
Человек и его ближайшие родственники

Не тигры или лошади, не волки или медведи, не слоны или дельфины. Крысы, тупайи, шерстокрылы, зайцы и обезьяны.
Не тигры или лошади, не волки или медведи, не слоны или дельфины. Крысы, тупайи, шерстокрылы, зайцы и обезьяны.

В мире птиц часто происходят удивительные и необъяснимые явления, заставляющие нас вновь и вновь удивляться сложности и изяществу природы. Одним из таких примеров является поведение американской, или воробьиной пустельги (Falcosparverius), особенно интересное на зимовках в Калифорнии, США.
Эта маленькая, но изящная хищная птица, известная своей ловкостью и скоростью, демонстрирует поразительное явление: в зимний период самцы и самки ведут раздельный образ жизни. При этом интересно, что такое разделение не является случайным или временным, а обусловлено строгой сегрегацией территорий.
Самки американской пустельги выбирают для зимовки открытые угодья, где в изобилии встречаются мелкие грызуны. Это идеальная местность для охоты, где они могут полноценно питаться и поддерживать свои силы. Наблюдения показывают, что самки не только предпочитают эти территории, но и активно оттесняют самцов от них, создавая своеобразные «женские» зоны.
Самцы же, в свою очередь, вынуждены уклоняться в более труднодоступные для охоты лесные угодья. Здесь корм для птиц добывать значительно труднее, что, возможно, влияет на их физическое состояние и поведение. Это явление ставит перед учеными ряд вопросов относительно причин такого разделения. Возможно, это связано с конкуренцией за ресурсы или же с различиями в поведенческих стратегиях между самцами и самками.
Такое поведение американской пустельги вновь напоминает нам о сложности и многообразии природных явлений. Исследования продолжаются, и каждый новый сезон наблюдений приносит ученым новые данные, помогающие лучше понять этот уникальный аспект жизни этих удивительных птиц.

Причиной, по которой у большинства самцов млекопитающих (исключения: кони, крысы и мыши) [1] есть вроде бы бесполезные для них соски, как у самок, является то, что неразвитые молочные железы не относятся к репродуктивной системе животного и потому образуются независимо от пола особи наряду с другими "унисекс" органами: сердцем, кишечником и т.д.
И только позднее, в процессе полового созревания у самок молочные железы начинают развиваться и входят в состав их репродуктивной системы, у самцов же они так и остаются неразвитыми и никуда не входят (исключения: самцы даякских фруктовых летучих мышей и летучих лисиц в маске Бисмарка (да, такое название странное) кормят молоком своё потомство наряду с самками) [1].
Не совсем равноценной аналогией в качестве иллюстрации может послужить хвост павлина (хотя на самом деле ярким оперением покрыт не сам хвост, а надхвостье птицы. Хвостик же находится под массивным покровом «веера» и только поддерживает его и обеспечивает функциональность).
До полутора лет самки и самцы павлина практически ничем друг от друга не отличаются, их хвосты не играют роли в репродуктивной функции и образованы примерно одинаково.
А вот уже по достижению возраста в полутора лет в процессе полового созревания хвосты самцов начинают развиваться, обрастают верхними кроющими перьями и, условно, входят в их репродуктивную систему к трём годам. [2]
На вопрос, почему же молочные железы изначально не мутировали как исключительно материнская прерогатива, можно ответить, обратившись к ранним предкам млекопитающих.
Согласно гипотезе, высказанной в ряде работ Олавом Офтедалем и его коллегами, предковые формы молочных желез возникли у древних синапсид (предков млекопитающих) около 310 миллионов лет назад и первоначально использовались с целью сохранения необходимой влажности в кладке яиц [3].
Вполне вероятно, что в то время, по аналогии с современными птицами, и самцы, и самки синапсид делали примерно равный вклад в заботу о кладке, в том числе и в вопросе сохранения влажности, поэтому работоспособные железы развились у обоих полов предков млекопитающих.
Ведь после откладки яиц в дальнейшем развитии детенышей нет ничего такого, что требовало бы какой-то специфической помощи со стороны матери: самец способен ухаживать за яйцами не хуже самки, а после того, как птенец вылупится, самка точно так же, не хуже самца, может добывать корм и приносить его в гнездо [4].
Когда же молочные железы развились до своей современной функции - кормления потомства - и стали использоваться преимущественно только самками, сделать эти железы исключительно материнскими стало затруднительно и слишком затратно, да и не особо нужно.
Затруднительно и затратно, так как генетически молочные железы уже были двуполым органом (как какая-нибудь поджелудочная), да и плюс, чем старше какая-либо особенность организма, тем меньше она подвержена генетической изменчивости.
А не нужно потому, что и самец с сосками, и самец без них имеют примерно одинаковые шансы на выживание, практически никакого эволюционного преимущества исчезновение молочных желез самцу не дает.
Однако, у самцов некоторых млекопитающих, таких как кони, крысы и мыши [1], все же произошло исчезновение молочных желез: они хоть и возникают на определённом этапе эмбрионального развития, но позже рассасываются и при рождении уже отсутствуют.
Источники:
[1] Зачем самцам молочные железы? Оказывается, есть виды, у которых мужчины могут вскармливать потомство! // Книга животных (19.10.2022);
[2] Хвост павлина // Наши птички (2023);
[3] Даниил Симонов. Маммализация териодонтов, или Как появилась лактация // XX2 ВЕК (18.05.2023);
[4] Джаред Даймонд. Глава 3. Почему мужчины не кормят грудью? «Нулевая» эволюция мужской лактации / «Почему нам так нравится секс» // Библиотека (2014).
Дельфины относятся к самым умным и известным животным на Земле. Существует около 40 различных видов дельфинов, и они обитают в океанах по всему миру. Сложная система эхолокации дельфинов позволяет им эффективно ориентироваться и добывать пищу в зачастую темной и мутной среде. Это удивительное проявление биологической инженерии исследуется с помощью высокоскоростной съемки и детальной компьютерной анимации. Мы многое узнали, однако вопрос присхождения эхолокации до сих пор остаётся без ответа: механизм эхолокации необычайно сложен, для его осуществления нужно огромное количество структур, каждая из которых сама по себе абсолютно бесполезна.
Некоторые выдержки из книги жалоб и предложений)

Блювота:) Нативная реклама

Самые большие вопросы были к этому пикабутянину.

Практически всем русскоговорящим людям, а думаю и не только им, знакомы такое народное творчество как пословицы и поговорки, и в некоторых из них, речь часто заходит о таком анатомическом образовании на лице или морде, как нос. Например – «не суй нос не в своё дело», «нос – через реку перерос», «любопытной Варваре, на базаре нос оторвали», так же, иногда употребляют более грубую форму – замарать или воротить рыло. Во всех приведённых мною выражениях, нос или его аналог рыло, представлены как нечто выпирающее относительно лица, что собственно так и есть, и даже если мы прекратим разглядывать человеческие носы, а перейдём к братьям нашим меньшим, например собакам или кошкам, да даже к свиньям, бурундукам, кенгуру и тем более слонам, во всех случаях нос, так или иначе, будет находиться чуть впереди, обладателя этого носа! Но если пойти чуть дальше и посмотреть на птиц, змей, крокодилов, лягушек и даже утконоса, то случится чудо, и выпирать всегда будет верхняя челюсть, а нос будет расположен немного позади.

Привет синапсиды, меня зовут Даниил, и я веду канал «Помни о предках. Палеонтология», поэтому для тех, кто не хочет много читать, внизу будет прикреплён ролик на эту же тему, а для любителей длиннопостов, я разберу небольшую, но очень интересную статью, опубликованную 2 ноября 2022 года в журнале «Работы национальной академии наук» или просто PNAS. В этом труде достаточно подробно анализируется выше озвученный парадокс, ну а точнее, как же терии или живородящие млекопитающие, дошли до жизни такой, да ещё и с выпирающим шнобелем! Для желающих почитать оригинал, будьте готовы к ГМО-эмбрионам, которых потом будут травить циклопамином мерзкие учёнусы! Ну а для остальных, пожалуй, начнём.
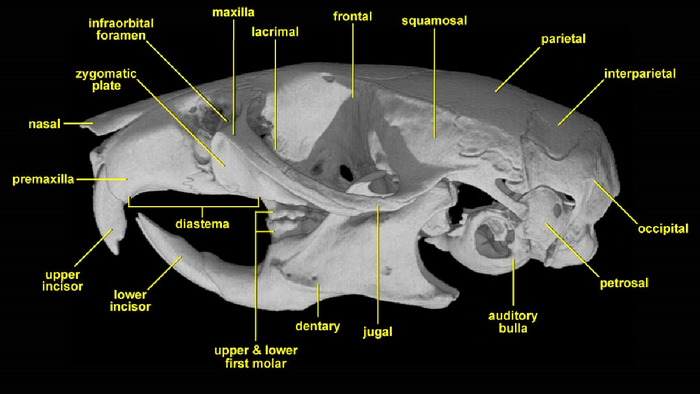
Итак, весь смысл в том, что у териев, то есть плацентарных и сумчатых млекопитающих, строение скелета черепа таково, что премаксилла или верхнечелюстная кость, смещена несколько вниз, а над ней располагается назальная или носовая кость, которая, например, у всех завропсид находится за премаксиллой, а у монотремат, они же однопроходные, мы можем наблюдать некий переходный вариант, но что это я, разве это доказательство эволюции, гораздо проще объяснить создание всего сущего сверхсуществом за 6 дней! Но, не будем о грустном. Статью можно поделить на две части, в первой проводится тот самый бесчеловечный опыт с эмбрионами, правда не человеческими, а лягушки, геккона, ехидны и мыши, а во второй части, сравниваются эволюционные изменения носовой части синапсид, как вымерших, так и ныне живущих. Начнём естественно с первой!
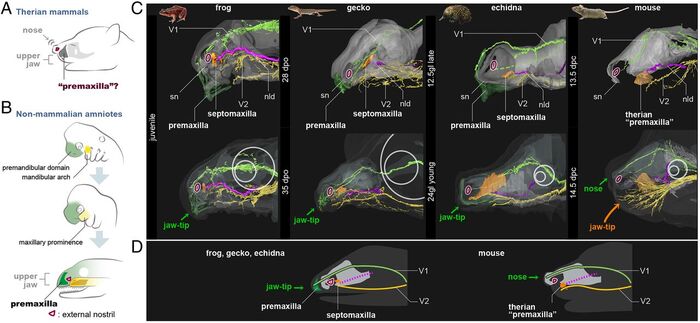
Собственно, для людей интересующихся эволюционной изменчивостью и происхождением видов, не будет секретом тот факт, что морфология эмбрионов тетрапод, весьма сходна до определённого периода, когда начнут проявляться специфические особенности анатомии конкретного вида, что является одним из доказательств эволюции. Именно поэтому учёные в составе Хироки Хигасямы, Тацуя Хиросавы, Ингмара Вернебурга и других, решили начать с начала, и сравнить анатомию эмбрионов. В результате были получены следующие данные: у зародышей лягушки, гекона и ехидны, премаксилла окостеневает медиально, то есть с середины, у первых двух рост наблюдается на протяжении всего времени, у монотремат же, рост замедляется, кость становится сравнительно меньше, а оставшееся пространство замещается верхнечелюстной перегородкой, а у зародыша мыши, окостенение происходит снаружи, относительно носовых капсул. Так же, у первой тройки верхнечелюстная кость возникает в области глазного нерва, а у териев, она развивается в области верхнечелюстного нерва. Таким образом, учёные заключают, что предчелюстная кость териевых млекопитающих, может быть сравнима с нетериальной перегородкой верхней челюсти, которая у териев отсутствует.
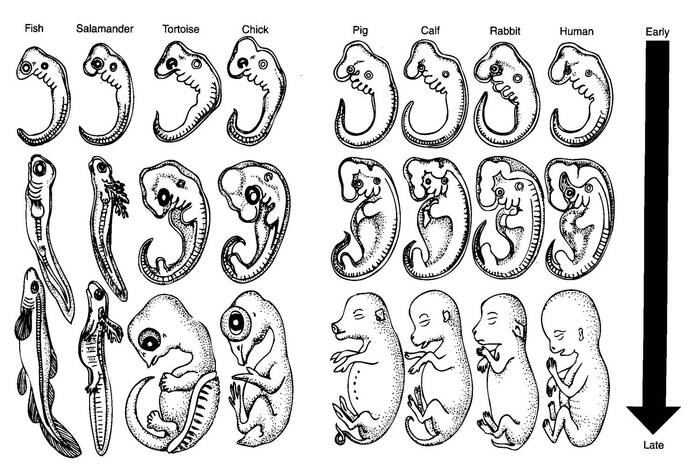
Чтобы понять какой из лицевых зачатков даёт начало верхнечелюстной кости и носовой перегородке у разных амниот, исследователи сравнили куриный и мышиный эмбрионы, искусственно замедлив развитие морды и создав условия для появления «заячьей губы» или расщеплённых губы и нёба, с помощью циклопамина, что позволяет ингибировать мезенхимальную пролиферацию нервного гребня, то есть запустить процесс увеличения числа мазинхимальных клеток, из которых потом вырастут соединительные и мышечные ткани, а так же некоторые клетки крови. На поздних стадиях развития глотки, слияние лобно-носовых и верхнечелюстных выступов, успешно подавлялось у обеих эмбрионов, так же контрольным экземпляром служил эмбрион геккона со спонтанно расщепившимся нёбом.
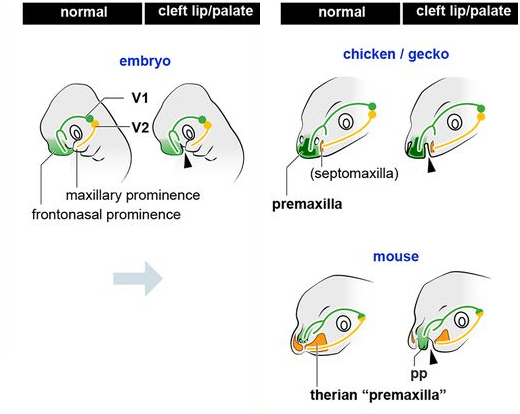
Расщелины у этих трёх животных сохранялись в одном и том же положении на протяжении всего развития, деля морду на три части: две боковые части верхнечелюстного выступа и среднюю лобно-носовую часть, где у эмбрионов геккона и курицы разделение совпадало, имея шов между верхнечелюстной костью и верхней челюстью, а перегородка верхней челюсти была обнаружена в области верхнечелюстного отростка, когда как у мышиного эмбриона верхняя челюсть также была разделена на медиальную и латеральную области, но в отличие от курицы и геккона, основное тело предчелюстной кости было обнаружено на боковой стороне расщелины. Таким образом, премаксилла у териев не является полностью лобно-носовой производной, а основная её часть, вероятно, происходит от верхнечелюстной перегородки.
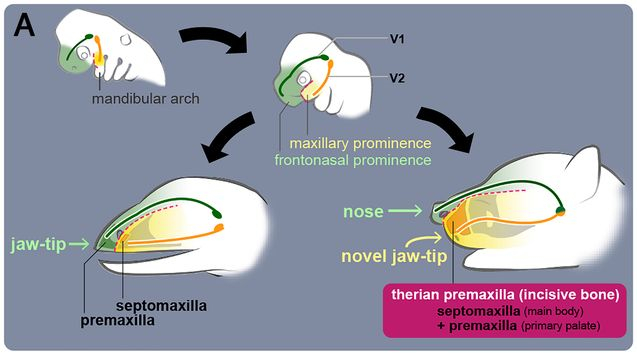
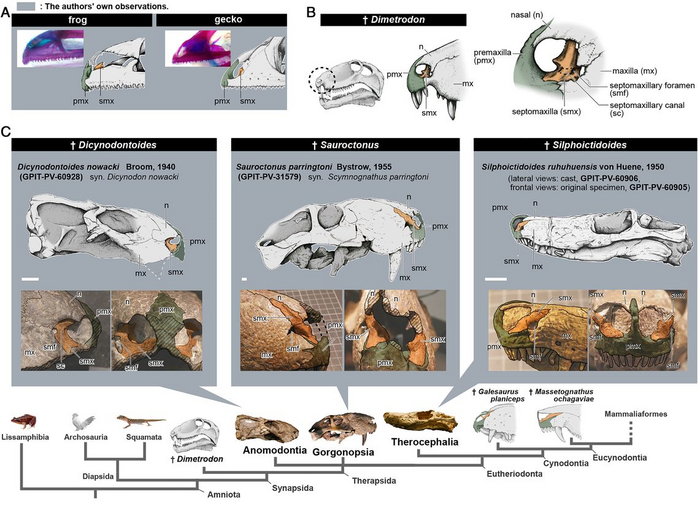
Во второй части работы, авторы сравнили окаменелости таких животных как: диметродон, дицинодонт, завроктон и сильфиктидоид, с представителями современных териев, как броненосец и мышь, естественно в области носоглотки. Основное внимание было обращено на септомаксиллу или верхнечелюстную перегородку, о которой говорилось в первой части работы. Такие перегородки наблюдаются как у молодых гекконов и лягушек, так и у базальных синапсид, представляя собой небольшой костный элемент в носовом отверстии пронизанный носо-слёзными протоками. У развитых синапсид, более близких к маммалиям как горгонопс завроктон и тероцефал сильфиктидоид , внешняя часть верхнечелюстной перегородки начала переходить на боковую поверхность морды, а предчелюстная кость и перегородка оставались разделёнными у большинства цинодонтов и не териевых млекопитающих. И только у териев, наблюдается утрата внутренней части предчелюстной кости с появлением единственного носового отверстия в черепе. Так как терианская премаксилла присутствует как у метатериев, так и у эутериев, то есть сумчатых и плацентарных, слияние перегородки и предчелюстной кости, должно было произойти у их общего предка. Возможно, это уже лично мои мысли, не последнюю роль здесь мог сыграть вирус, сделавших яйцекладущих млекопитающих – живородящими.
Таким образом, авторы продвигаются в давней загадке сравнительной анатомии, где ростральная или передняя часть верхней челюсти, обеспечивается разными нервными ветвями у териев и всех остальных тетрапод. Данное исследование показывает, что предчелюстная кость не представлена в передней части верхней челюсти не териевых четвероногих, а у териев образует подвижный нос, выдающийся наиболее вперёд относительно всего остального тела.
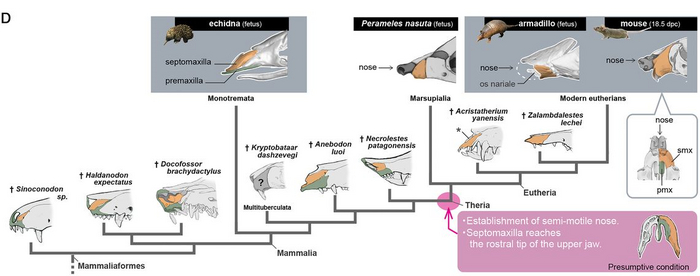
Так же, в ходе эволюции, создаётся териевая конфигурация верхней челюсти, с ростральным расположением вибрисов, что способствует наиболее быстрому получению информации посредством осязания. Думаю что через некоторое время, данный маркер можно будет использовать как ещё один признак для отличия териев от всех остальных, наряду с размером мозга и строением среднего уха.
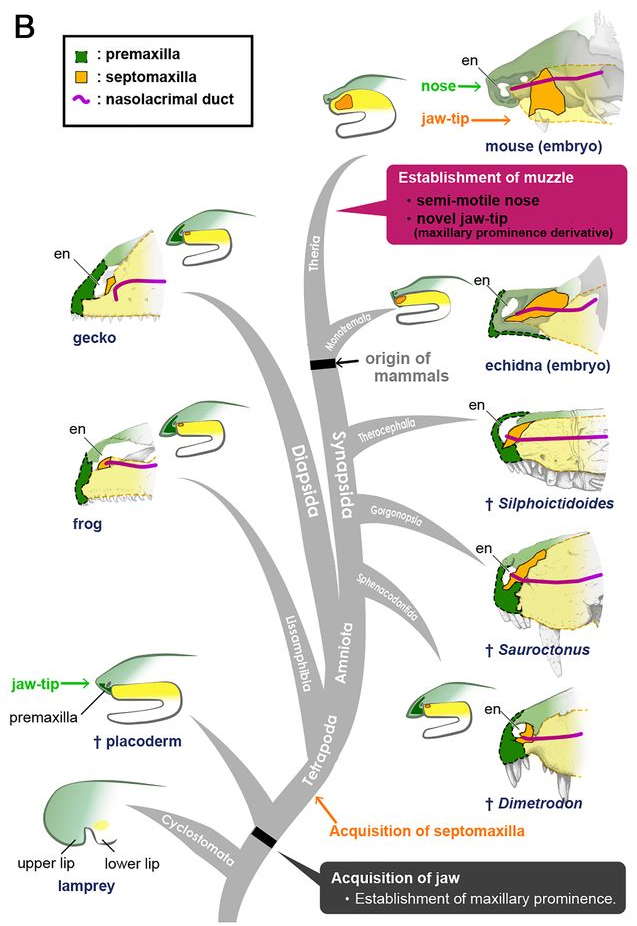
Вот так териевые млекопитающие оказались с носом, что безусловно дало им ряд преимуществ, перед всеми остальными тетраподами и вероятно сыграло свою роль около 65 миллионов лет назад. Ниже приведена ссылка на статью и оригинальный ролик на Ютубе, буду благодарен за его просмотр и оценку! Да скорого, мои синапсидные бро!
Оригинал статьи - https://www.pnas.org/doi/abs/10.1073/pnas.2111876118
Такой процесс появления потомства на свет как вивипарность или живорождение, возник независимо у более 150 видов позвоночных: сто пятнадцать раз у завропсид, тринадцать раз у костистых рыб, девять раз у хрящевых рыб, восемь раз у земноводных и один раз у плакодерм. Но самыми известными представителями живородящих являются, конечно же, маммальные синапсиды или млекопитающие, к коим относимся и мы с вами.

Вивипарность у синапсид появляется примерно 160-200 миллионов лет назад, по оценкам различных исследователей, но наиболее новые работы, всё больше отдаляют это событие к границе Триасового и Юрского периодов. Эта датировка примечательна тем, что примерно в это же время произошло одно из самых массовых вымираний Фанерозоя – Триасово-Юрское, стёршее со страниц палеонтологической летописи последних конодонтов, представлявших на тот момент около 20% морской фауны, многие виды амфибий, одну из доминирующих хищных групп архозавров - орнитозухий, тем самым высвободив пищевую нишу для динозавров, которые будут править миром ещё 135 миллионов лет, и остатки терапсид, за исключением недавно появившихся млекопитающих и, по сути реликтовых, цинодонтов и дицинодонтов.

Стоит сказать, что разделение поздних синапсид на млекопитающих и цинодонтов, достаточно условно, и определяется в основном по строению костей внутреннего уха и размеру мозга, с развитием его зон, отвечающим за обоняние, осязание и координации движений, так как с отпечатками мягких тканей синапсид у нас совсем не густо, как и с находками их яиц, определить способы выкармливания потомства не представляется возможным, но если исходить из гипотезы о том, что яйца синапсид были покрыты мягкой оболочкой как у современных однопроходных, то забота о потомстве в целом, и выкармливание с помощью видоизменившихся в молочные, потовых желёз, вероятно возникших у синапсид с появлением волосяного покрова, представляется наиболее вероятным вариантом. Именно поэтому живородящие млекопитающие, появляются на филогенетическом древе практически сразу после выделения маммалиаморф в отдельную группу, около 220 миллионов лет назад, в конце Триассового периода.
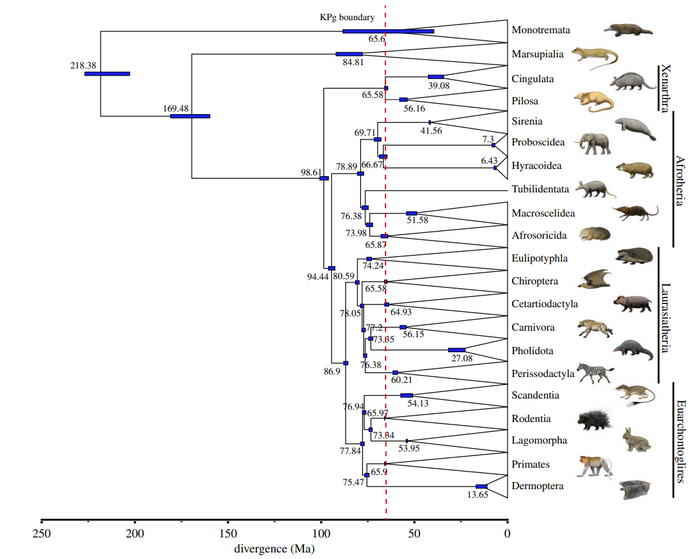
Млекопитающих принято разделять на четыре группы: аллотерии – не до конца изученная, но представляющая для нас интерес, по вопросу живорождения группа млекопитающих, монотремы или однопроходные и собственно терии – сумчатые и плацентарные. В данной статье, мы рассмотрим все четыре группы, проследив эволюцию появления потомства от однопроходных к плацентарным и сравним живорождение у плацентарных, сумчатых и как не странно - аллотериев.
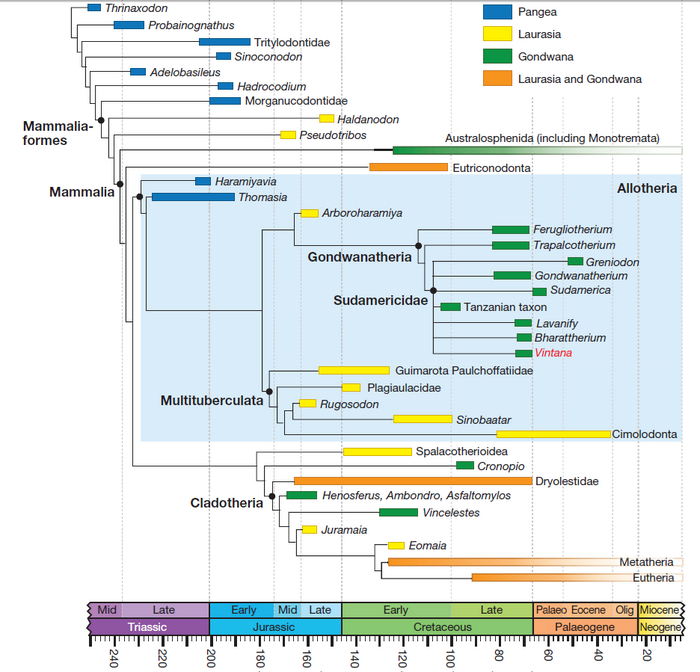
Выше, я упоминал об уникальности живорождения у млекопитающих, которое возникло всего единожды или дважды, завропсиды в этом плане вне конкуренции! Так вот, мало того что оно может примерно совпадать с Триасово-Юрским вымиранием, так и виной вивипарности у млекопитающих стали очень дальние родственники эпидемии охватившей человечество в 80-х годах ХХ века, вызванной микроскопическим, даже не совсем организмом, который нам известен как - вирус иммунодефицита человека, являющийся экзогенным ретровирусом, то есть вирусом распространяющимся горизонтально.

По одной из гипотез, где-то в конце Триасового периода, оставшихся на тот момент в живых синапсид охватил ряд пандемий неопределённых ретровирусных инфекции, но не только экзогенного, а эндогенного характера. Эндогенные ретровирусы, применяют вертикальный способ распространения, а их генетическая информация, стала составной частью всего генома организма животного. Эндогенные ретровирусы, обычно ведут себя как группа генов, находящихся под контролем клетки, а активатором к производству новых вирионов, может стать любой внешний фактор, влияющий на биом, как крупное извержение вулкана, падение метеорита, резкое изменение уровня моря и другие. Судя по всему, именно такое событие, вероятно приведшее к массовому вымиранию около 200 миллионов лет назад и привело к активации некого эндогенного ретровируса, практически у всех млекопитающих, за исключением маммального населения нынешней Австралии, а поражённые инфекцией животные подверглись мутациям, из-за привнесённых в геном животного, генетической информации вируса, вследствие чего, произошел метаморфоз клоаки, и вместо одного общего прохода, образовалось три прохода у особей женского пола и два у мужских особей.
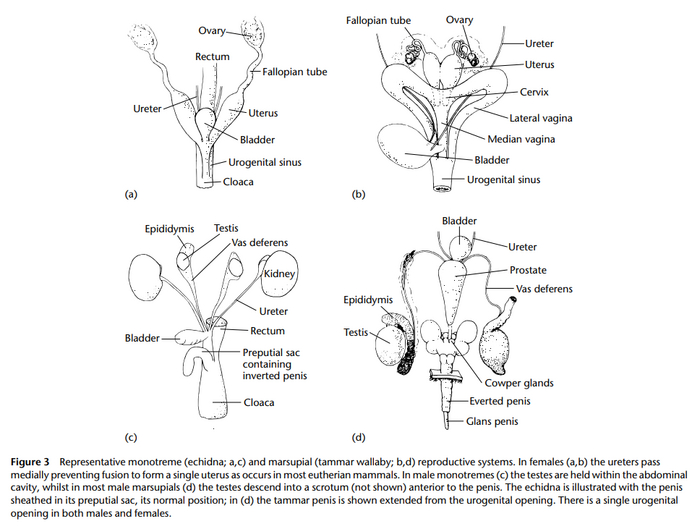
Говорить с точностью о данном метаморфозе не представляется возможным, ввиду неполноты палеонтологических находок, но предположительно, инфекция могла привести в выпадению клоаки наружу и формированием внешних мочеполовых органов в достаточно короткие сроки, в несколько сотен или тысяч поколений, а ввиду того, что речь идёт о некрупных раннемезозойских млекопитающих с быстрым половым созреванием у потомства, данный переход мог сложиться за считанные сотни лет. Австралийской же фауне, удалось каким-то образом избежать заражения инфекцией, толи ввиду наличия иммунитета, толи отделения Австралийского континента от Гондваны в начале Юры, а может быть и по ряду факторов. Так или иначе, мы имеем на данный момент совершенно не живородящих млекопитающих из группы монотрематы, как утконосы и ехидновые, а так же их ископаемых мезозойских предков с территорий Австралии. Однопроходные млекопитающие, по ряду признаков таковыми не являются и наиболее, напоминают морфологически непосредственных предков млекопитающих – цинодонтов.

Ну да вернёмся к нашим ретровирусам, собственно, откуда мы знаем об их роли в появлении живорождения у млекопитающих? Всё дело в плаценте, которой обладает каждое живородящее млекопитающее, как сумчатое, так и собственно - плацентарное. Плацента, это довольно любопытное образование, формирующее так называемый синцитий, это когда клетки сливаются между собой в как бы одну большую многоядерную клетку. Плацента выполняет ряд серьёзных функций необходимых для нормального протекания беременности, как то: газообмен, снабжение питательными веществами, выведение продуктов метаболизма плода, секреция ряда гормонов и… плацента умеет подавлять иммунитет матери.
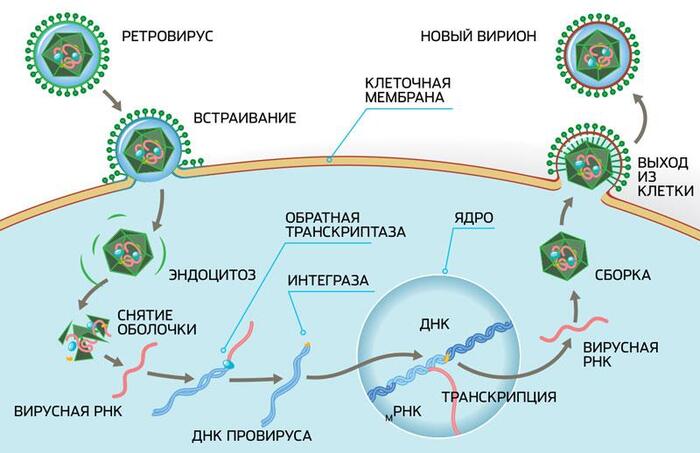
Это требуется для того, что бы организм матери не отторг находящийся внутри него чужеродный объект, в виде будущего чада. Срок беременности у териев гораздо больше, нежели инкубационный период у завропсид и даже однопроходных млекопитающих, ведь нахождение в утробе матери создаёт для развития плода наиболее комфортные условия - температура вокруг всегда одинаковая, кислорода и питательных веществ в достатке, да и ядовитые метаболиты выводятся исправно, а значит можно и не торопится с появлением на свет и спокойно развиваться до приемлемых размеров!

Итак, выделим два основных интересующих нас пункта свойственных плаценте: первое это слившиеся клетки, а второе – подавление иммунитета. Эти свойства удивительно напоминают действие одного из ретровирусных белков – ENV (от английского envelope - оболочка). Конечно же, вирус использует этот белок не для пользы матери и ребёнка, а с целью подавить иммунитет и, заставив клетки слиться, тем самым расширив свои охотничьи угодья и воспользовавшись всеми ресурсы синцития.
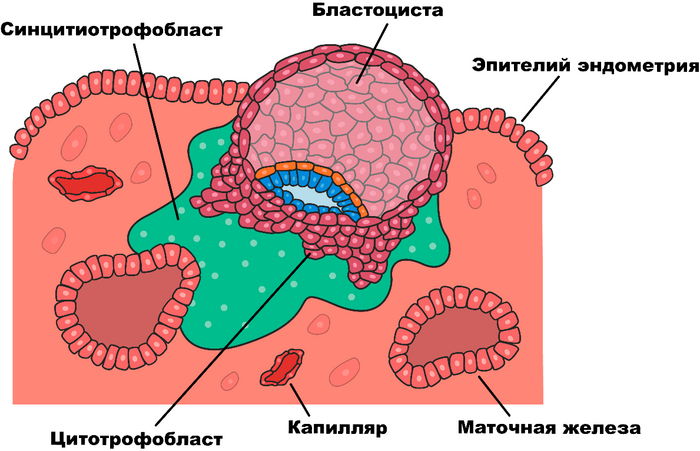
В 2000-м году, две группы учёных возглавляемых Джоном Мак-коем из США и Франсуа Малле из Франции, одновременно обнаружили в плаценте человека эндогенный ретровирус, который назвали Human Endogenous Retrovirus-W или HERV-W, белку оболочки вируса было присвоено имя синцитин, так как его функция напрямую связана с образованием синцития в плаценте. Так же обнаружилось, что если отключить ген, отвечающий за производство данного белка, то беременность протекает с патологиями вызванными дисфункцией сосудов. В 2003-м году был открыт ещё один ретровирус участвующий в образовании плаценты - HERV-FRD и его белок, названый синцитином второго типа.
Эти примеры показывают, что в ходе эволюции, млекопитающие научились вступать в симбиоз с ещё недавним смертельным врагом, научившись, в процессе естественного отбора, сохранять полезные функции, привнесённые в организм вирусом. Правда, как говорилось выше, у вируса нет задачи, помочь животному, клетки которого он поражает, а скорее наоборот. Так выяснилось что многие тяжелые заболевания человека, появляются у людей со следами активации эндогенных ретровирусов, это волчанка, сахарный диабет и рассеянный склероз. В случае последнего, подозрение падает на уже упоминавшийся выше синцитин, то есть, белок, без которого вивипарность невозможна, может оказаться причиной тяжелейшего заболевания. Получается, что польза от использования синцитина в образовании плаценты, превысила риск отдалённых негативных последствий, образовав своеобразный компромисс между млекопитающими и ретровирусом.
Теперь рассмотрим некоторые физиологические изменения у ранних млекопитающих. Как говорилось ранее, вероятно, выкармливать потомство неким секретом, пусть ещё и не полноценным молоком, синапсиды научились ещё задолго до появления у них живорождения, в пользу этого говорит невозможность появления развитого потомства, как у завропсид, из яйца как у однопроходных, следовательно, потомство нуждается в уходе и выкармливании.

Появление живорождения способствовало возможности более длительного вынашивания плода, а так же более эффективно защищало потомство от вероятной гибели до рождения, по сути, за некоторым исключением, сохранение жизни матери равнялось сохранению жизни детей, вплоть до рождения и с большой долей вероятности достижения ими пубертатного периода.
Но что бы появилось живорождение, необходимо избавится от такого органа как клоака, где отходы пищеварительной системы смешиваются с продуктами выделительной системы, сюда же выходят и протоки яйцеводов у женских особей. Так вот, клоака, присутствует абсолютно у всех млекопитающих, правда у большинства на зародышевой стадии, например у человека она исчезает в районе четвёртой-седьмой недели развития плода, после чего анальный канал отделяется и клоака образует мочеполовой синус в котором формируется мочевой пузырь, а так же внешние и внутренние половые органы.
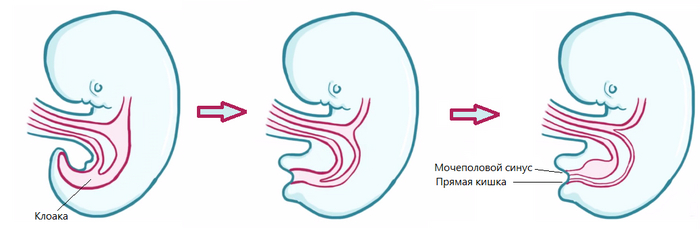
Итак, как уже говорилось выше, на фоне последствий от различных ретровирусных инфекций, клоака у ранних млекопитающих, могла начать трансформацию, разделив анальное отверстие и мочеполовой синус, что привело к образованию матки у женских особей. Точнее, маток сначала было несколько, что можно проследить по анатомии сумчатых и некоторых плацентарных, например: у сумчатых целых две матки и два-три влагалища, позволяющие вынашивать одновременно двоих детёнышей, на время, гормонально затормаживая, развитие младшего, вплоть до появления на свет старшего, при этом самцы сумчатых обладают раздвоенным пенисом, для более эффективного оплодотворения. У плацентарных двойная матка наблюдается у афротериев и ксенантров, а так же у многих грызунов, двурогая матка есть у хищных, насекомоядных, кито-парнокопытных и других эутериев, а вот простая матка присутствует только у приматов и рукокрылых.
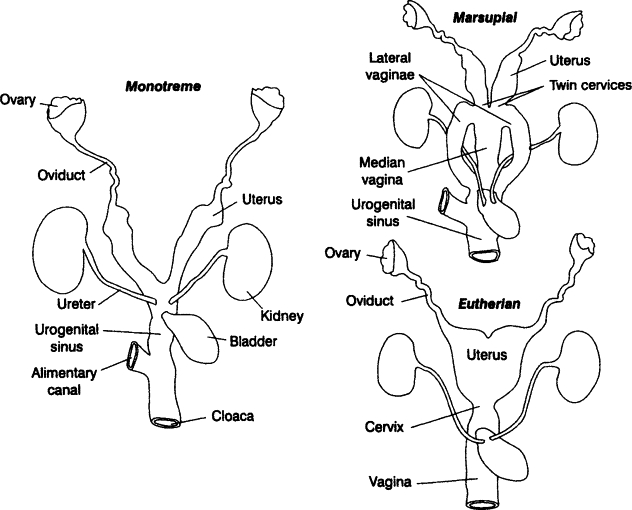
Так же, на примере сумчатых хорошо видна эволюция плаценты, по сути, беременность сумчатых частично напоминает этот процесс у яйце-живородящих завропсид: сначала зародыш начинает своё развитие в яйце внутри организма матери и лишь незадолго до родов вылупляется из яйца и прикрепляется слабой плацентой к эндометрию матки матери и через не очень длительное время происходят роды. Период имплантации зародыша к стенке матки, является самым опасным для жизни зародыша, около 75% неудачных беременностей у плацентарных приходятся именно на него.
Дело в том, что в момент прикрепления зародыша и начала образования плаценты, происходит воспаление стенки матки и наши добрые друзья Т-лимфоциты, спешат на помощь и с шашкой наголо, мчатся уничтожать непрошеного гостя, именно эта реакция вызывает тот самый токсикоз у беременных. Но тут, в дело вступает ретровирусный белок, который заставляет Т-лимфоциты не только не атаковать свежезакрепившийся плод, но и защищать его от чужеродных элементов. Затем, к моменту родов, воспаление возобновляется и происходит отторжение плаценты маткой, и непосредственно - роды. Но вот у сумчатых, плацента практически не влияет на иммунный ответ организма, поэтому основную часть своего развития, их детёныш проводит в яйце. Это подтвердили исследования домового опоссума, принадлежащего к сумчатым млекопитающим, опубликованные в 2000 году профессором биологии из Австралии Линни Селвуд.
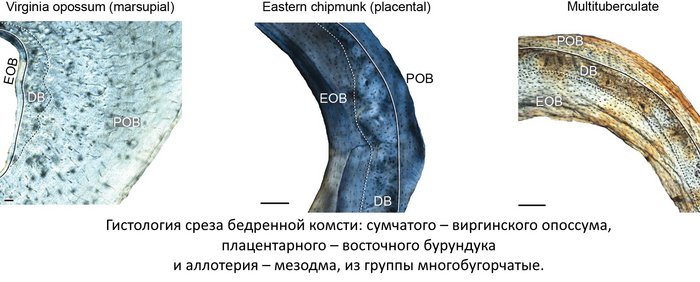
Другое же исследование, проведённое уже в 2022 году, группой американских учёных под руководством Лукаса Вивера, привело результаты сравнения гистологии бедренной кости трёх групп млекопитающих: сумчатых, плацентарных и аллотериев. Всё дело в том, что компактная костная ткань, делится на два состояния: незрелая костная ткань – формирующаяся у животного в периоды быстрого роста и зрелая костная ткань – образовывающаяся в периоды медленного роста организма и напрямую связанна с периодом молочного вскармливания, который у сумчатых, длится намного дольше чем у плацентарных. Сравнение срезов костей у плацентарных и аллотериев, оказалось во многом схожим по этим параметрам, и разительно отличающимся от таковых у сумчатых, таким образом, данная работа предполагает, что аллотерии – группа, считающаяся более древней, чем сумчатые и плацентарные, вероятно развивалась именно по плацентарному сценарию. Конечно, данное исследование, лишь предполагает плацентарность у аллотериев, но и в таком случае, это ещё один аргумент, в пользу появления живорождения на рубеже Триасового и Юрского периодов.
В итоге, мы имеем удивительный пример эволюционных изменений, произошедших в ходе ряда уникальных мутаций вызванных ретровирусной инфекцией, за весьма недолгое, по меркам эволюции время, около 20-40 миллионов лет, сделавших из яйцекладущих цинодонтов, более развитых – живородящих млекопитающих. И пусть относительно точная дата появления живорождения у синапсид пока ещё не определена, но у учёных уже всё-таки есть вероятные временные границы, когда данное событие могло произойти, а уж насколько ближе это будет к 200 или 160 миллионам лет назад, покажет только время, новые находки и исследования.
Даниил Симонов, «Помни о предках. Палеонтология», ноябрь 2022 г.
Список источников:
1. N. de Parseval , V. Lazar, J.-F. Casella, L. Benit, T. Heidmann: «Survey of Human Genes of Retroviral Origin: Identification and Transcriptome of the Genes with Coding Capacity for Complete Envelope Proteins», October 2003, DOI: 10.1128/JVI.77.19.10414-10422.2003
2. G. P. Wagner, E. M. Erkenbrack, A. C. Love: «Stress-Induced Evolutionary Innovation: A Mechanism for the Origin of Cell Types», September 2018, DOI:10.20944/preprints201809.0193.v1
3. L. N. Weaver, H. Z. Fulghum, D. M. Grossnickle, W. H. Brightly, Z. T. Kulik, G. P. Wilson Mantilla, M. R. Whitney: «Multituberculate Mammals Show Evidence of a Life History Strategy Similar to That of Placentals, Not Marsupials» September 2022 DOI: 10.1086/720410
4. C. J. Burgin, J. Widness, N. S. Upham: «Introduction to Illustrated Checklist of the Mammals of the World», November 2020, DOI:10.5281/zenodo.4270050
5. N. M. Foley , M. S. Springer, E. C. Teeling: «Mammal madness: is the mammal tree of life not yet resolved?», April 2016, https://doi.org/10.1098/rstb.2015.0140
6. M. C. Wallis, P. D. Waters, M. L. Delbridge, P. J. Kirby, A. J. Pask, F. Grützner: «Sex determination in platypus and echidna: autosomal location of SOX3 confirms the absence of SRY from monotremes. Chromosome Research», November 2007, doi:10.1007/s10577-007-1185-3
7. M. Renfree, G. Shaw: «Reproduction in Monotremes and Marsupials», May 2001 DOI:10.1038/npg.els.0001856
8. R. Motani , D. Jiang, A. Tintori, O. Rieppel, G. Chen: «Terrestrial Origin of Viviparity in Mesozoic Marine Reptiles Indicated by Early Triassic Embryonic Fossils», February 2014, doi:10.1371/journal.pone.0088640
9. D. W. Krause, S. Hoffmann, J. R.Wible, E. C. Kirk, J. A. Schultz, et al: «First cranial remains of a gondwanatherian mammal reveal remarkable mosaicism», November 2014, doi:10.1038/nature13922
10. J. A. Sharp, V. Modepalli, A. K. Enjapoori, H. E. Abud, C. Lefevre, K. R. Nicholas: «Milk: Milk of Monotremes and Marsupials. Reference Module in Food Science», 2016, doi:10.1016/b978-0-08-100596-5.00910-0
11. L. Selwood: «Marsupial Egg and Embryo Coats», May 1999, doi.org/10.1159/000016733








