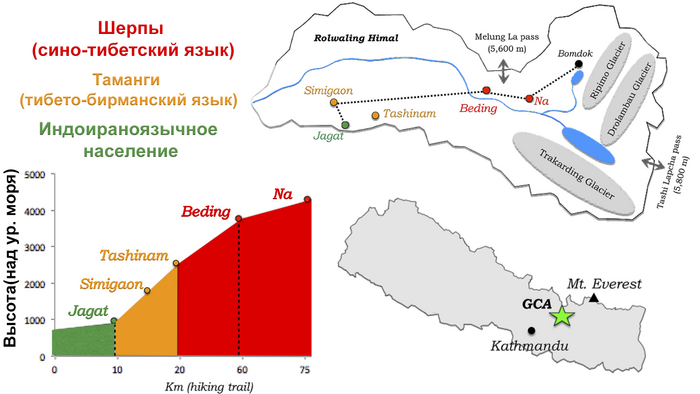
В то время как шерпы/тибетцы Непала, вероятно, прибыли в Гималаи с нагорья (т.е. северным путём), предыдущее генетическое исследование предложило отдельный южный маршрут для миграции групп тибето-бирманцев средних высот.

Однако, как нетибетские группы, говорящие на тибето-бирманских языках, связаны друг с другом и с тибетской родословной, оставалось неясным. В данной работе авторы указывают на две генетические клины: тибетскую и нетибетскую тибето-бирманскую.
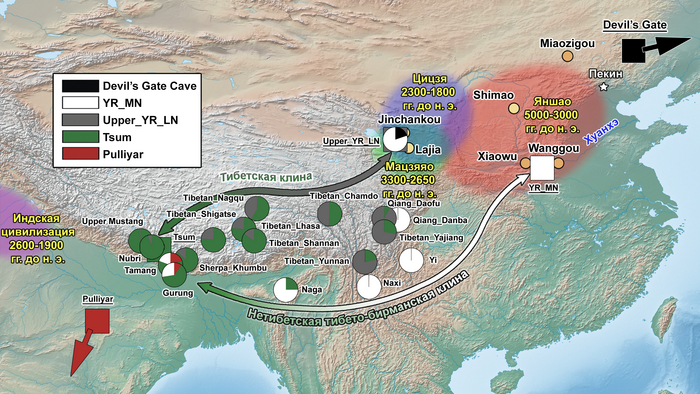
Тибетская клина соединяет древних и современных непальских тибетцев (которые в моделях представлены жителями Цумской долины) с обитателями поздненеолитических стоянок культуры Цицзя в верховьях реки Хуанхэ (Upper_YR_LN). А нетибетская тибето-бирманская клина отражает примесь вдоль маршрута вокруг Тибетского нагорья и соединяет популяции средних высот, такие как наси, йи, нага, таманги и гурунги с представителями среднего неолита культуры Яншао на Центральной равнине (YR_MN). Помимо этого, таманги и гурунги, имея более высокий вклад от тибетской линии 60-63%, дополнительно демонстрируют генетическую примесь из Южной Азии от 9 до 19%, как и популяции Бутана из предыдущих исследований.
Примечательно, что для населения Непала со значительным южноазиатским вкладом (например, барамцев, чантьяльцев, гурунгов и представителей народа чепанг), южноиндийские группы, такие как пулаяр, лучше подходят для моделирования их южноазиатского происхождения, чем соседние группы Северной Индии. Эти результаты подчеркивают сложную и многослойную историю смешения тибето-бирманских популяций в Гималаях.
Длительный положительный отбор на участках генов EPAS1 и EGLN1 у тибетцев
В своём предыдущем исследовании авторы сообщали, что производные аллели для положительно отбора в гене EPAS1 наблюдались только в менее древних образцах участка Самдзонг, в отличие от более древних жителей Чохопани и Мебрака. Проанализировав новые геномы древних жителей из районов Мананг и Мустанг, авторы по-прежнему не обнаружили производных аллелей в блоке гаплотипов EPAS1 в образцах из Чохопани и Суила, однако они наблюдались с промежуточной частотой на 5 других участках (от 25 до 58%).
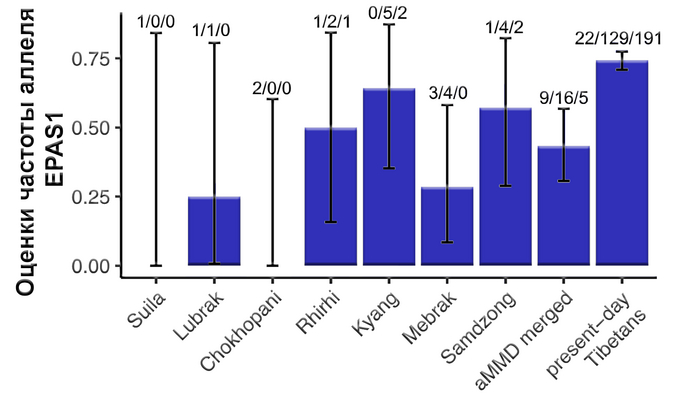
Интересно, что частота производных аллелей в древних образцах в целом ниже, чем у современных тибетцев (75%), а это указывает на то, что отбор все еще воздействовал на эти аллели в недавнем прошлом.
Итоги
В новой работе генетические данные 38 древних гималайцев указывают на то, что родословная, обнаруженная у современных тибетцев и шерпов, уже присутствовала у жителей высокогорья к 1494-1317 годам до нашей эры, отчётливо отличая их от жителей низменностей. Тем самым делая старше свидетельства существования тибетского генофонда по крайней мере на 500 лет по сравнению с предыдущими данными из Чохопани.
При этом исследование демонстрирует, что тибетский генофонд хорошо моделируется как смесь двух источников генетической родословной. Один из источников на 82-92% состоит из генетических компонентов связанных с жителями низменностей, которые проживали на северо-восточной окраине Тибетского нагорья во времена позднего неолита. А другой источник представляет собой древний и ранее неизученный палеолитический субстрат на долю которого приходится до 18%. Палеолитический субстрат, по-видимому, внес свой вклад исключительно в тибетский генофонд среди современных популяций, изученных на сегодняшний день.
Моделирование современных тибетских популяций и их нетибетских соседей, носителей тибето-бирманских языков, демонстрирует две генетические линии, которые, по-видимому, отражают два различных маршрута миграций – северный и южный, как видно на карте. Тибетские популяции смешаны вдоль северной клины, а генетическое, культурное и языковое разнообразие современных носителей тибето-бирманских языков вдоль южного склона Гималаев отражает слияние древних популяций, прибывших по этим двум маршрутам после их разделения со времен позднего неолита.
В отличие от предыдущих, весьма противоречивых работ, основанных на анализе ДНК современных тибетцев и ханьцев, результаты новой работы, благодаря древним геномам, показывают, что предки современных тибетцев присутствовали в Гималаях, по крайней мере, с 1420 года до н.э., во времена, когда на участках Суила и Лубрак появляются самые ранние прямые свидетельства постоянного присутствия человека. Помимо этого, исследование подтверждает тесную связь между ранними гималайскими популяциями и группами позднего неолита, жившими вдоль северо-восточной окраины Тибетского нагорья около 2300-1800 гг. до н.э. (Upper_YR_LN).
Эти неолитические группы с территорий современных провинций Ганьсу и Цинхай, вероятно, включали в себя популяции, которые позже заселили нагорье, однако точные даты этого события ещё не ясны.
Долгое время считалось, что выращивание ячменя, который лучше, чем просо подходит для более прохладного и сухого климата нагорья, способствовало неолитической экспансии в регион. Хотя новые результаты могут предположительно соответствовать давней гипотезе о распространении ячменя по нагорью около 1650 г. до н.э., такая массовая демографическая диффузия из районов Ганьсу и Цинхай всего за 200 лет вряд ли будет единственным объяснением древней генетической связи между Тибетским нагорьем и этими районами. Поэтому авторы предлагают альтернативный сценарий, в котором генетическая связь между популяциями нагорья и низменностей могла сформироваться намного раньше и, следовательно, не могла быть связана с внедрением ячменя или других одомашненных растений или животных западноевразийского происхождения.
Стоянка Karou в восточном Тибете (около 5000-3000 лет назад) и стоянка культуры Qugong близ Лхасы (около 3800-3000 лет назад) демонстрируют местную археологическую традицию и имеют композицию сборки и керамические мотивы, отличные от таковых в культуре Цицзя.

Karou, Qugong, ZongriКроме того, свидетельства с участка Zongri (около 2600-2000 гг. до н. э.) предполагает, что охотники-собиратели Тибетского нагорья обменивались просом с жителями равнин намного раньше предполагаемого появления ячменя.
А отсутствие следов отбора EGLN1 в верховьях реки Хуанхэ, у людей, принадлежащих к культуре Цицзя (Upper_YR_LN) в сочетании с предполагаемой селективной разверткой EGLN1, примерно 8000 назад, предполагает, что две популяции, возможно, уже разделились задолго до прибытия ячменя в район Ганьсу и Цинхай.
Ячмень выращивался как второстепенная культура в этих районах еще ок. 2000 г. до н.э., оставляя открытой возможность более ранней экспансии людей, выращивающих ячмень, на Тибетское нагорье ещё до 1650 г. до н.э., но археологических свидетельств, подтверждающих такой сценарий, не хватает.
Авторы отмечают, что их нынешние данные не могут полностью отвергнуть гипотезу, основанную на распространении ячменя, поэтому они призывают к поиску более древних геномов с Тибетского нагорья, а именно старше 1650 года до н. э., чтобы непосредственно проверить гипотезу.
Наконец, новое исследование показывает длительное воздействие естественного отбора на формирование генофонда высокогорных жителей Восточной Азии. Следует отметить, что увеличение частоты аллеля EPAS1 за период времени, от древних жителей из районов Мананг и Мустанг до современных тибетцев, подчеркивает медленное, но устойчивое действие положительного отбора на этот генетический вариант денисовского происхождения.
Будущие исследования позволят лучше понять популяционную историю этого региона, хотя и не только этого!
Источник:
Liu, CC., Witonsky, D., Gosling, A. et al. Ancient genomes from the Himalayas illuminate the genetic history of Tibetans and their Tibeto-Burman speaking neighbors. Nat Commun 13, 1203 (2022). doi.org/10.1038/s41467-022-28827-2
Вспомогательные источники: Gnecchi-Ruscone, G.A., Jeong, C., De Fanti, S. et al. The genomic landscape of Nepalese Tibeto-Burmans reveals new insights into the recent peopling of Southern Himalayas. Sci Rep 7, 15512 (2017). doi.org/10.1038/s41598-017-15862-z
Ren L, Yang Y, Wang Q, et al. The transformation of cropping patterns from Late Neolithic to Early Iron Age (5900–2100 BP) in the Gansu–Qinghai region of northwest China. The Holocene. 2021;31(2):183-193. doi.org/10.1177/0959683620941137
Jeong C, Peter BM, Basnyat B, Neupane M, Beall CM, Childs G, et al. (2017) A longitudinal cline characterizes the genetic structure of human populations in the Tibetan plateau. PLoS ONE 12(4): e0175885. doi.org/10.1371/journal.pone.0175885
Ancestral Origins and Genetic History of Tibetan Highlanders Dongsheng Lu, Haiyi Lou, Kai Yuan, Bing Su, Longli Kang, Shuhua Xu doi.org/10.1016/j.ajhg.2016.07.002
Genetic Evidence of Paleolithic Colonization and Neolithic Expansion of Modern Humans on the Tibetan Plateau, Molecular Biology and Evolution, Volume 30, Issue 8, August 2013, Pages 1761–1778, Xuebin Qi, Chaoying Cui, Yi Peng, Xiaoming Zhang, Zhaohui Yang, Hua Zhong, Hui Zhang, et. all doi.org/10.1093/molbev/mst093
Показать полностью
20

Если вы профи в своем деле — покажите!
Такую задачу поставил Little.Bit пикабушникам. И на его призыв откликнулись PILOTMISHA, MorGott и Lei Radna. Поэтому теперь вы знаете, как сделать игру, скрафтить косплей, написать историю и посадить самолет. А если еще не знаете, то смотрите и учитесь.
Раса и адаптация. Часть 3. Негры (продолжение)
Начало: Раса и адаптация. Часть 1
Продолжение 1: Раса и адаптация. Часть 2. Негры
Еще раз здравствуйте, дорогие читатели. Не предполагал, что первые посты по теме вызовут такое бурное обсуждение и интерес. Хотя поднимающиеся в комментариях темы очень интересны, я бы предпочел не уходить в антропологию слишком сильно - во первых я ее знаю не слишком хорошо, а во вторых - если удариться в антропологию, частота публикаций упадет, а с неграми мы и за 10 публикаций не закончим. Так же - генетику я планирую затрагивать так мало, как возможно и примесь денисовцев и неадндертальцев буду указывать только там, где это сильно повлияло на малую расу - и очень мельком.
В целом именно благодаря этим обсуждениям серия задумывавшаяся как 5-6 постов увеличилась едва ли не вдвое - но мне все же хочется удовлетворить любопытство всех читателей, так что будем идти как идет.
И ах да - африканские негры и впрямь не имеют неандертальской или денисовской крови. Общий предок - гейдельбергский человек покинул Африку еще до того, как в ней сформировались люди современного типа. В то же время черные австралийской расы имеют значительную примесь денисовцев. А теперь - давайте продолжать.
Это все чем отличаются негры?
Нет. Другие выраженные отличия касаются обмена веществ и адаптаций к перегреву. Пот и моча негров, в особенности прожавающих в самых жарких частях Африки, например - на границе с Сахарой и другими пустынями, а так же в жарких саванная субэкваториального пояса, содержат меньше соли, да и потоотделение у них в целом ниже - зато и моча и пот содержат больше продуктов обмена - и соответственно сильнее пахнет. Это тоже является адаптацией к перегреву - любой человек, независимо от расы, в условиях жары интенсивно потеет и теряет соль, которую нужно компенсировать - в противном случае наступает истощение, слабость и, в перспективе, смерть - ведь соль, а точнее ионы натрия, необходима на клеточном уровне - они обеспечивают нормальную работу клеточных мембран и транспорт питательных веществ в клетки. Но на материке (любом) мало источников соли, необходимой для человека, а вот потери с жарой не компенсируется ее приходом с пищей (в естественных условиях - то есть без возможности где-то купить соль) отсюда и адаптация. Мочеиспускание так же происходит реже за счет более узких почечных сосудов. Однако несмотря на эти адаптации перегрев может быть смертелен. При любом раскладе при перегревании потовых желез где-то до +39 градусов по Цельсию наступает тепловое утомление потовых желез, пот перестает выделяться, тело - охлаждаться и температура тела начинает расти. В перспективе - может достичь летальной. Защитой именно от этого является биоритмы негров - падение активности днем, потребность залезть куда-то в тенёк и спать.
Кроме того негроиды и близкие к ним по адаптациям австралоиды - единственные расы у которой не отмечается сильного угнетения пищеварения при перегреве организма. Замечали, что в жару часто не хочется есть? Это происходит потому, что при перегреве снижается выработка желудочного сока и желчи, нарастает вязкость слюны. Эта проблема имеются так же и у белых, и у монголоидов. А негры к этому более адаптированы, чем остальные расы. Что как бы указывает, что монголоиды живущие в джунглях появились там гораздо позже, чем негры. Да и решение они нашли такое, что адаптироваться им и не пришлось.
Экотипы мелкого ранга? Что, не все негры одинаковы? Это про малые расы? Да, про малые расы и не только. Разные школы антропологии выделяют разное число малых рас негров, наиболее типично разбиение черной расы (экотипа) на 3 подрасы (мезоэкоптипа в экологическом смысле): широко распространенную негрскую, центральноафриканскую (негрилльскую), южноафриканскую (бушменскую). Все эти расы так же являются продуктами естественного отбора и по своему уникальны.
Кроме того, имеется ряд рас, происхождение которых условно спорно или является смешанным. К примеру эфиопская (восточно-африканская) раса, населяющая Эфиопию, Сомали, Судан и ряд других восточно-африканских стран имеет смесь негроидных и европеоидных черт, при этом про ее происхождение является спорным: неизвестно, является ли она реликтом давних эпох, возникшим еще до разделения на белую и черную расы, и предком и тех и других - или является ли она продуктом смешивания европеоидных соседей (древних белых племен, потом - древних египтян, позднее - арабских завоевателей) и негров. Однако ее набор адаптаций в целом довольно схож с типичными негроидами, хотя и чуть менее ярко выражен. Однозначно смешанными являются все черные, проживающие к северу от Сахары (всяческие берберы). Этот регион регулярно завоевывался белыми, практически никогда не был изолирован от европейцев и смешение происходило неизбежно и постоянно.
Вместе с тем часть откровенно черных ребят (веддоидная, меланезийская (негритосская/папуасская - да, негритос и папуас, это не оскорбительные прозвища, а именно названия нескольких малых рас!) и австралийская малые расы) относятся к большой австралоидной расе -- четвертой из больших рас человека. Их очень долго объединяли с негроидной расой, но при внешнем сходстве и крайне сходных адаптациях, они покинули Африку очень давно - около 75-60.000 лет назад, в период, когда формирование современных рас еще не было завершено. Они - одна из наиболее рано обособившихся ветвей человеческой расы. О ней мы вкратце поговорим в отдельной небольшой публикации, т.к. экология представителей этой расы весьма сходна с таковой же у негроидов.
А теперь давайте поговорим о тех малых расах, в чисто африканском происхождении которых сомнений нет.
Негрская раса
Default-негр, в сущности. Все сказанное в первой части абсолютно применимо к представителям этой, сложившейся в саваннах северной части Африки, малой расы. В целом эту малую расу можно охарактеризовать двумя словами: типичный негр с темной, защищающей от солнца кожей, темной же радужкой глаз, с кучерявыми черными волосами, толстыми губами, широким и плоским носом, выступающими вперед челюстями. В пределах этой расы выделяется целая куча подтипов (суданский - наиболее чистый и типичный негр; центрально-африканский, с явной примесью негрилльной расы и более мелкий; восточно-бантоидный тип - типичный негр, но с более выступающим и узким носом; южно-африканский, с примесью бушменской расы; и восточно-африканский - с примесью эфиопской расы.
Негрилльская, пигмейская, или центрально-африканская раса
В принципе эта малая раса настолько своеобразно. что заслуживает отдельного поста. Именно ее описание раздуло пост настолько, что включение в него австралийской раса, которую я хотел объединить с черной раздуло бы пост, до утраты разумного размера.
Характерной чертой этой малой расы является очень низкий рост - до 145 см у мужчин и 140 у женщин. Эта раса - одна из немногих малых рас, которые смогли адаптироваться к жарким экваториальным дождевым лесам - и с антропологической и с экологической точки зрения она очень сильно обособлена от остальных негров. По данным генетических исследований предки современных пигмеев обособились от остальных негроидов около 60.000 лет назад. и населяли в основном бассейн Конго.
Сам облик представителей пигмейской расы значительно отличается от остальных негров - у них более светлая кожа, так как в тенистых дождевых лесах не нужно столько мелатонина для выживания. Однако сам темный цвет кожи не исчез, т.к. в целом черная раса сформировалась уже к моменту миграции их предков.. У них более, чем у остальных черных, развит третичный волосяной покров (борода, усы, волосы в нижней части живота), более тонкие губы и крайне широкий нос - хотя в принципе широкие носы так или иначе свойственны практические всем негроидам. По видимому широкий нос является исходным для человека, а вот носы других типов - уже являются адаптацией к холодному и/или сухому климату. Они не столь сильны и выносливы, сколь остальные негры, но притом имеют большую гибкость, нужную для жизни в лесу.
Условия дождевых лесов в целом одни из самых жестких и непригодных для человека на планете. По жесткости с ними может сравниться разве что Крайний Север, к примеру - северная Чукотка или Аляска. Охотники и, в первую очередь, собиратели легко могут найти здесь много добычи - личинки, какие-то коренья и фрукты доступны здесь почти каждый год. А вот сельское хозяйство, способное прокормить больше населения и более устойчиво, чем собирательство, осложнено сезоном дождей смывающим плодородный слой, и частыми паводками. Да и то сельское хозяйство что там есть обычно является подсечным - вырубаем кусок леса, сжигаем деревья, сеем без удобрений, пока не выродится почва - кочуем на новое место и начинаем все сначала. Скотоводство практически невозможно из-за сонной болезни и переносящей ее мухи-цеце. В результате всего этого недоедание и недополучение белков и по сей день является типичной проблемой региона. Смерть от голода вполне обычна во всей Африке, и центральная Африка - не исключение. Другой крупной угрозой жизни является множество тропических болезней. Сонная болезнь, тропические лихорадки (желтая лихорадка, болезнь Эбола и др.), жесточайшие гельминтозы (проще - глисты) поражающие не только кишечник и печень, но и кожу, глаза и другие органы, куча других экзотических болезней, не говоря уже о малярии. До кучи - присутствует множество ядовитых тварей.
В целом комплекс условий в этих краях таков, что в естественной среде, до прихода белого человека, средний срок жизни представителя этой расы составлял около 20 лет, и не сильно вырос даже в наши дни. Поэтому одна из первых целей адаптации негроидов к условиям экваториальных дождевых лесов являлась компенсация сверхвысокой смертности. А какой могла быть эта компенсация? Только наращивание рождаемости! И здесь мы подходим к одному из самых типичных для этой группы негров признаку: малорослости. Очень долго считалось что малорослость - генетическое отклонение обусловленное долгой изоляцией и инбридингом или последствия жаркого и душного климата ( с более мелкого тела проще отвести лишнее тепло) или из поколения в поколение скверным питанием. Однако последние исследования показали, что это не так. И здесь я даже сошлюсь на работу, поскольку открытие это недавнее - но очень хорошо все объясняющее (https://www.nationalgeographic.com/science/article/short-liv...): из-за короткого срока жизни пигмейки период размножения смещается в сторону молодого возраста. Из-за чудовищной детской смертности от болезней, голода и укусов всяких тварей для поддержания популяции требуется, чтобы к 20-22 годам у женщины было уже 5-6 детей. Как этого добиться? Правильно - снизить возраст полового созревания. А значит вся энергия растущего организма направляется именно на этот процесс. Физический рост тела практически останавливается в 11-12 лет, примерно в это же время начинается завершение полового созревания - и к 15 годам у женщин наступает пик фертильности - период в который вероятность зачатия и успешного вынашивания ребенка максимален. То, что для европейца воспринимается как педофиилия (или в крайнем случае эфебофилия), для центральноафриканской расы - условие выживания, и их организмы к этому полностью адаптированы. Однако при этом менопауза (потеря женщиной способности к размножению, обусловленная возрастом) наступает к возрасту около 40 лет - считай что на 10 лет раньше чем у белых. Впрочем, поскольку подавляющее большинство местных жителей на протяжении всей истории, кроме последнего века, не доживает до этого возраста, большого значения это никогда не имело.
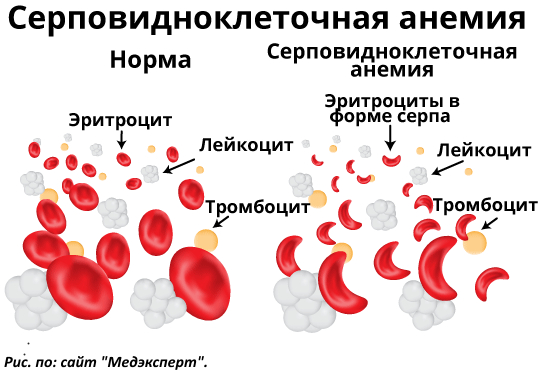
Одним из сильнейших факторов ограничивающих рост и поддержание популяции является малярия. В условиях отсутствия лекарств (а лекарства для негров в тех краях стали доступны лишь в XX веке) она смертельна с вероятностью 100%, а область расселения негрилльской расы и по сей день является ее основным природным очагом - на долю этого и близких к нему регионов приходится подавляющее большинство случаев этой болезни. (Чтобы узнать о малярии больше - прочитайте предыдущую заметку (Раса и Адаптация. Необходимое отступление: некоторая информация об основах генетики и малярии, как лимитирующем факторе). На практике это означает, что при заражаемости сотни миллионов в год, к 20, а скорее к 10 годам жертвой малярии станет (а значит - умрёт) каждый. В принципе условием заселения дождевых лесов Африки в общем и бассейна Конго было наличие адаптации - иммунитета к этой страшной болезни. Без него люди живущие в том районе вымерли бы еще тысячелетия назад - смертность в любом раскладе обгоняла бы рождаемость. Как ни странно такой адаптацией стала потенциально смертельная наследственная болезнь - серповидноклеточная анемия. Эритроциты человека больного этой болезнью неполноценны. Нормальные эритроциты имеют дисковидную двояковогнутую форму, эритроциты же при этой болезни имеют форму серпа (рисунок в конце поста). Эритроцит такой формы недостаточен для того, чтобы маляриный плазмодий внедрившийся в него поглотил достаточно питательных веществ для деления - а внедрится в новый эритроцит у него уже не получится.. В крови больного этой болезнью плазмодии, поражая эритроциты, теряют способность размножаться - и погибают. То есть - человек выздоравливает от малярии после первого приступа. Платой за это является смертность от серповидноклеточной анемии.
Чтобы понять почему негры, защищенные этой болезнью от более опасной (для популяции в целом) малярии еще не вымерли уже от нее придется ненамного заглянуть в генетику (подробнее о генетике - тут: Раса и Адаптация. Необходимое отступление: некоторая информация об основах генетики и малярии, как лимитирующем факторе). Обозначим как А аллель, отвечающий за нормальную форму эритроцита, а как a - аллель, отвечающий за серповидную форму эритроцита. Практически все жители бассейна Конго, где эта болезнь распространена тотально, достигшие 20 лет - гетерозиготны (Aa) по этому признаку. Доминирование аллеля нормальной формы эритроцита - неполное. В итоге потомство всех этих гетерозигот распределяется как:
25% гомозиготны по доминантному признаку (AA). Это здоровые люди, у которых нет этой болезни. Вот только в условиях экваториальной Африки эти здоровые люди мрут от малярии как мухи - чаще еще в детском возрасте, редко доживая до 10-15 лет.
50% - гетерозиготы: обычные гетерозиготы (Аа), как родители. Имеют более неполноценные эритроциты в форме широкого серпа. Они иммунны к малярии, но всю жизнь страдают анемией, из-за которой редко переваливают за 40 лет. Более слабые и вялые, они тем не менее выживают в этом малярийном аду и дают потомство - в то время как гомозиготные люди (АА) - сделать это в 99% случаев не успевают.
25% - гомозиготны по рецессивному признаку (aa). Эта часть потомства обречена на смерть. Их эритроциты имеют форму узкого серпа и их не хватает для того, чтобы переносить достаточно для выживания организма кислорода. Они умирают еще в младенчестве.
Следует так же отметить, что больные этой болезнью очень чувствительны к кровопотере. Их организмы уже функционируют на пределе - серповидные эритроциты переносят лишь немногим больше того минимума кислорода, который базово нужен для выживания и активной жизни. Европеец (да и нормальный негр, если честно) гарантировано погибают при потере 70% циркулирующей крови, для центральноафриканских негров смертельны куда меньшие уровни кровопотери - 35-50% от объема циркулирующей крови. Что тоже увеличивает вероятность кончины до достижения хотя бы 30 лет.
Нужно отметить, что с потерей негрилльской расой изоляции (это случилось несколько тысяч лет назад) эта болезнь помаленьку распространилась по всей Африке (больше - у негров центрально-африканского подтипа негрской расы, меньше - у всех остальных), но ее распространение все же не слишком велико - да и в среде людей с нормальными генами (AA) гетерозиготы будут довольно редки, а гомозиготы (aa) будут крайне редки - так что младенческой смертности просто не будет. Просто время от времени рождаются чахленькие и менее сильные и живучие негритята-гетерозиготы (Aa), а детская смертность у гетерозигот реально невелика.
Однако даже в США этой болезнью страдает около 100.000 негров. При получении адекватного лечения и регулярных переливаний крови 90% больных (включая гомозигот (аа)) доживают до возраста 20 лет и 50% (тут правда все - гетерозиготы) до 50 лет.
Конвергентная эволюция
Перед тем как обсуждать следующие расы нам необходимо ввести понятие "конвергентная эволюция" - процесс когда у разных таксонов (семейств, родов, видов, подвидов) под действием сходных условий среды и общего образа жизни образовываются одни и те же адаптации. Хорошими примерами могут служить такие пары как "пума (кошачьи) - фосса (виверровые)" или "еж (насекомоядные) - дикобраз (грызуны)". Эти животные - не родственники, но они вырабатывали сходные признаки: сходную форму тела, сходные стратегии выживания.. Первые - для эффективной охоты ( фоссы процветают на Мадагаскаре) занимая там экологическую нишу отсутствующей там пумы. Вторые - приобрели свои черты для эффективной защиты от более крупных хищников. В целом примеров конвергентной эволюции очень много. Но действует она и на уровне людей, отдельные малые расы которых вырабатывали сходные приспособления к среде независимо друг от друга.
Бушменская или южно-африканская раса
Бушменская раса крайне своеобразна с генетической и антропологической точек зрения - настолько, что некоторые антропологи предпочитают рассматривать ее как отдельную большую расу, тем паче что остальные негроиды куда ближе к европейцам, нежели к бушменам. Находки свидетельствуют о том, что эта раса сложилась где-то в южной Африке и одно время была широко распространена, продвинувшись по горным саваннам восточной Африки в Северное полушарие - но впоследствии была вытеснена оттуда негрской расой. Представители этой расы имеют не черный или темно-коричневый, а желтовато-бурый или бурый цвет кожи. Это указывает, на то, что формирование этой расы шло в зоне с меньшим воздействием солнечного света - скорее всего в субтропических степях и полупустынях южной оконечности Африки. Сухость климата, ветры, влияние холодных воздушных масс привносимых зимой с умеренного пояса, привели к тому, что включились механизмы конвергентной эволюции - и в результате у этой малой расы успешно сформировался ряд монголоидных признаков - как уплощение лица, так и эпикантус - складка верхнего века, в примыкающем к носу уголке глаза, прикрывающая выходы слезных желез и, в условиях засушливой южной Африки, защищающая их от забивания пылью и иссушивания ветром. Однако отсутствие сильных холодов сохранило у них типичный крупный негритянский нос.
Кроме того у представителей этой расы, особенно у женщин, присутствует запас жира, откладывающегося в первую очередь на ягодицах (т.н. стеатопигия) - а кроме того сильно искривлен позвоночник (выраженный лордоз). Фото внизу.
Для этой малой расы - стеатопигия и лордоз - это нормально и присуще практически всем ее представительницам, и в чуть меньшей степени - представителям.. Чтобы не ставить тег NSFW грудь женщины на скромной в остальном фотографии отцензурена - хотя я и сомневаюсь, что у кого-то возникло бы желание рукоблудить на откровенно научное фото, тем паче столь странное. Зачем эта черта выработалась - не знает никто. В экологии человека - свои загадки.
В целом же большинство адаптивных признаков негроидной расы в той или иной степени присуще и бушменам - и если защита от солнца присутствует в сниженной форме, то защита от потери соли, высокая выносливость, повышенная сила, позволяющая быстро бегать и двигаться присутствуют в почти полном объеме - хотя и не столь выражены, как у негрской расы. Ввиду изоляции это малой расы от остальных негроидов на протяжении десятков тысяч лет все эти признаки сформировались опять же в ходе конвергентной эволюции.
В заключении обсуждения этой расы хочется развеять один маленький миф - постоянную эрекцию у мужчин-бушменов. Нету ее. Член обычно частично эрегирован, больше насыщен кровью, чем у всех остальных - но упершись в небеса не стоит. Иначе было бы слишком много травм и местные негры тупо бы вымерли.

Показать полностью
2


На поляне траву полёвки в полночь косили, и при этом напевали странные слова
Биологи заметили необычное поведение грызунов. Полёвки косят траву, чтобы спастись от пернатых маньяков.

В степях Монголии, Китая и Забайкалья живут небольшие грызуны – полёвки Брандта (Lasiopodomys brandtii). Чаще всего они селятся именно в степях, иногда обитают по периферии солончаковых низин и в долинах мелких степных речек, а также заселяют предгорные и горные массивы и иногда проникают даже в пустыни.
Излюбленные местообитания полёвок Брандта – разнообразные луга в котловинах степных озер, обычно служащие пастбищем для бесчисленных стад. Живя в сухих открытых степях, полёвка постоянно находится в опасности, которая исходит от многочисленных хищников.
Учёный из Эксетерского университета Дирк Сандерс с коллегами заметил, что полёвки Брандта срезают блестящий ковыль (Achnatherum splendens), чтобы не проморгать одного из своих врагов — сорокопута.
Кто такие сорокопуты?
Этот пернатый убийца орудует по всему миру, в основном в Евразии и Африке. Он распинает своих жертв на острых предметах – ветках и сучках, колючей проволоке, шипах – а затем разрывает их на клочки.
Нет, это не портрет маньяка, а описание сорокопута – маленькой и на первый взгляд безобидной птахи, от которой мороз по коже. Натуру этих пернатых отражает их название на латыни – Lanius («палач», «мясник», «приносящий жертву»).
Хотя сорокопуты и относятся к воробьинообразным, клюв у них острый и крючковатый – такой скорее можно увидеть у ястребов или сов.
Поймав слишком крупную добычу (ящерку, например, или мышку), сорокопуты аккуратно нанизывают её на ветку, устраивают на шипах растений или в других подходящих местах. А потом вся эта кровавая инсталляция разрывается острым клювом на части – настоящий пир для небольшой птички.
Сорокопут со своими жертвами



Как выяснилось, полёвки Брандта из рода Брандтовы полёвки блестящий ковыль не едят и не используют для обустройства своих жилищ – они именно срезают длинные стебли для собственной безопасности: отсутствие растения позволяет им вовремя заметить подлетающего хищника и успеть юркнуть в убежище.
Это любопытный пример инженерии экосистемы, отмечают исследователи.
Когда над головами грызунов летали сорокопуты, они резко сокращали объём ковыля в районе своих местообитаний. Это привело к тому, что сорокопуты реже залетали в эти районы, видимо, считая участки со скошенной травой плохими охотничьими угодьями.
«Подобная деятельность обходится полевкам дорого с точки зрения затраты энергии, поэтому для такого поведения должно быть высокое "давление отбора": укорачивание травы должно значительно повысить их шансы на выживание», – пишут учёные.
Исследователи также проверили, как влияет отпугивание птиц, установив сети над определенными участками. И тут их ждало второе открытие: поскольку сорокопуты совсем перестали летать над головами грызунов, полёвки перестали косить траву.
«Иногда мы недооцениваем способность диких животных реагировать на изменения в окружающей их среде, – говорит доктор Сандерс.
– В этом случае полёвки смогли изменить своё поведение в ответ на исчезновение хищников. Наши результаты напоминают о том, что некоторые виды демонстрируют замечательную адаптацию».
«Это исследование является хорошим примером того, что животные могут активно изменять свою среду обитания, чтобы снизить риск нападения хищников», — отмечают учёные.
Исследование опубликовано в журнале Current Biology.
Показать полностью
4

Предок ягуара и других больших кошек был похож на леопарда
Ягуар — самая большая дикая кошка, распространенная на территории Северной и Южной Америк. Поскольку животному грозит вымирание, исследователи решили изучить ягуаров, чтобы понять, как их можно спасти.
Биологи из Федерального университета Риу-Гранди-ду-Сул (Бразилия) расшифровали геном ягуара (Panthera Onca) и сравнили его с геномами четырех других больших кошек: тигра, льва, снежного барса и леопарда. Все они принадлежат роду Panthera.
Ученые пришли к выводу, что 4,6 млн лет назад все большие кошки разошлись от общего предка, похожего на современного леопарда.
Исследование опубликовано в Science Advances.
Расшифровав ДНК, исследователи выяснили, что 4,6 млн лет назад пять больших кошек разошлись от общего предка, который распространился из Азии на большую части планеты. Напоминал он современного леопарда. В период плейстоцена (2,5 млн — 11,7 тыс. лет назад) численность популяций всех видов уменьшилась, а значит, их генетическое разнообразие стало скудным.
Ягуар (Panthera onca)

Ученые обнаружили, что скрещивание помогало животным выжить и развить новые способности. Например, ягуар после скрещивания с львом стал более крупным, что сказалась на силе его укуса. Благодаря этому животное смогло охотиться на крокодилов и черепах. Также ягуар унаследовал у льва два гена, участвующих в формировании зрительного нерва.
Другой пример эволюционной адаптации продемонстрировал снежный барс. Животное научилось справляться с нехваткой кислорода и приспособилось жить в Гималаях, Тибете и других горных хребтах.
Ученые уверены, что результаты исследования помогут остановить сокращение популяций.
Показать полностью
1


Биохимия,которую мы заслужили

С просторов.
баянометр всякую хрень выдал.
Показать полностью
2