



ИСТОРИЯ В ЛИЦАХ: «ЦЕНТР САДОВОДСТВА: ПО ПУТИ ПРОГРЕССА И ИННОВАЦИЙ»
Фильм из цикла «История в лицах» рассказывает о достижениях Федерального научного селекционно-технологического центра садоводства и питомниководства. Многопрофильное научное учреждение, основанное в тридцатых годах прошлого века, сегодня динамично развивается по пути прогресса и инноваций. В лабораториях, садах, теплицах, на полях сотрудники ФГБНУ ФНЦ Садоводства рассказывают об особенностях своей работы и о перспективах развития отрасли. Телекомпания Ирины Храбровой "Х-ТВ", выпуск 2023 г. Контакты: e-mail: x-tv.info@mail.ru
Показать полностью


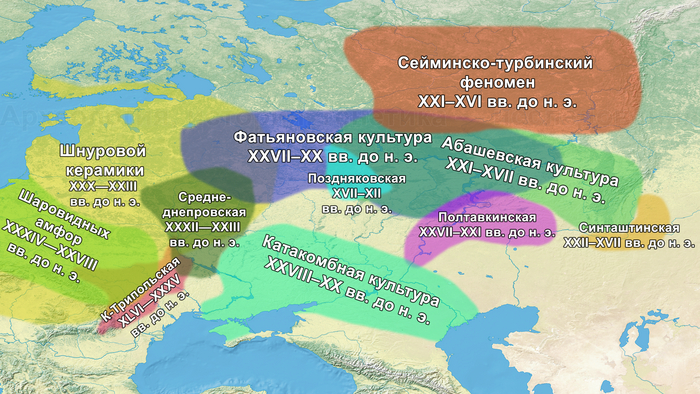
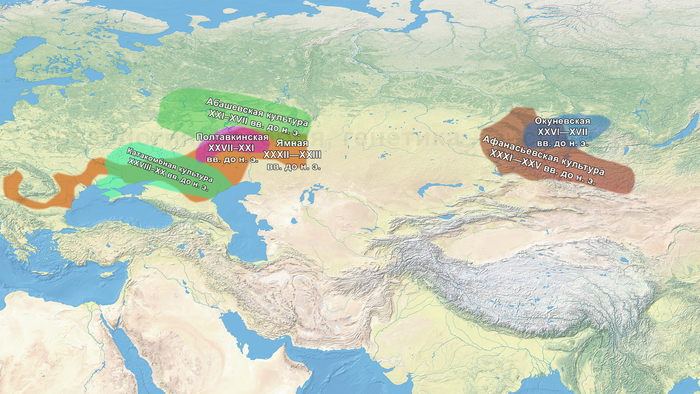
ДНК носителей фатьяновской и абашевской культур. Миграции на Русской равнине в бронзовом веке
Вступление
Приветствую! Сегодня речь пойдёт о научной работе, авторы которой рассматривают фундаментальный вопрос о происхождении и особенностях миграций носителей фатьяновской и средневолжской абашевской культур. Цель работы заключалась в том, чтобы на основании новых лабораторных данных о древней ДНК рассмотреть гипотезы о пришлом происхождении палеопопуляций фатьяновской и средневолжской абашевской археологических культур на Русской равнине.
В исследовании были проанализированы образцы 25 человек, относящихся к фатьяновской культуре (20 образцов было получено от захороненных в пределах Волосово-Даниловского могильника, два из раскопок в Никульцино и по одному из Воронково, а также Голузинского и Горицкого могильников). Стоит отметить, что Волосово-Даниловский могильник — наиболее репрезентативный погребальный комплекс фатьяновской археологической культуры.
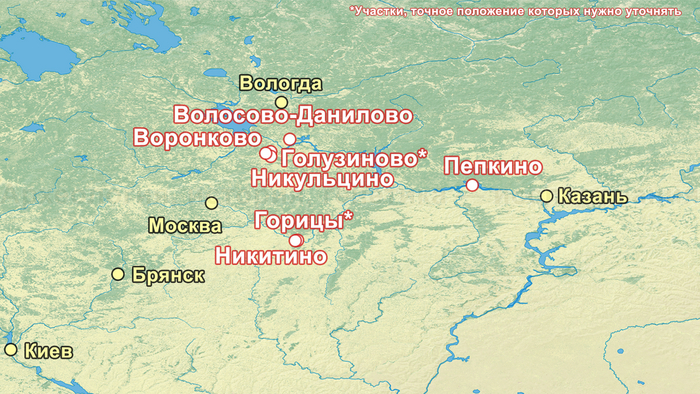
Помимо этого, впервые был проведен палеогенетический анализ 14 человек, относящихся к абашевской культуре (тринадцати человек из Пепкинского кургана и одного из Старшего Никитинского могильника). Пепкинский курган, открытый у деревни Пепкино, представляет собой коллективное захоронение группы молодых мужчин, не менее 27, средневолжской абашевской археологической культуры, погибших в бою.

Кстати, для исследования авторы работы намеренно отбирали представителей мужского пола разного возраста, так как для них возможен анализ однородительских маркеров как по мужской, так и по женской линиям.
Здесь сразу хотелось бы выразить благодарность авторам работы за их вклад в наше понимание прошлого.
Введение
Обнаружение в геномах европейских групп компонента восточных охотников и собирателей, соотносимого с генетическим наследием населения ямной археологической культуры, в своё время распространенной на обширных территориях евразийских степей, стало одним из наиболее спорных вопросов в российской археологии.

И хотя между распространением культуры и генов нельзя ставить знак равенства, генетический вклад от носителей ямной культуры зафиксирован во многих регионах Европы. Наличие этого степного генетического сигнала отличает представителей культур шнуровой керамики и колоколовидных кубков, а также более поздних жителей Европы эпохи бронзы от ранних неолитических земледельцев этого региона. При этом на территории европейской части современной России подобные генетические исследования до недавнего времени отсутствовали, пока в 2021 году не была опубликована работа, обзор которой также есть на канале, под названием: Генетическая история населения запада России при переходе от каменного века к бронзовому. В этом, более раннем, исследовании авторы пришли к выводу о зарождении носителей фатьяновской культуры благодаря миграции смешанного населения, потомков носителей ямной культуры и европейских ранних земледельцев.
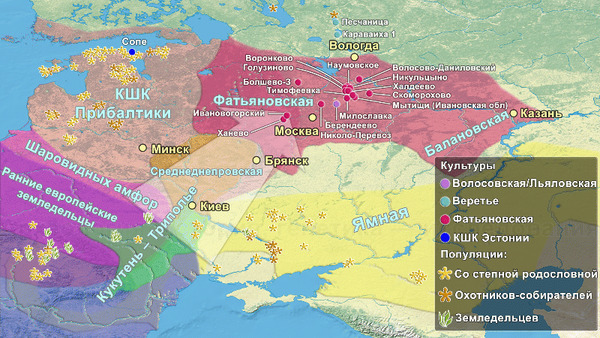
Тогда авторы сообщали, что в моделях только две популяции, а именно из культуры шаровидных амфор Украины и Польши, а также трипольской культуры Украины, оказались потенциальными источниками для смешения со степными скотоводами с генетическими профилями как у представителей ямной культуры Самары или Калмыкии, для образования генетической структуры популяции фатьяновцев. Также была отмечена близость этих людей с представителями культуры шнуровой керамики.
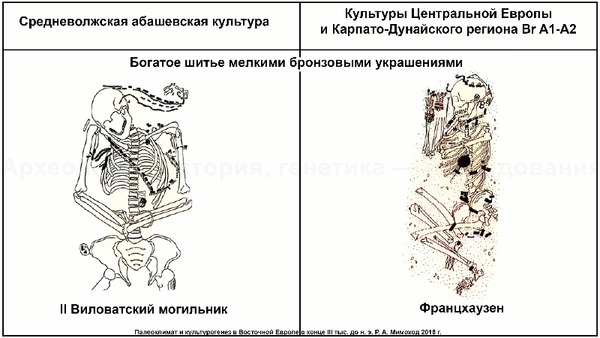
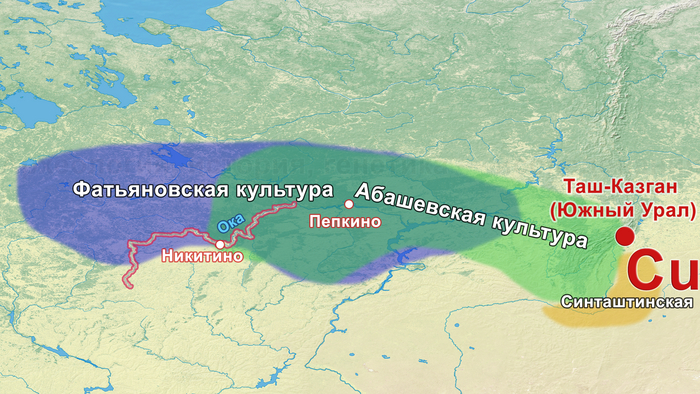
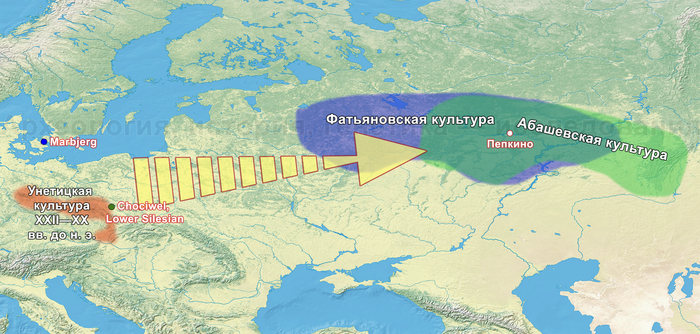
На основании данных археологии в последние годы уже высказывались предположения, что на территории Восточно-Европейской равнины в конце III тыс. до н. э. происходили масштабные миграции, с первоначальным импульсом из центральноевропейского региона. В пользу этого довода говорит схожесть керамического комплекса средневолжской абашевской культуры, следы которой выявлены непосредственно в Поволжье на территориях Брянской, Московской и Ярославской областей, с керамикой поздней фазы культуры колоколовидных кубков в Южной Германии.
Результаты
Анализ Y-хромосомы
По результатам микросателлитных исследований и анализа Y-хромосомы по методу секвенирования нового поколения (NGS) для образцов, относящихся к фатьяновской культуре, получена единственная гаплогруппа R1a (Z93).
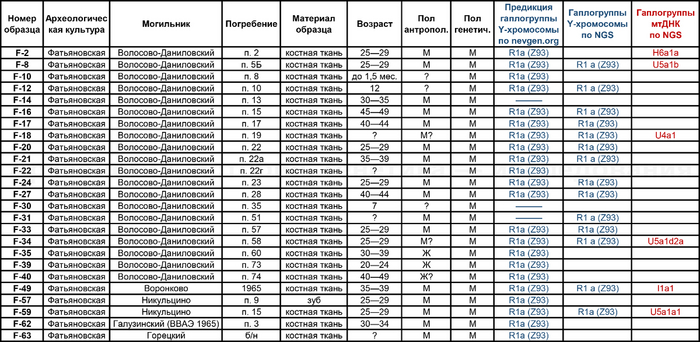
Вероятно, такая однородность гаплогрупп Y-хромосомы связана с эффектами основателя и дрейфа генов в изолированной древней группе неместного происхождения.
Эффект основателя — вариант генетического дрейфа, при котором снижение и смещение генетического разнообразия происходит из-за заселения малым количеством представителей вида новой географической территории.
Дрейф генов — Изменение частот аллелей и генотипов в популяции, имеющее случайный характер. Такие случайные отклонения в частотах аллелей происходят из поколения в поколение, особенно при небольшом эффективном размере популяции.
На данный момент линии гаплогруппы R1a1a1b2 Z93 с заметной частотой встречаются в Южной Сибири среди алтайцев и хакасов (>30 %), также сравнительно небольшие частоты встречаются среди татар Поволжья (5 %), в Кыргызстане (6 %) и во всех популяциях Ирана (1—8 %).
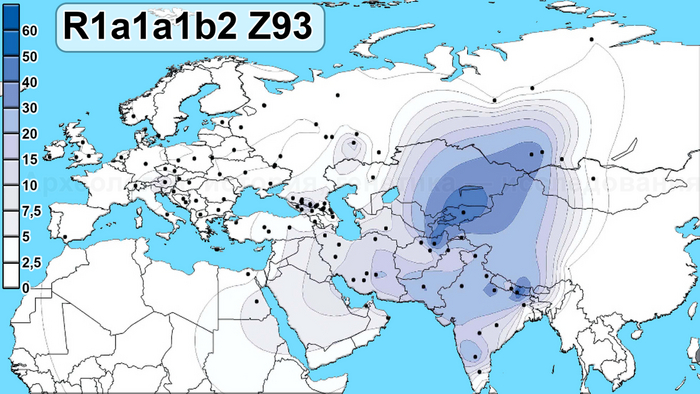
Среди археологических культур указанная гаплогруппа обнаружена, в частности, в сагсайской культуре XV—XII вв. до н. э. Субветвь этой гаплогруппы R1a1a1b2a Z94 представлена рядом образцов, относящихся к археологическим культурам средней бронзы центральной и западной части степного коридора Евразии — андроновской, полтавкинской, синташтинской и также сагсайской (XX—XV вв. до н. э.).
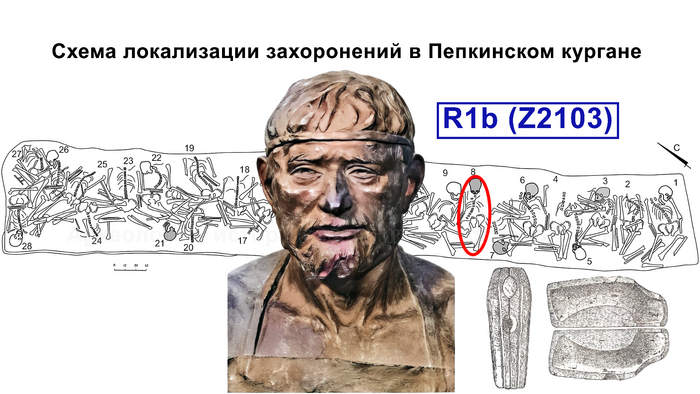
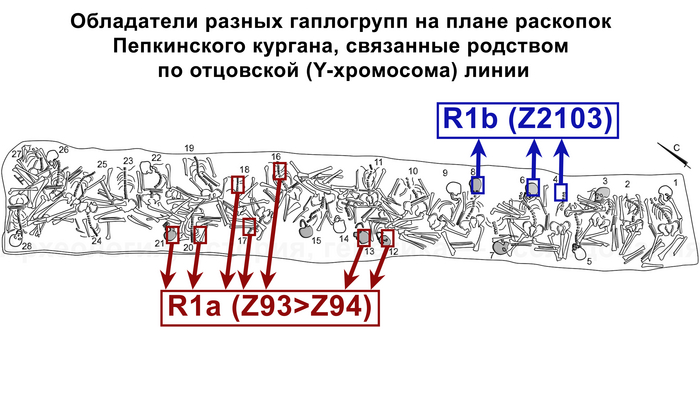
А вот для образцов абашевской культуры полученные результаты менее однородны. Для большинства образцов из Пепкинского кургана определена гаплогруппа R1a (Z93>Z94), однако у троих мужчин с этого участка была гаплогруппа R1b (Z2103).
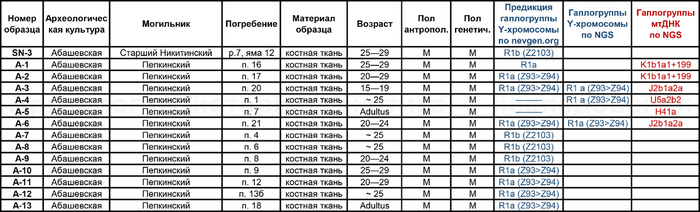
Примечательно, что для абашевца захороненного на значительном удалении от Пепкинского кургана в Старшем Никитинском могильнике, был получен достаточно полный гаплотип, также предсказывающий принадлежность к той же гаплогруппе R1b (Z2103).
Указанная гаплогруппа была выявлена у мужчин, относящихся к ямной, афанасьевской, полтавкинской, катакомбной и окуневской культурам III—II тыс. до н. э.
Микросателлитные исследования в палеогенетике являются важным инструментом, особенно при исследовании групповых захоронений, поскольку позволяют выявить близость с точки зрения происхождения по мужской линии, а иногда и прямого родства, когда гаплотипы по всем считанным STR локусам совпадают, при этом результат тем точнее, чем выше число считанных локусов. Такие совпадения были найдены как для фатьяновцев погребенных в Волосово-Даниловском могильнике, так и для абашевцев из Пепкинского кургана, а также Старшего Никитинского могильника.
Что указывает на возможное близкое родство по отцовской линии людей с совпадающими STR локусами. А у людей, которые продемонстрировали минимальные отличия в значениях аллелей в одном или двух локусах, общий предок по отцовской линии мог жить несколькими поколениями ранее.
Анализ митохондриальной ДНК
Материнские линии фатьяновцев были более разнообразны, чем отцовские. При этом основной гаплогруппой мтДНК была U с сублиниями — U5a1b, U5a1d2a и U5a1a1. А среди редких, так называемых минорных линий, определены H6a1a и I1a1
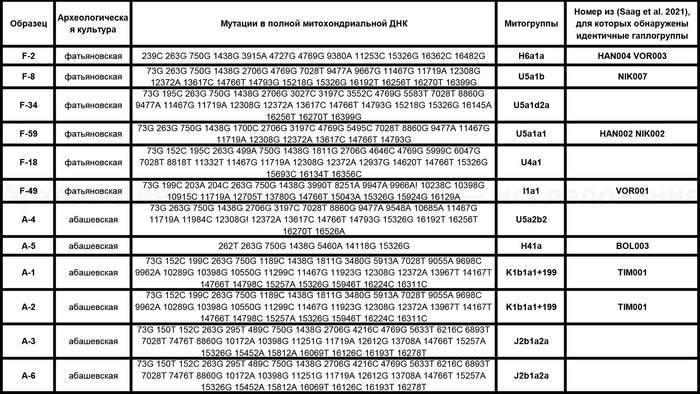
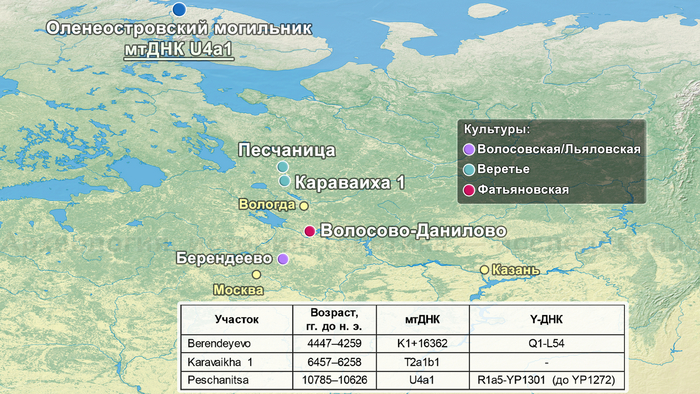
В большинстве линии гаплогрупп совпадают с задокументированными и в работе 2021 года, но с некоторыми дополнениями, как линии U5a1d2a и U4a1 у фатьяновцев из Волосово-Даниловского могильника. Однако в предыдущем исследовании линия U4a1 определена у одного из охотников-собирателей культуры Веретье эпохи мезолита, захороненного в могильнике Песчаница 1 на юго-западе Архангельской области. А ещё ранее эта линия была обнаружена у восточных и скандинавских охотников-собирателей. В частности, у охотников и собирателей эпохи неолита Большого Оленьего острова в Кольском заливе.
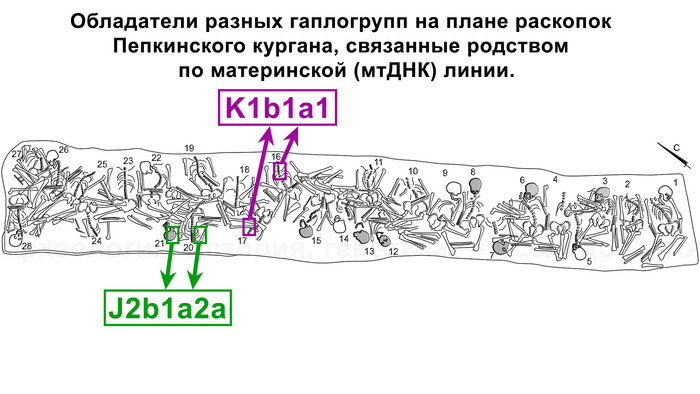
А что касается хронологически более поздних представителей абашевской культуры, то часть их линий, таких как U5a2b2, H41, ранее были обнаружены и у фатьяновцев. Однако неожиданностью для авторов стало наличие гаплогрупп J2b1a2a и K1b1a1+199 у абашевцев из Пепкинского кургана, ведь распространение этих линий связывают с неолитическими земледельцами. При этом пары с этими линиями имеют полностью совпадающий набор мутаций, что указывает на их родство по материнской линии.
Анализ аутосомных маркеров
Сравнительный анализ аутосомных генетических вариантов с использованием метода понижения размерности проводился для 19 образцов в сочетании с ранее опубликованными данными 75 человек. По результатам анализа большая часть исследованных в новой работе образцов фатьяновской и абашевской культур объединились между собой в одном кластере.
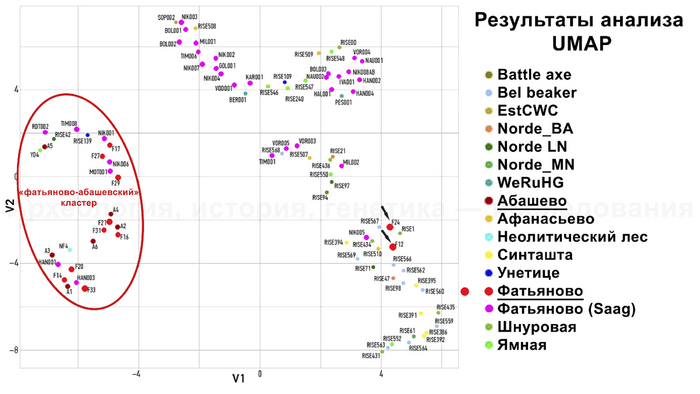
Туда же вошли некоторые фатьяновцы из предыдущей работы. Неожиданный результат был получен для двух фатьяновцев из Волосово-Даниловского могильника, которые оказались ближе к скоплению образцов из контекста различных археологических культур эпохи бронзы, а именно из культур шнуровой керамики, колоколовидных кубков, афанасьевской и синташтинской. Что указывает на генетическую близость жителей эпохи бронзы, несмотря на их разную хронологию и географию.
Обсуждение и выводы
В новой работе у представителей фатьяновской культуры не обнаружено других отцовских линий, кроме R1a (Z93), что, вероятно, связано с эффектом основателя (когда небольшая группа отделяется от первоначальной популяции и формирует новую) и с дрейфом генов (когда одна линия сохранилась из-за случайных событий).
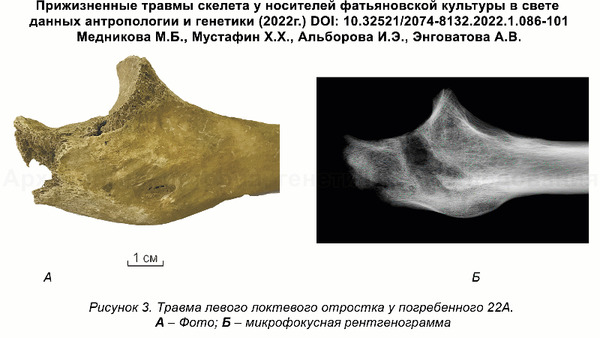
Это подтверждает миграцию группы представителей культуры шнуровой керамики из Центральной Европы и отсутствие связей с представителями местной волосовской культуры, что также упоминалась и в предыдущем исследовании, хотя его авторы использовали другие образцы и методы. Примечательно, что ранее, по результатам анализов травм левой руки в области локтевого сустава и кисти у мужчин из Волосово-Даниловского могильника, авторы исследования, опубликованного в 2022 году, пришли к выводу о схожей тактике боя у фатьяновцев Верхней Волги и шнуровиков Германии и Польши. Что служит независимым индикатором их принадлежности к единой культурной общности.
Помимо этого, совпадение митохондриальных гаплогрупп в обоих упомянутых генетических исследованиях также подтверждает гипотезу о неместном происхождении носителей фатьяновской культуры. При этом однообразный погребальный обряд фатьяновцев, различный для мужчин, женщин и детей, а также их низкое генетическое разнообразие, добавляют аргументов в пользу существования строгих правил внутри общества, которые можно объяснить его сплочённостью в связи с миграцией в иную среду. А учитывая некоторую близость по материнской линии захороненных в Волосово-Даниловском могильнике, с преобладанием ветвей гаплогруппы U, можно сделать вывод, что фатьяновцы переселялись семьями.

Датировка образцов из Волосово-Даниловского могильника указывает на его использование в течение более трёхсот лет, минимум с середины третьего тысячелетия до н. э. (интервал калиброванных дат: 2563-2037 гг. до н. э.).
При этом Волосово-Даниловский могильник всеми исследователями рассматривался как наиболее поздний.
А спустя несколько сотен лет после первого появления фатьяновцев возникла абашевская археологическая культура, в лесостепных районах среднего Поволжья и верхнего Дона, предположительно, основанная другими мигрантами. При этом Пепкинский курган, на котором были захоронены мужчины, погибшие в одном сражении, на данный момент является главным археологическим памятником средневолжской абашевской культуры. На первый взгляд это могли быть воины одного племени, потому как они захоронены в одной могиле и по схожему обряду, между 2130-1950 гг. до н. э.
Однако анализ древней ДНК выявил как минимум две группы разного происхождения. Экспертиза бронзовых изделий из Пепкинского кургана и Старшего Никитинского могильников показала, что по составу металла оба памятника относятся к единому этапу средневолжской абашевской культуры. А мышьяк и серебро в составе их металла, могут указывать на связь с рудными источниками Южного Урала. При этом для образца из Старшего Никитинского могильника была получена несколько более поздняя дата чем для Пепкинского кургана, отличающаяся на время жизни одного поколения абашевцев (3557 ± 31 BP).
Примечательно, что в Пепкинском кургане, один из абашевцев, погребенный с набором предметов кузнеца-бронзолитейщика с гаплогруппой Y-хромосомы R1b (Z2103), по данным изотопного анализа провел свои юные годы в геохимических условиях, которые соответствовали степным районам Южного Урала.
К тому же, известно о происхождении металла некоторых изделий, найденных в Пепкинском кургане, из Таш-Казганского месторождения меди.
При этом по соотношениям изотопов стронция в зубной и костной ткани другой группы мужчин из Пепкинского кургана, все они оказались «неместными» для территории современной республики Марий Эл, где они были похоронены. Примерно до 10 лет они жили в регионе соответствующему западному ареалу проживания абашевцев в бассейне Оки. Похожие значения характерны и для захороненных в пределах Старшего Никитинского могильника. Интересно, что абашевцы с гаплогруппой R1b(Z2103) были более мобильны, но при этом связаны достаточно тесным родством. Можно предположить, что именно эта группа в составе абашевского населения была тесно связана с металлургическим производством. Однако другая, более многочисленная часть мужчин из Пепкинского кургана с гаплогруппой R1a (Z93>Z94), очевидно, имела иное происхождение. Отцовские линии и аутосомные маркеры сближают их с фатьяновцами, прежде всего, из Волосово-Даниловского могильника. Однако неясно, связано ли происхождение абашевцев с фатьяновцами, что в целом не подтверждается археологически, или же с новой миграцией из Европы спустя несколько веков после миграции предков фатьяновцев. И хотя митохондриальные линии поддерживают обе гипотезы, отдельные линии обнаружение у некоторых людей из Пепкинского кургана, такие как J2b1a2a и K1b1a1+199, скорее, свидетельствуют о недавней миграции из Европы, где генетический след неолитических земледельцев был очень заметен.
Ранее была предложена гипотеза общей энеолитической подосновы населения абашевской культуры и культур шнуровой керамики.
А по черепам из Пепкинского кургана антропологами была отмечена высокая изменчивость у абашевских мужчин.
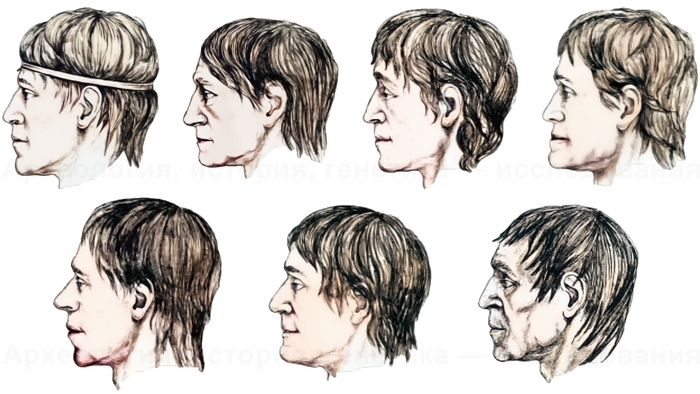
Графические реконструкции мужчин погребенных в Пепкинском кургане (Реконструкции Г. В. Лебединской).
Но, в целом, метод главных компонент выявил сближение выборки мужчин, погребенных в Пепкинском кургане, с южноевропеоидными формами, ранее отнесенными к грацильным представителям средиземноморской расы. Что больше свидетельствует в пользу недавней миграции из Европы.
На графике с использованием метода понижения размерности (UMAP) люди из Волосово-Даниловского могильника сближаются со второй, более многочисленной группой из Пепкинского кургана, что может указывать на преемственность. Однако абашевец из погребения 7 Пепкинского кургана близок и к мужчине унетицкой культуры из Польши, а также к представителю позднего неолита из Марбьерга в Дании, которые жили примерно в то же время, что и он. А это уже аргумент в пользу недавней миграции.
У носителей фатьяновской культуры, на протяжении всего периода существования Волосово-Даниловского могильника, наблюдается достаточно стандартный погребальный обряд, который роднит их с культурами шнуровой керамики Европы, чего нельзя сказать для набора инвентаря поселений фатьяновцев. Но обряд свидетельствует о формировании могильника представителями пришлого населения, несущего традиции европейских групп шнуровой керамики.
А вот вопрос о происхождении представителей средневолжской абашевской культуры решается не столь однозначно. В ее составе присутствуют две составляющие — с гаплогруппами R1a (Z93>Z94) и R1b (Z2103).
Для носителей линии R1a (Z93>Z94) типично захоронение в могиле рядом близких родственников по материнской линии (причем здесь выделяются пары с европейскими «раннеземледельческими» митохондриальными гаплогруппами: как J2b1a2a из погребений 20, 21 и K1b1a1+199 из погребений 16,17).
В то время как захороненных рядом мужчин с гаплогруппой Y-хромосомы R1b (Z2103) сближает родство по отцовской линии. Это указывает на то, что система родства в группах разного происхождения отличалась.
Примечательно, что абашевцы из Пепкинского кургана с гаплогруппой R1a (Z93>Z94), в том числе, связанные тесным родством, всегда имели с собой костяные поделки и украшения, в отличие от носителей линии R1b (Z2103), у отдельных носителей которой прослеживаются связи со степными районами Южного Урала.
Однако решение вопроса о происхождении двух составляющих абашевского населения лежит в плоскости будущих исследований.
Источник: А. В. Энговатова, И. Э. Альборова, Х. Х. Мустафин, В. Ю. Луньков, Ю. В. Лунькова (Москва, Россия), А. А. Канапин, А. А. Самсонова (Санкт-Петербург, Россия), М. Б. Медникова (Москва, Россия) Древняя ДНК носителей фатьяновской и абашевской культур (к вопросу о миграциях населения эпохи бронзы в лесной полосе на Русской равнине) doi.org/10.55086/sp232207228
Показать полностью
23

Животные с синдромом Дауна

Тигр Кенни
Из всех "животных с синдромом Дауна", наиболее известен Кенни- редкий белый тигр, спасенный от недобросовестного заводчика в 2002 году заповедником дикой природы Терпентин-Крик в Арканзасе, где он жил до своей смерти в 2008 году.
Белые тигры чрезвычайно редки, но помимо своей красивой белой шерсти, Кенни имел различные уродства, включая аномально короткую и широкую морду. Увидев Кенни на фото, многие пользователи соцсетей пришли к выводу, что у него синдром Дауна. На самом деле это ложь.
Уродства Кенни являются результатом нескольких поколений инбридинга, а не хромосомной мутацией. Поскольку белые тигры чрезвычайно редки в природе, но особенно желанны из-за своего уникального меха, большинство из них, живущих сегодня, являются результатом агрессивных программ разведения, интенсивно использующих инбридинг между белыми тиграми.
Кенни далеко не единственный представитель семейства кошачьих, которого ложно рекламируют, как страдающего синдромом Дауна.

Котенок Отто стал интернет-сенсацией в своей родной Турции в 2014 году. Когда крошечный кот скончался в возрасте чуть более двух месяцев, интернет-издания сообщили, что его ранняя смерть была связана с последствиями синдрома Дауна. Однако здесь имеется одна маленькая неувязка: у кошек любого вида, как и практически у всех животных, синдром Дауна не развивается.
Объяснение:
Каждая клетка здорового человека содержит 23 пары хромосом. Синдром Дауна возникает у людей из-за генетической мутации, в результате которой образуются три копии 21-й хромосомы вместо двух. Генетический состав клеток животных кардинально отличается от человеческого, поэтому утроение одной и той же хромосомы не будет иметь подобного эффекта. Кроме всего прочего, у многих животных вообще нет 21-й хромосомы: кошки, к примеру, имеют только 19 пар хромосом.
"Животные с синдромом Дауна", обнаруживаемые сегодня по всему Интернету, на самом деле имеют лишь внешние признаки болезни, подобно людям с синдромом Дауна. Широко расставленные глаза тигра Кенни и его короткая морда были вызваны инбридингом, аномальные черты лица котенка Отто никогда не были окончательно объяснены, но могли быть вызваны генетической мутацией, дефицитом гормонов, и так далее.

Лев ошибочно причисляемый к животным с синдромом Дауна.
В то время как представления о животных с синдромом Дауна являются мифом, человекообразные обезьяны, похоже, единственная группа животных, которые иногда демонстрируют генетические дефекты, по крайней мере сравнимые с синдромом Дауна. У человекообразных обезьян 24 пары хромосом, а не 23, как у людей. И у некоторых обезьян была диагностирована дополнительная копия 22-й хромосомы, которая в целом похожа на 21-ю хромосому у людей.
Первый подтвержденный случай трисомии 22 у шимпанзе был зафиксирован в 1969 году. Однако та обезьяна прожила меньше года. А недавно японские исследователи подтвердили второй случай шимпанзе с этим хромосомным дефектом.

Шимпанзе по имени Канако с подтвержденной трисомией по 22 хромосоме.
Канако родилась в неволе в 1992 году в заповеднике, который в 2011 году был передан Киотскому университету и назван заповедником Кумамото, Центром исследования дикой природы. Когда ей было 156 дней, Канако разлучили с матерью Канаэ, после чего её воспитанием занимался исключительно персонал заповедника.
В течение первого года Канако часто страдала от кашля, простуды, лихорадки, диареи и отека вокруг правого глаза, хотя эти симптомы не редкость у молодых шимпанзе. С раннего возраста у неё наблюдалась задержка роста, врожденный порок сердца и недоразвитые зубы. Проблемы с сердцем были обнаружены в 2014 году во время обычного медицинского осмотра. Эхокардиограмма показала, что у Канако дефект межпредсердной перегородки - отверстие в стенке, разделяющей две верхние камеры сердца.
В течение первого года жизни у Канако развилась катаракта и она полностью ослепла к тому времени, когда ей исполнилось 7 лет. Также у Канако косоглазие и заболевание, вызывающее постепенное истончение роговицы. Похожие проблемы со здоровьем иногда встречаются у людей с синдромом Дауна.
Слепота Канако затрудняет общение с другими шимпанзе, но не делает его полностью невозможным. По началу Канако жила отдельно от других шимпанзе, чтобы избежать агрессии с их стороны, но впоследствии ей дали возможность раз в месяц общаться с другой самкой шимпанзе по имени Роман, всегда спокойной и дружелюбной по отношению к другим обезьянам.

Канако (справа) и Роман
В 2015 году сообщалось, что в Танзании обнаружен детёныш шимпанзе с синдромом Дауна, однако случай не подтвержден, поскольку не удалось провести хромосомный тест.
"Трудно оценить вероятность редкого события, используя небольшую популяцию. Но учитывая, что около 500 шимпанзе родились в неволе в Японии, вероятность этой аутосомной трисомии у шимпанзе может быть сравнима с вероятностью трисомии 21 у людей, которая встречается в 1 из 600 рождений", — сказал Хитоши Хирата, ведущий автор исследования и директор заповедника Кумамото.
Вывод: на самом деле животных с синдромом Дауна не существует.
Вольный перевод статьей:
Показать полностью
4
2


Пришло время Грегора Менделя
До конца семидесятых годов XX века генетика в основном двигалась по пути, проложенному Грегором Менделем. Только расшифровка нуклеиновых оснований в молекулах ДНК, наследственность позволили опираться на физико-химические методы, не анализ результатов гибридизации. Исследования Менделя 1865 года были переоткрыты только в 1900 году, принесли посмертную славу и послужили началом новой науки генетики.

1 Portrait of Gregor Johann Mendel by Daniel J. Fairbanks
Ещё в начальной школе Грегор обнаружил математические способности и по настоянию учителей продолжил образование в гимназии города Опава. Денег на обучение не хватало. С большим трудом удалось завершить гимназический курс, а помощь младшей сестры Тереза, которая пожертвовала своим приданным, помогли проучиться только некоторое время на курсах по подготовке в университет.
Профессор математики Франц посоветовал Менделю вступить в августинский монастырь города Брно, где аббат Кирилл Напп был человеком широких взглядов, поощрявший занятия наукой. В 1843 году Мендель поступил в этот монастырь и получил имя Грегор (при рождении ему было дано имя Иоганн). Через четыре года монастырь направил двадцатипятилетнего монаха Менделя учителем в среднюю школу. Затем с 1851 по 1853 год он изучал естественные науки, особенно физику, в Венском университете, после чего стал преподавателем физики и естествознания в реальном училище города Брно.
В 1856 году Мендель начал классическую работу по гибридизации и анализу наследования признаков, удачно выбрал для своих экспериментов горох. Он высевал горох на отведенных ему 2,5 сотки в течение восьми лет. Мендель провёл десять тысяч опытов с разными по окрасу цветков и внешнему виду семян. Для проверки закономерности полученных в результате скрещиваний передачи лишь одного признака формы семян («гладкие — морщинистые»), он подверг анализу 7324 горошины рассматривая каждое через лупу, сравнивая их форму и делая записи.
2 Моногенетические признаки, которые Грегор Иоганн Мендель исследовал в своих экспериментах с горохом
Благодаря большой набранной статистики удалось сделать выводы и увидеть целостную картину исследования — буквенно «помеченные» альтернативные признаки открыли ему, что определенные типы скрещивания в разном потомстве дают соотношение 3:1, 1:1, или 1:2:1.
Грегор Мендель не был признан современниками. Слишком уж простой казалась им предложенная схема, которая могла объяснить и обосновать практическими данными сложные явления. Поколебать господствующее тогда представление о пирамиде эволюции не удалось. Мендель говорил: «Мое время еще придет». Эти слова начертаны на его памятнике, установленном перед монастырским садиком, где он ставил свои опыты.
Интересно? Еще можно почитать
1) «Другая часть живого вещества исповедует иную стратегию. Эти виды пластичны, изменчивы. И творят они самих себя, черпая новую информацию благодаря активным взаимодействиям между собой и со средой обитания. Каким образом происходит обогащение информацией, во многом остаётся загадкой. Это тема особая, непростая и для наших целей второстепенная.
Важно отметить сам факт поисковой стратегии у значительной части растений и животных. Среди них ярко проявляется стремление к разнообразию, неожиданным решениям, свободе творчества».
Этот и еще 190 материалов VIKENT.RU по теме Эволюционные идеи
2) Видео: ТВОРЧЕСКАЯ ЭКСПАНСИЯ
3) Видео: # КАК СТАТЬ НЕУДАЧНИКОМ с САМОМНЕНИЕМ ?..
Источники
Грегор Иоганн Мендель на портале VIKENT.RU https://vikent.ru/author/423/
Самин Д. К., 100 великих ученых. — М.: Вече, 2004. — 592 с. (100 великих) с.265-270
Фото:
Показать полностью
2
2

А что знаете вы о лососе? Рыбка-то эта - загадочная!
Познавательный и очень обаятельный фильм "Загадка лососей" рассказывает о работе учёных Института биологии моря Дальневосточного научного центра АН СССР по изучению генетической структуры лососевых рыб.
Дальтелефильм 1976. Источник: канал на YouTube «Советское телевидение. Гостелерадиофонд России»


Генетика будущего
Девушку зовут Fatima Diame, родилась в Сенегале, выступает за Испанию.

Сможете найти на картинке цифру среди букв?
Справились? Тогда попробуйте пройти нашу новую игру на внимательность. Приз — награда в профиль на Пикабу: https://pikabu.ru/link/-oD8sjtmAi

Гаплогруппы Y-ДНК из Центральной Азии. Филогения Y-хромосом в свете новых данных
Центральная Азия расположена на пути расселения людей современной анатомии с Ближнего Востока в Северную Евразию, что отражено в геномах древних людей, таких как усть-ишимский человек, обитатели Янской стоянки на севере Якутии и стоянки Мальта в Иркутская область. При этом климатические изменения, связанные с последним ледниковым максимумом (LGM) существенно изменили распределение популяций древних людей в Центральной Азии и прилегающих регионах. Древние носители ботайской культуры благодаря предковым линиям древних северных евразийцев генетически связаны с представителями стоянок Мальта в районе озера Байкал и Афонтова гора у современного Красноярска. Последующие миграции земледельцев с Ближнего Востока и скотоводов из степи, дополнили разнообразие в регионе. Распространение на восток в Центральную Азию и восточно-евразийские степи индоевропейских популяций привело к возникновению энеолита и бронзового века. Известны довольно примечательные Бактрийско-Маргианский археологический комплекс и андроновская культурно-историческая общность, предковая, как считается, ираноязычным популяциям.

Реконструкции: www.ancestralwhispers.org/reconstructions
Карасукская культура возникла в Южной Сибири и распространилась по обширным территориям, дав начало ряду преемников. Позднее подъём скифов вызвал широкомасштабные миграции и примесь через евразийские степи, включая Центральную Азию.

Реконструкции: www.ancestralwhispers.org/reconstructions
В более поздний исторический период тенденция к расселению людей с запада на восток изменилась, юэчжи, усуни и эфталиты мигрировали с востока на запад и создали ряд государств в Центральной Азии. А хунну/гунны массово мигрировали на запад и значительно изменили этническую структуру в Центральной Азии и Восточной Европе. Позже древние тюркские и монгольские племена распространились на запад и рассеялись по всей Центральной Азии, Ближнему Востоку и Восточной Европе. При этом потомки вышеупомянутых и не упомянутых, в сильно сокращенной предыстории, древних людей в течение длительного исторического периода смешивались и формировали современные популяции в Центральной Азии.
Предыдущие генетические исследования современных людей пролили свет на происхождение и генетическую структуру популяций из Центральной Азии. А анализ древней ДНК останков различных исторических периодов выявил закономерности смешения и генетическую структуру древних людей, а также их вклад в генофонды современных популяций.
В целом отцовский, материнский и аутосомный генофонды большинства популяций этого региона характеризуются высоким разнообразием и примесью с популяциями Ближнего Востока, Европы, а также Южной, Восточной и Северной Азии. Также у популяций этого региона выявлена сильная племенная структура, которая привела к значительным различиям в доминирующих отцовских линиях между племенами. Однако внутри племён разнообразие коротких тандемных повторов на отцовских Y-хромосомах (Y-STR), как правило, невелико.
И хотя поток генов является обычным явлением, генетические взаимоотношения между популяциями по-прежнему тесно связаны с языком. Например, таджики генетически родственны другим индоевропейским популяциям, в то время как у кыргызов обнаружено много генетических компонентов из Сибири. Кроме того, были оценены последовательности Y-хромосом конкретных племен или семейных кланов для изучения формирования родственных популяций. Тем не менее, демографическая история населения Центральной Азии сложна, поэтому любые новые данные дополняют и уточняют наши знания об истории региона. Так и в новой работе авторы дополняют данные полноразмерными последовательностями Y-хромосом 187 человек. В выборку вошли казахи, туркмены, узбеки, таджики, кыргызы, каракалпаки, хазарейцы, карлуки, уйгуры и дунгане.
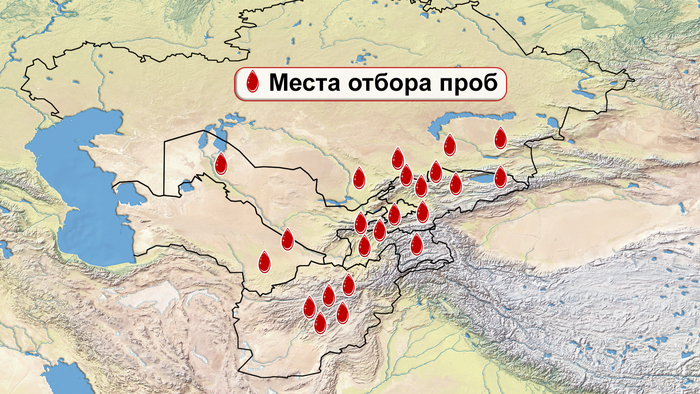
Основной целью авторов работы было выявление на основе полноразмерных последовательностей Y-хромосом общих закономерностей в отцовских генофондах. После чего идентифицировать отцовские линии, которые почти уникальны для какой-то конкретной популяции, и проанализировать их демографическую историю. В частности, исследователи оценили различные периоды генетического обмена и расхождения среди древних популяций в Центральной Азии.
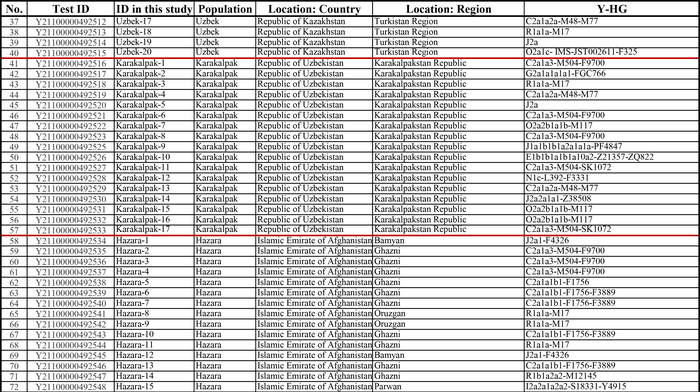
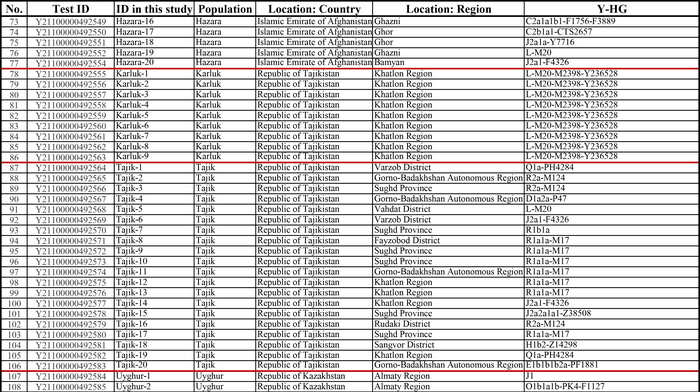
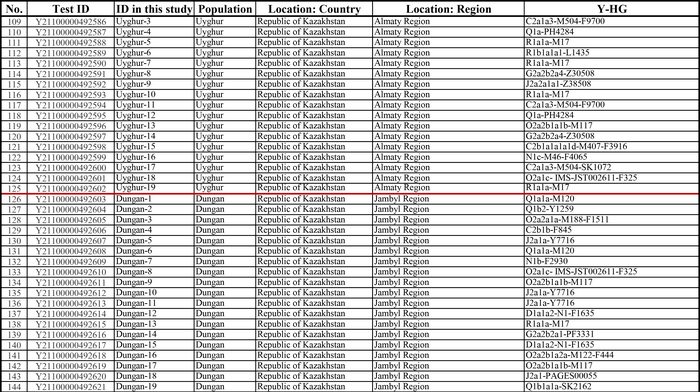
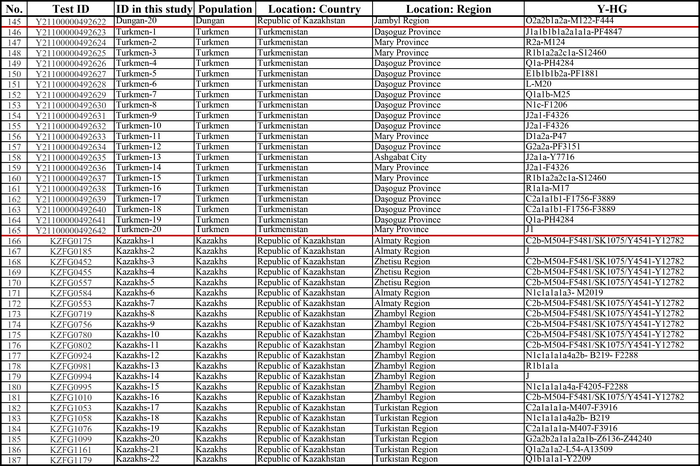
Результаты
Пересмотренное филогенетическое древо, основанное на Y-хромосомах популяций Центральной Азии
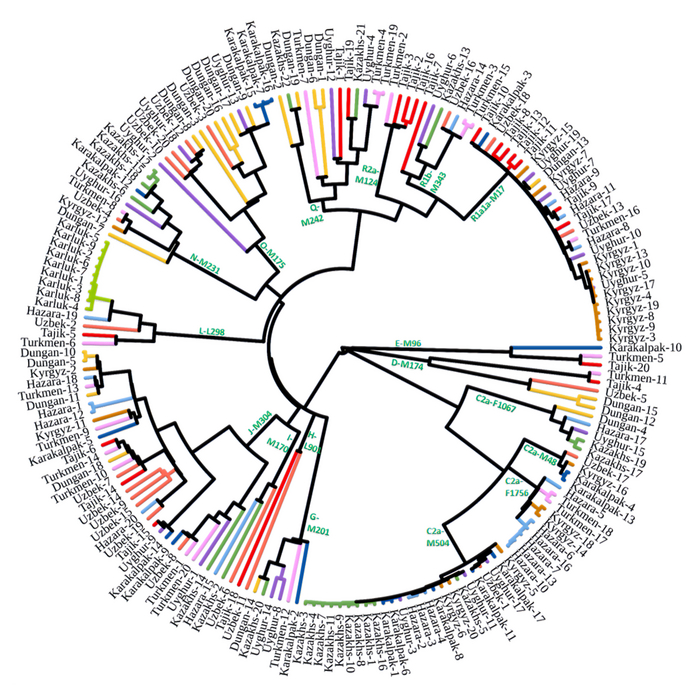
Гаплогруппа C2a1a1b1-F1756 была преобладающей отцовской линией древних племен дунху и сяньби, а появление этой линии у хазарейцев, кыргызов и туркмен указывает на то, что их предки по отцовской линии из Восточной Евразии. Гаплогруппа C2a1a2a-M48-F6379 имела высокую частоту в Младшем жузе казахов и ойратов — западных монголоязычных народов. Появление этой линии у каракалпаков свидетельствовало о близком родстве этого населения с казахами и древними монголами. Считается, что экспансия в Центральной Евразии С2а1а3- M504 напрямую связана с деятельностью древних монгольских племен в прошлом тысячелетии. В исследовании отмечена высокая частота этой линии в популяциях хазарейцев, каракалпаков, казахов, кыргызов и уйгуров, что согласуется с предполагаемым монгольским происхождением этих популяций из предыдущих исследований разных авторов. Линии N-M231 у казахов связаны с древнетюркскими племенами. А линии D-M174 и O-M175, обнаруженные в популяциях Центральной Азии, появились в результате смешения за последние 2000 лет с популяциями из Восточной Азии.
Как уже известно, гаплогруппы E-M96, G-M201, H-L901, I-M170 и L-M20 были широко распространены с низкой частотой в популяциях из Центральной Азии. Эти линии представляют собой смесь из Южной Азии, Ближнего Востока и Европы. Тем не менее, исследователи обнаружили особую сублинию L-M20 у 9 карлуков, а именно L1a2a-M2398-Y236528, которая около 1500 лет назад отделилась от линий, обнаруженных в Индии.
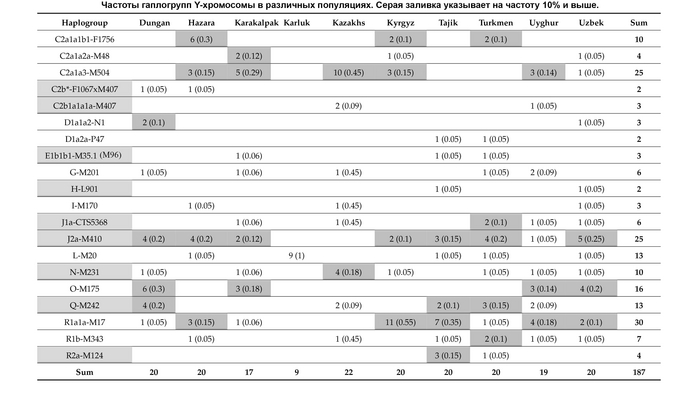
Предыдущие исследования показали, что Южная Сибирь, вероятно, является центром распространения гаплогруппы Q-M242, начавшейся около 30 тысяч лет назад. Кроме того, многие второстепенные сублинии Q-M242 были обнаружены в популяциях из Внутренней Евразии, включая монголо- и тюркоязычные популяции. В новой работе авторы также обнаружили различные сублинии Q-M242.
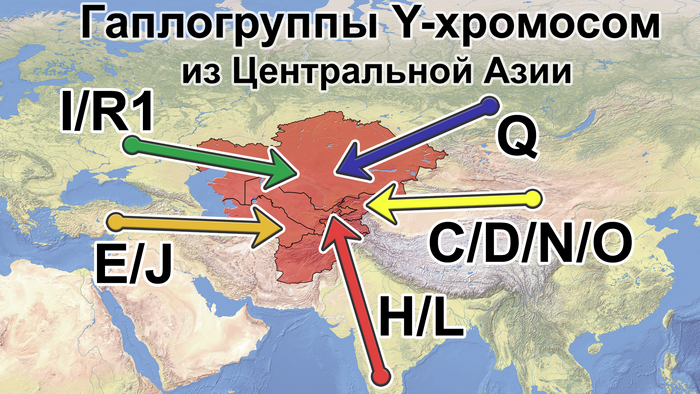
Также известно, что R1a1a-M17 и R1b-M343 являются двумя основными отцовскими линиями индоевропейского населения и большинство современных популяций в Центральной Азии по-прежнему демонстрируют высокую частоту R1a1a-M17. Пересмотренное филогенетическое дерево высокого разрешения с оценкой возраста дает подробный обзор вклада по отцовским линиям в популяциях Центральной Азии. Высокое разнообразие линий макрогаплогрупп предполагает интенсивную примесь из разных периферийных областей Евразийского континента, так линии C/D/N/O из Восточной Евразии, E/J с Ближнего Востока, H/L из Южной Азии, I/R1 из Западной Евразии и Q из Южной Сибири.
Однако наиболее важным открытием этой работы является идентификация уникальных редких, так называемых, минорных линий, которые в Центральной Азии распространились недавно. В том числе 33 такие линии моложе 2000 лет.
Выводы
Множественные события миграций и различные линии гаплогрупп
Центральная Азия не является регионом зарождения земледелия эпохи неолита или технологических систем эпохи бронзы. Поэтому роста населения, вызванного этими технологическими революциями, в Центральной Азии не было. Однако генетические данные показывают, что источниками популяций для Центральной Азии были различные регионы Евразии, включая Южную Азию, Ближний Восток, Европу, Северную и Восточную Азию. Этот общий сценарий согласуется с географическим положением Центральной Азии как центра Евразии. Причём эти миграции происходили постепенно в разные периоды. Так, неолитическая революция на Ближнем Востоке привела к глобальному распространению гаплогруппы J2-M172 и её дочерних линий, параллельно с появлением первых неолитических поселений в южной части Центральной Азии.
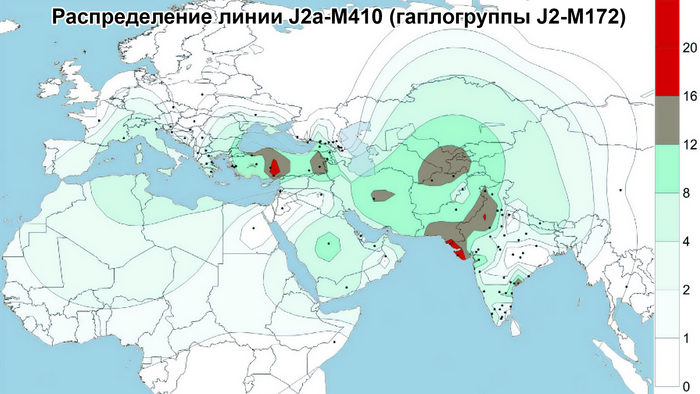
А позже непрерывная миграция с Ближнего Востока привела к появлению дополнительных сублиний J2-M172.
Многие исследования древней ДНК показали, что распространение отцовских линий R1a1a-M17 и R1b-M269 согласуется с экспансией индоевропейских популяций, и это может быть связано с разнообразными сублиниями R1a1a-M17 в Центральной Азии. Кроме того, гаплогруппа G-M201, вероятно, также происходит от индоевропейцев. Однако основываясь на ранее опубликованных генетических данных древних и современных популяций, авторы предполагают, что наличие в Центральной Азии некоторых линий Q-M242, N1a1-M46, R1a1a-M17 и C2a1b1-F1756 может быть связано с миграцией тюркских народов. А монголоязычные популяции привнесли в Центральную Азию линии C2a1a3-M504, отдельные C2a1b1-F1756 и сублинии C2a1a2a-M48. Во время миграции с Востока на Запад, на протяжении сотен лет, многие отцовские гаплогруппы распространились из Восточной Азии в Центральную Азию, включая несколько сублиний D-M174, C2a-F1096 и O-M175. В целом из-за высокой скорости мутаций Y-хромосома легко образует специфические для популяции сублинии или субклады, как кому удобнее.
Однако в комментариях люди часто неверно трактуют предковые гаплогруппы опираясь только на те популяции, у которых встречаются их сублинии. Не говоря про тех, которые им приписывают какие-то невероятные свойства.
Недавние миграции и формирование современного населения
Авторы работы предполагают, что недавнее появление редких минорных предковых линий напрямую способствовало формированию современных популяций Центральной Азии.
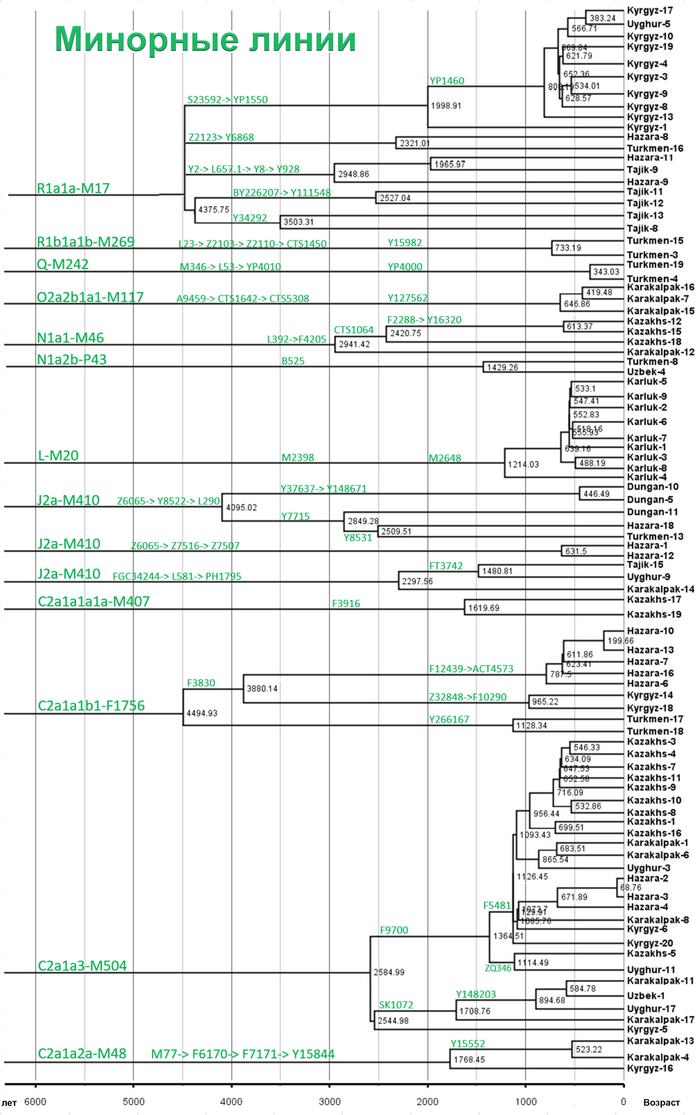
Основные отцовские гаплогруппы популяций в разных регионах Евразии включают C-M217, D-M174, E-M96, G-M201, H-L901, I-M170, J-M304, L-L298, T-M184, N-M231, O-M175, Q-M242, и R-M207. Эти макрогаплогруппы распространены и в популяциях Центральной Азии, однако их частоты существенно различаются. На уровне сублиний население Центральной Азии имеет определенные доминирующие линии.
Но как показано на графике, среди населения Центральной Азии было и много мелких минорных предковых линий характерных для некоторых этнических групп, как к примеру, R1a-M17-Y34292 и R1a-M17-Y111548 у таджиков. Наблюдается множество минорных предковых линий, которые расширились за последние 1000 лет, таких как C2a1a3-M504-F5481, L-M20-M2648 и R1a1a-M17-YP1460. Многие такие линии от R1a1a-M17 у таджиков связаны с их индоевропейскими предками. А в целом минорные линии наблюдались у казахов, кыргызов, карлуков, каракалпаков и хазарейцев.
Авторы считают, что расширение этих специфических линий напрямую связано с формированием современных этнических групп в Центральной Азии.
Таким образом, построено филогенетическое дерево линий Y-хромосом с высоким разрешением для популяций из Центральной Азии. Где значительное разнообразие отцовских линий, указывает на примесь из различных регионов Евразии в разные периоды времени. А множество минорных предковых линий указывает на недавние события примеси, которые могут соответствовать формированию современных популяций Центральной Азии за последнюю тысячу лет.
Источники: Maxat Zhabagin, Lan-HaiWei, Zhaxylyk Sabitov, Peng-Cheng Ma , Jin Sun, Zhanargul Dyussenova, Elena Balanovska, Hui Li and Yerlan Ramankulov. Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosome from Central Asia // Genes. 2022, 13, 1776. doi.org/10.3390/genes13101776
Показать полностью
13