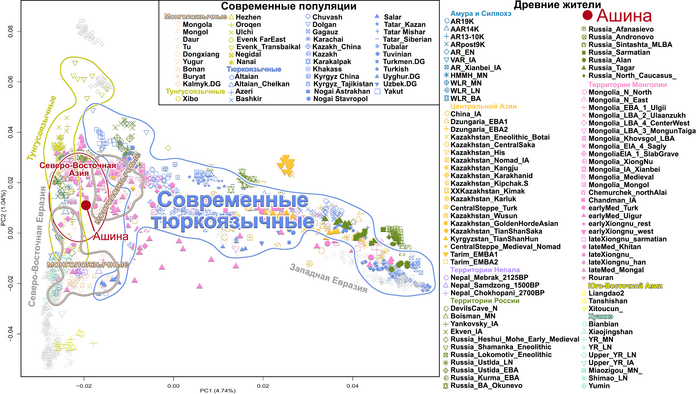
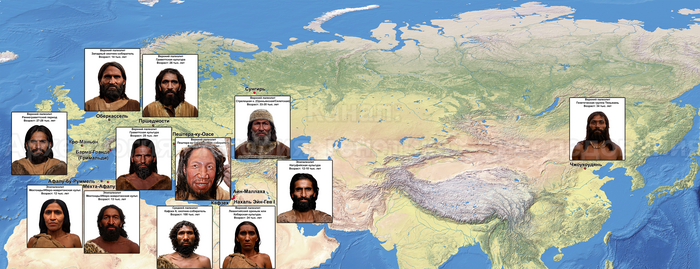
В статье речь пойдёт о генетических данных представительницы древних тюрков – принцессы, а после брака с императором китайско-сяньбийской династии Северной Чжоу - У-ди, императрицы – Ашины из одноимённого рода, дочери Мукан-кагана, третьего кагана древних тюрков – Ашина Сыцзинь, при котором Тюркский каганат расширил свои границы от Жёлтого до Чёрного моря, став крупнейшей империей того времени.
Кстати, о происхождении и внешности супруга Ашины - третьего императора китайско-сяньбийской династии Северной Чжоу - У-ди (Юйвэнь Юна) будет в следующей статье.
А в данном исследовании авторы, проанализировав генетический профиль императрицы Ашины, пытаются ответить на три ключевых вопроса:
1. Какое было происхождение у древних тюрков?
2. Есть ли генетическая связь между Ашиной и другими тюркскими народами, а также различными популяциями Центральной и Восточной степи после железного века?
3. Есть ли генетическая связь между древними тюрками и современными тюркоязычными народами?
Вступление
Первый расцвет тюркских народов пришелся на период с VI по VIII век, когда возникла могущественная кочевая империя в виде Тюркского каганата, основанного правителями из рода Ашина, простиравшегося от Монгольского плато до Каспийского моря. Появившись вслед за хунну, сяньби и другими кочевыми конфедерациями, влияние тюрков ощущалось по всей Евразии, что повлияло на языковой ландшафт значительной её части в течение следующих столетий. Тюркский язык, состоящий из более чем 40 языков, является самой многочисленной группой в гипотетической алтайской языковой семье, и около 200 миллионов человек говорят на тюркских языках более чем в 10 странах.
К сожалению, исторические сведения о Тюркском каганате носят разрозненный характер и часто противоречивы. Происхождение древних тюрков является предметом споров, но если не брать мифические истории с тотемными предками по типу появления от хуннского царевича и волчицы, их происхождение может быть сведено к трем конкурирующим гипотезам:
1. Представители племени Ашина выходцы из одноимённого племени хунну в Северо-Восточной Азии.
2. Их происхождение связано с Причерноморско-Каспийской степью или Центральной Азией с последующей миграцией на восток.
3. Сложносоставное происхождение в окрестностях Пинляна или Гаочана в Турфанской впадине, охватывающее как восточные, так и западные евразийские этнические группы.
Поэтому происхождение представителей правящего рода из племени Ашина вызывает большой интерес и является ключом к разгадке происхождения древних тюрков. Тем более с учётом того, что среди тюркской знати была широко распространена кремация.
Результаты
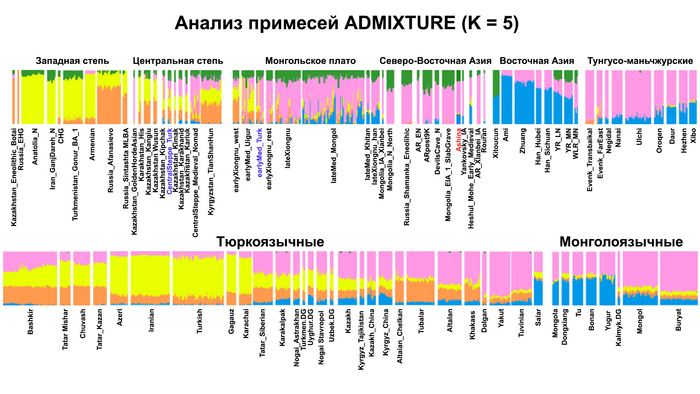
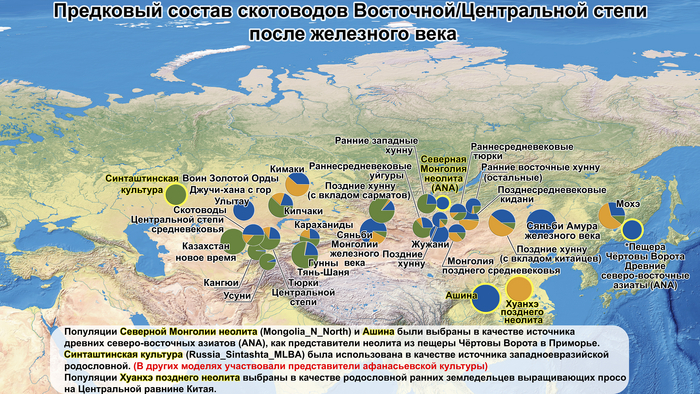
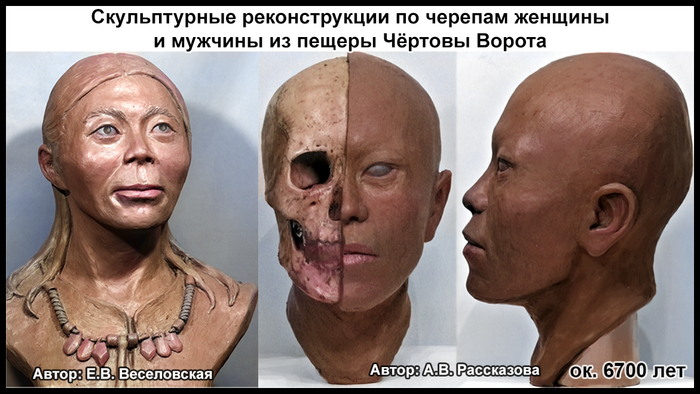
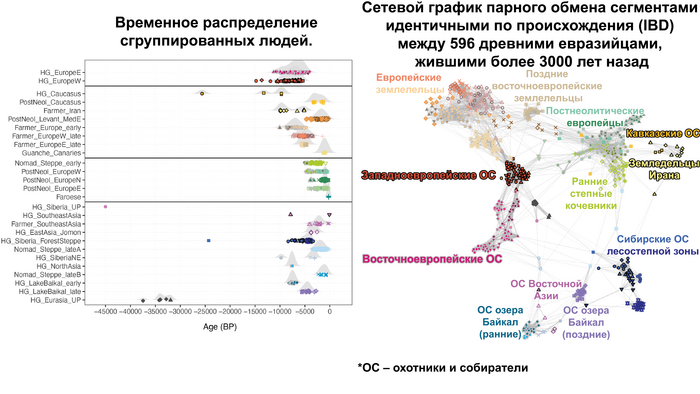
По данным мтДНК императрица Ашина принадлежала к гаплогруппе F1d, распространенной в Северо-Восточной Азии. А по результатам полногеномных анализов она группировалась с древними популяциями Северо-Восточной Азии и востока Монгольского плато, но особенно, с древними северо-восточными азиатами, как представители неолита из пещеры Чёртовы Ворота, возрастом около 6700 лет и носители бойсманской культуры среднего неолита из археологического памятника Бойсмана-2, Приморья, представителями неолита Амура и Северной Монголии, а также с кочевниками Восточной степи после железного века, включая сяньбийцев, жужаней, киданей и части монгольского населения. Из современных популяций она была генетически близка с современными тунгусо-маньчжурскими и монголоязычными народами.
При этом вклада от древних китайцев, связанных с жителями реки Хуанхэ у Ашины не было обнаружено. Однако отмечен небольшой вклад от жителей Западной Евразии – 2,3-3,9%, включающий степную родословную с примесью анатолийских земледельцев, который произошел у предков Ашины за 1566 ± 396 лет до её жизни. В целом генетические данные опровергают гипотезы о западно-евразийском и множественном происхождении представителей знатного рода тюркских каганатов – Ашина в пользу восточноазиатского происхождения.
Однако стоит отметить, что у других образцов в тюркский период наблюдалось смешение компонент древних северо-восточных азиатов, которых в среднем было более половины, с компонентами западноевразийских популяций Бактрийско-Маргианского археологического комплекса и кочевников степи железного века, что также соответствует заметному увеличению гаплогрупп Y-хромосомы R и J, в течение тюркского периода в Монголии.
Есть ли генетическая связь между Ашиной и другими тюркскими народами, а также популяциями Центральной и Восточной степи после железного века?
Начиная с железного века в Восточной Степи возникали и рушились, выходящие далеко за её пределы, различные государственные образования кочевников: хунну, сяньби, жужани, тюрки, уйгуры, кидани, кыргызы и – наиболее агрессивные и успешные из всех – монголы. Соседняя средняя часть Евразийской степи или Центральная степь, также была свидетелем перемен в судьбах кочевых образований: гунны, усуни, кангюи, карлуки, кимаки, караханиды и кыпчаки.
Однако представительница элиты Тюркского каганата продемонстрировала генетическую связь только с древнемонгольскими племенами сяньбийцев, киданей и жужаней, а также с представителями тунгусо-маньчжурских племён мохэ. А вот монголы, уйгуры, карлуки, кимаки, кипчаки, а также караханиды были отдалены от кластера Ашины. Хунну вообще был союз разных по происхождению племён, они сильно отличались между собой по географии и по времени. Некоторые западные их представители вообще на 68,4% имели компоненты как у людей из афанасьевской культуры.
Кстати в моделях надо адекватно воспринимать компоненты, они подразумевают схожие генофонды, а не непосредственных представителей культур.
Среди скотоводов Центральной степи усуни, кангюи и гунны Тянь-Шаня большую часть своего происхождения (62,4–73%) ведут от западных степных групп афанасьевской культуры.
А тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, имеют 35–50,6% компонент как у афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но доля восточно-евразийского происхождения увеличилась у средневековых скотоводов Центральной степи с 67,3% до 82,5%, за некоторым исключением. Однако стоит отметить, что ещё в новое время на территории Казахстана встречались люди с генетическим профилем на 75,5% происходящем от западных степных скотоводов. При этом Ашина не продемонстрировала близкого генетического родства с тюрками Центральной степи или раннего средневековья. Но в отличие от Ашины, тюрки Центральной Степи и раннесредневековые тюрки продемонстрировали высокую, но вариативную степень западно-евразийского происхождения, что указывает на существование генетической подструктуры ещё у древних тюркоязычных народов.
Есть ли генетическая связь между древними тюрками и современными тюркоязычными народами?
В скотоводческих образованиях Восточной Степи утверждалось, что распространение монгольских языков было связано с сяньбийскими, жужаньскими, киданьскими и монгольскими группами, в то время как хунну, тюрки и уйгуры считались связанными с распространением тюркских языков. В последнем случае были выдвинуты гипотезы о двух волнах распространения: булгаро-тюркской диффузии, начавшейся в гуннский период, спровоцированной более ранней экспансией хунну, и последующей демографической экспансии, связанной с Тюркским каганатом. При этом во втором и третьем веках нашей эры Центральная степь была населена ираноязычными группами, которые постепенно переходили на тюркские языки.
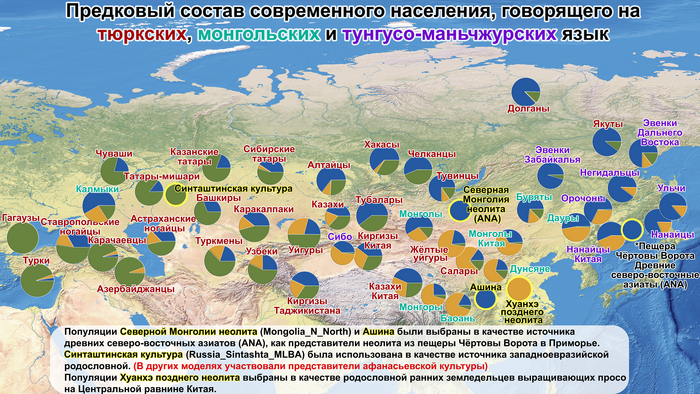
Поэтому генетические данные не подтверждают связи между древними и современными тюркоязычными группами и указывают на значительные различия между тюркоязычными популяциями и популяциями, связанными с распространением тюркских языков.
А пропорции предков древних северо-восточных азиатов, которые составляли основу древних тюрков, варьировались у тюркоязычных групп и даже полностью отсутствовали у некоторых самых западных их представителей. Это указывает на то, что распространение тюркских языков было в основном обусловлено культурными факторами, а не демографической диффузией и интеграцией населения. Поэтому натягивать современные тюркоязычные популяции на тюркизированные древние в качестве доказательства тюркоязычности этих древних популяций абсолютно не имеет смысла и противоречит всем данным.
Пояснение
А подвести итог хочу простым языком, отклонившись от строгой научной линии повествования. Ещё раз: это не научная часть!
Древние тюрки, и тем более из рода Ашина, по происхождению были близки к древним северо-восточным азиатам, с незначительным вкладом от западных степных скотоводов. По реконструкциям понятно даже как они выглядели. Это характерные отличия.
Древние и современные тюрки имеют разное происхождение. И многие современные тюркоязычные народы объединяет лишь язык.
Отдельные представители тюркских народов в поисках своих предков почему-то обращаются к древним тюркам, однако схожих с ними компонент у них не так много, а у некоторых вообще почти нет. Их генофонд состоит из смеси различных древних популяций, говоривших на разных языках, при этом у разных народов и разный генетический коктейль и своя интересная история. Но если древних тюрков начинать искать от современных, то можно выйти на разные популяции, которые внесли свой вклад в современные и намеренно или искренне заблуждаясь, объявить их тюрками.
Характерным примером таких манипуляций выглядит якобы доказательство тюркоязычности скифов, основанное только на словах из исследования 2017 года, что современные потомки западно-скифских групп встречаются среди различных современных групп на Кавказе и в Центральной Азии, в то время как сходство с восточно-скифскими группами встречается более широко, но почти исключительно среди современных тюркоязычных групп, особенно из кыпчакской ветви тюркского языка, которые в прошлом были кочевниками. Что не удивительно с учётом огромного генетического разнообразия этих групп, значительно большего чем у всех народов Европы вместе взятых. Трудно промахнуться в стадо слонов из дробовика. К тому же ещё и с учётом того, что сами тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, как уже было сказано выше, имеют 35–50,6% предков от афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но они же по определению тюрки, и мы не называем кыпчаков индоевропейцами или монголами с китайцами! С такой логикой если в Анадыре с вами поздоровался и разговаривает чукча на чисто русском языке, то его предки по мнению таких вот специалистов в кавычках, должны быть исключительно русскими.
Работа о скифах 2017 года: Unterländer, M., Palstra, F., Lazaridis, I. et al. Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat Commun 8, 14615 (2017). https://doi.org/10.1038/ncomms14615
И это исследование 2017 года в комментариях, не цитировал разве что только ленивый защитник тюркского происхождения скифов и не только. И продолжат это делать, вот увидите. При этом само исследование они не читали и вряд ли понимают…
Толкование скифских слов из тюркского несостоятельно хотя бы потому, что фонетика засвидетельствованных скифских слов противоречит фонетическим характеристикам пратюркского (распад которого по глоттохронологическим данным относится к I в. до н. э., т. е. много позже появления на исторической арене скифов – соответственно, сравнивать скифские слова имело бы смысл только с пратюркскими, а не с их рефлексами в отдельных древних и современных тюркских языках). Так, пратюркское слово не могло начинаться на сонанты *m
и *l, тогда как нам известны скифские имена Мадий и Липоксай; невозможными в пратюркском были и скопления согласных в анлауте (начале слова), засвидетельствованные в самоназвании скифов и таких именах, как, например, Спаргапиф. Одним словом, попытки объявить скифов тюрками представляют интерес разве что с точки зрения современной этнопсихологии. Сергей Кулланда http://генофонд.рф/?page_id=3267
1/3
Интервью одного из авторов работы о скифах 2017 г., руководителя межинститутского сектора молекулярной палеогенетики Института цитологии и генетики СО РАН, Института археологии и этнографии СО РАН Александра Пилипенко на научно-популярном сайте elementy.ru
К примеру, от союза русского и китаянки положение их детей может быть на том же графике где-то около ногайцев или ближе к казахам.
*Очень грубый пример
И по пропорциям главных компонент они будут на них похожи, но ведь нельзя же будет их ребенка назвать казахом или ногайцем. Часто вижу непонимание этого элементарного биологического аспекта в комментариях. Но когда нужно что-то доказать о себе любимом и, тем более, великом, тут логика ломается.
Или упор часто делают на то, что если кто-то до наших дней сохранил традиции, связанные с кочевым образом жизни, питанием и тому подобно… То только его предки в прошлом могли этим заниматься. Мне вот интересно, чем занимались по такой логике предки тех, кто сейчас компьютерные технологии развивает или в космос ракеты запускает? А так и бомжам можно сказать, что они продолжатели славных традиций охотников и собирателей… Уж пусть не обижаются на меня современные кочевники и скотоводы, а также их потомки. В сохранении традиций нет ничего плохого, если не сочинять себе историю…
Примечание: текста много, тема сложная и состоит из двух постов. Разделы – "Социокультурные изменения" и "Итоги" (Появление и сокращение генетического разрыва), во второй части.
Вступление
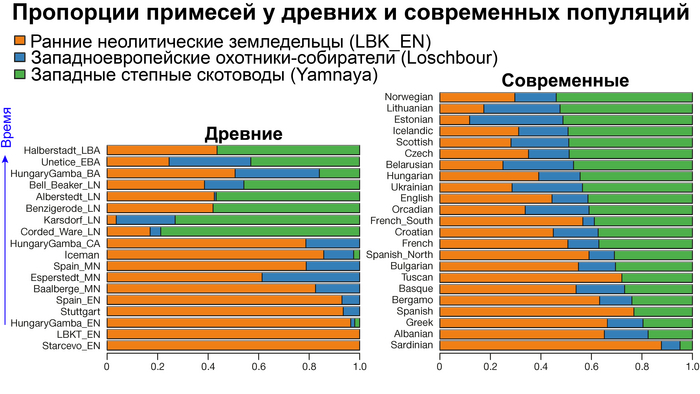
Генетическое разнообразие человеческих популяций Западной Евразии в значительной степени сформировалось в результате трех основных доисторических миграций:
1. Миграции охотников-собирателей, а именно анатомически современных людей, находящихся на ранней стадии социально-экономического развития, которые занимали территорию, примерно, с 45 тыс. лет назад. При этом вклад в последующие популяции внесли выжившие их линии после последнего ледникового максимума около 25-19 тыс. лет назад.
2. Далее, гораздо ещё более существенный вклад, внесли ранние неолитические земледельцы, пришедшие с Ближнего Востока примерно с 11 тыс. лет назад.
3. Степные скотоводы, распространившиеся из Причерноморско-Каспийской степи около 5000 лет назад, привнесли третью основную генетическую компоненту в популяции Западной Евразии.
Аутосомный генофонд практически всех европейских популяций по большей части состоит из различных пропорций этих предковых компонент, при этом независимо какие у них гаплогруппы.
Европейский континент заселялся людьми современной анатомии в несколько волн, при этом не все линии этих людей были предками современных. Первые «сапиенсы» Европы (более 40 тыс. лет назад) пересекались и скрещивались с неандертальцами, пока те совсем не вымерли. Об этой примеси свидетельствуют геномы людей из пещеры Бачо Киро в Болгарии, связанных с археологической культурой начального верхнего палеолита, и со стоянки Пештера-ку-Оасе в Румынии. Другие люди того периода, такие как женщина из пещеры Златый кунь Чехии и человек из Усть-Ишима России, не несут в своих геномах значительного количества неандертальского вклада, чем другие неафриканские группы. При этом люди, жившие в Европе более 40 тыс. лет назад, не оставили существенных следов в генетическом составе современных евразийских популяций, что говорит о том, что они в значительной степени вымерли или были ассимилированы последующими группами людей современной анатомии.
В те времена, на демографию древних людей влияли различные факторы, в том числе и климатические. Пик похолодания во время последнего ледникового максимума (LGM) был около 21 тыс. лет назад, когда Северная Европа была в значительной степени покрыта льдом, а южная граница ледниковых щитов проходила через Германию и Польшу. Многие участки Европы стали холодными пустынями с тундростепями и лесостепями на побережье Средиземного моря. А люди и животные переместились в южные широты, о чем свидетельствует, например, солютрейская культура на Пиренейском полуострове и на юге Франции. Другими предложенными рефугиумами или климатическими убежищами в тот период были Апеннинский полуостров, Балканы и юго-восточная часть Европейской равнины.
А в результате послеледниковой колонизации, палеогенетический анализ выявил разделение популяций охотников-собирателей на западных и восточных. Источники родословной западных охотников-собирателей (WHG) связаны с носителями эпиграветтской, азилийской и других эпипалеолитических культур генетического кластера Виллабруна, появление которого обусловлено миграциями в конце ледникового периода и появлением новой генетической компоненты, связанной с Ближним Востоком того времени, начиная с бёллинг-аллерёдского потепления около 14 тыс. лет назад, что археологически коррелирует с культурными переходами.
А родословная восточных охотников-собирателей (EHG) демонстрирует дальнейшую примесь с верхнепалеолитическими популяциями Сибири - древними северными евразийцами (ANE).
Предковый состав западных охотников-собирателей (WHG) был регионально изменчивым в мезолитических популяциях. Если охотники и собиратели из Британии и северо-западной континентальной Европы продемонстрировали однородный состав от палеолита и мезолита, а также вплоть до неолитической экспансии около 6000 лет назад, то их коллеги из Испании, представляли собой смесь людей из двух рефугиумов, когда к генетическому профилю кластера Виллабруна, добавляется примесь от более ранних местных пиренейских охотников-собирателей, связанных с солютрейской и мадленской культурами верхнего палеолита.
Этот генетический профиль сохранился во время последнего ледникового максимума в Юго-Западной Европе около 25-19 тыс. лет назад.
А время появления примеси сформировавшей восточных охотников-собирателей (EHG), оценивается в 15–13 тыс. лет назад, однако её пропорции в популяциях, по-видимому, соответствуют клине, которая в целом коррелирует с географией, так охотники-собиратели Прибалтики и Украины демонстрируют большее сходство с предками кластера Виллабруна, чем охотники и собиратели западной части современной России, у которых было больше вклада от древних северных евразийцев (ANE).
В свою очередь скандинавские охотники и собиратели продемонстрировали различные пропорции смеси западных и восточных охотников и собирателей (WHG и EHG) в позднем мезолите.
Стоит отметить, что охотники и собиратели мезолита и тем более палеолита, были очень разнообразны и неоднородны, но из-за ограниченного количества данных, наши знания о мезолитической структуре населения и процессах демографического смешения долгое время были скудны и содержали существенные хронологические и географические пробелы в информации. Отчасти это объясняется малым количеством хорошо сохранившихся мезолитических человеческих останков возрастом более 8000 лет, а отчасти и потому, что большинство исследований древней ДНК, посвященных периодам мезолита и неолита, были ограничены преимущественно находками из Европы.
1/3
Хотя давно напрашивается более детальная классификация охотников и собирателей по их характерным отличиям. Так, археологические данные указывают на условную границу от восточной Балтики до Черного моря, к востоку от которой сообщества охотников и собирателей существовали гораздо дольше, чем в Западной Европе, несмотря на аналогичное расстояние до центра распространения раннего земледелия на Ближнем Востоке.
Компоненты восточного и западного происхождения у охотников-собирателей в этом пограничном регионе сильно различаются, но более точные представления о демографических процессах, связанных с разделением на восток и запад и о динамике численности населения к востоку от Европы, включая Северную и Центральную Азию за тот же период времени, весьма ограничены. В этих регионах термин «неолит» характеризуется культурными, экономическими и социальными изменениями, а также использованием керамики. К примеру, неолитические культуры степей Центральной Азии и таежного пояса России уже изготавливали керамику, однако продолжали вести прежний образ жизни с присваивающей экономикой, как у предшествующих мезолитических культур охотников-собирателей, с различными инновациями в зависимости от условий среды обитания. Однако пока недостаточно данных для понимания того, как неолитизация отличалась по своим срокам, механизмам и последствиям в разных уголках Евразии.
В Европе, кроме восточной её части, переход от охоты и собирательства к земледелию сопровождался прибытием мигрантов ближневосточного происхождения, практикующим производящее хозяйство на основе одомашненных растений и животных. С последующим значительным ростом населения. В Юго-Восточной, Центральной и Западной Европе анатолийские земледельцы впоследствии в некоторой степени смешались с местными охотниками и собирателями, а на территории Британии, прибывшие с континентальной Европы ранние европейские земледельцы полностью заменили местных охотников и собирателей.
А вот в восточно-балтийском регионе наблюдается заметно иная модель неолитизации, с появлением одомашненных животных только при появлении культуры шнуровой керамики (CWC) около 4800 лет назад. Аналогичным образом в Северном Причерноморье люди с генетическим профилем мезолитических охотников и собирателей тысячелетиями сосуществовали с земледельческими группами, расположенными дальше на запад.
И третий крупнейший генетический пласт в истории европейских популяций связан со степными скотоводами раннего бронзового века, как представители ямной культуры около 5 тыс. лет назад. Этот генетический профиль, быстро распространился по Европе посредством экспансии как носителей культуры шнуровой керамики (CWC), так и родственных ей культур.
Хотя предыдущие исследования выявили эти крупномасштабные миграции в Европу и Центральную Азию, основные аспекты, касающиеся демографических процессов, не решены. Ямная или степная родословная в общих чертах охарактеризована как смесь восточноевропейских (EHG) и кавказских охотников-собирателей (CHG), образовавшаяся в результате гипотетической смеси «северного» степного и «южного» кавказского источников. Однако точное происхождение этих предковых источников не было установлено. Более того, за некоторым исключением, известные гаплогруппы Y-хромосомы представителей ямной культуры не соответствуют гаплогруппам, обнаруженным у европейцев после 5000 тыс. лет назад и происхождение этой отцовской линии также не установлено. К тому же "степная" родословная в Европе до сих пор была выявлена только в смешанной форме, а детали и механизмы, с помощью которых эта родословная впоследствии распространилась вместе культурой шнуровой керамики (CWC), оставались неясными.
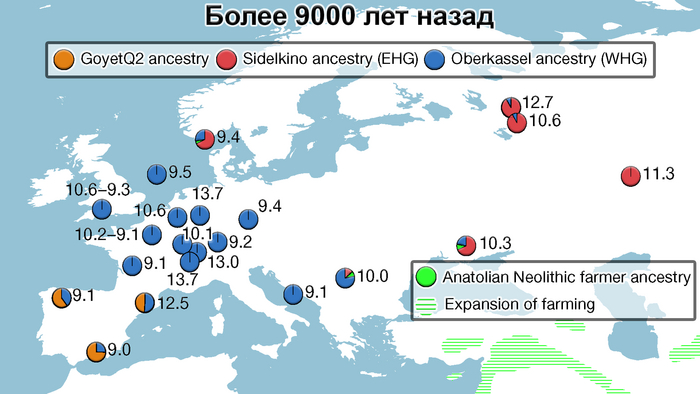
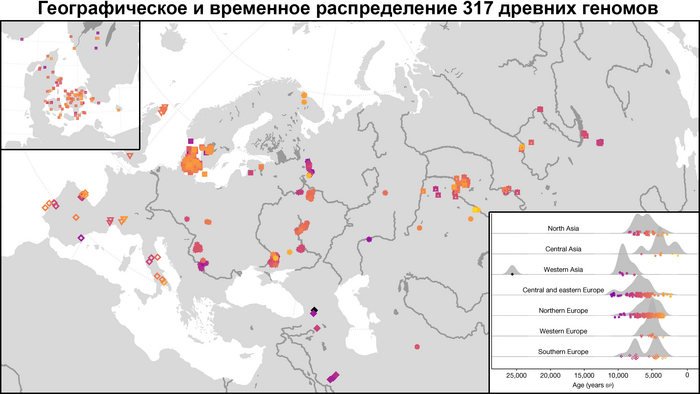
В новой работе учёные под руководством Эске Виллерслева из образцов, датированных радиоуглеродным методом при помощи ускорительной масс-спектрометрии, секвенировали геномы 317 человек, 309 из них датируются периодом от 11 до 3 тысяч лет назад.
А совместно с ранее опубликованными данными получилась выборка из 1600 древних геномов, секвенированных с помощью метода дробовика.
Географически 317 отобранных образцов охватывают обширную территорию Евразии, по долготе от Атлантического океана до озера Байкал, а по широте от Скандинавии до Ближнего Востока, и происходят из контекстов, которые включают курганы, пещеры, болота и дно водоёмов.
Географически образцы можно разделить на три региона:
1. Центральная, Западная и Северная Европа.
2. Восточная Европа, включая западную часть России, Беларусь и Украину.
3. Урал и Западная Сибирь.
Образцы включают представителей многих ключевых культур Западной Евразии мезолита, неолита и последующих периодов, такие как маглемозе, эртебёлле, воронковидных кубков (TRB) и шнуровой керамики или одиночных погребений в Скандинавии; импрессо или кардиумной керамики в Средиземноморье; кёрёш и линейно-ленточной керамики (LBK) в Юго-Восточной и Центральной Европе; и многие археологические культуры Украины, западной части России и Зауралья (как веретьё, льяловская, волосовская, китойская и др.). Выборка была особенно плотной в Дании, где сотня геномов, охватывала период от раннего мезолита до бронзового века. Обширная выборка была также получена из Украины, Западной России и Зауралья, охватывая период от раннего мезолита до неолита, примерно до 5000 лет назад.
Результаты
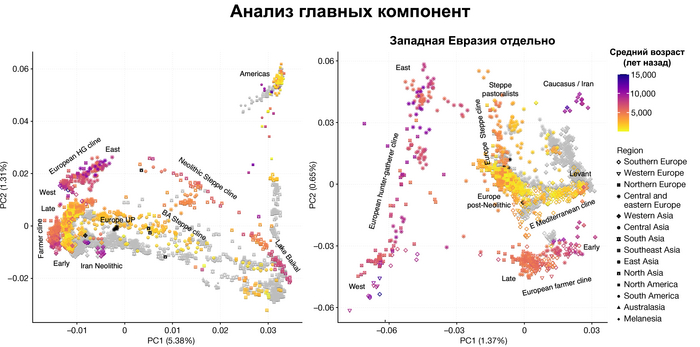
Полученный набор генетических данных позволил при анализе главных компонент использовать за основу древние геномы, и уже на них спроецировать положение современных популяций, а не наоборот, как это было во многих предыдущих исследованиях.
И хотя результаты согласуются с тем, что древние люди сильнее друг от друга генетически отличались, чем современные, такой подход позволил выявить ещё более значительные генетические различия между древними людьми, которые ранее не наблюдались, что отражает меньший эффективный размер их популяции и большую генетическую изоляцию.
Чтобы получить более точную генетическую характеристику в пространстве и времени, авторы использовали попарную идентификацию сегментов ДНК идентичных по происхождению, чтобы сгруппировать анализируемых людей в связанные кластеры сходного генетического происхождения, а не по культурным или территориальным признакам.
В более крупном масштабе они отражали ранее описанные группы западных и восточных охотников-собирателей, а вот в более мелком масштабе они сгруппировались не только в пределах ограниченных пространственно-временных диапазонов и/или археологических контекстов, но также продемонстрировали ранее неизвестные связи в более широких географических диапазонах.
Полученные кластеры далее использовались для моделирования состава предков последующих популяций.
Популяционная структура охотников и собирателей после максимума последнего оледенения (LGM)
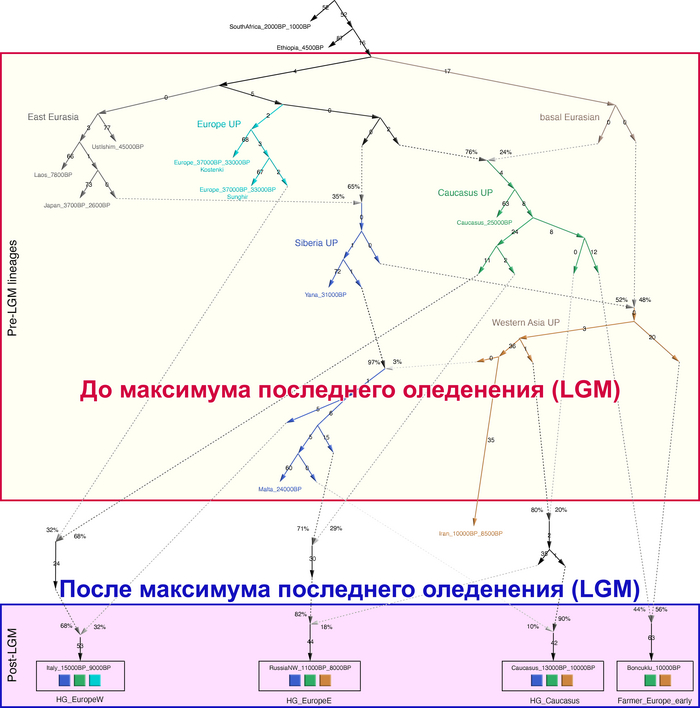
До последнего ледникового максимума, популяции Евразии довольно сильно отличались и в разной степени смешивались с неандертальцами. Некоторые популяции пока остаются гипотетическими, как к примеру, так называемые, «базальные евразийцы» с незначительной или нулевой примесью от неандертальцев, их прямых образцов, подтвержденных генетикой, на данный момент нет, но генетические следы этой популяции были обнаружены у охотников-собирателей Кавказа и Западной Азии верхнего палеолита, а также у более поздних западноевропейских охотников и собирателей.
В данной работе авторы сообщают о генетических данных полученных из зуба женщины верхнего палеолита, жившей около 26-25 тыс. лет назад в окрестностях карстового грота Котиас Клде в современной Грузии, не путать с более молодыми образцами кавказских охотников-собирателей из этого археологического комплекса, у которых было менее 20% этой кавказской родословной верхнего палеолита. А сам геном этой представительницы Кавказа верхнего палеолита представляет собой смесь 76% западноевразийских охотников-собирателей верхнего палеолита (т.е. до максимума последнего оледенения) и 24% «базальных евразийцев». Кстати популяции Ближнего Востока верхнего палеолита, почти наполовину произошли от «базальных евразийцев», которые, практически не имели примеси неандертальцев и отделились от других неафриканских линий ещё до их разделения между собой. При этом западноевропейские охотники-собиратели, жившие после последнего ледникового максимума, моделируются как итоговая, за несколько событий, смесь 46% популяций, связанных с Кавказом верхнего палеолита, 22% европейцев, живших около 33-37 тыс. лет назад, связанных со стоянками Сунгирь и Костёнки и 32% древних популяций, связанных с Сибирью верхнего палеолита, как представитель стоянки Мальта, возрастом 24 тыс. лет, т.е. древних северных евразийцев.
В свою очередь восточноевропейские охотники-собиратели моделируются как смесь 58% популяций, связанных с обитателями стоянки Мальта в Прибайкалье, 28% популяций, связанных с Кавказом верхнего палеолита, как женщина из Котиас Клде (но в разные периоды, сначала 24%, а потом 4%) и 14% представителей Западной Азии верхнего палеолита. Эта смесь также проходила в несколько этапов. Тем самым новые данные подтверждают генетическое разделение охотников и собирателей в Восточной и Западной Европе после максимума последнего оледенения, когда люди снова расселялись по Евразии, при этом эти различия сохранялись на протяжении всего мезолита.
В анализе примесей, эти сложносоставные генофонды представлены отдельными компонентами. А различия между ними, а именно между западноевропейскими и восточноевропейскими охотниками-собирателями мезолита максимально выражены у представителей Италии, возрастом от 15 до 9 тыс. лет и Северо-Запада современной России возрастом от 11 до 8 тыс. лет.
Обратите внимание, что генетический состав одинаков даже у людей с разными гаплогруппами и наоборот, по всему геному люди с одинаковыми линиями гаплогрупп отличаются.
Кстати, в представлении сложных процессов, не стоит забывать о временных промежутках, о которых идёт речь.
Что касается североевропейских охотников и собирателей, то они отличались по пропорциям смеси компонент как у западноевропейских и восточноевропейских охотников-собирателей, за исключением Дании, где популяции продемонстрировали южноевропейское происхождение и были генетически схожи с охотниками-собирателями из Италии, возрастом от 15 до 9 тыс. лет. При чём эта родословная сохранялась в регионе на протяжении 5 тыс. лет.
В целом результаты указывают на то, что существовало по меньшей мере три отчетливые волны миграций охотников и собирателей в Скандинавию:
1. Первая волна связана с миграцией людей южноевропейского происхождения на территорию Дании и прибрежной части Юго-Западной Швеции.
2. Вторая волна в Прибалтику и Юго-восточную Швецию связана с популяциями из Юго-Восточной Европы.
И 3. Третья волна миграций на крайний север Скандинавии, а затем на юг вдоль атлантического побережья Норвегии связана с охотниками и собирателями с северо-запада современной России.
Эти миграции, вероятно, представляют собой послеледниковую экспансию из рефугиумов или районов-убежищ, общих как для людей, так и для многих видов растений и животных.
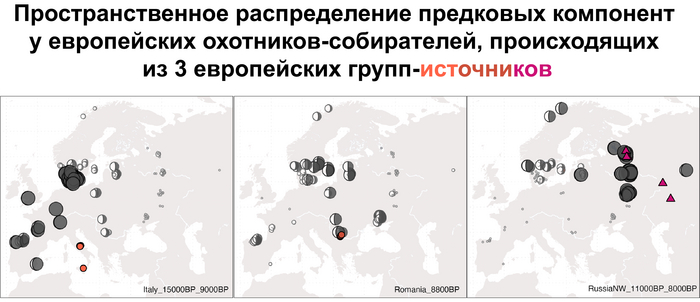
Стоит отметить, что генофонд европейских охотников-собирателей верхнего палеолита, живших до максимума последнего оледенения был в значительной степени заменён генофондом, происходящим из рефугиума в Южной Италии. На Пиренейском полуострове самые древние образцы из этого исследования, в том числе охотника-собирателя, возрастом около 9500 лет (NEO694) из бухты Санта-Майра на востоке Испании, показали преимущественно южноевропейское происхождение с незначительным вкладом охотников и собирателей верхнего палеолита, который, вероятно, отражает генофонд, связанный с мадленским периодом до максимума последнего оледенения.
А более поздние жители севера Пиренейского полуострова, были ближе к охотникам и собирателям Юго-Восточной Европы, демонстрируя около 40–30% вклада балканских охотников-собирателей.
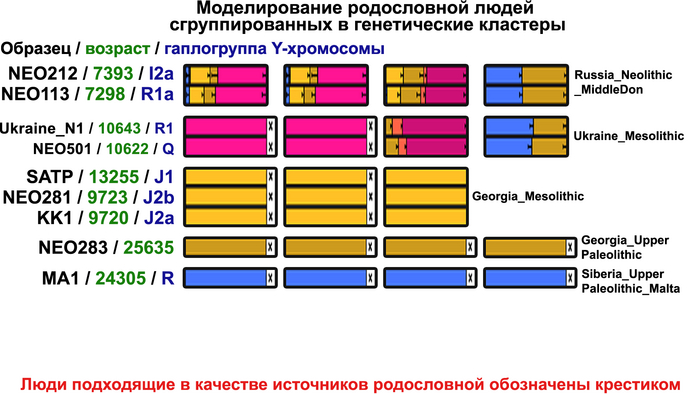
Поток генов, связанных с охотниками-собирателям Юго-Восточной Европы был направлен и в Восточную Европу после мезолита, на территорию современной Украины и далее. А у двух человек из могильника мариупольского типа раннего энеолита (NEO212, NEO113 ) близ хутора Голубая Криница на юге Воронежской области, возрастом около 7300 лет, генетический профиль состоял уже из смеси более ранних охотников-собирателей с территории Украины с вкладом от 18 до 24% из источника, связанного с охотниками-собирателями Кавказа.
1/2
Поэтому на графике анализа главных компонент они отклонены в сторону Ирана и Кавказа. По расчётам эта примесь произошла примерно 8300 лет назад. Результаты указывают на более ранний генетический контакт между популяциями Кавказа и степи ещё до появления культур кочевников в Причерноморско-Каспийской степи. Помимо этого, этот контакт задокументирован западнее, чем предполагалось ранее.
Основные генетические изменения в Европе
Предыдущие исследования древней геномики задокументировали несколько эпизодов крупномасштабной смены населения в Европе за последние 10 тыс. лет, но 317 новых геномов из данной работы, заполняют важные пробелы в наших знаниях.
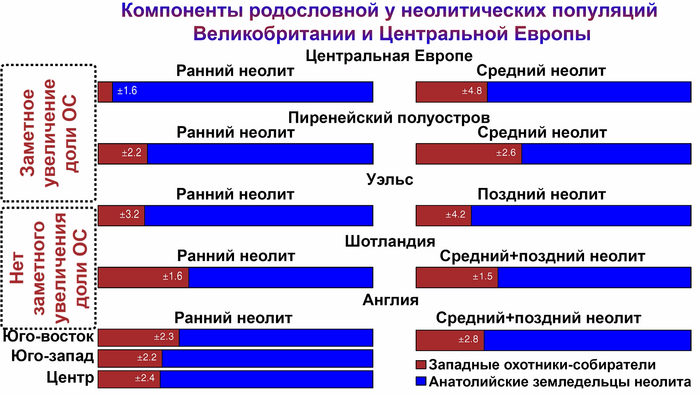
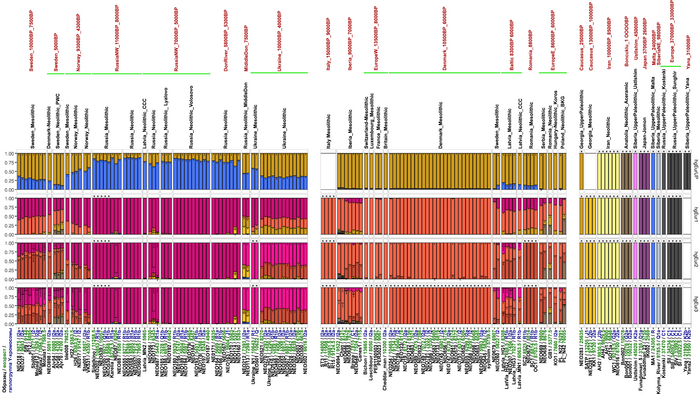
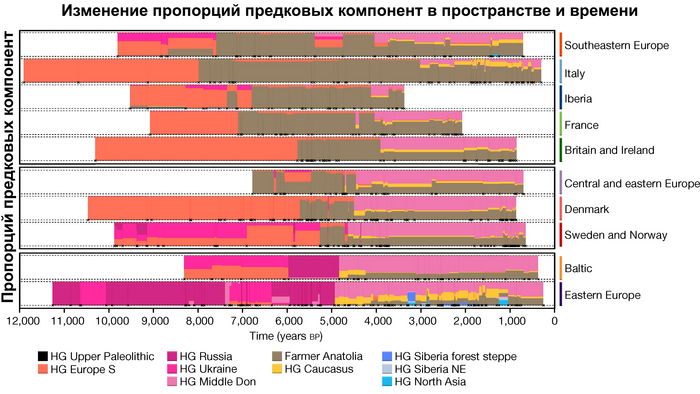
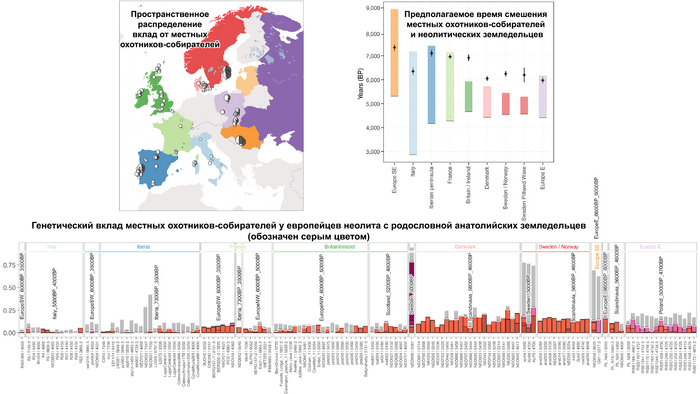
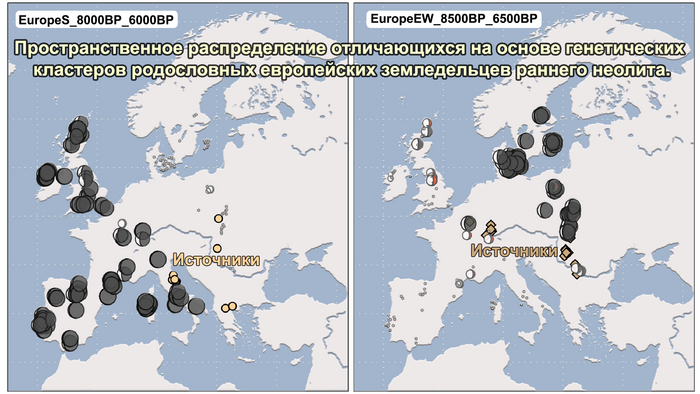
Анализ выявил глубокие различия в пространственно-временной динамике неолитизации между восточной и западной частями Европы вдоль пограничной зоны, простирающейся от Черного моря до Балтийского. К западу от этой пограничной зоны, переход к неолиту сопровождался крупномасштабными сдвигами в генетической родословной, от местных охотников-собирателей к земледельцам анатолийского происхождения (Boncuklu_10000BP). Этот переход в зависимости от региона охватывал обширный период времени, более 3 тыс. лет, от самых ранних свидетельств на Балканах около 8700 лет назад до 5900 лет назад в Дании.
В последующие периоды смешение между ранними европейскими земледельцами и местными охотниками-собирателями привело к частичному увеличению родословной охотников и собирателей во многих регионах Европы, но этот вклад редко превышал 10%, помимо заметных исключений, наблюдаемых у людей из района Железных ворот на границе современных Сербии и Румынии, а также у носителей культур ямочной керамики из Швеции и кардиумной керамики раннего неолита из Португалии, которые, по оценкам, содержат от 27 до 43% вклада от охотников и собирателей, как представители Пиренейского полуострова, возрастом от 9 до 7 тыс. лет (Iberia_9000BP_7000BP).
Обитатели эпохи неолита с территории современной Дании продемонстрировали до 25% генофонда охотников-собирателей (EuropeW_13500BP_8000BP), но преимущественно не местных.
Также обнаружены региональные различия в происхождении последующих неолитических групп. Так последующие неолитические популяции Западной Европы происходят от ранних земледельцев Южной Европы. А в последующие неолитические группы Восточной Европы и Скандинавии внесли свой вклад преимущественно ранние земледельцы Центральной Европы.
А вот с восточной стороны упомянутой пограничной зоны, на территории Прибалтики, Украины и западной части России, не наблюдается существенных генетических изменений, а предковые линии местных охотников и собирателей, без заметного вклада от анатолийских земледельцев,сохраняются примерно до 5 тыс. лет назад. Эта генетическая преемственность согласуется с археологическими данными, которые свидетельствуют о сохранении в этом обширном регионе групп охотников, рыболовов и собирателей, которые использовали керамику, но не практиковали земледелие и животноводство ещё на несколько тысяч лет.
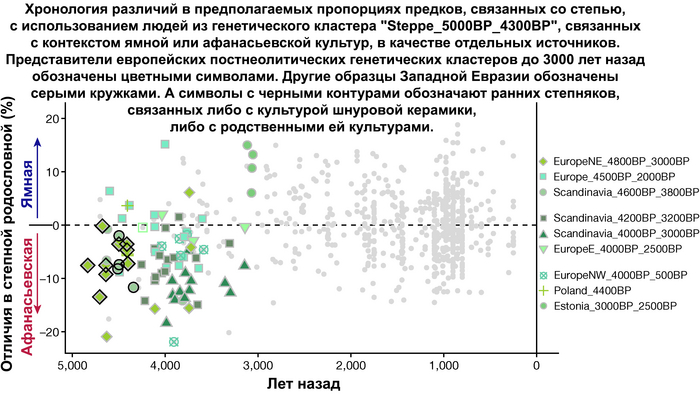
Но несмотря на то, что влияние степной родословной на популяции Евразии хорошо известно, вопросы о её происхождении оставались не решенными. В данной работе авторы показывают, что состав степной родословной (Steppe_5000BP_4300BP) может быть смоделирован как смесь примерно 65% компонент, связанных с охотниками-собирателями, использующими керамику, из района Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет (MiddleDon_7500BP) и 35% компонент, связанных с охотниками и собирателями Кавказа, возрастом от 13 до 10 тыс. лет (Caucasus_13000BP_10000BP).
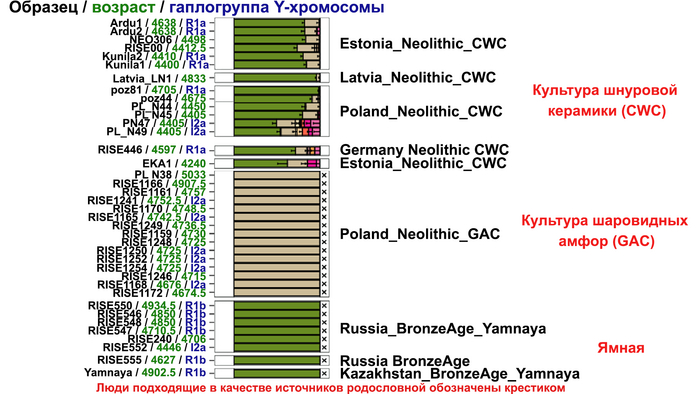
Таким образом, охотники-собиратели Среднего Дона, которые уже имели родословную, связанную с кавказскими охотниками-собирателями, являются неизвестным до сих пор ближайшим так называемым северным источником большей части родословной у людей, связанных с ямной культурой. При этом генетический состав людей из могильника Голубая Криница, отличается от такового у других древних людей из могильников мариупольского типа Приднепровья. Однако распространение степной родословной в Европе связно с культурой шнуровой керамики, носители которой представляют собой смесь предковых линий, связанных со степью и неолитическими земледельцами, происхождение которых может быть связано исключительно с культурой шаровидных амфор.
Это наблюдение имеет большое значение для понимания возникновения культуры шнуровой керамики. Хотя уже давно высказывались предположения о стилистической связи между керамикой культур шаровидных амфор и шнуровой керамики, включая использование сосудов в форме амфор и узоры, выполненные шнуром. При этом незадолго до появления самых ранних групп культуры шнуровой керамики восточные группы культуры шаровидных амфор и западные группы ямной культуры обменялись культурными элементами в лесостепной переходной зоне к северо-западу от Черного моря, где керамические амфоры и кремневые топоры культуры шаровидных амфор были включены в погребения ямной, а типичное для ямной использование охры встречалось в погребениях культуры шаровидных амфор, что указывает на тесное взаимодействие между этими группами. Предыдущие генетические анализы нескольких людей из зоны контакта предполагали, что эти контакты были ограничены культурным влиянием, а не примесью. Однако, в свете новых генетических данных, похоже, что эта зона — и, возможно, другие подобные зоны контакта между культурой шаровидных амфор и группами из степи (как из ямной культуры) — были ключевыми в формировании культуры шнуровой керамики, благодаря которой генетические компоненты, связанные со степью, и культурой шаровидных амфор, распространились далеко на запад и север, что привело к регионально разнообразным ситуациям взаимодействия и смешения.
При этом распространение культуры шнуровой керамики происходило преимущественно благодаря связям, которые были выстроены культурой шаровидных амфор в предшествующий период. Но несмотря на сходство по аутосомным данным, различные линии гаплогрупп Y-хромосомы у представителей культур шнуровой керамики и ямной предполагают, что известные на данный момент образцы представителей ямной культуры не подходят в качестве непосредственного источника степной родословной у носителей культуры шнуровой керамики.
Это предположение было подтверждено моделированием предковых компонент у европейцев со степным происхождением возрастом от 3 тыс. лет назад с участием представителей родственных ямной и афанасьевской культур, потому как модели показывают, что представители афанасьевской культуры, хоть и незначительно, но оказались более лучшим источником степной родословной у европейцев, несмотря на разное географическое распространение.
Стабильность популяций охотников-собирателей к востоку от Урала
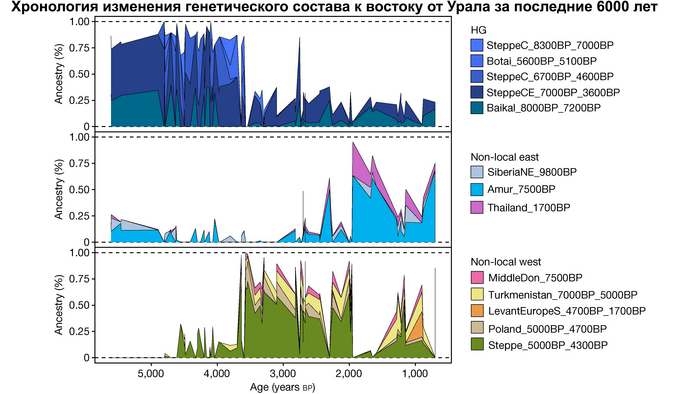
В отличие от значительного числа геномов древних охотников-собирателей из Западной Евразии, которые были изучены до настоящего времени, геномные данные охотников-собирателей к востоку от Урала были скудны. Эти регионы характеризуются ранним появлением керамики из районов, расположенных дальше к востоку, они были населены сложными обществами охотников-собирателей с постоянными, а иногда и укрепленными поселениями. В данной работе сообщается о 38 новых образцах, 28 из которых от охотников-собирателей, использующих керамику 8300-5000 лет назад. Большинство этих геномов попадает на неолитическую степную клину, которая охватывает сибирские лесостепные зоны бассейнов рек Иртыш, Ишим, Обь, Енисей, а также озера Байкал.
Эти охотники и собиратели могут быть смоделированы как смесь компонент из трёх генетических кластеров:
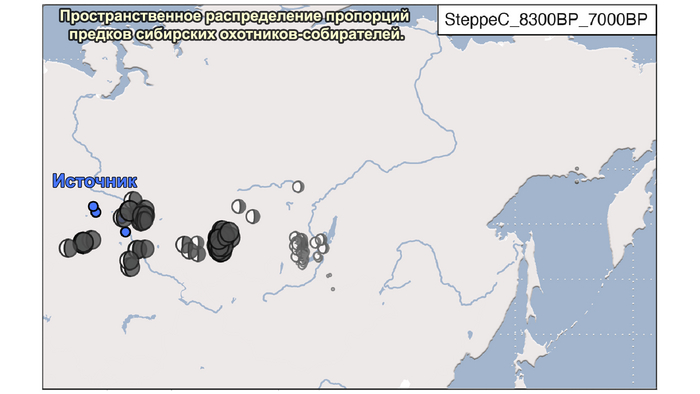
1. Компонента западносибирских охотников-собирателей возрастом от 8300 до 7000 лет (SteppeC_8300BP_7000BP), доминировала в западной части лесостепной зоны.
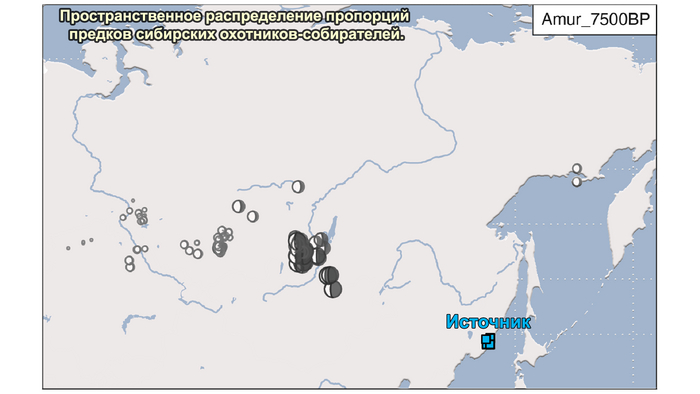
2. Компонента охотников-собирателей Северо-Восточной Азии возрастом около 7500 лет (Amur_7500BP) или древних северо-восточных азиатов из пещеры Чёртовы ворота в Приморье, была максимально выражена у популяций Байкала.
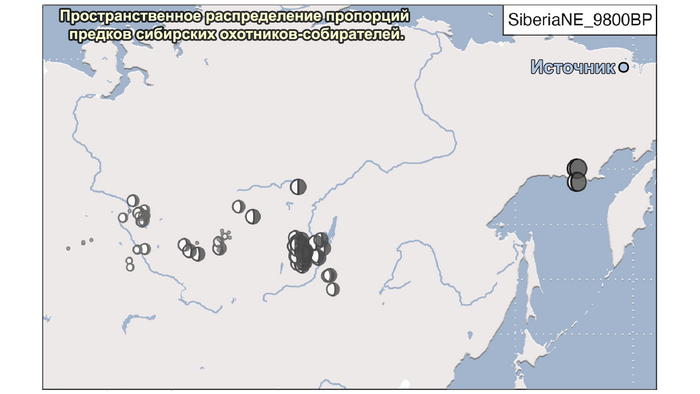
И 3. Компонента представлена палеосибирской родословной возрастом 9800 лет (Siberian_9800BP) как у обитателя Дуванного Яра в низовьях реки Колымы, она снижались по мере удаления от озера Байкал на запад.
1/5
Авторы использовали кластеры также и для исследования пространственно-временной динамики генетического состава в степи и районе озера Байкал после неолита. Результаты подтверждают увеличение компоненты охотников-собирателей лесостепи возрастом от 7000 до 3600 лет (SteppeCE_7000BP_3600BP) у жителей Байкала позднего неолит-ранней бронзы (кластеры Baikal_5600BP_5400BP и Baikal_4800BP_4200BP). Однако родословная, относящаяся к этому кластеру, уже наблюдалась у вновь описанных неолитических охотников-собирателей как у озера Байкал (NEO199 и NEO200), так и вдоль Ангары на севере (NEO843) около 7000 лет назад. У двух мужчин Байкала была определена гаплогруппа Y-хромосомы Q1b1, характерная для последующих групп позднего неолита-ранней бронзы из того же региона. Что совместно с расчётной датой примеси около 7300 лет назад, позволяет предположить, что поток генов между охотниками-собирателями Байкала и лесостепей Южной Сибири уже был во времена раннего неолита, что согласуется и с археологическими данными.
Новые образцы также проливают свет на генетическое происхождение носителей окуневской культуры раннего бронзового века в Минусинской котловине на юге Сибири. Её представителей можно смоделировать как смесь сибирских охотников-собирателей лесостепной зоны из кластеров возрастом 6700-4600 и 7000-3600 лет (Steppe_6700BP_4600BP и SteppeCE_7000BP_3600BP) со степными скотоводами возрастом от 5000 до 4300 лет (Steppe_5000BP_4300BP), связанными исключительно с афанасьевской культурой, время этой примеси датировано около 4600 лет назад.
1/2
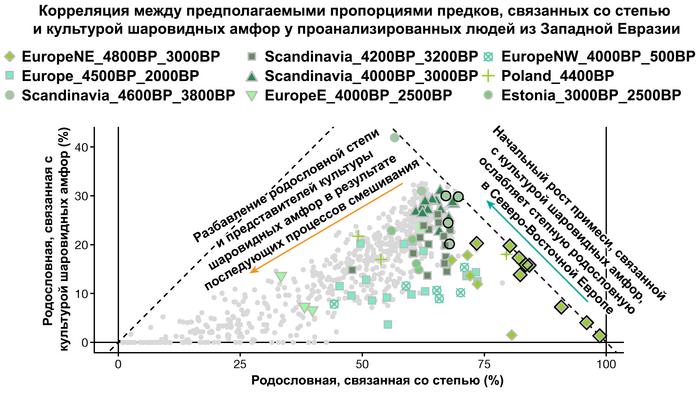
А примерно 3700 лет назад жители в степном регионе и у озера Байкал демонстрируют уже заметно отличающиеся генетические профили. Ранние стадии этого перехода характеризуются притоком предков, связанных со степью, количество которых снижается со временем, от 70% на начальном этапе.
Аналогично динамике в Западной Евразии, происхождение, связанное со степью, здесь соотносится с таковым как у представителей культуры шаровидных амфор с территории современной Польши, возрастом 5000-4700 лет, подтверждая ранее задокументированный поток генов от групп из культуры шаровидных амфор в соседние группы степи и лесостепи, а также поток этой уже смешанной с европейскими земледельцами родословной дальше на восток уже с синташтинской и андроновской культурами в бронзовом веке. Однако у людей из окуневской культуры отсутствует вклад от людей культуры шаровидных амфор, а люди со степным происхождением начиная от 3700 лет назад демонстрируют чуть большую близость к ямной, чем к афанасьевской культуре, что указывает на две волны миграций западных степных скотоводов на восток. Первая волна в раннем бронзовом веке была связана с ямной культурой, а вторая, позже, была связана с синташтинской и андроновской культурами. Более поздние этапы переходного периода характеризуются увеличением родословной, связанной с иранскими земледельцами Центральной Азией (Turkmenistan_7000BP_5000BP) и древними северо-восточными азиатами Приморья (Amur_7500BP).
Суммарно, эти результаты показывают, что на востоке Евразийской степи родословная охотников и собирателей со сложной генетической структурой сохранялась и преобладала значительно дольше, чем в западной Евразии. Пока менее чем 4 тыс. лет назад по степи не прокатились волны миграций, которые привнесли в регион одомашненные породы лошадей с последующим появлением нового снаряжения для верховой езды и колесницы со спицами. Также эти миграции способствовали распространению проса в качестве сельскохозяйственной культуры.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
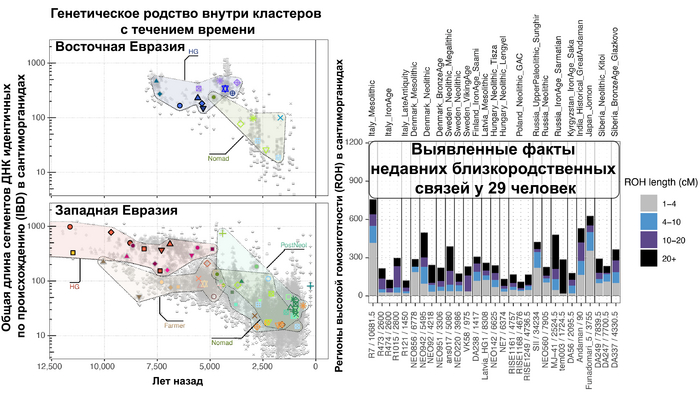
Закономерности распределения сегментов ДНК идентичных по происхождению между людьми, продемонстрировали снижение родства внутри кластеров с течением времени как в западной, так и в восточной частях Евразии. Что согласуется с увеличением эффективного размера популяции в течение этого времени. Однако наблюдаются заметные демографические различия между западной и восточной Евразией. Если на западе во время перехода к неолиту около 9000-6000 лет назад, люди с родословной анатолийских земледельцев демонстрируют снижение доли сегментов ДНК идентичных по происхождению, по сравнению с охотниками-собирателями, то на востоке генетическое родство оставалось высоким примерно до 4000 лет назад, что согласуется с гораздо более длительным существованием небольших локализованных групп охотников-собирателей.
Помимо этого, анализ сегментов гомозиготности (ROH) не выявил значительного числа близкородственных связей, только 29 из 1396. При этом каких-то пространственно-временных или культурных закономерностей в распространенности близкородственных связей обнаружено не было. Зато авторы задокументировали первый случай однородительской дисомии по древнему геному (когда человек получает две копии хромосомы или части хромосомы от одного родителя и ни одной копии от другого родителя). Эта особенность обнаружена у представителя из Темясовских курганов позднесарматской культуры возрастом около 1700 лет.
Также были выявлены люди с различной степенью родства.
Итоги
Появление и сокращение генетического разрыва
После максимума последнего оледенения около 25-19 тыс. лет назад во время расселения древних людей по Евразии, установилось чёткое генетическое разделение между востоком и западом по линии от Черного моря до Балтийского, что отражено и в археологических данных. Это разделение сохранялось на протяжении всего мезолита и неолита. В Восточной Европе распространение земледелия происходило с опозданием примерно на 3000 лет, и эта задержка может быть связана с особенностями окружающей среды, поскольку регионы к востоку от условной границы имеют более континентальный климат и более суровые зимы, возможно, менее подходящие для ближневосточных методов ведения земледелия. На этих территориях сохранились высокоразвитые сообщества охотников-собирателей со стабильными, сложными, а иногда и укрепленными поселениями, с обменом на большие расстояния и крупными кладбищами. Рацион, включающий пресноводную рыбу, зафиксирован как по изотопным данным из этого исследования, так и по анализам остатков липидов в керамике.
В северных лесных районах этой пограничной зоны сообщества охотников-собирателей сохранялись вплоть до появления культуры шнуровой керамики около 5 тыс. лет назад, тогда как в южных и восточных степных регионах охота и собирательство со временем были дополнены животноводством с крупным рогатым скотом и овцами и, возможно, коневодством в Центральной Азии, но этот факт ещё под вопросом. Некоторые из этих групп, такие как представители хвалынской культуры Северного Кавказа и Среднего Поволжья, стали свидетелями появления мужских сообществ, вовлеченных в широкомасштабные обменные связи на основе медных изделий с Восточной и Центральной Европой, а также Кавказом. Поселения в то время были приурочены в основном к равнинным поймам и долинам рек, тогда как степной пояс оставался в значительной степени неиспользованным.
Окончательное исчезновение этой генетической, экономической и социальной границы было вызвано событиями, развернувшимися в степном регионе. Здесь археологически можно наблюдать две временные фазы технологических инноваций: широкое распространение колесных повозок, запряженных волами, около 5500 лет назад и более позднее развитие колесниц со спицами и верховой езды.
Начиная ещё с первой фазы инноваций, в сочетании с возможными изменениями условий окружающей среды, степь стала выгодной экономической зоной, которая позволила группам по типу ямной культуры эксплуатировать степь в качестве кочевых степных скотоводов около 5000 лет назад. А поселения эпохи энеолита вдоль речных долин были заменены этой новой формой мобильной экономики на основе скотоводства, которая окончательно разрушила генетическую границу, существовавшую в предыдущие тысячелетия.
Далее, после второй фазы инноваций около 4 тыс. лет назад, изобретение боевых колесниц и использование проса в качестве продовольственной культуры позволили андроновцам и родственным им группам окончательно распространиться на восток в Среднюю Азию и за ее пределы, оставив глобальное наследие в виде распространения индоиранских языков.
Это исследование внесло свою лепту в наши знания о прошлом, особенно касательно степных миграций.
Во-первых, авторы выявили до сих пор неизвестный источник родословной, который способствовал появлению степных скотоводов, на его роль хорошо подходили охотники-собиратели Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет.
А во-вторых, было определено, что степная родословная, которая распространилась по Европе, благодаря представителям культуры шнуровой керамики, сформировалась в результате культурного и генетического слияния степных групп, родственных ямной культуре и культуре шаровидных амфор. При этом благодаря своему смешанному культурному и генетическому происхождению, носители культуры шнуровой керамики практиковали смешанную экономику, используя различные стратегии выживания в различных условиях. Такая гибкость в значительной степени способствовала их успеху в расселении и адаптации к очень разным экологическим и климатическим условиям за очень короткий период времени.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
Их есть у нас! Красивая карта, целых три уровня и много жителей, которых надо осчастливить быстрым интернетом. Для этого придется немножко подумать, но оно того стоит: ведь тем, кто дойдет до конца, выдадим красивую награду в профиль!
Турки по генам оказались ближе к армянам, чем к азербайджанцам (картинка из ОИ)
Палеогенетические исследования, проведенные учеными на территории современной Турции привели к крайне неожиданным для них результатам. В своем большинстве население Турции имеет 4 основных предковых для них компонентов, которые достались им вовсе не от тюрков, а в основном от индоевропейцев (армян, курдов, греков и славян) и в меньшей степени от арабов. Тюркские же наследственные компоненты в структуре жителей Турции не превышают всего на всего 6-10%. Так "Фонд доноров костного мозга" выявил высокое сходство генетических показателей армян и турок. "Армяне и турки генетически близки друг к другу" - пишут руководители этого Фонда. В настоящее время многие турки, прошедшие ДНК-тесты, отказываются верить, что у них явно преобладают армянские, итальянские, славянские или греческие корни.
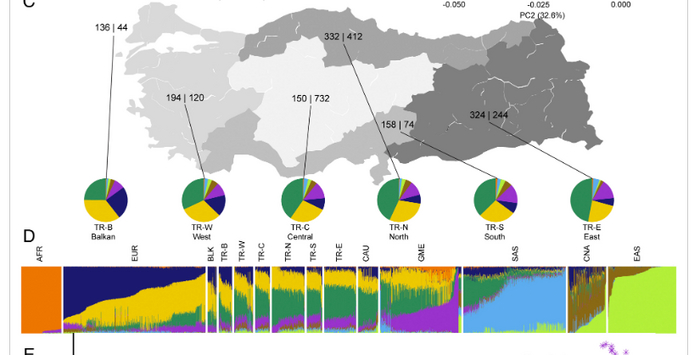
В конце 2021 года в научном журнале PNAS вышло новое подробное генетическое исследование населения Турции, в котором было выявлено, что турки имеют четыре основных предковых компонентов в их геноме: европейский (синий), балканский (желтый), армянский (зеленый) и еще ближневосточный (фиолетовый). Все остальные предковые компоненты: африканский, восточноазиатский, среднеазиатский и южно-азиатский у турков представлены в незначительном количестве. Было установлено, что все предковые компоненты неодинаково распространены в самых разных регионах Турции и довольно сильно коррелируют с проживанием в этих местах более древних групп населения, которые там жили до завоевания Анатолии тюрками-сельджуками. Например, в TR-B (балканский кластер) представлено около 80% европейского, балкано-славянского и кавказского (армянского) предковых компонентов. Примерно такое же соотношение
Районы Турции, имеющие различия по предковым компонентам (M Ece Kars et al., 2021)
отмечено в Западном кластере (TR-W) с некоторым увеличением доли армянского и уменьшением европейского и балкано-славянского ДНК компонентов. Еще больше доля армянского предкового компонента наблюдается в северных, центральных и восточных регионах Турции, в которых он достигает 50%. Суммарная же доля восточноазиатского и среднеазиатского предковых компонентов нигде не превышет 10%, а меньше всего этого предкового компонента на востоке TR-E (менее 7%) и юге Турции TR-S, где в основном проживают курды. Зато в этих районах увеличивается доля ближневосточного предкового компонента (цвет фиолетовый).
Как такое могло произойти?
Оказывается все очень просто и банально. Один из авторов книги "Внуки" Фетхие Четин, которую она написала совместно с академиком Айше Гюль Алтынай отмечает, что в Турции сейчас проживает огромное количество "скрытых" армян, которые даже не догадываются о своем происхождении из-за того что их пращуры в начале XX века испытали мощное давление и в дальнейшем просто скрывали свое происхождение даже от своих детей. Многие современные турки после прохождения тестов ДНК оказываются в сильном шоке и отказываются верить в то, что они не имеют даже капли тюркской крови. Примерно такая же ситуация связана и с некоторыми из других народов, которые издревле населяли территорию Анатолии или же окрестности Константинополя (Стамбула) - греками, славянами и курдами.
Почему турки по ДНК ближе к армянам, чем к азербайджанцам?
Как показали последние исследования азербайджанских палеогенетиков (Алиев, Ибрагимов, Халилова и др., 2018) современные азербайджанцы несут преимущественно ближневосточные предковые компоненты (55%) и в меньшей степени другие (20% - европейские, балканские и армянские, 18% - центральноазиатские и восточноазиатские, 6% - южноазиатские). У турков же соотношение этих предковых компонентов другое (около 70% - европейские, балкано-славянские и армянские), 20% - ближневосточные и 10% - восточноазиатские и южноазиатские), что сходно с предковыми компонентами большинства населения Армении.
Таким образом, турки имеют большое сходство с азербайджанцами по языку и культуре, немного меньшее по религии (проповедуют ислам, но азербайджанцы - шииты, а турки - сунниты), а вот по происхождению и генам турки имеют большее сходство с армянами, чем азербайджанцами.
Армянка и турчанка (картинка из ОИ)
Если Вам понравилась наша статья не забывайте ставить лайк (👍 - палец вверх), комментируйте ее и подписывайтесь на наш канал.
История происхождения славян была запрещена Ватиканом
Первое наиболее объемное исследование по истории славян, написанное итальянским священником (хорватом по происхождению) Мавро Орбини "Славянское царство", вскоре после его публикации в 1601 году было запрещено Ватиканом и внесено в папский «Индекс запрещенных книг». Этой книгой очень интересовался российский император Петр I.
В 1846 году польский археолог Тадеуш Воланский издал книгу "Письма о славянских древностях". Разгневанное католическое духовенство вскоре вознамерилось расправиться с археологом крайне жестко — сжечь его на костре из собственных книг. Поляка спас русский император Николай I, который не позволил разделаться с ним и повелел охранять археолога.
Мавро Орбини (1563 - 1610 гг) и обложка его книги "Славянское царство"
Почему католическая церковь запретила эти труды ученых?
Для того, чтобы это понять обратимся к истокам католической церкви. Как известно из истории, в 1054 году произошел церковный раскол, после которого случилось окончательное разделение христианской Церкви на Римско-католическую церковь на Западе, с центром в Риме, и церковь Православную на Востоке, с центром в Константинополе. Обе церкви предали друг друга анафеме. Византия подтвердила свою самостоятельную от Запада цивилизацию, но лишилась политической поддержки Рима и в целом всего Запада. Напомню, что католическая церковь была духовным рупором средневековой Священной Римской Империи, которая тогда представляла союз разных западноевропейских государств и народов (германских, франкских, итальянских и др.), который существовал с 962 по 1806 годы. Элита и знать этого негосударственного объединения считала себя потомками Великой Римской Империи, а их католическая церковь во главе с Папой всячески поддерживала данную идеологию. В реальности западные европейцы были потомками тех самых варваров с которыми Древний Рим постоянно вел войны и в конце концов был ими разрушен. С начала XI века западноевропейские завоеватели стали нападать и на территории, которые тогда находились под контролем Восточной Римской империи (Византии). Это еще более усилило конфликты и противостояние Западной (католической) и Восточной (православной) церквей, которое не преодолено и до настоящего времени.
Что же такого крамольного было написано в этих книгах, которые так сильно напугали католическую церковь?
Во-первых, оба автора проанализировав все исторические документы, которые были известны на тот момент времени пришли к выводу, что славяне имели очень древнее происхождение, а их влияния и широкое расселение по всей Европе замалчивалось и всячески скрывалось. Во-вторых, оба автора указывали на то, что предки славян имели отношение к образованию Древнего Рима. Тадеуш Воланский напрямую высказывался о том, что этруски не только являлись предками славян, но именно они и были первыми основателями Рима. Мавро Орбини писал о том, что предки славян в древности покорили Грецию, Македонию, Иллирию, Италию, Испанию и даже Египет.
Почему гипотеза о происхождении славян была запрещена Ватиканом?
Так почему же католическая церковь запретила эти книги?
Все очень просто, эти книги, по мнению Римской-католической церкви, выдвигали гипотезу, которая напрямую противопоставлялась принятому на Западе мнению, что западно-европейцы являлись законными потомками древних римлян. Из этих книг могло сложиться мнение, что, наоборот, скорее славяне вместе с Православной церковью являются наследниками Великой Римской Империи.
Но так ли это или нет на самом деле?
1) Согласно современным данным палеогенетиков завершение эпохи бронзы совпало с общемировой войной (нашествием народов моря) или катастрофой бронзового века. После этого мир в Европе разделился на три основных лагеря: 1. Западная Европа - носители в основном гаплогрупп R1b-L151, R1b-P312, R1b-U106 и I2a-Isles (по аутосомным компонентам в основном - WHG и CHG), 2. Восточная Европа - носители гаплогрупп R1a-Z645 (все ее ветви), I2a-Din и N1a (по аутосомам в основном EHG и SHG), 3. Южная Европа (позже Древний Рим) - носители самых разных гаплогрупп - I2a (разные ветви), G2a, J1, J2, E1b, R1b-Z2103, R1a и других (по аутосомным анализам в основном EF - это потомки неолитических земледельцев и в меньшей степени народы с другими предковыми компонентами).
2) Как показали последние исследования различных ученых, с которыми вы можете познакомиться в моих предыдущих статьях (перечень смотрите в ссылках ниже), славяне по своему генетическому происхождению являются смесью потомков восточно-европейцев и южно-европейцев. То есть именно славяне, а не представители современной Западной Европы, в большей степени являются наследниками или потомками населения Древнего Рима, что подтверждает предположения указанных выше авторов и причину противодействия им со стороны Ватикана.
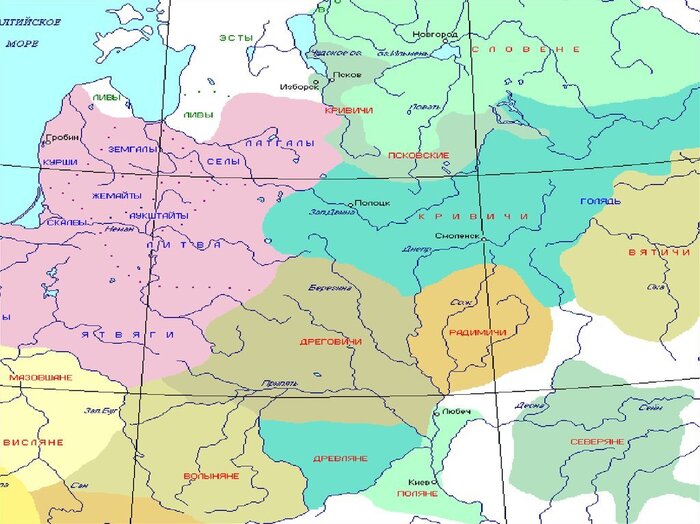
Из истории известно, что самое крупное объединение восточно-славянских племён в VI-XII веках нашей эры располагалось в центральной части Русской равнины (от Смоленска и верховьев рек западной Двины и Днепра на западе до рек Волги и Оки на востоке, Пскова и Новгородчины - на севере). До сих пор нет среди ученых единого мнения откуда пришли эти люди - то ли с севера на юг, то ли с юга на север. А звался этот загадочный народ - кривичи. По мнению одних ученых - Владимира Топорова и академика Андрея Зализняка "кривичи" пришли с севера Польши, так как их язык был близок к языкам лютичей и невров. Историк Нина Васильева считает их автохтонами Русской равнины, которую кривичи заселили еще в бронзовом веке, а историк Сергей Алексеев считает, что кривичи пришли с юга, а на север их вытеснили гунны на рубеже IV–V веков нашей эры. Другие ученые вообще не относят кривичей к славянам, а видят в них потомков балтов, финнов, скандинавов или германцев.
Распространение племен кривичей в конце первого тысячелетия нашей эры (картинка из ОИ)
Много легендарных событий связано с этим народом. Это был один из самых могущественных и удивительных племенных союзов на Русской ранине.
Очень необычным является и само название "кривичи". Существует не менее пяти версий его происхождения: 1) от имени первого вождя Кривъ или Криве, 2) от извилистой (неровной, кривой) местности, на которой жил этот народ, 3) от слова кровные - близкие по крови, 4) от одноглазого (кривого) скандинаво - германского бога Одина, потомками которого могли быть кривичи, 5) от литовского слово kirvis, которое обозначает боевой топор или секиру, что возможно указывает на культуру "боевых топоров" бронзового века. Имеются и некоторые другие версии происхождения славянского племени кривичи.
Интересно, что с этим словом у некоторых народов восточной Европы до сих ассоциируются русские. Так по литовски русские - это kriẽvai, а по латышски - krievs, Россия - Krievija. Слова с корнем krȋv, kríva (неправильный, необычный, неверный в отношении веры, кривой, странный) есть и в славянских и финских языках. Что же такого неправильного, необычного или кривого было в истории этого древнего племени?
Историки и археологи пришли к выводу, что кривичи не были чисто славянским племенем, а их могучий союз образовался, вероятно, в процессе смешения и взаимной ассимиляции нескольких разных народов - балтов, финнов, славян и даже скандинавских и восточно-германских племен. На это указывают и разные археологические находки этнических артефактов, найденные в их захоронениях. Важно также отметить, что захоронения кривичей отличались от их ближайших соседей (других славян, балтов, финнов или скандинавов). Отличительной чертой погребений кривичей являлись их длинные курганы — валообразные земляные насыпи. Все длинные курганы содержат захоронения по обряду трупосожжения.
Длинные курганы кривичей (картинка из ОИ)
Кривичи были одним из самых крупных восточнославянских племенных союзов и, по-видимому, по мере своего постепенного расселения на восток кривичи ассимилировали существенную часть древне балтского, а также часть финно-угорского населения. Кривичи делились на несколько племен, которые заселяли разные районы Русской равнины - на севере Беларуси, в Смоленской области и в соседних с ними районах Подвинья и Поднепровья жили полоцко-смоленские кривичи, на севере - на Псковщине и в западной части Новгородчины были распространены племена псковских кривичей, а в верховьях Волги и Оки, в современных Владимирской, Тверской, Московской, Рязанской, Нижегородской и Ярославской областях - восточные кривичи. Кроме этого племена кривичей упоминаются, как одно из славянских племён на полуострове Пелопоннес, в греческом сочинении «Хроника Мореи». На этот же факт указывает и известный российский лингвист О. Н. Трубачёв.
По антропологическим данным для кривичей был характерен высокий рост, долихокефалия (длинное и узкое лицо), крупный выступающий волнистый нос, очерченный подбородок, в основном русые волосы, которые характерны для северного валдайского типа. В. В. Бунак в 1932 году пришёл к выводу о сходстве кривичей с алеманнами (древнегерманский племенной союз). В захоронениях (длинных курганах) этого племенного союза обнаружены разные генетические маркеры (Y-гаплогруппы) по отцовской линии: N1a1a, R1a, I2a, I1, E1b1b и J2a1, что подтверждает многонациональное происхождение этого могущественного племенного союза. В отцовской линии R1a у кривичей широко распространен был субклад R1a-CTS3402, который определяется у многих носителей на Русской равнине. Прародина этого субклада, скорее всего находится где-то в Карпатах и приходится на эпоху Бронзы. В железном веке к началу нашей эры его носители уже широко заселяли многие территории Русской равнины. С возникновением восточнославянских языков в современной их форме в V-VI веках н.э. большая группа носителей CTS-3402 стала их использовать. Другие же носители этой Y-гаплогруппы влились в балто-финские или окско-волжские группы, а также в северо- и восточногерманские племена (субклад встречается в захоронениях викингов в Salme, на Готланде и в ряде других мест). Типичные представители субклада R1a-CTS3402 до V века могли говорить на днепровско-окском языке. Но какие-то ветви перешли на германские языки, а другие - на волго-окские языки и вошли в состав эрзи, мокши, мери и муромы. Недавние исследования европейских и российских ученых РАН принесли сенсацию. Древняя ДНК с археологического памятника Пужалова гора возле г. Гороховец на Клязьме IX века дала неожиданную для вымершего народа мери ветвь - R1a-M458-L1029. Этот субклад традиционно ассоциируется с Центральной Европой.
Кривичи - реконструкция учёного-антрополога, доктор исторических наук М. М. Герасимова
После образования Древней Руси кривичи вместе с вятичами приняли активное участие в славянской колонизации Волго-Окского междуречья (современные Владимирская, Костромская, Рязанская, Ярославская, Нижегородская, Тверская, Московская, а также Вологодская области), где, вероятно, ассимилировали или оттеснили местные финские племена дьяковской культуры.
В Ипатьевской летописи кривичи упомянуты в последний раз под 1128 годом, а полоцкие князья названы кривичскими под 1140 и 1162 гг. После этого кривичи больше не упоминаются в русских летописях. Племя кривичей довольно часто связывалось с варягами и долгое время употреблялось в различных зарубежных источниках вплоть до самого конца XVII века. Кривичские князья оставили след в генеалогическом древе первых руских князей Рюриковичей и связаны с ними по материнской линии через сына Рогнеды (кривичская княжна) - Ярославом Мудрым, будущим великим князем Древней Руси.
Если Вам понравилась статья подписывайтесь на канал, делитесь ею в социальных сетях и ставьте лайк (👍 - палец вверх)
Исследования ДНК жителей древней Ярославии удивили ученых (картинка из ОИ)
Город Ярославль и Верхнее Поволжье исторически было зоной контактов самых разных этнических групп населения: славян, балтов, финно-угров и скандинавов. По летописям в X-XI веков здесь проживал ныне вымерший народ мери. Сюда мигрировали прибалтийские финны (предки карел и вепсов), а в конце первого тысячелетия нашей эры в этот регион проникли славяне, причем эта миграция происходила из двух различных потоков: ростово-суздальского («низового») и новгородского («верхового»). Поэтому вполне предсказуемо среди жителей древней Ярославии могли быть представители всех этих этнических групп.
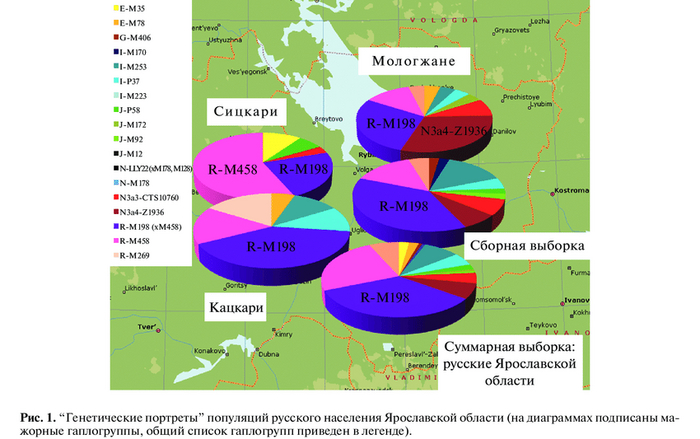
Современные исследования разных авторов подтверждают это предположение (смотри рис.1 и рис.2 ниже). Исследования ученых РАН (М.Чухряева и др., 2017) показали, что в этом регионе явно преобладало население с отцовской Y-DNK гаплогруппой R1a (более 50%) и в меньшей степени были представлены другие отцовские Y-DNK гаплогуппы - N, I, J, E и R1b (смотрите ниже рисунок №1).
Более поздние генетические исследования из захоронений населения города Ярославля периода его штурма ханом Батыем в 1238 году подтвердили в целом предыдущие исследования с несколько увеличиной пропорцией в части носителей Y-ДНК I1 и R1b, которые могли быть от скандинавов (рис.2). Однако, новые данные вполне объяснимы с исторической точки зрения, так как тогдашние дружинники русских князей, среди которых было немало скандинавов, жили в основном в городах. Были определены мажорные (R1a, R1b, I1, N3)и минорные(E, I2, G2a, J2b, J1c3) гаплогруппы Y-хромосомы населения Древнего Ярославля.
Рис. 2. Генетический портрет жителей древнего Ярославля, убитых во время штурма 1238 года.
С развитием генетики, интерес вызывают не только вопросы о происхождении популяций, но также и отдельных людей, особенно если они принадлежали знатному роду. В частности, вопросы о происхождении элиты средневековой Руси IX–XVI вв., где важнейшими действующими лицами были представители правящего рода Рюриковичей, долгое время были предметом дискуссий. Ранее отцовские линии в виде гаплогрупп Y-хромосомы Рюриковичей были реконструированы по биологическим материалам современных людей, чьё происхождение, c разной степенью достоверности восходит к Рюрику. А что касается исследований древних геномов, то их осложняют трудности в идентификации останков Рюриковичей и другой знати. Поскольку до начала XV в. имена погребенных отсутствовали на погребальных сооружениях, надгробиях и саркофагах. Места погребений той эпохи устанавливаются по летописным сообщениям и спискам имён для поминовения XVI–XVII вв. Помимо этого практики повторного использования старых погребальных сооружений, перемещение почитаемых останков и их изъятие в ходе антирелигиозной кампании в советское время, привели к тому, что значительная часть останков XI–XIV вв. из погребений в древнерусских храмах и не только, оказалась утрачена или не может быть достоверно связана с определенными историческими личностями.
1/2
Рака с мощами св. Александра Невского
К примеру, в 1967 году на территории Черниговского Спасо-Преображенского собора во время строительных работ был случайно обнаружен череп мужчины 25-35 лет со следами ран от меча, без археологического документирования. Останки были предположительно отнесены одному из внуков Ярослава Мудрого - Глебу Святославичу, князю тмутараканскому и новгородскому, жившему в 11 веке. Однако точно определить кто был похоронен в каменном саркофаге возле Черниговского Спасо-Преображенского собора не представляется возможным. Особенно в свете недавнего анализ ДНК этого человека, который показал, что он был носителем гаплогруппы Y-хромосомы I2a1a2b1a1a и обладателем митохондриальной линии H5a2a.
Датировки (только археологические): Саркофаг найден на заднем дворе собора. Глубина верхней плиты саркофага составляла 1,9-2,0 м от современной поверхности. Стратиграфия несколько пострадала от более позднего нарушения грунта: трубы водоснабжения проложены на высоте 0,2 м над изголовьем саркофага. Археологи относят его к XI веку нашей эры. гаплогруппа – Y-хромосомы I2a1a2b1a1a гаплогруппа мтДНК – H5a2a
Поэтому особую ценность представляют те немногочисленные захоронения, где костные останки по археологическому контексту, антропологическим данным и комплексу исторических свидетельств надёжно отнесены к представителям рода Рюриковичей.
К таким достоверным захоронениям относится погребение в Спасо-Преображенском соборе Переславля-Залесского князя Дмитрия Александровича, второго сына Александра Ярославича Невского. Прямые сообщения о погребении Дмитрия Александровича в Спасо-Преображенском соборе содержит ряд летописей, в том числе Новгородская четвёртая, Воскресенская, Никоновская летописи, а также Московский великокняжеский свод конца XV века.
А поиск места погребения был облегчен ещё и тем фактом, что после захоронения князя Дмитрия Александровича в 1294 году и его наследника бездетного старшего сына Ивана в 1302 году с последующим переходом Переяславского княжества Москве, этот храм как престижная усыпальница использовался реже чем многие другие в средневековье. Изучив все доступные документы, можно было полагать, что погребение Ивана Дмитриевича находилось в юго-западной части собора, под хорами, а погребение князя Дмитрия Александровича – в юго-восточной части, в южной алтарной апсиде, где до реставрации 1891 г. находились отмечавшие места погребений более поздние кирпичные надгробия XVII в. В результате сам саркофаг с останками, был обнаружен отрядом Института археологии РАН под руководством Владимира Валентиновича Седова в 2014 г.
1/2
Князь Дмитрий Александрович унаследовавший после смерти великого князя Александра Ярославича Невского – Переславское княжество, в разные годы также был новгородским князем и великим князем владимирским. Князь умер в 1294 году на Ламском Волоке, возвращаясь в Переславль из Твери, и был похоронен в Спасо-Преображенском соборе Переславля.
Результаты
Палеогенетический анализ
В результате полногеномного секвенирования фрагментов древней ДНК из наиболее информативного образца - ладьевидной кости стопы Nev2.3, сгенерировано более 15 млн чтений, определены 532154 однонуклеотидных полиморфизма (ОНП). В процессе обработки данных секвенирования установлено, что это был мужчина с митохондриальной гаплогруппой F1b1 и гаплогруппой Y хромосомы N1a1a1a1a1a1a7a~ N-Y59140.
1/2
Анализ Y-хромосомы князя Дмитрия Александровича и других предполагаемых представителей рода Рюриковичей
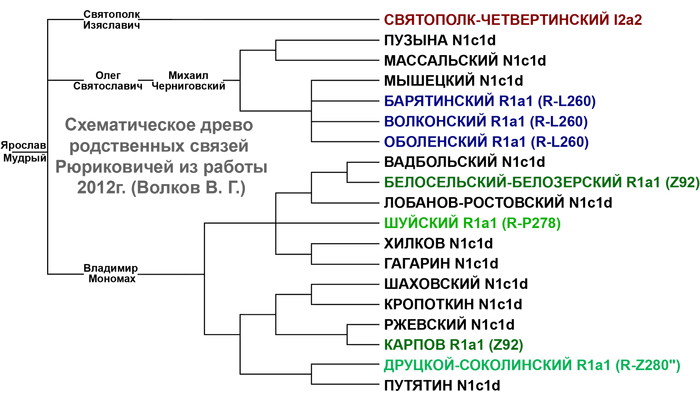
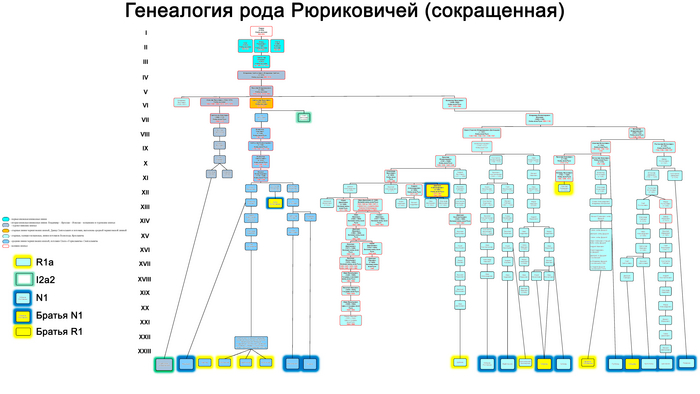
На основании результатов генетических исследований трех древних потомков Рюрика и предполагаемых 43 современных Рюриковичей выдвинуты гипотезы о трех возможных отцовских линиях Рюрика, а именно N1a, R1a и I2a.
В результате филогенетического позиционирования Y хромосома князя Дмитрия Александровича кластеризовалась с Y хромосомами современных вероятных потомков Рюриковичей, принадлежащих к гаплогруппе N1a, которые произошли из различных аристократических родов: Мстиславичей и Юрьевичей, потомков Мономаха от великого князя Мстислава Владимировича и великого князя Киевского Юрия Долгорукого, а также Ольговичей, черниговских Рюриковичей, потомков князя Олега Святославича или «Гориславича».
1/2
При этом важно отметить, что вероятные захоронения князя Глеба Святославича Черниговского, из рода Ольговичей, с линией I2a и Изяслава Ингваревича Луцкого, из рода Мстиславичей, с линией R1a, не подтверждены археологическими данными, что ставит под сомнение и гипотезы, которые вытекают из генетического анализа указанных образцов.
В целом как древние, так и современные предполагаемые Рюриковичи с линиями гаплогрупп R1a и I2a, обладают высокой гетерогенностью нуклеотидной последовательности Y хромосом и не указывают на единое происхождение по прямой мужской линии. В то время как большинство ныне живущих потомков Рюрика по прямой мужской линии согласно их родословным, с гаплогруппой N1a, обладают максимально похожими вариантами Y хромосомы как между собой, так и с Y хромосомой князя Дмитрия Александровича. Что совместно с полногеномными данными средневековых и современных Рюриковичей указывает на то, что, как минимум, со времени великого князя Ярослава Мудрого (XI в), мужчины этого рода были носителями гаплогруппы Y-хромосомы N1a.
Поиск живших в прошлом людей наиболее близких к князю Дмитрию Александровичу по Y-хромосоме
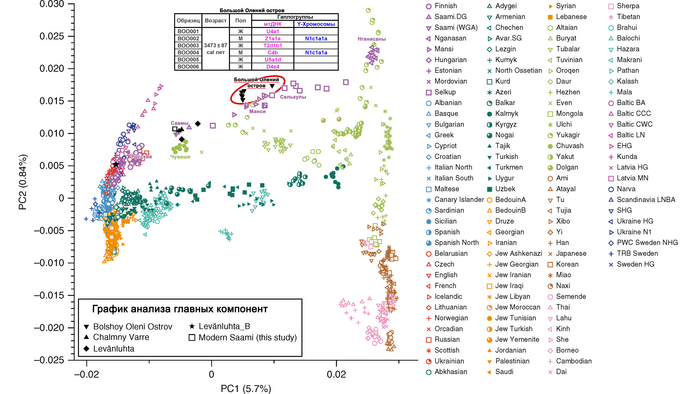
На филогенетическом дереве Y хромосомы видно, что помимо современных Рюриковичей, линия Дмитрия Александровича группируется с древними людьми середины II тысячелетия до н.э. Оленеостровского могильника в Кольском заливе Баренцева моря.
В предыдущих работах на примере этих образцов был показан поток генов из Сибири на север и восток Европы (восточно-евразийская компонента).
1/3
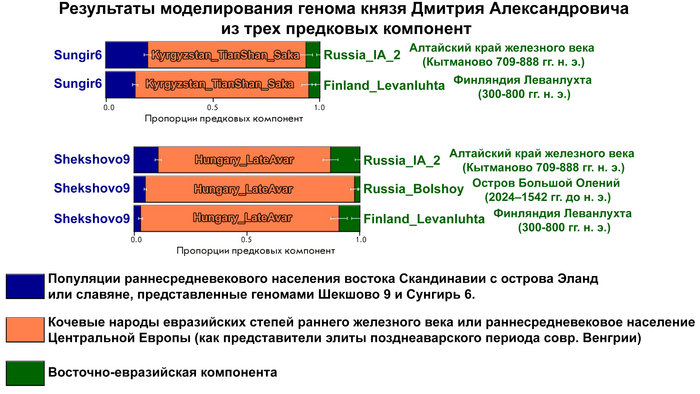
Высокая степень биологического соответствия Y-хромосомы представителя русского княжеского рода и людей эпохи раннего металла привела авторов работы к гипотезе о возможном вкладе восточно-евразийского генофонда в формирование североевропейского населения раннего средневековья, поскольку, согласно летописям, Рюриковичи имели с ними генетическую связь.
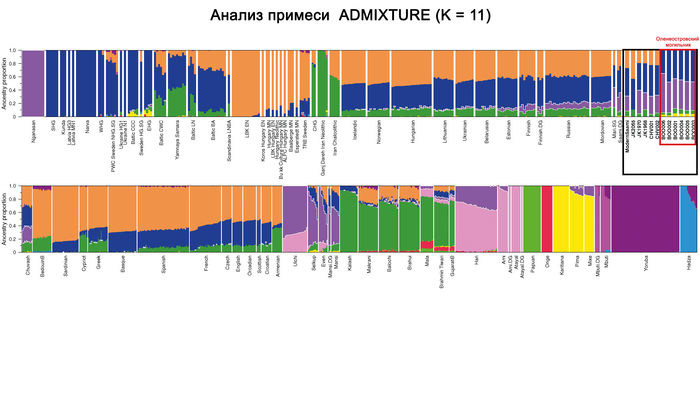
Авторы работы исследовали вклад людей с Большого Оленьего острова в формирование средневекового населения, жившего вдоль побережья Балтийского моря из современных Финляндии, Дании, Швеции и Норвегии, используя геномы периода «викингов» из работы 2020 года, обзор которой тоже есть на канале. В статистических анализах удалось выявить существенный вклад геномов древних представителей Большого Оленьего острова в формирование генофонда популяций периода «викингов», скорее всего, благодаря контактам последних с финно-угорским населением прибалтийского региона. При этом у представителей периода «викингов» с гаплогруппой N1a этот вклад выше чем у носителей линий R1a. Результаты показывают, что в формировании генофонда отдельных жителей из Швеции, Норвегии и Балтийского региона участвовали и люди из Сибири с восточноазиатскими корнями. При этом в наиболее подходящих моделях, помимо европейских охотников-собирателей, неолитических земледельцев и степных скотоводов, скандинавские популяции моделировались с участием обитателей Большого Оленьего острова или представителей хунну железного века.
Здесь стоит адекватно воспринимать компоненты в моделях. В данном случае хунну добавлены для проверки географического происхождения компоненты из Сибири, если говорить упрощённо.
А что касается предков князя Дмитрия Александровича по мужской линии, то с большой вероятностью, среди них были мужчины, оставившие могильник на Большом Оленьем острове в Кольском заливе Баренцева моря около 3600 лет назад.
Анализ мтДНК князя Дмитрия Александровича
Митохондриальную гаплогруппу F1b1, определенную у князя Дмитрия Александровича относят к восточно-евразийскому кластеру, она встречается с разной частотой у большинства исследованных древних и современных популяций Байкальского региона и сопредельных территорий Центральной Азии.
Погребения мужчин с митохондриальными линиями F1b1b и F1b1f, а также с линией Y-хромосомы N1a1a1a1a3a (F4205) характеризовались довольно богатым погребальным инвентарем с золотыми и позолоченными предметами, что указывает на их принадлежность к знати.
● A1811 – Женщина, захороненная с костями овец или коз. мтДНК F1b1f ● A1812 – Мужчина из богато обставленной могилы, с золотыми серьгами, золотыми заколками для волос, ремешками из золотой фольги с переплетающимися узорами из серебра и мечом, украшенным золотыми пластинами. мтДНК – F1b1f, Y-хромосомы – N1a1a1a1a3a (F4205) ● A1821 – Пожилой мужчина, захороненный с исключительно богатыми погребальным инвентарём, включающим позолоченный меч и пояс. мтДНК – F1b1b, N1a1a1a1a3a (F4205)
Однако с уверенностью говорить о происхождении материнской линии князя Дмитрия Александровича довольно сложно, так как практически во всех исторических периодах прослеживается повышенная изменчивость и разнообразие митохондриальных гаплогрупп в одном регионе.
Здесь также важно понимать разницу между генофондом популяций и отдельно взятых людей, особенно представителей знатных родов, у которых была широко распространена практика привлечения самых разных по своему происхождению женщин для союзов, официального и неофициального характера, в качестве выгодных или вынужденных политических шагов. Поэтому материнская линия F1b князя Дмитрия Александровича могла быть связана как с древними миграциями из Сибири, так и с миграциями кочевников раннего средневековья. При этом у этих разных по времени групп могло быть общее происхождение этих линий.
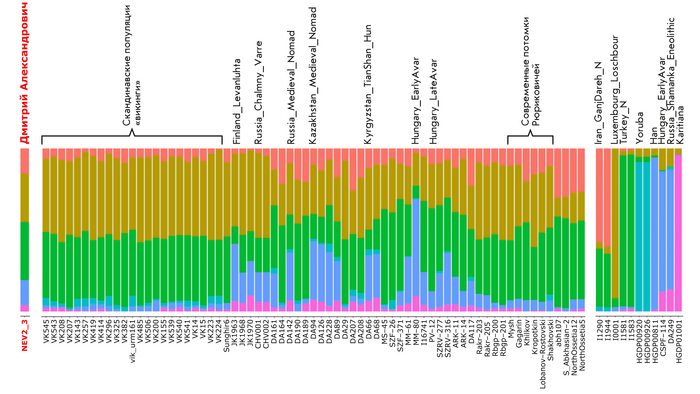
Результаты анализа главных компонент
Генетическую близость генома князя Дмитрия Александровича к другим известным древним и современным популяциям оценили с помощью метода главных компонент (PCA).
Сразу стоит отметить, что надо адекватно понимать положение на этом графике отдельных представителей популяций, особенно из знатного рода. Если положение большинства образцов сочетается с их географией, то положение отдельно взятых людей от смешанных союзов будет где-то между положением его родителей. При этом генетическая схожесть в этом анализе не подразумевает общую историю, а лишь схожие пропорции генетических компонент.
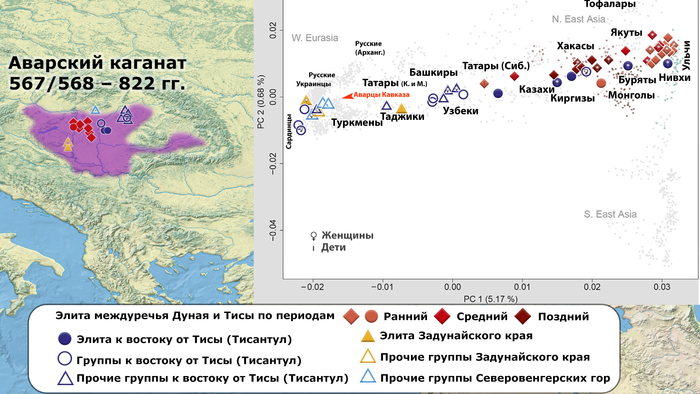
В нашем случае геном князя Дмитрия Александровича занимает промежуточное положение между древними и современными образцами из Европы и Центральной Азии. Наиболее близкими генетически и хронологически к князю Дмитрию Александровичу, расположены некоторые представители авар позднеаварского периода из междуречья Дуная и Тисы, а также из предполагаемого вторичного центра власти Аварского каганата к востоку от Тисы.
Здесь стоит напомнить, что в Аварском каганате (VI–IX вв.), помимо европеоидного большинства ярко выраженный монголоидный тип, имели, преимущественно, представители элиты, который размывался со временем к позднеаварскому периоду.
Как известно из работы 2022 года, популяции аварского периода демонстрируют значительную генетическую изменчивость, а их образцы разбросаны на графике анализа главных компонент от популяций Западной Евразии до популяций Северо-Восточной Азии.
Однако несмотря на эту общую неоднородность, существуют четкие закономерности генетической подструктуры, соответствующие как хронологии и географии Карпатского бассейна, так и социально-археологическим категориям. Почти все представители элиты раннеаварского периода образуют плотный кластер с высоким содержанием компоненты «древних северо-восточных азиатов» (АНА) до 98%. Однако образцы представителей элиты позднеаварского периода уже сильнее смещены в сторону Западной Евразии, а количество родословной «древних северо-восточных азиатов» у них сократилось уже до 70-80%.
При этом среди простых жителей раннего и позднего аварского периодов преобладал более западный генетический профиль, демонстрирующий сильную связь с населением Карпатского бассейна доаварского периода, с нулевым или небольшим вкладом компоненты «древних северо-восточных азиатов». И как раз именно к этой группе образцов относится расположенный рядом с князем Дмитрием Александровичем представитель позднеаварского периода со смешанным генетическим профилем (Hungary_LateAvar), примерно, на 80% состоящим из компоненты максимально выраженной у предшествующих местных жителей Карпатского бассейна и на 20% из компоненты «древних северо-восточных азиатов» из восточной степи.
Анализ примесей (Admixture)
В анализе примесей, по предковым компонентам отмечено генетическое сходство князя Дмитрия Александровича с представителями раннесредневекового населения востока Скандинавии периода викингов.
Что может свидетельствовать в пользу версии о «варяжском» (скандинавском) происхождении Рюрика, родоначальника княжеского рода, призванного на Русь, о чем прямо говорит Летопись. При этом авторы, используя термин «викинги», подчёркивают, что это неоднородная и сложная по своему историческому формированию европейская популяция, объединенная лишь образом жизни и ареалом проживания. Однако есть и отличие генома князя от скандинавских популяций эпохи «викингов», в том числе с территории современной России, это отличие связано с наличием у него дополнительной восточно-евразийской компоненты (обозначенной синим цветом на графике), отсутствующей в таком количестве у подавляющего большинства «викингов». Эта компонента максимально выражена у нганасан, финно-угорского народа Западной Сибири манси, представителей народа хань в Китае, у элиты авар из междуречья Дуная и Тисы (Hungary_DanubeTisza_MLAvar), а также у более древних представителей эпохи раннего неолита Байкала и Монголии (Russia_Shamanka_Eneolithic. SG и Mongolia_North_N). В меньшей степени эта компонента обнаружена у жителей раннего средневековья с территории современной Финляндии (Finland_Levanluhta), у кочевника Прикаспийской степи (Russia_Medieval_Nomad), современника князя Дмитрия Александровича, а также у более древних ираноязычных степных кочевников железного века с территорий современных Казахстана и Кыргызстана (Kazakhstan_TianShan_Saka, Kyrgyzstan_TianShan_Hun). Суммарно, на основании данных анализа главных компонент, анализа примесей и информации о митохондриальной ДНК можно утверждать, что в геноме князя Дмитрия Александровича имеется существенный вклад восточной предковой компоненты, что отличает его от раннесредневекового населения востока Скандинавии, «викингов» и средневекового славянского образца из Владимира (Сунгирь6), но сближает в первую очередь с древним населением Финляндии, Кольского полуострова и раннесредневековым населением Центральной Европы с этой компонентой. Вероятно, этот вклад получен и по мужской, и по женской линиям, что соответствует процессам древней миграции из Сибири по степному евразийскому коридору в 1 тыс. до н.э.–1 тыс. н.э.
Моделирование генома князя Дмитрия Александровича из геномов предковых популяций
После анализа различных комбинаций предковых компонент наиболее подходящая модель указывает на вклад в происхождение князя Дмитрия Александровича трех компонент, как у следующих популяций:
Раннесредневекового населения востока Скандинавии с острова Эланд. Или славян, представленных геномами Шекшово9 и Сунгирь6.
Представителей степных кочевых народов евразийских степей железного века или раннесредневекового населения Центральной Европы (степных кочевников с территории Венгрии).
Восточно-евразийской компоненты.
Предсказание фенотипических признаков князя Дмитрия Александровича на основании генетических данных
С большой вероятностью глаза князя Дмитрия Александровича были карими (P = 0.962), волосы темными (P = 0.810) или коричневыми (P = 0.555), а оттенок кожи промежуточным (0.635), т.е. не светлым и не темным.
Выводы
Благодаря палеогенетике и применению статистических методов проверки гипотез, подтверждается связь Рюриковичей со Скандинавией по мужской линии, хотя отсутствие генетических данных от населения с территории севера Руси в период, непосредственно предшествовавший становлению княжеского правления, не позволяет предметно судить о присутствии предков князя Дмитрия Александровича и других Рюриковичей на материковом севере и северо-востоке Европы. Анализ генеалогического древа Рюриковичей показал, что современные представители этого рода, с гаплогруппой Y-хромосомы N1a, группирующиеся с образцом князя Дмитрия Александровича, относятся к трем разным ветвям этого рода – Ольговичам, Мстиславичам и Юрьевичам. Которые восходят к князю Ярославу Мудрому, как к ближайшему их общему предку.
В свою очередь митохондриальная гаплогруппа F1b1 у князя Дмитрия Александровича может свидетельствовать о вкладе восточных популяций в его генотип, что подтверждается и аутосомными данными. Согласно результатам анализа главных компонент и анализа примесей, геном князя Дмитрия Александровича имеет сходство с геномами представителей скандинавских популяций эпохи «викингов», но отличается от них заметным вкладом восточной компоненты. Что может быть связано с распространенной практикой браков русских князей с дочерями половецких ханов с конца XI в. Такие браки скрепляли союзнические отношения и усиливали политическое взаимодействие.
Мать Дмитрия, супруга Александра Невского – Александра Брячиславна происходит из рода полоцких Изяславичей. Сведения о женах этих князей скудны, имя и происхождение матери Александры неизвестно. Однако, известно, что представители полоцкой ветви Рюриковичей не избегали брачных союзов с половчанками. Из половецкого рода происходила вторая жена полоцкого князя Святополка Изяславича (1050–1113), Елена, дочь хана Тугоркана.
Однако восточная компонента в геноме Дмитрия Александровича могла быть получена одновременно и по мужской, и по женской линиям, отражая браки Рюриковичей с представительницами династий Центральной и Южной Европы, как сербские Вукановичи или венгерские Арпады, к примеру. Элиты этих территорий могли в большей степени, чем «простое» местное население, сохранить восточные гены, привнесенные миграционными потоками первого тысячелетия нашей эры. Исходя из перечисленных вероятностей, объяснения присутствия «восточной компоненты» в геноме Дмитрия Александровича, пока остаются гипотетическими, как и состав его предков по материнской линии.
Но несмотря на оставшиеся вопросы, впервые на примере представителя древнего рода Рюриковичей показана генетическая составляющая сложного характера межэтнических взаимодействий в формировании знати средневековой Руси.
Источник: Жур К.В., Шарко Ф.С., Седов В.В., Добровольская М.В., Волков В.Г., Максимов Н.Г., Сеславин А.Н., Макаров Н.А., Прохорчук Е.Б. Рюриковичи: первый опыт реконструкции генетического облика правящего рода средневековой Руси по данным палеогеномики // Acta Naturae. - 2023. - Т. 15. - №3. - C. 50-65. doi: https://doi.org/10.32607/actanaturae.23425
Выспаться, провести генеральную уборку, посмотреть все новые сериалы и позаниматься спортом. Потом расстроиться, что время прошло зря. Есть альтернатива: сесть за руль и махнуть в путешествие. Как минимум, его вы всегда будете вспоминать с улыбкой. Собрали несколько нестандартных маршрутов.