


Виды антропологической выставки в Москве 1879 года























Показать полностью
23
Аланы появляются на Северном Кавказе в начале первого тысячелетия н. э. и упоминаются в древнеримских источниках с I века, а во второй половине II в. н. э. отмечена уже полностью сложившаяся аланская археологическая культура, которая включает как керамический и погребальный комплексы, так и появление в равнинной и предгорной части Центрального и Северо-Восточного Кавказа значительного числа больших (от 0,5 до 1,2 км2) городищ со сложной структурой и четко выделенной цитаделью.
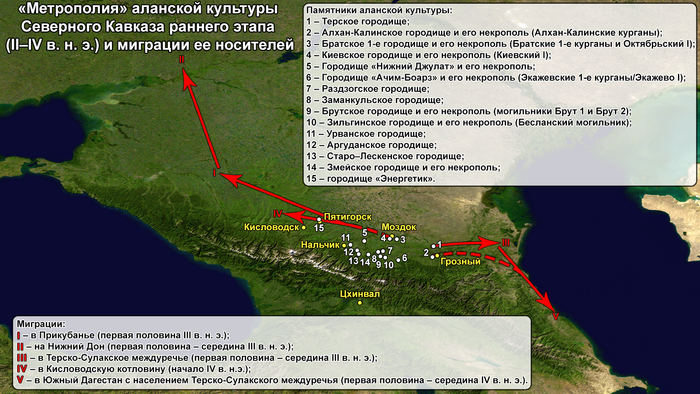
Благоприятные климатические условия являются важной причиной расцвета культуры. Высокая урожайность зерновых культур и увеличение поголовья скота способствовали росту численности населения и его плотности (несоизмеримых с другими периодами древней и средневековой истории региона), а также появлению центров городского типа с развитыми ремеслами. Это привело к распространению носителей аланской культуры по обширным территориям, параллельно с накоплением значительных отличий от исходных вариантов культуры по мере удаления от «метрополии». В конце IV–V вв. н. э. события начала эпохи Великого переселения народов и засушливость климата, усиливающаяся к VII в. н. э., привели к постепенному исчезновению аланских памятников городского типа на равнинах. При этом население перемещалось в горную и предгорную зоны, а также на территорию Среднего Дона. Хронологически аланская культура прослеживается до золотоордынского времени в начале XIII века. После чего потомки алан сохранились в горных районах, язык которых, как считается, и унаследовали современные осетины. Разгром алан золотоордынцами в свою очередь способствовал продвижению адыгских племен на обширные пространства Центрального Кавказа в XII–XIV веках.
Согласно гипотезе аланского происхождения осетин, аланы и ассимилированные ими горные племена сложились в средневековую осетинскую народность, которая сохранила некоторые культурные аланские традиции и иранскую речь. Палеоантропологические данные указывают на преобладание аланского антропологического варианта в раннесредневековом населении Северной Осетии, которое сформировалось на основе слияния ираноязычных племен и местного населения, восходящего к кобанской культуре. Однако после разгрома Алании чингизидами эти антропологические особенности затушевываются из-за миграций населения с равнин и увеличения роли местного горского населения в формировании антропологического облика популяций на территории Осетии.
И хотя в предыдущих палеогенетических исследованиях вопрос аланского генетического наследия специально не рассматривался, но данные о геномах 6 алан V в. н. э. из современной Северной Осетии изучены в контексте генетической истории Евразийской степи бронзового века и более поздних периодов.
При этом у изученных древних жителей не было обнаружено генетической связи с носителями савроматской и сарматских культур из Прикаспийской степи V в. до н. э. – I в. н. э., однако отмечено наибольшее сходство с населением Армении железного века. Впрочем, очень скоро эта ситуация изменится: сейчас сразу несколько научных коллективов изучают древнюю ДНК алан и их соседей по времени и пространству. И хотя каждый коллектив – в зависимости от объема добытого финансирования – изучает небольшую выборку образцов, но в совокупности они могут создать вполне объективный образ алан. Так что с нетерпением ждем их результаты.
В данной работе анализ современного населения Кавказа по очень большой полногеномной панели, в 4,5 млн маркеров, проведен с целью поиска генетического наследия алан у осетин, балкарцев, карачаевцев, ингушей и чеченцев. Потому как именно эти народы, проживая в основном ареале аланских племен, и могли сохранить в своих генофондах некоторое генетическое наследие алан. Основания для оптимизма дает выявленная тесная связь между генетической и лингвистической структурой народов Северного Кавказа.
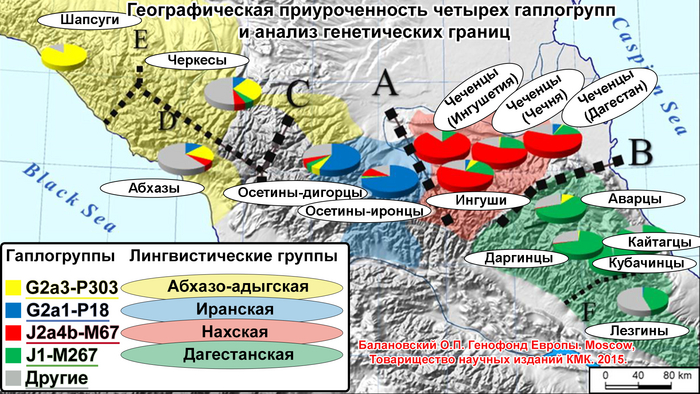
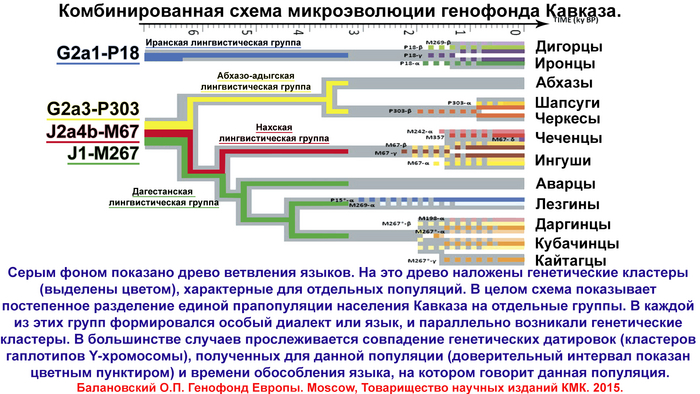
Особенно детально рассмотрен генофонд осетин, поскольку они помимо прочего являются и наиболее вероятными лингвистическими наследниками алан. Поэтому в анализ включены разные субэтносы – общества из Северной Осетии (алагирцы, дигорцы, иронцы, куртатинцы, тагаурцы) и Южной Осетии (ксанцы, кударцы, туальцы). В сравнительный анализ, конечно же, вовлечены и другие народы, которые могли внести свой вклад в генофонды предполагаемых наследников алан.
Анализ 140 независимых моделей предковых компонент показал, что оптимальная по детализации и устойчивости структура генофонда Северного Кавказа проявляется при 10 и 11 предковых компонентах.
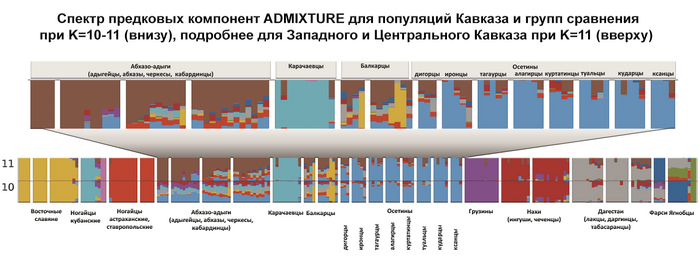
Но поскольку при 11 проявляется новая предковая компонента абхазо-адыгских народов, авторы приводят детальное описание генофонда в предположении 11 компонент – то есть предполагается, что 11 предковых популяций внесли свой вклад в анализируемую совокупность 207 геномов.
При этом в очередной раз стоит напомнить, что только для удобства интерпретации, каждой предковой компоненте дано условное название по той популяции, в геномах представителей которой отмечен её наибольший вклад.
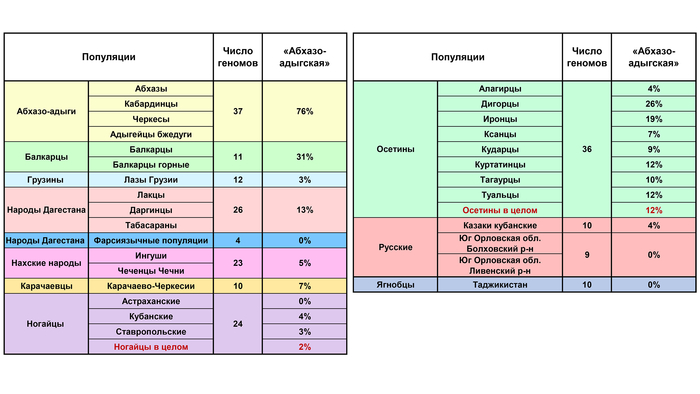
Составляет 76% генофонда абхазо-адыгских народов – адыгейцев, абхазов, черкесов и кабардинцев. Эта предковая компонента примечательна тем, что она единственная из 11 обнаруживается практически у всех представителей проанализированных популяций. Ее нет лишь у фарсиязычных жителей Дагестана, ягнобцев Таджикистана и южных русских. Она составляет треть генофонда балкарцев. У осетин чаще встречается у дигорцев (26%) и иронцев (19%), варьируя в остальных обществах осетин в интервале 4–12%. Ее вклад снижается у чеченцев и ингушей до 5% и вновь возрастает до 13% в Дагестане (в этой работе он представлен геномами даргинцев, лакцев и табасаран). В целом эта компонента доминирует у жителей Западного Кавказа, и с низкими частотами встречается на Центральном и Восточном Кавказе.
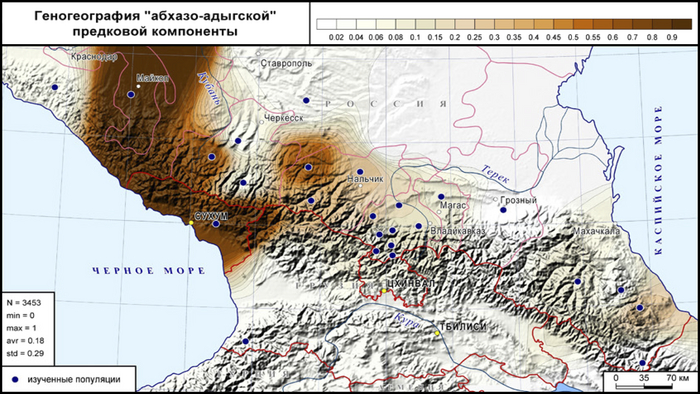
Что вероятно отражает генетические следы обширных миграций носителей этой предковой компоненты в середине второго тысячелетия н.э., но не исключает и след более древнего субстрата в населении Северного Кавказа.
«Осетинская» предковая компонента
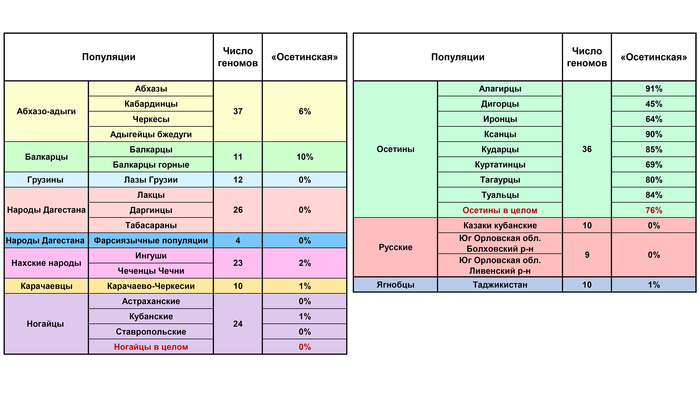
В среднем составляет 76% генофонда осетин, но очень варьирует в разных субэтносах – от 45% у дигорцев до 90% у ксанцев и 91% у алагирцев.
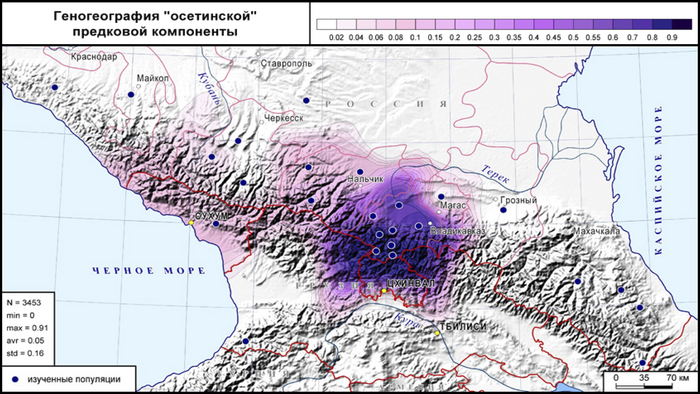
Эта компонента охватывает не только этнический ареал осетин, но и с невысокими частотами область Западного и Центрального Кавказа, составляя 9–10% в генофондах абхазов, балкарцев и кабардинцев, 6% у черкесов, 3% у ингушей и 1% у чеченцев.
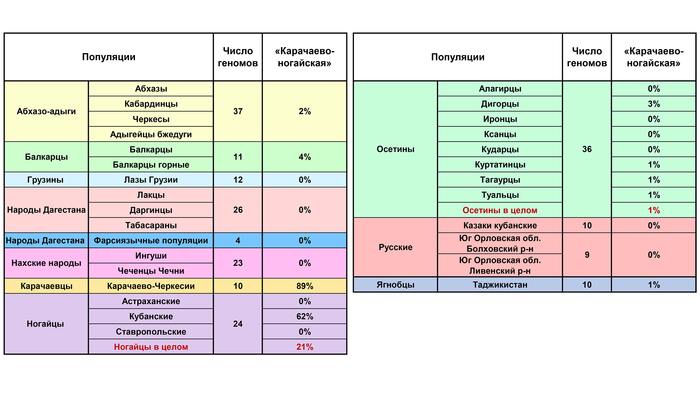
Соответственно доминирует у карачаевцев (89%) и у кубанских ногайцев (62%). На карте её ареал как лоскут одного одеяла подшит к «Абхазо-адыгской» предковой компоненте с севера и востока.
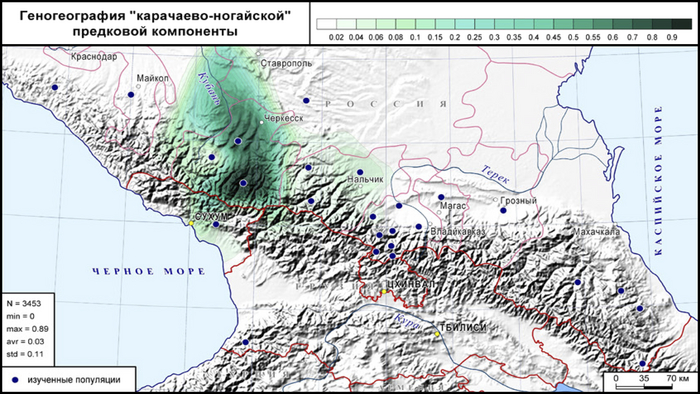
Однако её вклад в соседние популяции черкесов и равнинные популяции балкарцев, невелик (5–6%), а у других жителей Западного и Центрального Кавказа она встречается эпизодически.
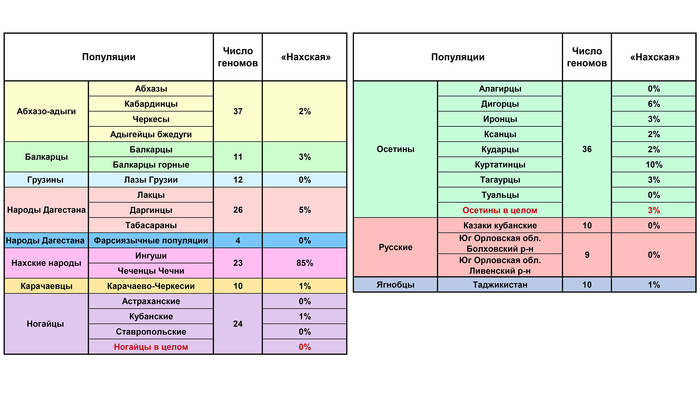
Вклад данной компоненты высок в генофонды ингушей (87%) и чеченцев (83%). Но географически она распространена от Западного до Восточного Кавказа.
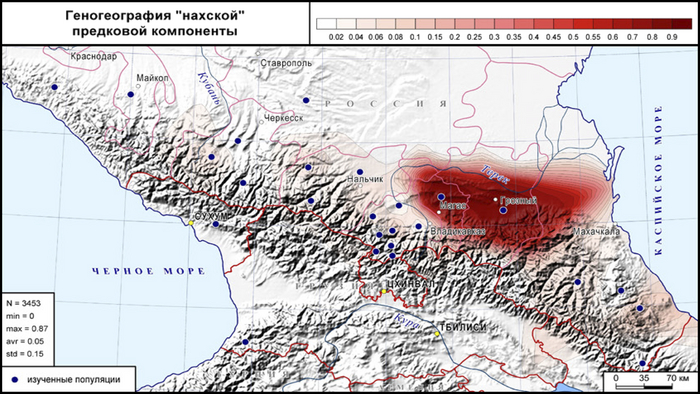
Ее вклад в генофонды осетин-куртатинцев и табасаран Дагестана достигает 10%, в генофонды осетин-дигорцев и лакцев Дагестана 6%, в генофонды Центрального Кавказа (иронцев, тагаурцев) и народов Западного Кавказа (кабардинцев, черкесов, балкарцев) 3–4%.
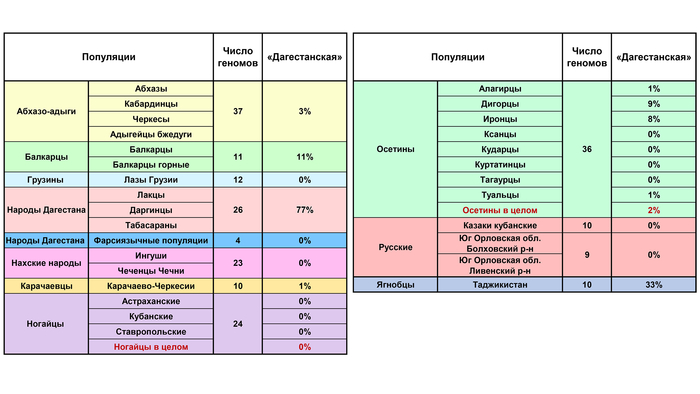
В среднем составляет 77% общего генофонда народов Дагестана, достигая максимума у даргинцев и практически отсутствует у соседних нахских народов Центрального Кавказа. Но отдельные её очаги обнаружены как в Осетии у иронцев с дигорцами (8-9%), так и на Западном Кавказе у балкарцев – 11%.
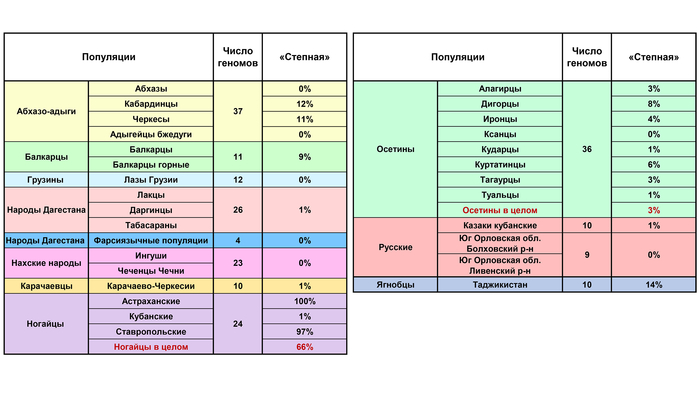
Составляет 97% генофонда ставропольских и 100% астраханских ногайцев. Но у кубанских ногайцев ее практически нет (1%) зато у них преобладает «Карачаево-ногайская» компонента (62%). Возможно, именно «степным» влиянием можно объяснить заметный вклад этой компоненты в генофонды Карачаево-Черкесии и Кабардино-Балкарии (11-12% у кабардинцев и черкесов, 9% у балкарцев), а также у северных осетин (3–8%.)
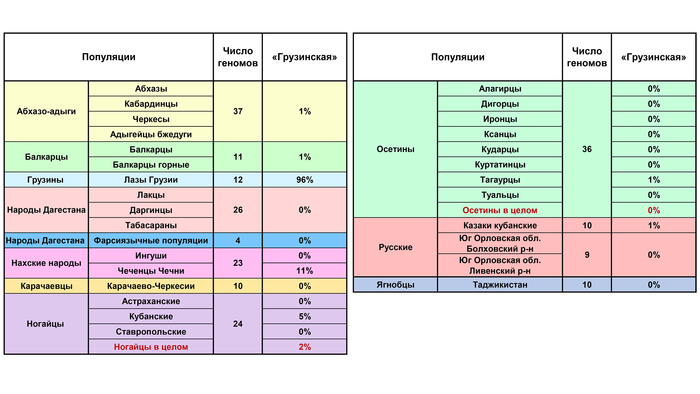
Кроме лазов Грузии, у которых вклад этой предковой компоненты достигает 96%, она на заметном уровне обнаружена у чеченцев (11%) и абхазов (5%).
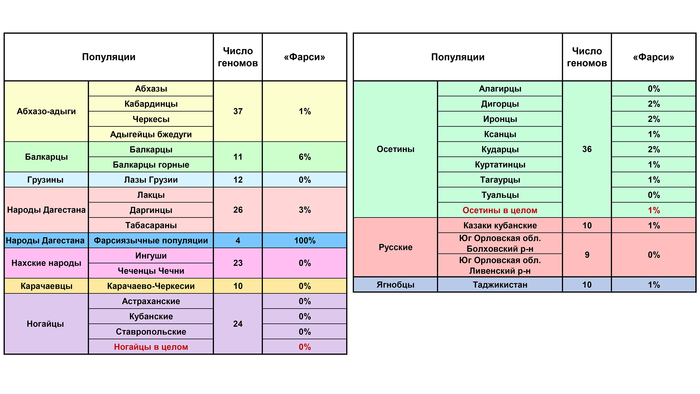
Кроме 100% у фарсиязычных жителей Дагестана (включенных в анализ в качестве представителей юго-западной ветви иранских языков), выявлена также с заметным вкладом у балкарцев – 6%.
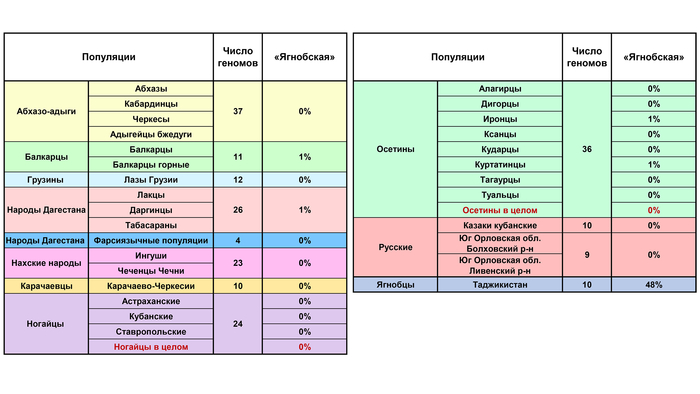
Даже у ягнобцев Таджикистана вклад этой компоненты составляет лишь 48%, отсутствуя в остальных популяциях. Географически далекие ягнобцы включены в анализ потому, что они близки лингвистически: ягнобцы и осетины являются сейчас единственными живыми представителями северо-восточной ветви иранских языков, в которую ранее входили и сарматские языки, и аланский.
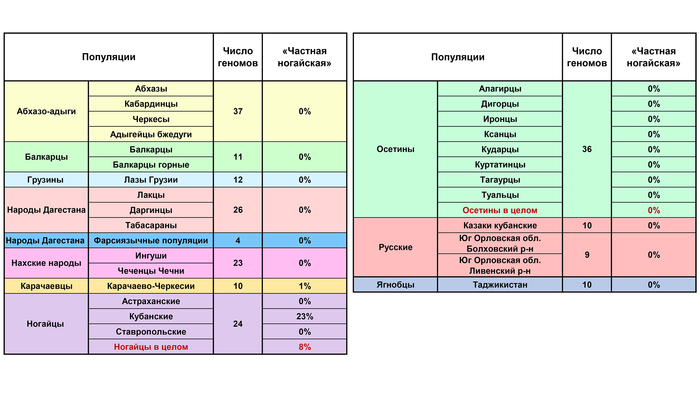
Ярко проявилась только в двух геномах кубанских ногайцев. Но её небольшой вклад обнаруживается и в отдельные геномы других народов, что, вероятно, отражает уже третий вариант тех прапопуляций, пришедших из евразийской степи, которые вместе с носителями «карачаево-ногайской» и «степной» компонент участвовали в сложении генофондов ногайцев, карачаевцев и балкарцев.
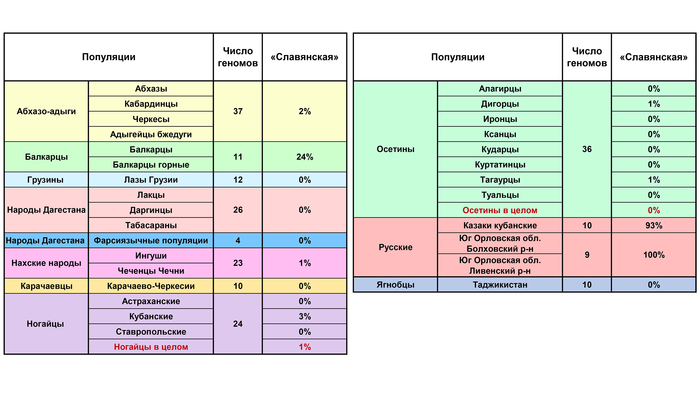
Эта компонента отражает поздний вклад славянского населения в генофонды коренного населения Кавказа. И ожидаемо ее вклад составляет 100% у южных русских и 93% у кубанских казаков. Однако она также составляет 24% генофонда балкарцев, 3% кубанских ногайцев, 2% абхазо-адыгских народов и около 1% у некоторых других народов.
Полученные результаты показывают, что в генофонде современного населения Кавказа отсутствует единственная предковая компонента, которую можно было бы напрямую связать с аланским генетическим наследием.
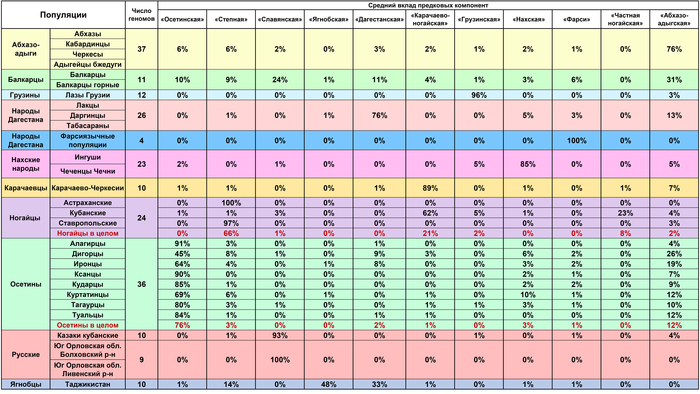
Ареалы всех четырех преобладающих предковых компонент, а именно «абхазо-адыгской», «карачаево-ногайской», «осетинской» и «нахской», складываются друг с другом как лоскуты одного одеяла, сшитые на периферии своих ареалов. Более того, суммарно эти четыре компоненты составляют основную часть генофондов тех современных народов, у которых предполагается наличие «аланского генетического следа». Общий вклад четырех «прапопуляций» составляет: 99% у ингушей, 98% у карачаевцев, 95% у осетин, 85% у чеченцев и только 49% у балкарцев.
Стоит отметить, что наличие даже заметного вклада какой-то предковой компоненты в генофонде популяции может не иметь отношения к её древней истории, а может быть связано лишь с отдельными недавними мигрантами. Но в этом случае данная компонента будет преобладать в одних геномах и полностью отсутствовать в других. И наоборот, даже небольшая доля предковой компоненты, но обнаруженная в большинстве геномов популяции, может отражать важный этап ее генетической истории, некий древний пласт ее генофонда. Учитывая эти особенности стоит рассмотреть каков вклад «абхазо-адыгской», «карачаево-ногайской», «осетинской» и «нахской» компонент в геномы основных номинантов на аланское генетическое наследие – балкарцев, ингушей, карачаевцев, осетин, чеченцев.
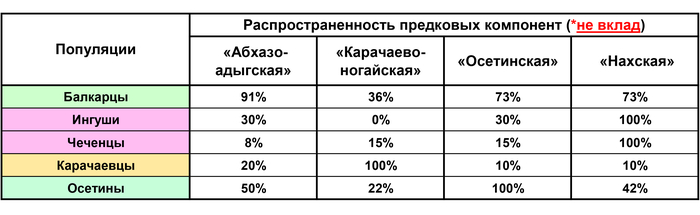
«Абхазо-адыгская» компонента, которая предположительно отражает вклад некоего автохтонного населения Кавказа, представлена практически у всех балкарцев (91% геномов), среди 50% геномов осетин, 30% ингушей, 20% карачаевцев, 8% чеченцев, а также у половины геномов Дагестана (причем у всех табасаран и у трети лакцев).
«Карачаево-ногайская» компонента присутствует у всех карачаевцев, среди 36% геномов балкарцев, 22% осетин, 15% чеченцев и отсутствует у ингушей. Она характерна почти для всех кубанских ногайцев и при этом отсутствует у астраханских и ставропольских ногайцев. Можно предположить, что эта компонента отражает след лишь одной из миграций тюркоязычных популяций золотоордынского времени, которые могли резко генетически различаться.
«Осетинская» компонента присутствует у всех осетин, у 73% балкарцев, 30% ингушей, 15% чеченцев, 10% карачаевцев и даже у 8% народов Дагестана. Наличие ее среди заметного числа геномов всех пяти этносов-«номинантов» позволяет выдвигать ее как кандидата на аланский генетический след.
«Нахская» компонента присутствует во всех геномах ингушей и чеченцев, у 73% геномов балкарцев, 42% осетин, 10% карачаевцев и даже у 38% геномов народов Дагестана. То есть и эта компонента оставила внушительный вклад в геномах всех пяти этносов-«номинантов» и потому с не меньшим основанием может служить кандидатом на аланский генетический след (хотя он является второстепенным по сравнению с более древним субстратом прото-чечено-ингушского населения). Это позволяет выдвинуть гипотезу, что обе предковые компоненты, а именно –«осетинская» и «нахская» – лишь отчасти отражают аланский генетический след при этом каждая по-своему. Поэтому есть смысл рассмотреть их общий вклад более детально.
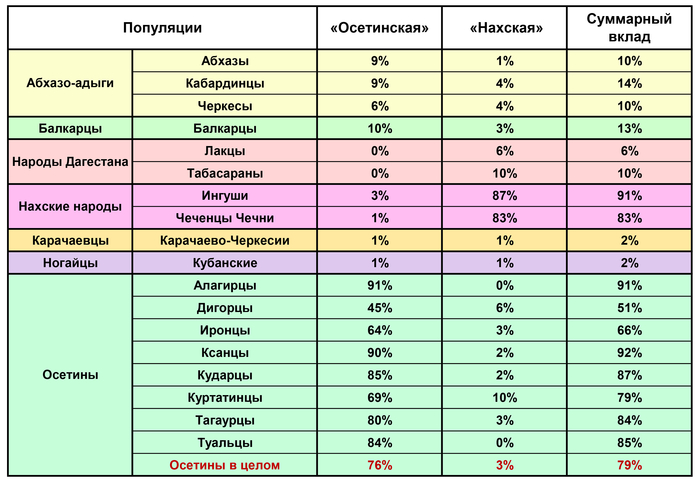
При рассмотрении индивидуальных геномов, в которых обнаруживается вклад и «осетинской», и «нахской» компонент, выявляет такой градиент: осетины, ингуши, чеченцы, балкарцы, карачаевцы, народы Дагестана. Однако их суммарный вклад выявляет иной градиент: ингуши (91%), чеченцы (83%), осетины (79%), кабардинцы (14%), балкарцы (13%), абхазы, табасараны, черкесы (10%), лакцы (6%), карачаевцы, кубанские ногайцы (2%).
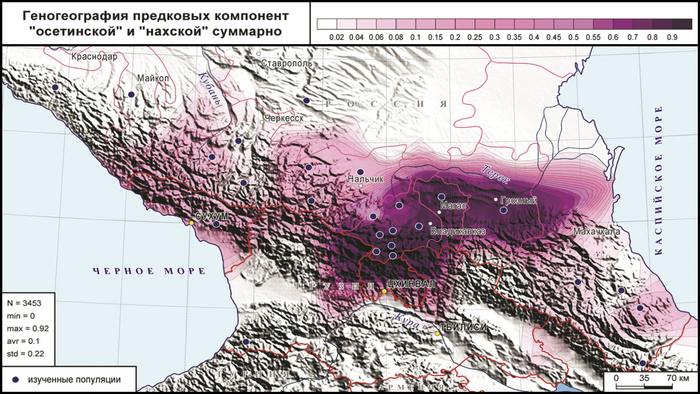
Причем в пределах генофонда осетин обнаружены важные различия: максимальный вклад (>90%) обеих компонент приходится на алагирцев и ксанцев, а наименьший – на дигорцев (51%) и иронцев (66%).
Особое положение занимают куртатинцы, у которых наибольший среди осетин вклад «нахской» компоненты (10%). При этом в популяциях осетин выявляется клинальная изменчивость «осетинской» компоненты: ее вклад возрастает с севера на юг (в среднем 70% у северных и 86% у южных).
Но эта изменчивость связана в основном с противоположным вкладом «абхазо-адыгской» компоненты: у северных обществ осетин в среднем 14% (с максимумом 26% у дигорцев), а у южных – 9%.
ЗаключениеАнализ вклада предковых компонент в генофонды современных народов Северного Кавказа, проживающих в основном ареале аланских племен, не обнаружил генетический след одной предковой прапопуляции, которую можно было бы связать с аланским генетическим наследием. Однако две предковые компоненты – «осетинская» и «нахская» – дают наибольший вклад и в индивидуальные геномы, и в популяционные генофонды основных номинантов на звание наследников алан – осетин, ингушей и чеченцев.
Генофонд балкарцев предстает как наиболее сложносоставной – с наибольшим вкладом «абхазо-адыгской» компоненты (31%) и с небольшим суммарным вкладом «осетинской» и «нахской» компонент (лишь 13%). Генофонд карачаевцев в основном состоит из предковой компоненты, общей с ногайцами, отражая след одной из миграций тюркоязычных популяций золотоордынского времени. Геногеографический анализ распределения вклада основных кандидатов на аланский генетический след («осетинской» и «нахской» компонент), выявленная неоднородность осетинских обществ и лингвистическое сходство осетин с аланами поддерживает гипотезу, что именно среди осетинских обществ сохраняется вероятность найти аланский генетический след.
Тем не менее, принимая в расчет и геногеографический ареал, и вклад в геномы других популяций, нельзя отвергнуть гипотезу, что обе предковые компоненты – «осетинская» и «нахская» – могут отражать генетическое наследие алан, хоть это и допускает совокупность очень маловероятных событий.
В любом случае, исходная гипотеза о наличие слабого общего аланского следа в генофондах народов, проживающих в ареале аланской «метрополии» и которых принято считать потомками алан, не подтверждена. Возможно, этот след в современных генофондах настолько слаб, что лишь анализ древней ДНК сможет его выявить.
Также стоит отметить, что помимо поиска наследия алан, в работе выполнен анализ селективных, фармакогенетических ДНК маркеров, которые демонстрируют популяционную структуру Северного Кавказа, сформированную под действием отбора. Фармакогенетические подходы применяются при разработке протоколов по подбору оптимальных лекарственных препаратов и их доз в зависимости от генотипа пациента.
Анализ выполнен с акцентом на его географический центр, приходящийся на наиболее детально изученных осетин: построена карта генетических расстояний Нея по совокупности 42 наиболее значимых фармакогенетических маркеров от популяции осетин до остальных популяций изучаемого региона. В отличие от карт предковых компонент, созданных по селективно-нейтральным маркерам, которые связаны с миграциями и дрейфом геном, по фармакогенетически значимым маркерам выявлены дополнительные закономерности генетического сходства, которые отражают действие отбора.
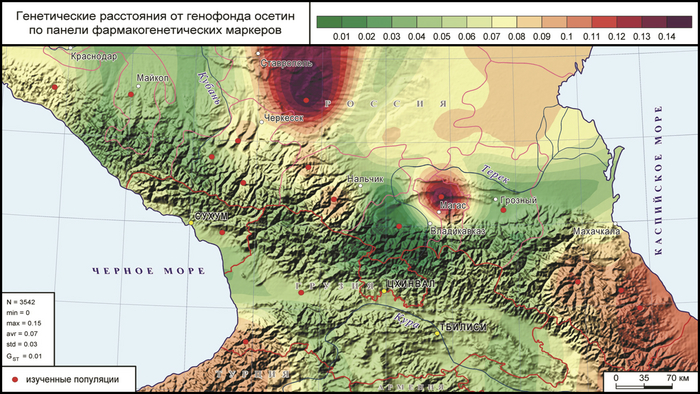
На первом месте по степени сходства с осетинами - чеченцы, на втором черкесы и адыгейцы, на третьем абхазы, а далее по снижению сходства: карачаевцы, кубанские ногайцы, кабардинцы, балкарцы и народы Дагестана. В целом ареал наибольшего сходства с осетинами тянется с востока на запад вдоль Кавказского хребта.
А наибольшие фармакогенетические отличия от осетин обнаружены у ставропольских ногайцев и, что было крайне неожиданно, у ингушей. Наиболее ярко выраженное сходство осетин по фармакогенетическому статусу с чеченцами и резкое отличие от ингушей, ярко контрастирует с данными, полученными по селективно-нейтральным маркерам в рамках метода примесей (ADMIXTURE), где генофонды чеченцев и ингушей почти неотличимы друг от друга. Такие особенности фармакогенетических ДНК-маркеров могут указывать на действие на них отбора в популяциях Центрального Кавказа и необходимость дальнейших исследований этого явления для последующего перехода к персонифицированной медицине.
Источник: Балановская Е.В., Агджоян Анастасия Т., Горин И.О., Петрушенко В.С., Пылёв В.Ю., Кулемин Н.А., Маркина Н.В., Черкасов Н.А., Кошель С.М., Агджоян Анна Т., Почешхова Э.А. В поисках аланского следа: генетическая история Северного Кавказа по полногеномным данным об аутосомном генофонде // Вестник Московского университета. Серия XXIII. Антропология, 2022; 3/2022; с. 48-62 (Published: September 15, 2022) DOI: 10.32521/2074-8132.2022.3.048-062
Удивился, когда искал информацию)
https://volga.news/article/317226.html
Оказывается, на Самарской набережной есть такая поделка. Мне трудно назвать это скульптурой.
Как взаимосвязаны неандертальцы и кроманьонцы со своими общими предками? Где, когда и от кого возникли эти параллельные человечества? Как современная антропология объясняет происхождение неандертальцев и кроманьонцев? Станислав Дробышевский, антрополог, кандидат биологических наук, доцент кафедры антропологии биологического факультета МГУ имени М. В. Ломоносова рассказывает, как шла эволюция неандертальцев и кроманьонцев, какие орудия труда они изготавливали, чем отличаются их культуры, как выглядели первые кроманьонцы и к чему привело их взаимодействие с неандертальцами.
ВКонтакте: https://vk.com/video-190320587_456240061
В современных учебниках эволюционная линия превращения обезьяны в человека претерпела значительные преобразования, точнее почти полностью исчезла. Как выяснилось, подавляющее большинство кандидатов на предки человека сосуществовали друг с другом, как сейчас мы и шимпанзе. А потому вместо единого "ствола" сейчас рисуют только "ветви". Эта учась постигла и "икону" антропологии - австралопитека афарского ("Люси"). По мнению антропологов он не имеет никакого отношения к нашей родословной и это после 40-летнего триумфального шествия по страницам учебников. В начале у учёных возникли сомнения в его прямохождении, затем в его скелете (через 40 лет) обнаружили кость современной обезьяна и, наконец, находка целого черепа наглядно продемонстрировала, что Australopithecus afarensis не может быть предком человека. Хорошая новость для научпопа. Ссылки на научные публикацию в видео.
Введение
Процесс неолитизации в Европе привел к серьёзным технологическим и культурным сдвигам, которые включали в себя новые методы ведения хозяйства. Существуют две основные группы моделей, которые объясняют процесс неолитизации. Модели демической диффузии описывают неолитизацию как процесс колонизации земледельцами, который был движим экспоненциальным ростом населения, характерным для неолита. Вторая группа моделей – аккультурации – описывает процесс, в котором, по крайней мере, переход к земледелию мог практиковаться местными охотниками-собирателями после периода (той или иной продолжительности) взаимодействия с соседними, пришлыми земледельцами.
Анатолийские земледельцы достигли Балкан и других регионов Юго-Восточной Европы в седьмом тысячелетии до нашей эры и впоследствии распространились дальше через Средиземное море, а затем через Дунайский коридор, существенно вытеснив коренное мезолитическое европейское население.
В отличие от Центральной Европы, области Восточной Европы, включая Украину, Молдову, Западную Россию и Румынию, не занимались сельским хозяйством до позднего неолита (~ 4500 г. до н. э.), хотя различные оседлые и полуоседлые группы охотников-собирателей этих регионов начали использовать глиняную посуду еще в 8500 г. до н. э.
Археологическая общность Триполье-Кукутень представляет собой группу нескольких взаимосвязанных археологических культур среднего неолита/энеолита на территории ряда областей современных Украины, Молдовы и Румынии. Этот комплекс простирается от Южных Карпат до Днепра и назван в честь типовых памятников Кукутень в Яссы (Румыния) и Триполье (Обуховский район Киевской области).
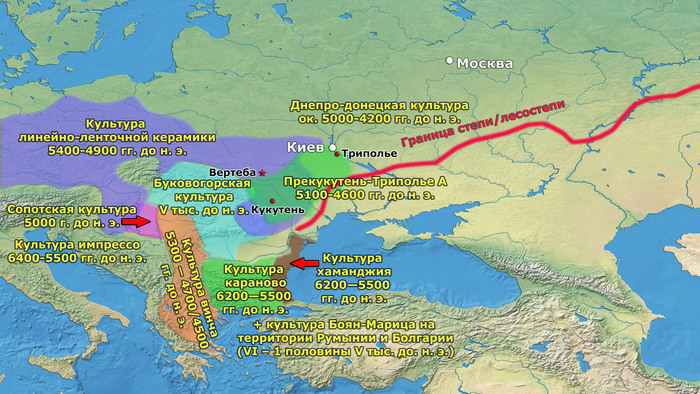
Культуры Кукутень и Триполье имеют общие корни в культуре Прекукутень; самые ранние стоянки археологической общности Триполье-Кукутень обнаружены в предгорьях Карпат, а самые ранние радиоуглеродные даты (от периода Прекукутень II) относятся примерно к 4800 г. до н.э. Археологическая общность Триполье-Кукутень возникла в результате взаимодействия нескольких дунайских неолитических групп, о чем свидетельствует сходство в строительстве домов, керамических стилях и производстве каменных артефактов.

Rassamakin, Y., & Menotti, F. (2011). Chronological Development of the Tripolye Culture Giant-Settlement of Talianki (Ukraine): 14C Dating vs. Pottery Typology. Radiocarbon, 53(4), 645-657. doi:10.1017/S0033822200039102
После возникновения этого культурного комплекса в Карпатском предгорье археологическая общность Триполье-Кукутень расширяла свой ареал и в конечном итоге заняла территорию, охватывающую большую часть современных территорий Украины, Молдовы и Румынии. Первые раннетрипольские (Триполье А) стоянки отошли от культуры Прекукутень ~ 4500 г. до н. э. в долине реки Днестр.
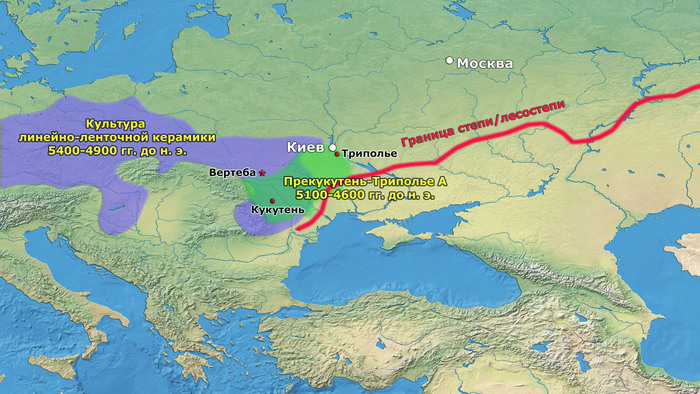
Более поздние перемещения населения, происходящие со среднего периода (Триполье BI) и далее, привели к тому, что трипольская культура распространилась на Волынь на западе и Днепр на востоке.
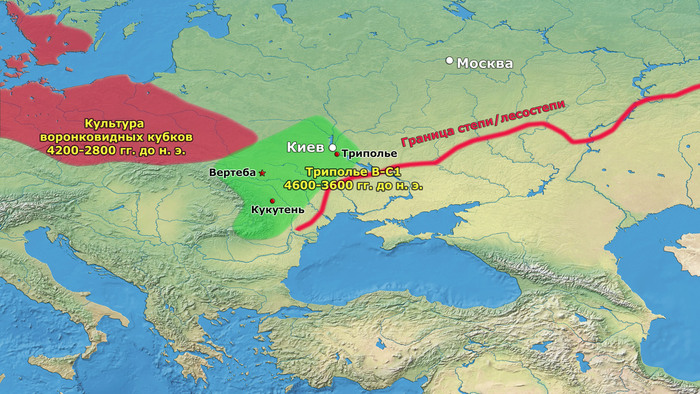
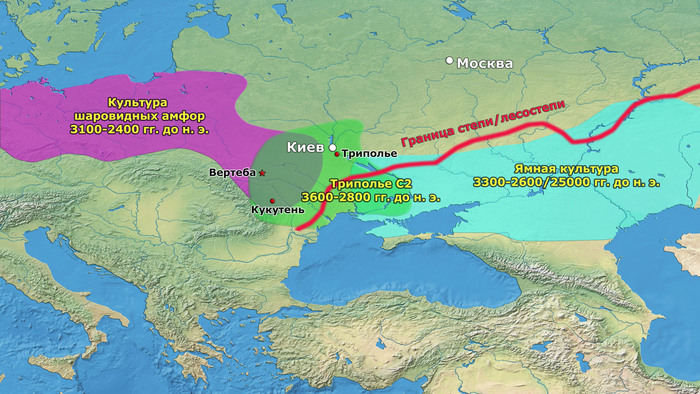
Считается, что эта территориальная экспансия была вызвана в первую очередь демографическим ростом, связанным с успешной стратегией земледелия и животноводства, с поиском новых пахотных земель для возделывания. Однако некоторый рост населения мог быть результатом включения в состав трипольцев местных групп охотников-собирателей, например, представителей буго-днестровской культуры.
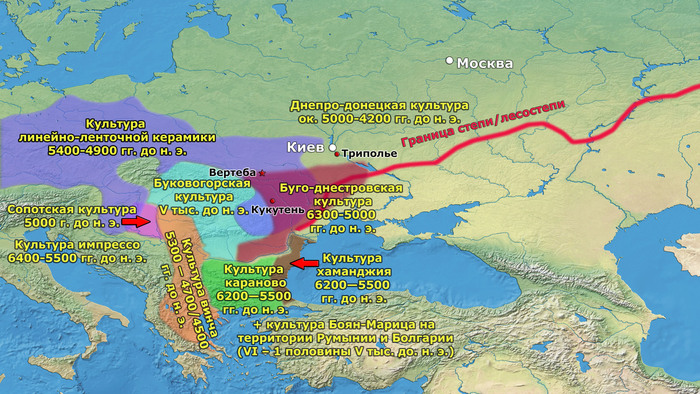
Другим каналом увеличения населения могла быть аккультурация переселенцев после краха неолита в Румынии, Венгрии и Болгарии. В средние и поздние периоды трипольской культуры (Триполье от BII до CI; 4100–3400 гг. до н. э.) некоторые группы трипольской культуры основали чрезвычайно крупные поселения в Центральной Украине, которые достигли размеров 100–320 га.

Быстрый демографический рост трипольцев на рубеже 4 тысячелетия до н. э. потребовал освоения новых территорий, ускорив миграцию в ранее периферийные районы.
Гипотезы возникновения мегапоселений разнообразны. Они могли быть защитным ответом на угрозы, исходящие от степных скотоводов или же от конкурирующих популяций внутри самой археологической общности Триполье-Кукутень. Они могли и просто быть временным эпизодом компактного скопления поселений из-за крупномасштабной миграции из Приднестровья.
Однако, несмотря на высокую плотность поселений в Западной и Центральной Украине, здесь обнаружено крайне мало захоронений. Лишь несколько кладбищ позднетрипольского периода были раскопаны в 1960-х и 1970-х годах, например, Чапаевка на территории Украины и Выхватинцы в Приднестровье. Хотя эти памятники дают некоторое представление о погребальных обрядах трипольцев, они ограничены по временному охвату и не подвергались современным лабораторным исследованиям.
Пример по могильнику в Выхватинцах:Чтобы лучше понять происхождение, связи и разнообразие населения археологической общности Триполье-Кукутень, авторы исследовали палеоантропологические образцы из трех камер на участке пещеры Вертеба в Тернопольской области Украины - одном из немногих мест, где они обнаружены.
Положение скелетов в могилах различно — наряду с преобладающими погребениями в скорченном положении на левом или на правом боку отмечено также несколько случаев погребений на спине, с согнутыми и поднятыми в коленях ногами. Ориентировка довольно устойчива — наибольшее число погребенных положено головой на северо-восток, с небольшим отклонением к востоку. В погребальном инвентаре численно преобладает керамика, представленная двумя типами: сосудами из розовой глиняной массы, украшенными черной и красной росписью; и нерасписными сосудами слабого обжига, орнаментированными веревочными оттисками.
Формы и стиль росписи керамики — типичные для позднего Триполья. Число сосудов в погребениях различно — от 1 до 8, чаще — 2-5. Помимо керамики, в мужских погребениях обнаружены роговые мотыги, кремневые вкладыши для серпов, костяные шила, а также каменный топор, кремневый нож, медное шило, глиняная и костяная антропоморфные статуэтки. Женские погребения сопровождались в основном сосудами, лишь в одном были найдены два пряслица и раковина. Выделяется своеобразием инвентарь детских погребений. Кроме сосудов, здесь были найдены антропоморфные глиняные погремушки; антропоморфные женские статуэтки были положены также главным образом в детские могилы. Помимо этого, обнаружены пряслица, бусы из раковин, кремневые отщепы, пластины, костяное шило.
М. С. Великанова «Палеоантропология Прутскоднестровского междуречья» (Издательство «Наука», 1975 г)
Результаты исследования методами масс-спектрометрии углерода-14 из останков людей и фауны указывают на присутствие трипольцев в пещере Вертеба в интервале между 3950 и 3520 гг. до н. э.
Lillie, M., Budd, C. E., Potekhina, I., Price, T. D., Sokhatsky, M., & Nikitin, A. G. (2017). First isotope analysis and new radiocarbon dating of Trypillia (Tripolye) farmers from Verteba Cave, Biche Zolote, Ukraine. Documenta Praehistorica, 44, 306–324. doi.org/10.4312p.44.18По данным керамических комплексов, присутствующих в пещере, и результатам жидкостно-сцинтилляционного метода анализа углерода-14 с более низким разрешением можно предположить, что заселение продолжалось в течение некоторого времени в период позднего Триполья (CII) и перехода к раннему бронзовому веку.
Помимо этого, масс-спектрометрия также выявила отложения в разных местах пещеры, относящиеся к мезолиту (7950–7490 гг. до н. э.), бронзовому веку, железному веку и средневековью.
Останки людей были извлечены из трех отдельных камер пещеры: участок 7, участок 17 и участок 20. Каждая из этих камер содержит материальную культуру, соответствующую археологической общности Триполье-Кукутень.

Однако захоронения в пещере носят вторичный характер, а нарушения, вызванные деятельностью человека в древности и процессом перемешивания останков животными или растениями, усложняют реконструкцию использования пещеры и хронологии. Большинство изученных образцов в этом исследовании происходят из участка 7, который был подробно задокументирован с помощью анализа керамики и радиоуглеродного датирования, причем пик использования этого участка древним населением датируется периодами CI и CII трипольской периодизации (~ 3900-3350 гг. до н. э.).

Фото: Педагог Светлана - CC BY-SA 4.0, commons.wikimedia.org/w/index.php?curid=92007799Интерпретации использования пещеры разнообразны: в качестве временного убежища, места проведения ритуалов или же погребения. Имеются дополнительные сведения, подтверждающие гипотезу о жертвах конфликтов или же человеческих жертвоприношений.
Палеогенетика трипольской популяции до сих пор включила анализ однородительских маркеров мтДНК и полногеномный анализ 8 представителей трипольской культуры.
Митохондриальные гаплогруппы, типичные для древних евразийских земледельческих групп (H, HV, T, K, J), выявлены у образцов из всех трех камер пещеры. В одной из камер обнаружена гаплогруппа W, характерная для степных популяций, связанных с культурами шнуровой керамики и унетицкой культуры Среднего Поволжья.
Полногеномный анализ представителей археологической общности Триполье-Кукутень показал, что предковые генетические компоненты преимущественно (60–80%) относятся к ранненеолитическим земледельческим группам, подтверждая, что ранние земледельцы, заселившие Западную и Центральную Украину, в значительной степени происходили из той же исходной популяции, что и земледельцы Анатолии и Западной Европы. Оставшиеся 40-20% предковых компонент определены с меньшей достоверностью.
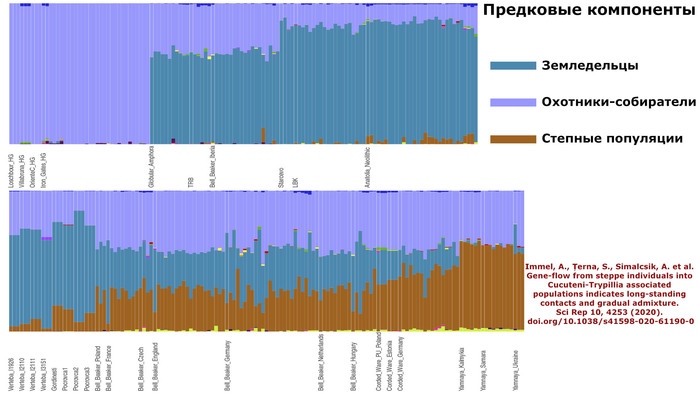
Immel, A., Țerna, S., Simalcsik, A. et al. Gene-flow from steppe individuals into Cucuteni-Trypillia associated populations indicates long-standing contacts and gradual admixture. Sci Rep 10, 4253 (2020). doi.org/10.1038/s41598-020-61190-0Авторы работы 2018 г. [Mathieson et al.] показали, что этот наследственный генетический компонент присутствовал у западных и восточных охотников-собирателей, населявших этот регион в эпоху неолита.
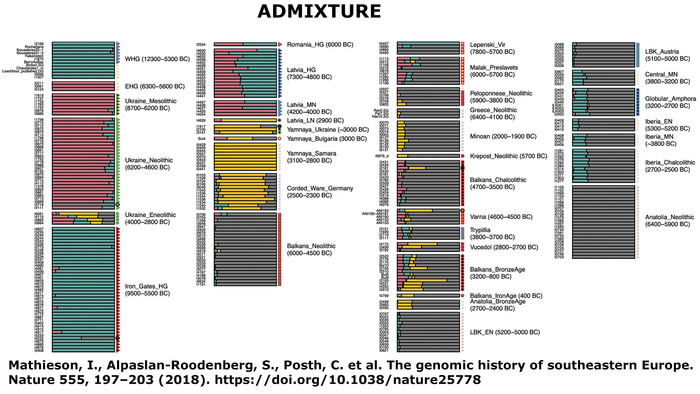
Mathieson, I., Alpaslan-Roodenberg, S., Posth, C. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018). doi.org/10.1038/nature25778Полногеномные данные четырех человек, происходящих из двух разных памятников на севере Молдовы (поздний трипольский период, 3500–3100 гг. до н. э., то есть на пять столетий моложе пещеры Вертеба), выявили у них значительный вклад степного населения (хотя и в разных пропорциях). Это может быть объяснено постепенной ассимиляцией местных мезолитических и неолитических групп охотников и собирателей трипольским населением, жившем на территории современной Молдовы и Приднестровья.
Ареал трипольцев расположен таким образом, что они контактировали как с центральноевропейскими, так и со степными популяциями. Археологические свидетельства взаимодействия с населением степей обнаруживаются в обожженной глиняной посуде. Некоторые из них выглядят почти идентично керамике, найденной в степях, в то время как другая керамика сочетает обжиг с декоративными мотивами трипольцев. Обнаружены и символические предметы, находящиеся под влиянием или непосредственно импортированные из степных сообществ. Например, каменные булавы обнаружены на некоторых стоянках среднего и позднего Триполья, а обмен глиняной посудой очевиден уже в Триполье BII.
Несомненно, существовала определенная степень взаимодействия между трипольским населением и днепро-донецкой культурой, в то время как взаимодействие трипольской культуры и последующей ямной, вероятно, было очень кратким.
Тем не менее, некоторые трипольские популяции, вероятно, находились в постоянном контакте со степными популяциями. После 3400 г. до н. э. трипольские мегаполисы были в значительной степени заброшены. Причина этого широко обсуждалась. Одна из гипотез - усиление конфликтов из-за расширения степных культур на запад. Такая гипотеза может найти подтверждение в частых свидетельствах насильственной смерти в пещере Вертеба.
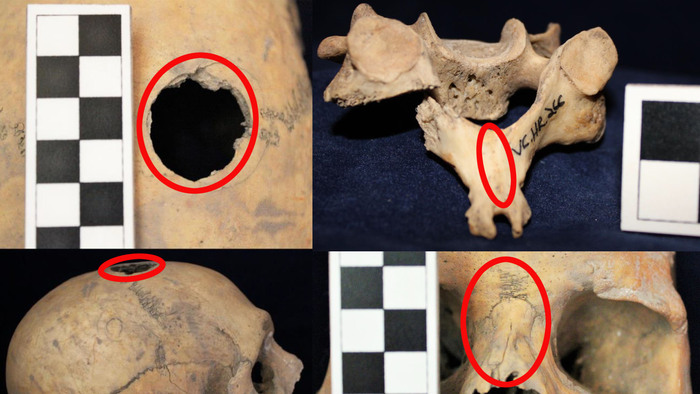
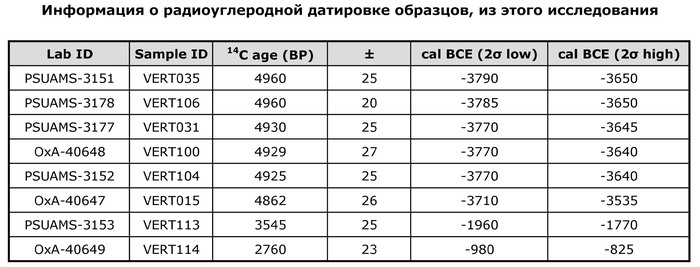
Примеры травм у захороненных в пещере Вертеба (Violence at Verteba Cave, Ukraine: New insights into the Late Neolithic intergroup conflict G.D. Madden, J.K. Karsten, S.H. Ledogar, R. Schmidt, M.P. Sokhatsky doi.org/10.1002/oa.2633)В данном исследовании авторы реконструировали последовательности генома 20 человек (8 женщин и 12 мужчин), захороненных в пещере Вертеба. Восемь образцов датированы масс-спектрометрией по углероду-14 в интервале от 3790 до 825 гг. до н. э, что соответствует трипольской культуре и позднее. Шесть человек (образцы ВЕРТ-035, ВЕРТ-106, ВЕРТ-031, ВЕРТ-100, ВЕРТ-104 и ВЕРТ-015) датируются поздним энеолитом (3790–3535 гг. до н.э.), один (VERT-113) из Зоны 7 датируется средним бронзовым веком (1960–1770 гг. до н.э.) и один из Зоны 17 (VERT-114) датируется поздним бронзовым веком (980–825 гг. до н.э.).
Используя полученные данные, авторы задались целью ответить на ряд вопросов:
1. Есть ли доказательства метисации с местными охотниками и собирателями, как было предположено в предыдущих работах?
2. Используя расширенный набор данных, можно ли уточнить неолитическую предковую составляющую трипольцев: сходны они с ранними земледельцами из Анатолии, представителями культуры линейно-ленточной керамики или иными?
3. Поскольку трипольцы жили в непосредственной близости от степных популяций, есть ли доказательства генетической близости к ямной или более ранним степным популяциям?
4. Имеет ли население более позднего бронзового века, заселившее регион, генетическую преемственность с представителями археологической общности Триполье-Кукутень из пещеры Вертеба?
Полученные результаты
Однородительские маркеры (мтДНК и Y-хромосомы)
Проанализированные образцы эпохи энеолита относятся к гаплогруппам митохондриальной ДНК T2b, H, HV, K1, N1, J1, U5 и T2c.
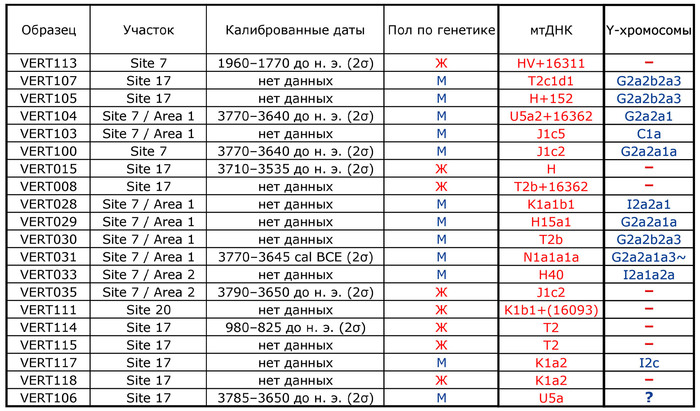
У представительницы среднего бронзового века (VERT-113) определена гаплогруппа HV, типичная для таких неолитических культур, как культура линейно-ленточной керамики к востоку от Дуная – Альфёльда, а также для жителей европейского бронзового века. Представительница позднего бронзового века (VERT-114) демонстрирует гаплогруппу T2, также связанную с многими культурами бронзового века. В целом эти гаплогруппы обычно встречаются в европейских популяциях эпохи неолита и бронзового века.
У мужчин выявлены гаплогруппы Y-хромосомы G2a, C1a и I2 (более подробно в таблице), которые также были выявлены в популяциях неолита и бронзового века Европы. При этом как гаплогруппы мтДНК, так и гаплогруппы Y-хромосомы всех образцов из Вертебы полностью согласуются с ранее опубликованными данными.
Полногеномные данные
На график анализа главных компонент вместе с 20 геномами из пещеры Вертеба авторы поместили 729 представителей современных популяций из Европы, а также 478 ранее опубликованных древних генома.

18 из 20 человек из Вертебы тесно связаны с европейскими популяциями эпохи неолита и энеолита: культура линейно-ленточной керамики, средний и поздний неолит Центральной Европы, а также трипольцы из Молдовы.
Анализ главных компонент также показал значительное сходство между 18 новыми образцами из Вертебы и четырьмя другими трипольцами из более ранней работы, также обнаруженных в пещере Вертеба. Поэтому их объединили в 22 образца, пометили на графике как Verteba_Trypillia и в дальнейшем проанализировали совместно.
А вот два генома бронзового века являются явными исключениями. Образец VERT-114 (позднего бронзового века) подпадает под разнообразие представителей культуры колоколовидных кубков и, по-видимому, занимает положение, близкое к чешской, венгерской и польской группам этой культуры. А образец, обозначенный как VERT-113 (среднего бронзового века), близок к европейским популяциям культур шнуровой керамики и срубной, демонстрируя значительное сходство со степным населением. Сразу стоит отметить, что статистический анализ указал на отсутствие доказательств связи со срубной культурой.
При этом все 22 трипольца из пещеры Вертеба содержат наследственный компонент, доминирующий у представителей анатолийского неолита (по методу ADMIXTURE).
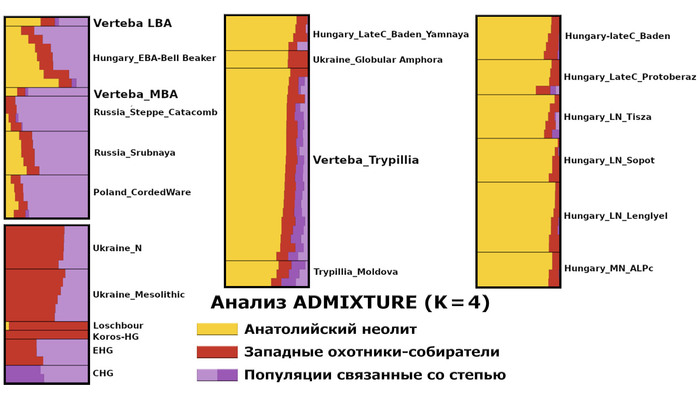
Это указывает на явную связь с европейскими неолитическими популяциями, как собственно и было показано в предыдущих исследованиях. Однако эти же образцы также демонстрируют присутствие компонентов восточных, кавказских и западных охотников-собирателей. Лишь один образец (I3151), по-видимому, не имеет предков восточных/кавказских охотников-собирателей.
В геноме образца эпохи поздней бронзы (VERT-114, укладывающимся в разнообразие представителей культуры колоколовидных кубков) преобладает компонент анатолийского неолита, но выявлен значительный компонент восточных охотников и собирателей. А геном среднего бронзового века (VERT-113) демонстрирует высокую степень сходства с геномами культуры шнуровой керамики и ямной культуры. При этом хотя моделирование геномов трипольцев свидетельствует об их связи со степными популяциями, статистически (f4) они не связаны с популяциями ямной культуры из ареалов современных России и Украины.
Моделируя величину вклада различных основных предков, которые могли бы внести свой вклад в генетический портрет трипольцев, показано, что 40% вклада связано с анатолийским неолитом, 20% - с западными охотниками-собирателями и 40% - с кавказскими охотниками-собирателями.
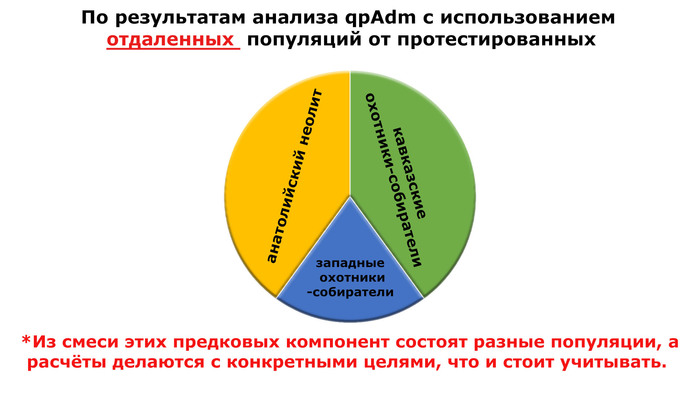
При этом охотники-собиратели юго-восточной Венгрии – Кёрёш (Koros_HG), являются источником западных охотников-собирателей у трипольцев из Вертебы. Также неолит Украины и западные охотники-собиратели являются двумя вероятными источниками генетического пласта, связанного с охотниками и собирателями, для трипольцев из Вертебы, в дополнение к смоделированному компоненту Венгрия-поздний энеолит-ранний бронзовый век-ямная (Hungary_LateC_EBA_Baden_Yamnaya).
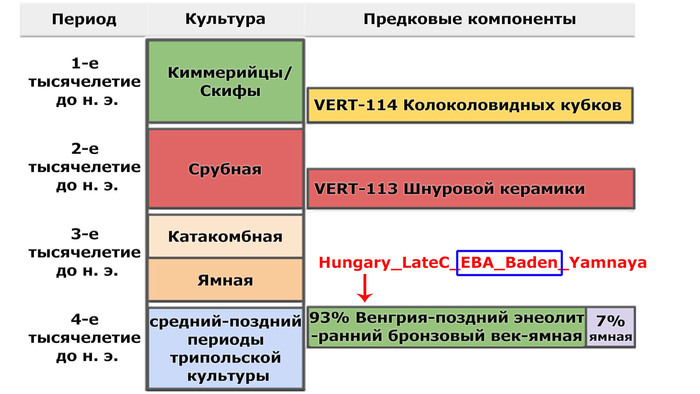
Это также указывает на очень незначительное присутствие среди предков трипольцев местных охотников-собирателей.
Кстати обратите внимание как происходит замена популяций на одной и той же территории и как сильно современные популяции отличаются от древних, вопреки распространяемым мифам.
Также стоит отметить, что большинство трипольцев из пещеры Вертеба не имеют статистически значимого сходства с молдавской популяцией археологической общности Триполье-Кукутень, но этих различий недостаточно, чтобы считать их разными популяциями.
Образец среднего бронзового века (VERT-113) является единственным, который демонстрирует четкий сигнал степного происхождения, причём наблюдается большая близость к ямной культуре с территории России, а не Украины
Кроме того, это единственный образец, который демонстрирует большую близость к представителям неолита на территории Украины, а не к западным охотникам-собирателям. В его геноме фиксируется до 33% вклад неолита Украины и 66% кавказских охотников-собирателей, что подтверждает большую долю степного происхождения.
Образец позднего бронзового века (VERT-114) генетически близок к геномам колоколовидных кубков на графике главных компонент и при моделировании «примесей» (ADMIXTURE). Однако, хотя модель с популяцией колоколовидных кубков в качестве единственного источника работает, есть и модели двух источников включающие 60% представителей культуры шаровидных амфор с территории Украины и 40% степных популяций.
Авторы также исследовали возможную связь генома поздней бронзы их Вертебы и киммерийцами, которые обосновались на территории современной Украины около 1000 г. до н. э. Однако геном из Вертебы оказался значительно ближе к носителям культуры колоколовидных кубков, чем к киммерийцам.
Анализ близкого родства в изученных геномах
Размеры гомозиготных участков ДНК указывают, что обитатели Вертебы жили в довольно многочисленных популяциях.
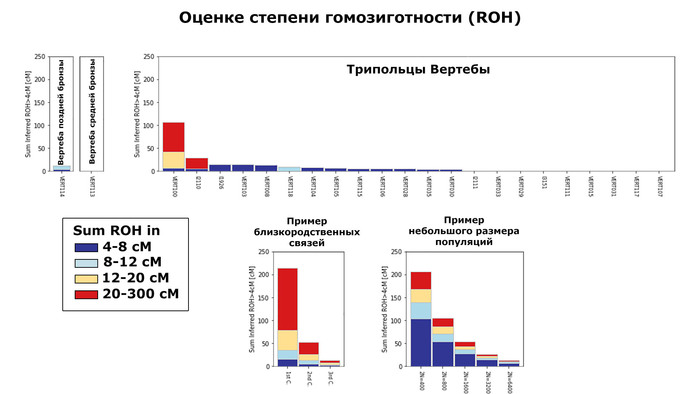
Исключением стал образец VERT-100, предки которого, вероятно, были родственниками.
Фенотипические и биохимические особенности
Ни один из проанализированных людей пещеры Вертеба не обладал толерантностью к лактозе.
Лишь два человека, видимо, были темноглазыми, но остальные обладали ДНК-маркером rs12913832, связанным с голубым цветом глаз.
Обсуждение
Генетический анализ 20 человек из пещеры Вертеба, которые датируются четвертым, вторым и первым тысячелетиями до нашей эры, выявил важные генетические изменения как в раннем бронзовом веке, так и в позднем. Эти датировки подтверждены радиоуглеродным датированием образцов 8 человек, поскольку пещера Вертеба многократно использовалась человеком, начиная с мезолита.
Предыдущие анализы митохондриальной ДНК представителей археологической общности Триполье-Кукутень показали их близкое родство по материнской линии с популяциями раннего неолита: гаплогруппы H, HV, T, V, J и K. И в новом анализе, за исключением двух человек с гаплогруппой U5a, все остальные 18 человек, имеют гаплогруппы, сходные с неолитическим населением Центральной Европы. Это разнообразие мтДНК резко контрастирует с населением более ранних неземледельческих неолитических памятников на территории Украины, которые имеют только гаплогруппу U, указывающую на преемственность с мезолитическими охотниками-собирателями.
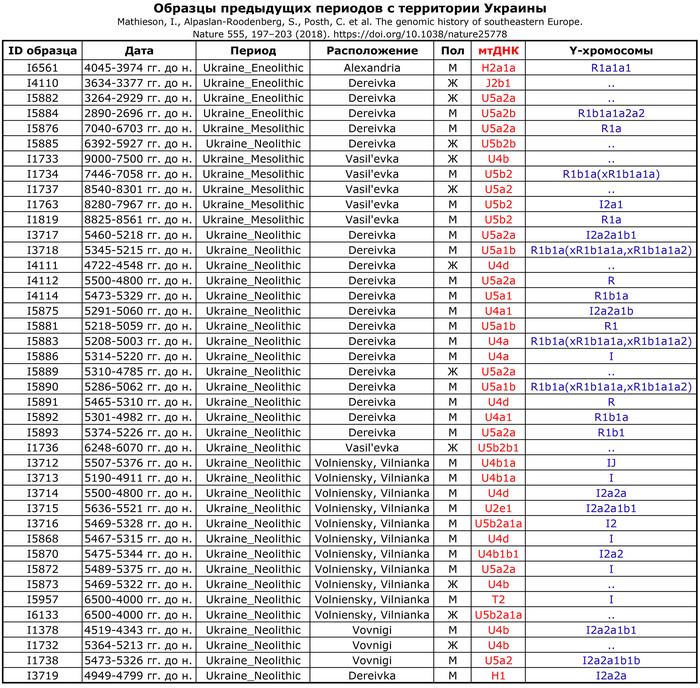
Mathieson, I., Alpaslan-Roodenberg, S., Posth, C. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018). doi.org/10.1038/nature25778Разнообразие гаплогрупп мтДНК предполагает, что местное население в значительной степени было заменено населением, связанным с трипольской культурой. Большинство представителей Вертебы имеют Y-гаплогруппу G2a2, которая широко распространена среди европейцев эпохи неолита, связанных с Анатолией.
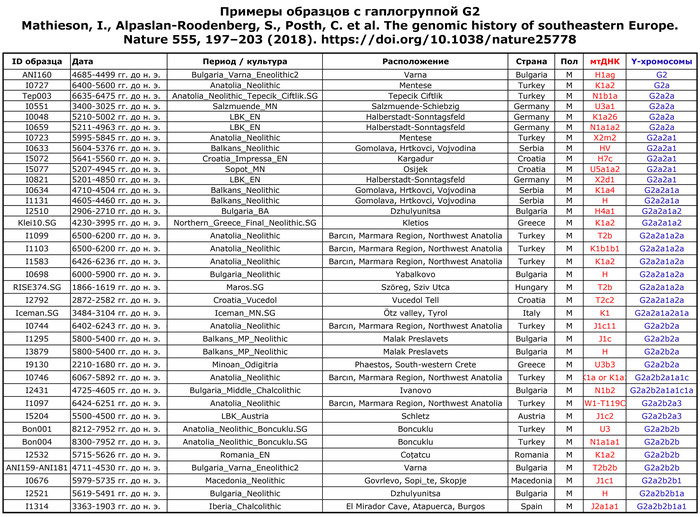
Другие выявленные гаплогруппы - C1 и I2 - также обнаружены среди европейских неолитических популяций.

Это указывает на миграцию и мужчин, и женщин, что контрастирует со степными миграциями во время бронзового века.
Данные популяционной генетики показывают, что люди, похороненные в пещере Вертеба в эпоху позднего энеолита (3790-3535 гг. до н. э.), генетически похожи на другие, ранее проанализированные, геномы из Молдовы. Эти наблюдения в целом предполагают, что люди энеолита археологической общности Триполье-Кукутень произошли от того же или близкородственного населения, которое распространило неолит по большей части Европы (в случае с трипольцами конкретно указывая на баденцев из Венгрии), и практически без признаков метисации с более ранним мезолитическим или неолитическим населением с территории современной Украины, связанным с охотниками-собирателями.
Фактически, большинство трипольцев могут моделироваться энеолитическими популяциями из Европы, имеющими степное происхождение, однако четверо из 20 человек могут быть смоделированы как трипольцы из Молдовы.
Результаты моделирования (qpAdm) предполагают, что существовали различия в составе предков трипольцев, которые могут быть связаны с долей охотников и собирателей, хотя, как отмечалось выше, эта изменчивость недостаточно существенна, чтобы разделить людей с разных участков на разные популяции.
При этом источником генетического пласта, унаследованного от охотников-собирателей у трипольцев, являются в основном западные охотники-собиратели. Значительная часть предков западных охотников-собирателей, обнаруженная у трипольцев (до 18%), может быть связана с увеличением доли этого компонента и в других популяциях среднего неолита Центральной Европы, вероятно, из-за смешения с популяциями на западе, которые уже имели более высокий компонент западных охотников-собирателей, полученный от неолитических групп, связанных с Анатолией до возникновения археологической общности Триполье-Кукутень.
Это также указывает на то, что неолитические популяции охотников-собирателей на территории Украины не внесли большого вклада в происхождение трипольцев. Кроме того, как и в Молдове, выявлен генетический вклад степного происхождения, но в Вертебе он ниже. Это может быть связано с их разными датировками, если предполагать непрерывный импульс движения с востока на запад, постепенно увеличивающий связь с ямной культурой в течение четвертого тысячелетия до н. э.
А вот геном из среднего бронзового века (VERT-113, 1960-1770 гг. до н. э.) значительно отличается от более ранних трипольцев: для него характерен больший вклад кавказских охотников-собирателей/ямной и восточных охотников-собирателей. Результаты (qpAdm) предполагают связь между геномом VERT-113 и представителями культуры шнуровой керамики из Польши. Кроме того, это единственный геном, который ближе к неолиту Украины, чем к западным охотникам и собирателям. Это позволяет предположить, что популяция, возникшая во втором тысячелетии до н. э., возможно, имела общие черты с популяциями неолита на территории Украины.
Интересно, что у генома поздней бронзы (VERT-114) нет значимой связи с геномом средней бронзы (VERT-113). Геном VERT-114 предполагает связь с культурой колоколовидных кубков, несмотря на то, что этот человек почти на 2000 лет моложе конца этого надкультурного феномена, и с датировкой ближе к киммерийцам или скифам. Это подтверждает данные, приведенные в более ранних работах, о притоке западного населения в степь в конце бронзового века.
Однако для понимания более полной картины необходимо получить ещё больше образцов разных периодов из этого региона.
Результаты новой работы имеют важное значение для понимания процесса неолитизации в Восточной Европе. Поскольку население археологической общности Триполье-Кукутень распространилось с современных территорий Румынии и Молдовы в лесостепные районы Западной и Центральной Украины, оно должно было вступить в контакт с населением, связанным с местной буго-днестровской культурой, чья система жизнеобеспечения была сосредоточена главным образом на охоте, рыбалке и собирательстве. Эта группа, вероятно, представляла собой потомков мезолитических охотников и собирателей. Анализ геномов населения пещеры Вертеба предполагает, что местные мезолитические охотники-собиратели не внесли существенного вклада в генофонд более поздних трипольцев. Иными словами, процесс неолитизации в Западной Украине был в большей степени результатом миграции (демической диффузии), а не принятия местными жителями практики земледелия (аккультурации).
Результаты также подтверждают гипотезу длительно существовавшей границы между оседлыми земледельцами лесостепной зоны и соседними кочевыми скотоводами из Причерноморской степи. Эта граница характеризуется резкими контрастами в материальной культуре и способах жизнеобеспечения и, вероятно, сохранялась благодаря этим факторам и языковым различиям. Отсутствие метисации на этой культурной границе является ключом к пониманию контекста, в рамках которого произошла миграция представителей ямной культуры.
В целом результаты показывают, что пещера Вертеба представляет собой важный погребальный комплекс, соединяющий Восток и Запад. Генетическая структура представителей археологической общности Триполье-Кукутень включает предковые компоненты, связанные как с более ранними западными охотниками-собирателями, так и с земледельцами Ближнего Востока, а также те, которые генетически отличаются от предков молдавских представителей этой общности.
Отсутствие местных предков, связанных с охотниками-собирателями эпохи неолита на территории современной Украины, предполагает, что эти земледельцы в основном заменили местных охотников-собирателей и не смешивались с соседними степными популяциями. Кроме того, в течение бронзового века пещера Вертеба использовалась последовательными волнами кочевников-скотоводов с востока, которые в конечном итоге принесли значительные генетические, а также культурные изменения в Европу и в конечном итоге смешались с местными потомками населения трипольской культуры.
Дополнительные генетические данные из этих и более поздних временных периодов позволят дополнить картину, а также ответить на вопросы хронологии и, возможно, указать, причины краха трипольской культуры.
Источник: Gelabert, P., Schmidt, R.W., Fernandes, D.M. et al. Genomes from Verteba cave suggest diversity within the Trypillians in Ukraine. Sci Rep 12, 7242 (2022). doi.org/10.1038/s41598-022-11117-8
3 200 000 лет назад обезъянка Люси упала с дерева разбилась насмерть.
( прим: чего-то я подзабыл, что мало кто знает это на самом деле, хотя некоторые этот факт считает баяном )
Недавнее время, 1975 год.
Людми́ла Ива́новна Тури́щева — советская гимнастка. Четырёхкратная олимпийская чемпионка, многократная чемпионка мира и Европы. Заслуженный мастер спорта СССР
Ну и петля Корбут, чемпионки мира:
Фантазия, что ты делаешь, прекрати








