
Образование, ахаха, что ты делаешь, прекрати

Показать полностью
1
Сегодня будет сложно. И вам и мне. Но мы попытаемся) Итак…
Отсюда (1,2,3) вы уже знаете немного о генетике и что-как-кому-зачем передается. Настало время двигаться дальше! Не подчиняясь аутосомно-доминантному и рецессивному типам наследования, а также сцеплению с полом, наследуются следующие штуки:
1. Митохондриальные болезни
2. Болезни экспансии
3. Однородительские дисомии (ОРД) и болезни геномного импритинга
Обо всем по порядку.
Митохондриальные болезни.
Митохондрии. Наши маленькие друзья. Батареечки наших клеточек. По одной из приятных мне теорий, митохондрия - это такая бактерия, которая залезла в наши клетки много лет назад и стала нам помогать в конечном счете. Мне кажется это логичным, т.к. форма ее очень похожа на бактериальную. А еще у нее, как и у бактерии есть своя ДНК! Она не похожа на ДНК остальных наших клеток, она уникальна, и… она тоже мутирует.
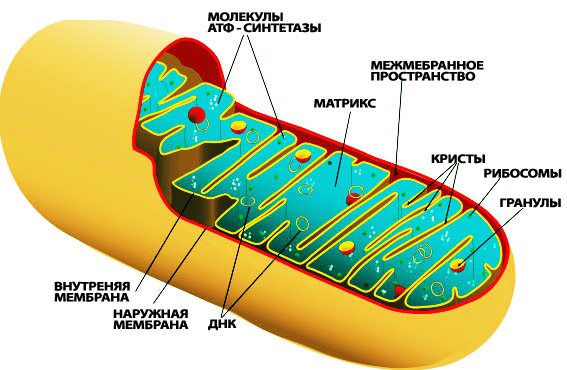
Крошечные желтые колечки - митохондриальная ДНК.
Когда сперматозоид влезает в яйцеклетку, он «отбрасывает коньки» - хвост остается на перроне, а поезд трогается без него. Нет, серьезно, хвост отваливается.
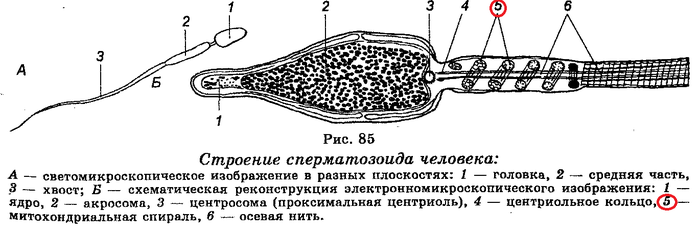
Видали, где митохондрии? Вот. Поэтому, мужская митохондриальная ДНК в жизни будущего зародыша не участвует. А вот митохондрии яйцеклетки – да. Это митохондрии нового организма.
Тое есть, получается, что митохондриальные заболевания передаются от мамы детям. Всем-всем любым детям, независимо от пола. 100%. А от больного папы – никому.
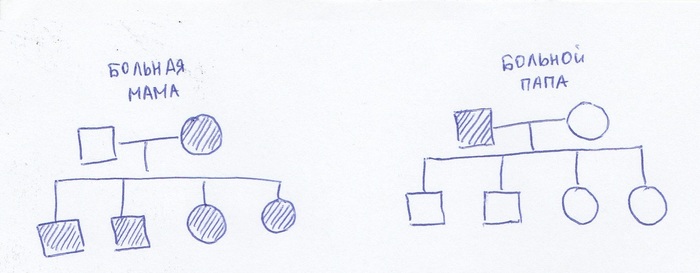
Вечно меня косит, ну да ладно.
Вот такой-вот простой механизм.
Единственной оговоркой в наследовании митохондриальных заболеваний становится то, что за годы совместного существования, мы взяли под контроль часть функций митохондрий. И если мутация произошла в ядерной ДНК, в той ее части, которая контролирует функцию митохондрий, механизм передачи заболевания, разумеется, будет другой.
Т.к. митохондрии – источник энергии, то при митохондриальных заболеваниях поражаются, первым делом, самые энергозависимые ткани: нервная система, эндокринные органы, мышцы скелета, сердце, почки, печень. Тяжесть состояния, как всегда, разнится при разных синдромах.
Болезни экспансии.
Снова болезни, начинающиеся с поражения ДНК. На сей раз – ядерной ДНК.
В нашем геноме есть повторяющиеся участки.
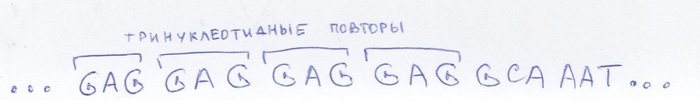
У меня в примере – трехбуквенные повторы, количество букв, на самом деле, может быть разным.
Тринуклеотидные повторы – мой осознанный выбор, т.к. большинство болезней экспансии – именно тринуклеотидные.
Вот есть себе повторы и есть. Что такое экспансия? По словарю Ожегова:
ЭКСПАНСИЯ, -и, ж, (книжн.). 1. Политика, направленная на экономическое и политическое подчинение других стран, на расширение сфер влияния, на захват чужих территорий.
2. Расширение, распространение чего-н. за первоначальные пределы. Э. эпидемии.
В нашем случае, больше второй вариант, иначе стоял бы тег политика. И написано бы было совсем о другом.
В результате сбоя при копировании ДНК (когда перед глазами постоянно мелькают одинаковые буквы – легко сбиться со счета), происходит увеличение количества этих повторов. Когда их количество переваливает за максимальный порог (для разных повторов он разный), возникает заболевание.
Болезни экспансии отличаются друг от друга и клиникой, и тяжестью. Обычно, чем больше повторов, тем тяжелее заболевание протекает.
Помните Тринадцать из Хауса, которую играла прекрасная Оливия Уайлд? У нее еще была болезнь Гентингтона. Ну вот. Хорея Гентингтона – экспансия CAG-повторов в гене HD, для развития болезни их должно быть больше 40 штук.

Вот такая вот фигня. Обычно, болезни экспансии наследуются по доминантному типу – достаточно увеличения числа копий только на одной хромосоме, но есть и рецессивные. Еще для них характерно «утяжеление» симптомов в последующих поколениях.
И наконец...
ОРД и геномный импринтинг.
В норме, мы наследуем 23 хромосомы от мамы и 23 хромосомы от папы. Но, бывает, что-то идет не так, и мы наследуем пару хромосом только от одного из родителей. Это и есть однородительская дисомия (ОРД). Варианты могут быть разные, а причина – нерасхождение хромосом. Оно может происходить на разных этапах клеточного деления и приводить к разным последствиям. Об этом я писала в прошлый раз.
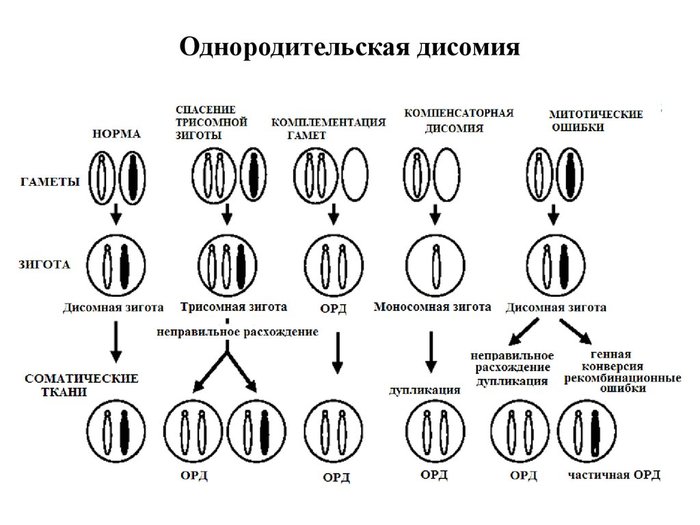
Вот сколько много всяких вариантов.
Пример. У человека есть две седьмые хромосомы. Но они не абсолютно одинаковые. Одна же ему досталась от мамы, другая от папы. Поэтому на них разные мутации и вообще. Разные они.
Если от этого человека ребенку достанется одна из этих хромосом в дублированном варианте – это изодисомия. Если обе – гетеродисомия. И то и то - неправильно, и будет считаться ОРД по хромосоме 7.
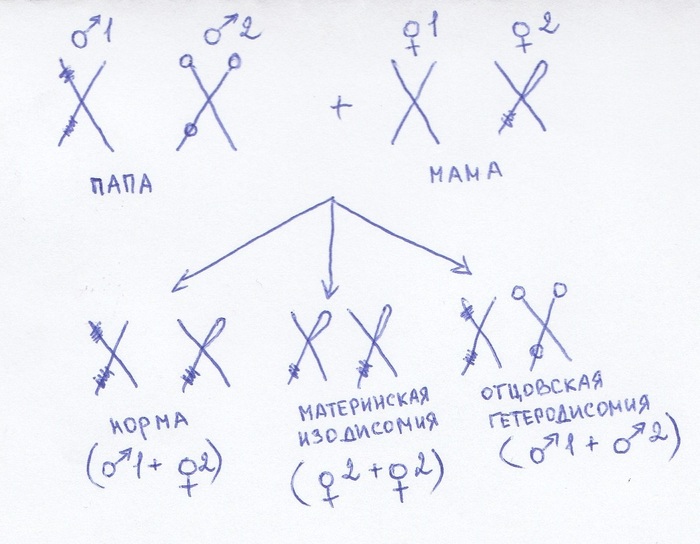
По схеме вы сами справитесь с достраиванием других вариантов деток, которые могут получиться, это просто пример)
Итог: от двух родителей – хорошо, от одного – плохо.
Не по всем хромосомам описаны ОРД, но, например, при отцовской ОРД по 14й хромосоме у ребенка отмечается умственная отсталость, патологии скелета, мышечная слабость.
Импринтинг же - след. Это следы на нашем геноме. Здесь была мама, а здесь был папа.
Хромосомы от папы несут «папский» отпечаток на себе, от мамы – «мамский». И вот мы состоим из 23 хромосом с «мамским» отпечатком и из 23 с «папским». Могли ли вы подумать?) Если инактивируется ген на одной из хромосом, то активным сразу станет второй такой ген, но вот чей отпечаток он несет? И в зависимости от того, чей он, эффект от его активации будет разным.
Не буду забивать вам более голову этим сложнючим механизмом, вы и так сегодня узнали много нового. Завершая все это дело, просто приведу интересный пример.
Есть два синдрома: синдром Ангельмана и синдром Прадера-Вилли. И они абсолютно разные.
При синдроме Ангельмана у детей грубая задержка в развитии, умственная отсталость, абсолютное недоразвитие речи, они худые, смеются без причины, у них специфическая походка, как у марионетки. Этот синдром еще называют синдромом "счастливой куклы".
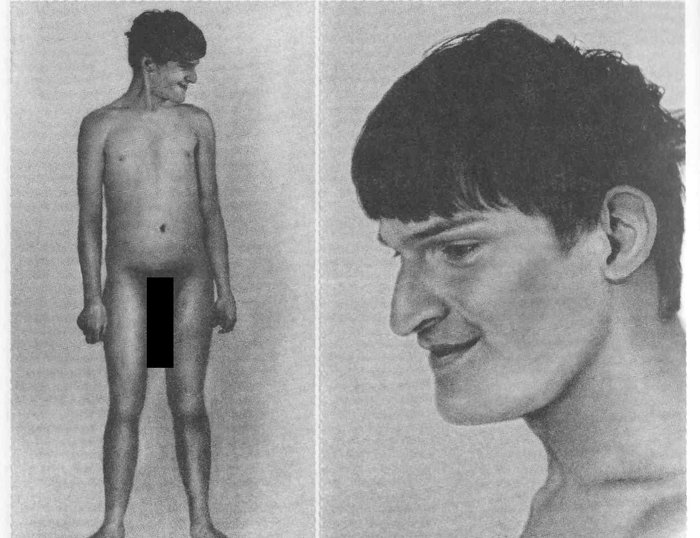
При синдроме Прадера-Вилли же – наоборот – умственно отсталые вялые детишки, с маленькими ладошками и стопами, отсутствием чувства насыщения, что ведет к ожирению. Обычно обладают добродушным характером и апатичны. Иногда мне встречается описание характера как «зловредного», но на практике пока подтверждается первый вариант. Будешь тут зловредным, когда тебя пытаются ограничить в еде.

Juan Carreno de Miranda, «La Monstrua vesti-da» и «LaMonstruadesnuda» («Одетый монстр» и «Раздетый монстр»). Считается, что на картинах изображен именно этот синдром.
Оба эти заболевания являют собой самый известный в генетической среде пример ОРД и импринтинга.
При синдроме Ангельмана – 68% всех случаев - это делеции 15q11-13 (область длинного плеча хромосомы 15) материнского происхождения (т.е. импринтинг), 7% - ОРД отцовского происхождения.
При синдроме Прадера-Вилли – 70% делеции 15q11-13 отцовского происхождения, 25% - ОРД материнского происхождения. Оставшиеся проценты от всех случаев наследуются несколько иначе.
Вот так - одна и та же мутации с разным отпечатком - абсолютно разные состояния.
И на сегодня все. Устали?) Надеюсь, что нет) Вы молодцы! Опять вы узнали что-то новое, опять образовались. Я ухожу со сцены, оставляя вас одних все это переваривать. Берегите свои повторы)









Привет подписчики и не очень) Без лишних слов, приступим.
По моему скромному мнению, базовое исследование в области генетики человека – кариотипирование.
Мы уже с вами выясняли, что кариотип – это наш хромосомный набор. Для чего может понадобиться его визуализация? Для определения пола, например. В онкологии. Или для выявления хромосомных аномалий (геномные и хромосомные мутации).
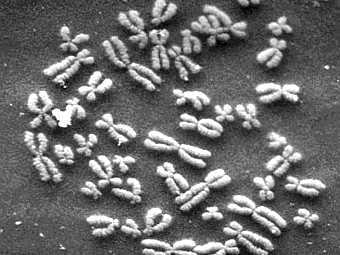
Для стандартного (и не только) кариотипирования необходимы делящиеся клетки, т.к. именно в таких клетках мы можем увидеть сформированные хромосомы. Чаще всего для этой цели используются лимфоциты – клетки крови, но на самом деле клетки могут быть из самых разных тканей.
Как это выглядит? Для начала, нужно сдать кровь. Можно кушать, уровень глюкозы измерять не будут, но необходимо не применять препараты, которые мешают клеткам расти (противоопухолевые препараты, антибиотики). Так же желательно дождаться, когда очередной ОРЗ отступит.
Итак, вы здоровы, не принимаете лекарственные средства и уже сдали кровь? Отлично! Уходите домой и ждите 2 недели. Все остальное за вас сделает лаборатория.
Ваша кровь помещается в специальную питательную среду, где лимфоциты будут расти в тепле и покое. По истечении времени (около 70 часов), в клеточную среду добавляют вещество, которое прерывает деление клеток на стадии метафазы митоза (эта та стадия деления, на которой хорошо видно хромосомы). Затем необходимо разорвать клеточную стенку, чтобы все хромосомки были видны, зафиксировать все это дело и покрасить.
Вариантов окраски может быть несколько. Чтобы просто посчитать хромосомы достаточно рутинного (сплошного) окрашивания:

Для более детального рассмотрения (что бы не пропустить внутрихромосомные аномалии – удвоение, делеции участков) изобретены другие - дифференциальные методы окрашивания. Например G-окрашивание, при котором на хромосоме хорошо видны темные и светлые полосы, на которые можно ориентироваться.

Потом в этой каше уже копается специалист – цитогенетик, который глазами и мозгом изучает ваши хромосомы и ищет там всякие ошибки.
FISH - fluorescent hybridization in situ, еще один интересный метод цитогенетики. Покажу, потому что красиво. Хромосома, вся или только интересующий участок, метится светящимися зондами разных цветов. И методика тут уже сложнее, чем при рутинном кариотипировании.

Пример нескольких зондов на одну хромосому (15). 2 световых сигнала – хромосома с вырезанным участком (делецией), 3 сигнала – здоровая хромосома. Синдром Прадера-Вилли (в другой раз)
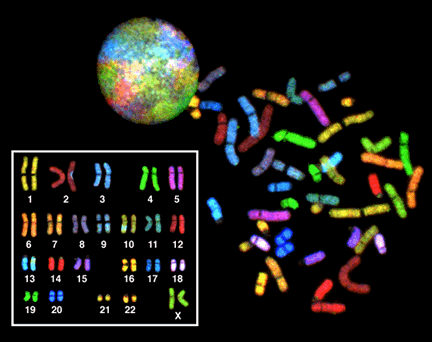
Мультицветный FISH.
Естественно, это далеко не все методы цитогенетики. Есть еще CGH, arrayCGH, но о них мы говорить не будем.
Зачем здоровым, казалось бы, людям сдавать кариотип? Парам с бесплодием, например, кариотип рекомендуют настоятельно. У них же не может быть лишней хромосомы, правда? Может, конечно. У женщины с кариотипом 47, ХХХ, хоть она и здорова, однако могут наблюдаться проблемы с фертильностью. Мужчины с синдромом Клайнфельтера (47,ХХУ), тоже могут выглядеть вполне себе здоровыми мужчинами, или просто не подозревать о своей болезни. Мозаицизм тоже никуда не делся (его мы рассматривали в прошлый раз). И вообще, мало ли чего.
Но есть одна нередкая штука, про которую я вам сейчас расскажу.
Транслокации – особенный вид мутаций, при котором происходит перемещение материала между негомологичными (разными из пар) хромосомами. Между 8й и 14й, 9й и 22й и т.д.
Обмен участками между гомологичными (одинаковыми) хромосомами называется кроссинговер (не уйти мне от него), и это нормальное явление. 4 и 4 хромосома могут преспокойнененько поменяться участками, и никто их не обвинит в транслокации - кроссинговер это хорошо.
Транслокация может быть сбалансированной (обмен, ничего не теряется, все остается в геноме, но на чужих местах), инсерционной (один участок с одной хромосомы переносится на другую, insert - вставить). По некоторым классификациям инсерцию вообще не относят к транслокациям, но это не суть. Выглядит это примерно так:

норма -------------------------------- сбалансированная ---------------------------------- инсерция
Сбалансированные транслокации – самые частые. Примерно каждый 1000й человек несет в себе такую перестройку и в ус не дует. Потому что носители таких транслокаций, как правило, абсолютно нормальные люди. Все же на месте, ничего не утрачено. Но у них высоки риски бесплодия, выкидышей, рождения детей с наследственной патологией, т.к. половина их половых клеток (гамет, клеток с половинчатым набором хромосом, что бы при слиянии снова получить полный набор) несут в себе несбалансированный материал.
И третий вариант. Робертсоновские транслокации – это обмен между акроцентрическими (как плохо-то получается без терминов) хромосомами, с потерей коротких плеч.
Ща попробую коротенько.
Смотрите:
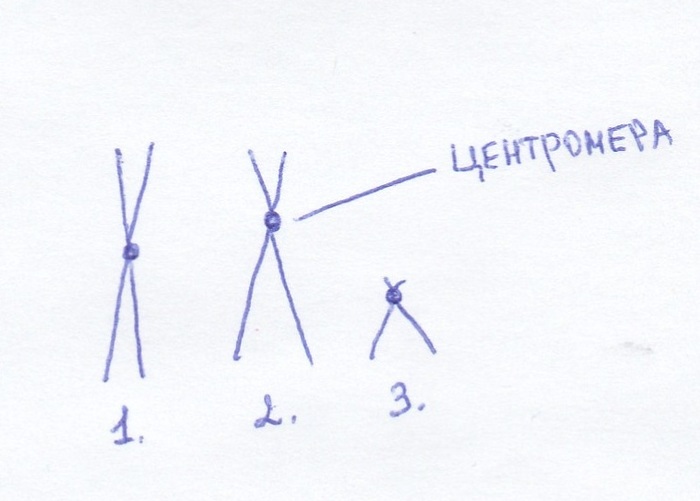
1. Метацентрическая хромосома. Центромера (серединка) делит ее на две одинаковые половины
2. Субметацентрическая хромосома – центромера смещена, четко видим длинное плечо и короткое.
3. Акроцентрические – центромера очень далеко от центра. Короткое плечо совсем короткое, а длинное совсем длинное.
Ну и вот. Робертсоновская транслокация (РОБ) выглядит примерно так:

Шесть типов гамет, которые может образовать человек с РОБ, если соединить их с половой клеткой здорового человека, получат такие варианты детей (статистически, господа):
1. 14,21 +14,21 = 14,21;14,21 – Это здоровый ребетенок - копия здорового родителя;
2. 21/14 + 14,21 = 14/21;14,21 – Носитель робертсоновской транслокации, как и один из родителей;
3. 21/14,21+14,21 = 21/14,21;14,21 – Считаем: три 21х хромосомы, две 14х. Это транслокационная форма синдрома Дауна;
4. 14+14,21 = 14;14,21 – Нет 21й хромосомы – такие дети не выживают;
5. 14,14/21+14,21 = 14,14/21;14,21 – Три 14х – летально;
6. 21 + 14,21 = 21;14,21 – Нет 14й – опять летально.
Не запутались? Очень надеюсь, что нет.
И сбалансированные и робертсоновские транслокации - подлая штука. Любое нарушение состава хромосом влечет за собой образование «неправильных» гамет. Согласитесь, обидно, быть полностью здоровым , но раз за разом натыкаться на неудачные беременности, которые очень тяжело воспринимаются психически, деморализуют, изматывают физически.
Поэтому всем парам с бесплодием необходимо делать кариотип. Предупрежден – значит вооружен.
На сегодня все. Всем спасибо, вы молодцы. Берегите кариотип смолоду)
1. РНК-полимеразы эукариот
Как мы помним, эукариоты - организмы имеющие оформленное ядро. Процесс транскрипции у эукариот по смыслу не отличается от прокариот, однако каждая из стадий обрастает все новыми подробностями.
Начнем с обзора эукариотических РНК-полимераз. Если в клетках бактерий все гены транскрибировала одна полимераза, то у эукариот существует, как минимум 3 типа РНК-полимераз, работающих в ядре:
1) РНК-полимераза типа I - синтезирует предшественницу рибосомальных РНК (рРНК), входящих в состав рибосомы
2) РНК-полимераза типа II - самая изученная полимераза, отвечает за синтез мРНК белков и малых ядерных РНК
3) РНК-полимераза III - синтезирующая тРНК, 5S рРНК и другие малые РНК, присутствующее в ядре и цитозоле.
Остальные РНК-полимеразы, работающие в митохондриях и хлоропластах. Например, РНК-пол IV – синтезирует siРНК в семенах растений, которая метилирует гены 5S; РНК-пол V – синтезирует РНК, которая вовлекается в siРНК-направленное формирование гетерохроматина у растений, а так же митохондриальная РНК-пол – 2 СЕ у дрожжей (145 и 43 кДа) и 1 СЕ у высших эукариотов (140 кДа)
Все РНК-полимеразы эукариот это мультисубъединичные белковые комплексы, с большой молекулярной массой. Гигантские машины для синтеза РНК.
2. РНК-полимераза II
Как отмечалось ранее, РНК-полимераза II является наиболее изученной полимеразой, а в 2006 году Роджер Корнберг был удостоен Нобелевской премии по химии за исследование механизма копирования клетками генетической информации, в частности за установление соотношения атомов в РНК-полимеразе эукариотов.
Как оказалось, эукариотические РНК полимеразы не способны сами по себе инициировать транскрипцию. Для этого им необходимы вспомогательные белки - факторы инициации, формирующие совместно с полимеразами преинициаторные комплексы.
В состав РНК-полимераз входят: самые большие СЕ (135 и 195 кДа) гомологичны СЕ бэта и бэта' бактерий, они формируют активный центр, 35-кДа СЕ – аналог альфа-СЕ бактериальной РНК-полимеразы, а так же некоторые другие СЕ отдаленно напоминающие СЕ бактериальных полимераз.
Теперь, чтобы начать транскрипцию необходимо наличие вспомогательных белков, которые называются транскрипционные факторы. Вначале с промотором взаимодействуют общие для всех генов класса II факторы транскрипции, они подготавливают промотор для связывания РНК-пол II. В результате формируется базальная машина транскрипции генов класса II - PIC.
Пока вы можете оценить всю мощь комплекса РНК-полимеразы и различных факторов транскрипции по видео (может немного рановато, но вдохновляюще)
3. Эукариотические промоторы
Я думаю сейчас самое время поговорить о промоторах. Как мы помним из прошлых постов, у прокариот промотор выглядел просто и состоял из двух блоков. Тут же дела обстоят несколько сложнее. Вообще для регуляции экспрессии генов существуют:
Цис-элемент (цис-действующий сайт, цис-регуляторный элемент) – последовательность ДНК, влияющая на транскрипцию гена, к которому она принадлежит территориально (на той же молекуле ДНК). Может располагаться в промоторе, в интронах и за кодирующей частью гена. Как правило, не транскрибируется
Транс-факторы (транс-действующий сайт) – последовательности ДНК, кодирующие белки, изменяющие активность генов, расположенных в любом месте генома. Эти последовательности транскрибируются и транслируются – транскрипционные факторы
Главными (коровыми) элементами промоторов генов класса II являются:
ТАТА-блок -25 п.н. с 5'-стороны от точки +1 - ТАТААА (это вполне объяснимая последовательность, т.к. между нуклеотидами А и Т образуются 2 водородные связи, и нити с такими последовательностями куда проще разъединить, чем пары Г и Ц с 3мя водородными связями)
Инициаторный (Inr) элемент располагается вокруг стартовой точки транскрипции
sINR – более консервативен и дополнительно имеет консервативные фланкирующие последовательности
Любого из них достаточно для правильной инициации транскрипции in vitro
Промоторы для РНК-полимеразы II могут включать в себя разное количество элементов:
1) ТАТА-бокс и Inr (очень сильный промотор)
2) Inr- последовательность и отсутствие ТАТА-бокса (ТАТА-less)
3) ТАТА-бокс
Если вы расслабились и подумали, что это всё, вынуждена вас огорчить...
С промоторами встречаются:
DCE – расположен по ходу транскрипции и состоит из трех субэлементов: SI (CTTC) располагается +6 - +11, SII (CTGT) располагается +16 - +21, SIII (AGC) – от +30 до +34
Цис-элемент В – участок ДНК, узнающий TFIIB - BREu и BREd (G/C G/C G/A C G C C), элементы, фланкирующие ТАТА-боксом, работают только с ТАТА-боксом
DPE – элемент промотора, расположенный по ходу транскрипции A/gGA/tC/tG/a,c (+28 - +32), характерен для промоторов, потерявших ТАТА-бокс
XCPE1 (10 п.н. -8+2, пурины) и ХСРЕ2 (11 п.н., пиримидины), найдены в промоторе гена Х вируса гепатита В, у человека 1% генов содержит эти элементы
UAS – элементы, усиливающие промотор: GC-бокс (GGGCGCC) и/или СААТ-бокс (GGCCAATCT), находятся слева от точки +1, на расстоянии примерно 300 пар нуклеотидов.
и многие другие. поэтому промотор теперь становится длиной от 100 до 1000 пар нуклеотидов.
Некоторые комбинации промоторов:

Наличие различных по виду промоторов является необходимым условием для жизнедеятельности клетки. С помощью различных вариаций элементов достигается контроль над экспрессией генов. Есть некоторые наблюдения, например: наличие различных элементов влияет не только на скорость транскрипции, но и трансляции; существует связь между типом корового промотора и длиной гена: гены, имеющие ТАТА-промоторы, в три раза короче генов, потерявших ТАТА-бокс; укорочение гена связано с укорочением длины интронов.
Так же к важнейшим цис-элементам можно добавить:
Энхансеры – регуляторные последовательности ДНК, расположенные на расстоянии тысяч пар нуклеотидов справа или слева от точки старта и усиливающие транскрипцию регулируемого гена.
Сайленсеры - регуляторные последовательности ДНК, расположенные на расстоянии тысяч пар нуклеотидов справа или слева от точки старта и ослабляющие транскрипцию регулирумого гена.
Таким образом, сборка инициирующего комплекса транскрипции у эукариот не ограничивается одним промотором, а включает в себя элементы, находящиеся на расстоянии тысячи пар нуклеотидов от регулируемого гена.
На рисунке изображен преинициирующий комплекс РНК-полимеразы, вы можете оценить его сложность:
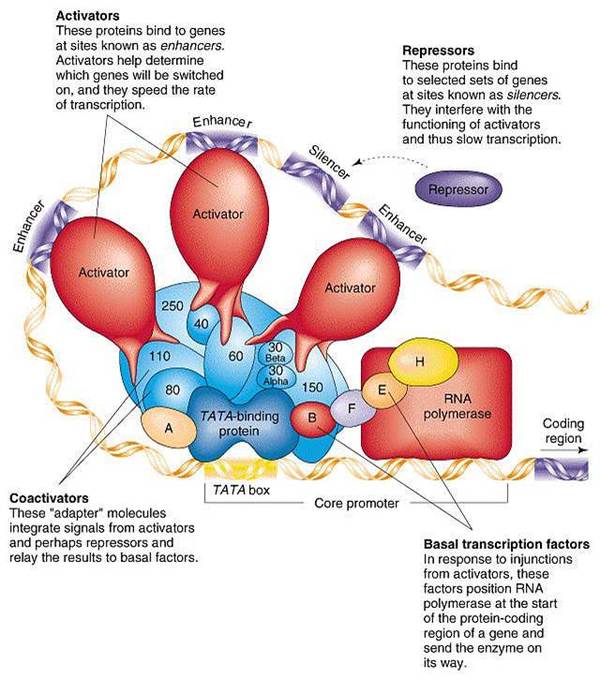
в следующем посте я планирую рассказать о том, как именно собирается преинициирующий комплекс РНК-полимеразы. Спасибо за внимание!
Продолжение постов о молекулярной генетике. (Эта часть получилась большой и менее красочной, я бы даже сказала, как задача со звездочкой)
Существует несколько моделей работы РНК-полимераз. Тут я хочу рассказать о двух моделях, которые наилучшим образом подтверждают экспериментальные исследования. Следует сказать, что мы переходим к рассмотрению следующей стадии транскрипции - элонгации, "осмысленному" движению РНК-полимеразы по ДНК и синтезу мРНК.
Что вообще требуется для работы РНК-полимеразы:
1) обязательным условием является наличие ионов Mg (2+), входящих в состав каталитического центра РНК-полимеразы.
2) рибонуклеозид-трифосфаты (АТФ, ЦТФ, УТФ, ГТФ)
3) и конечно, наличие самой ДНК.
Скорость движения РНК-полимераз измеряется в количестве присоединенных нукледотидов в секунду, так для полимеразы бактериофагов способны элонгировать in vitro растущую цепь РНК со скоростью 200–400 нт/с. Бактериальные РНК-полимеразы транскрибируют ДНК с промежуточной скоростью – 50–100 нт/с, эукариотические РНК-полимеразы элонгируют цепи РНК in vivo со скоростью ~20–30 нт/с. (тут прослеживается некоторая корреляция с размерами и количеством СЕ, ходящих в состав РНК-полимераз)
Основные черты структуры тройного комплекса, осуществляющего элонгацию цепей РНК, консервативны у всех ДНК-зависимых РНК-полимераз. В каждом элонгирующем комплексе имеются каталитический центр, одноцепочечная область ДНК-матрицы, а также несколько сайтов связывания ДНК и РНК. Для реализации принципа комплементарности при построении растущей цепи РНК участок матричной ДНК, входящий в состав комплекса, находится в расплавленном состоянии, и одна из его цепей служит матрицей при транскрипции. Этот участок ДНК, называемый транскрипционным пузырьком, или транскрипционной сферой, контактирует с каталитическим центром РНК-полимеразы. По обеим сторонам транскрипционного пузырька имеются участки ДНК, которые при перемещении фермента вдоль матрицы подвергаются плавлению (расплетанию) и повторному отжигу, в результате которого восстанавливается исходная структура ДНК. Считается, что этот процесс не является каталитическим, протекает самопроизвольно и связан с особенностями структуры РНК-полимеразы как таковой.
1. Модель прерывистого "червеобразного" движения
В 1992 г. М. Чамберлин с сотрудниками разработали модель элонгации РНК, в которой постулировалось, что процессы транслокации РНК-полимеразы вдоль ДНК и присоединение нуклеотидов к растущей цепи РНК в активном центре фермента разделены во времени. То есть простым языком можно сказать, что РНК-полимераза стоит из 2х частей, соединенных сжимаемой "пружиной". Существует 2 сайта связывания полимеразы ДНК: Сайт 1 в передней части по отношению к направлению транскрипции, а сайт 2 в передней.
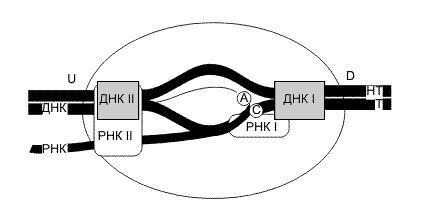
В соответствии с предложенной моделью, молекула РНК- полимеразы перемещается вдоль ДНК наподобие гусеницы: когда один из сайтов связывания ДНК фиксирован, другой может двигаться вперед.
В данной модели гибрид РНК/ДНК не вносит вклад в стабильность транскрипционного комплекса, а растущая цепь РНК удерживается благодаря наличию двух сайтов связывания РНК-полимеразы с РНК: участок прочного связывания (1) (5' конец) и слабого связывания (2) (3' конец).
Согласно предложенной модели элонгация цепей РНК представляется в виде циклического процесса.В начале цикла каталитический центр молекулы РНК-полимеразы располагается у задней границы РНК-связывающего сайта 1 в соответствии с положением 3'-ОН-конца РНК.Последовательно присоединяя нуклеотиды к растущей цепи РНК, каталитический участок перемещается относительно РНК-связывающего сайта I и в конце концов заполняет этот сайт десятью нуклеотидами вновь синтезированного участка РНК. Во время этой фазы элонгации ДНК-связывающий сайт 2 остается фиксированным на ДНК, тогда как ДНК-связывающий сайт 1 перемещается вперед синхронно с каталитическим участком на десять нуклеотидов. В конце фазы добавления нуклеотидов ДНК- и РНК-связывающие сайты 2 фиксируются на своих лигандах, а ДНК-связывающий сайт 1 переносится вперед на десять нуклеотидов в новое фиксированное положение. Это перемещение освобождает РНК-связывающий сайт 1, делая его готовым к повторению цикла транслокации.
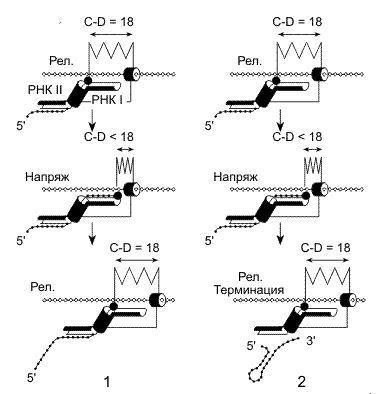
На рисунке показаны ДНК, синтезируемая РНК, а диагональными линиями - нарастающее напряжение белкового остова РНК-полимеразы.
При дальнейших исследованиях оказалось, что перемещение тройного комплекса вдоль транскрибируемой ДНК не всегда скачкообразно.
Большую часть транскрибируемых ДНК молекулы РНК-полимеразы проходят монотонно, регулярно присоединяя к растущим цепям РНК нуклеотид за нуклеотидом. Прерывистая, скачкообразная элонгация РНК имеет место лишь на участках матричной ДНК, в которых происходит задержка транскрипции или ее прекращение. Задержками могут быть последовательности-палиндромы, образующие шпильку в РНК , а так же последовательности, богатые урацилом.
2. Современное представление элонгирующего комплекса.
Дальнейшие исследования уточнили геометрию тройного комплекса и структуру каталитического центра. В ходе обширных биохимических и генетических исследований локализован активный центр РНК-полнмеразы, а также участки связывания нуклеиновых кислот, необходимых для стабильности элонгационного комплекса.
Выяснилось, что три остатка аспарагиновой кислоты в консервативном у всех многосубъединичных РНК-полимераз мотиве NADFDGD самой большой субъединицы (бэта') хелатируют каталитический нон Mg (2+). Субъединицы бэта’ и бэта формируют три сайта связывания: 1) сайт связывания дуплекса ДНК из 10 пар оснований (п.о.) перед активным центром. 2) сайт связывания гибридного участка РНК-ДНК длиной 8—9 п.о., образующегося в "транскрипционном пузырьке" из 12—15 п.о. расплавленной ДНК. и 3) сайт связывания одноцепочечного участка РНК. удаленного на 8—14 нуклеотидов от З'-конца РНК.
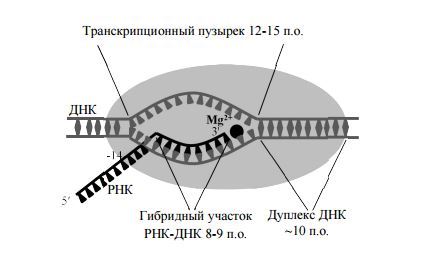
Выяснилось, что в элонгационном комплексе передний (по ходу движения РНК-полимеразы) дуплекс ДНК располагается в основном канале между субъединицами бэта' и бэта. Прямые контакты РНК-полимеразы с участком ДНК из ~10 п.о. образуют функциональный сайт связывания ДНК. Подвижный домен бэта', названный “зажимом” (clamp), удерживает ДНК в
канале. Каталитический центр РНК-полимеразы находится рядом с точной расплетания ДНК. Полипептидная петля на бэта СЕ препятствует прохождению двунитеой ДНК в активный центр, и, по-видимому, играет важдую роль в плавлении ДНК, тут цепь ДНК изгибается под углом примерно 90 градусов, освобождая место для формирования гетородуплекса ДНК-РНК. Сам дуплекс находится в канале, образованном бэта и бэта' СЕ. После синтеза 9 нуклеотидов цепь РНК отделяется от ДНК, параллельно с этим восстанавливается дуплекс ДНК, способствующий вытеснению РНК.
В ходе матричного синтеза РНК-полимераза катализирует нуклеофильную атаку 3'- гидроксила растущей цепи РНК на альфа-фосфат поступающего НТФ. в результате чего происходит включение очередного нуклеотида в цепь РНК и высвобождение пирофосфата. Активный центр фермента содержит участок связывания З'-конца РНК (i-сайт) участок связывания поступающего НТФ (i+1-сайт). За образованием фосфодиэфирной связи следует транслокация вновь образованного З’-конца РНК из i+1-сайта в i-сайт. при этом i+1-сайт освобождается для поступления нового НТФ (посттранслокационное состояние). Вместе с этим происходит разъединение пары оснований в переднем дуплексе ДНК. образование пары оснований в заднем дуплексе и разъединение последней пары оснований в гибриде РНК-ДНК.

В данном представлении становится возможным движение фермента в обратную сторону.
Осцилляции РНК-полимеразы назад и вперед по гибриду РНК-ДНК:
• Приводят к освобождению 3-конца РНК из гибрида
• Освобожденный нуклеотид может быть отщеплен
• Так преодолеваются «трудные» участки транскрипции – сигналы паузы и сигналы остановки, исправляются ошибочно встроенные нуклеотиды
Этот пост получился тяжелым, простите, но, надеюсь, это улучшит ваше понимание вопросов.
Спасибо за внимание.
подробнее в статье: С. А. ПРОШКИН, Г. В. ШПАКОВСКИЙ ЯДЕРНЫЕ РНК ПОЛИМЕРАЗЫ I, II И III: СТРУКТУРА И ФУНКЦИИ // Успехи биологической химии, т.45, 2005, с.269-306
Давно я не писала, простите. Соскучились? тогда продолжаем следующую часть.
В этом посте мы договорились обсудить сборку преинициирующего комплекса РНК-полимеразы (ПИК).
Принципиальными особенностями транскрипции эукариот являются:
• не способность РНК-пол(II) узнавать промоторы, теперь она нуждается в базальных факторах транскрипции
• в начале с промотором взаимодействуют общие для всех генов класса II факторы транскрипции, они подготавливают промотор для связывания РНК-пол II
• количество базальных факторов транскрипции превышает количество РНК-пол II
В результате формируется базальная машина транскрипции генов класса II - ПИК
Рассмотрим сборку ПИК на промоторах, содержащих ТАТА боксы.
В инициации транскрипции учавствуют транскипционные факторы:
TFIID (название формируется изTranscription factor, а римская цифра II, это класс полимеразы, с которой работает фактор) TFIIB TFIIF TFIIE TFIIH TFIIA
TFIID
Первым с ТАТА боксом взаимодействует TFIID - мультисубъединичный комплекс, в состав которого входит белок TBP (TATA-binding protein).
TBP имеет ряд собенностей:
• Имеет форму седла, сближается с ДНК благодаря остаткам лизина и аргинина
• Взаимодействует с ДНК в области первых и последних пар оснований ТАТА-блока 4 остатками фенилаланина
• R-группы фениаланина вставляются между первыми п. о. ТАТА-блока, и ДНК из В-формы переходит в частично раскрученную спираль. Вставка R-групп фенилаланина между последними п.о. в ТАТА-блоке приводит к обратному переходу ДНК в В-форму. Структура ДНК в ТАТА-блоке искажается, прилежащие участки сближаются, это способствует дальнейшей сборке ПИК.
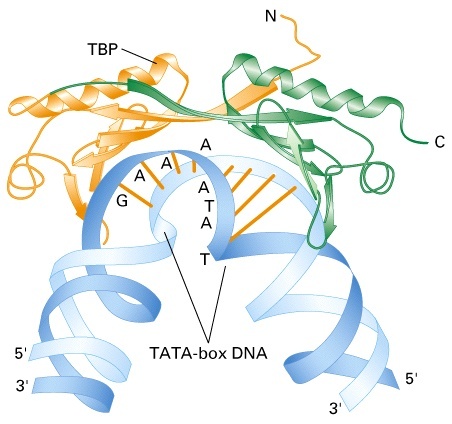
TBP является важным участником транскрипции, т.к. определяет положение РНК-пол II, вовлекается в плавление двунитевой ДНК, изгибая ДНК на 80°.
TFIIB
Следующим присоединяется TFIIB. С-конец TFIIB связывается под седлом комплекса ТВР-ТАТА с ТВР и с ДНК в областях BREu и BREd. Связывание TFIIB асимметрично, определяет положение РНК-пол II. Теперь, Промотор подготовлен для связывания с РНК-пол II.
TFIIF
Находится в комплексе с нефосфорилированной формой РНК-пол II без ДНК;
Содержит 2 СЕ: 30 кДа – Rap30 и 74 кДа – Rap74
Снижает неспецифическое связывание РНК-пол II и РНК-пол бактерий с ДНК
TFIIF обеспечивает контакт РНК-пол II с TFIIB-ТАТА.
Но и на этом все не заканчивается...
Затем присоединяются TFIIE и TFIIH
Подробнее хочется сказать о TFIIH. Это полифункциональный фермент:
TFIIH(р89) – ATP-зависимая геликаза (направление 3' 5' ) – совпадает с белком ERCC3 (мутации летальны, ответственны за развитие пигментной ксеродермы и синдрома Коккейна)
TFIIH(р80) – ATФ-зависимая геликаза (направление 5' 3') – совпадает с белком ERCC2 (мутации не летальны)
протеинкиназа для CTD-РНК-пол II, эта активность стимулируется фактором TFIIE
TFIIH связывается с РНК-пол II, CTD-домен которой не фосфорилирован
Фосфорилирование РНК-пол II ослабляет связь фермента с ПИК и она переходит к элонгации.

Далее происходит отсоединение фрагментов ПИК и РНК-полимераза продолжает свой путь только TFIIF. Транскрипция с участием минимального инициирующего комплекса происходит с недостаточной скоростью. Для эффективной транскрипции необходимы другие цис-элементы и транс-факторы, а также энхансеры, усиливающие транскрипцию в сотни раз (их мы обсуждали прошлый раз).
Спасибо за внимание!

В народе существует представление о неких врождённых математических способностях, благодаря которым одни дети усваивают математику лучше, чем другие. Мы проверили, есть ли у него научное основание.
Спойлер для ЛЛ: единого вывода на этот счет у ученых пока нет. Однако отмечается, что, независимо от задатков, для развития математических навыков намного важнее благоприятная жизненная среда, доступ к образованию, а также собственное желание и интерес ребёнка.
Вопрос, существует ли так называемое математическое мышление и врождённая склонность к этой науке, можно встретить на порталах для родителей и на женских форумах. Вопросы об этом задают пользователи сервиса «Яндекс.Кью». О генах математических способностей, которые наследуются от родителей, пишут СМИ (например, телеканал «Мир», РИА «Новости», «Комсомольская правда») и научно-популярные порталы.
В 2013 году психологи из Университета Питтсбурга провели эксперимент. Они исследовали несимволическое чувство числа (способность определять количество навскидку, без точного подсчёта) у шестимесячных младенцев и их последующие успехи в математике в три с половиной года. Младенцам показывали карточки, где были изображены разные предметы. На некоторых из карточек количество предметов отличалось. Некоторые дети замечали это отличие, другие — нет. Учёные смогли проследить это по тому, как долго дети задерживали на карточках взгляд. Спустя три года тех же детей проверили на знание цифр и умение считать до десяти. Оказалось, что чем лучше было развито чувство числа у детей, которые ещё даже не научились разговаривать, тем лучше они справлялись с простыми математическими заданиями в более осознанном возрасте. Это дало основания для гипотезы, что склонность к математике, действительно, может быть врождённой.
Несколько лет спустя американские учёные провели исследование, в ходе которого просили детей пяти-восьми лет и их родителей пройти несколько математических тестов. Они заметили прямую зависимость между скоростью выполнения заданий детьми и их родителями, что дало им повод утверждать: талант к математике может передаваться по наследству.
В 2020 году немецкие учёные отчасти подтвердили эту гипотезу. В ходе исследования они сделали магнитно-резонансную томографию (МРТ) 178 детям от трёх до шести лет, а затем предложили им выполнить ряд математических заданий. Они выявили, какие области мозга отвечают за успешное выполнение такого рода тестов. Затем они провели такое же исследование, но уже на детях в возрасте семи-девяти лет. Сравнение показало, что наиболее успешно задания выполняли те дети, у которых объём правой теменной коры головного мозга был увеличен. За его объём отвечает обнаруженный учёными ген ROB01, который передаётся по наследству. То есть, возможно, по наследству могут передаваться и математические способности.
По мнению американских учёных, существует ещё и корреляция между строением мозга и тем, насколько легко детям даётся изучение математики. В ходе эксперимента 24 ребёнка в возрасте восьми-девяти лет два месяца занимались с индивидуальными репетиторами по математике. Разумеется, значительное продвижение в изучении показали все дети, однако некоторые смогли обогнать других участников эксперимента. Всем детям перед исследованием сделали МРТ, и сравнение структуры мозга и успехов в обучении показало: легче и быстрее всего математика давалась тем детям, у которых объём гиппокампа изначально был больше. Также у наиболее успешных в математике детей наблюдались более крепкие нейронные связи между гиппокампом и зонами мозга, отвечающими совсем не за арифметику, как можно было бы подумать, а за формирование долговременной памяти.
Тем не менее далеко не все учёные придерживаются мнения, что способности к математике — врождённое качество, и у них есть свои аргументы. Так, например, исследование учёных из разных стран, в том числе из России, отчасти опровергло результаты питтсбургских исследователей о связи чувства числа и успехов в математике. Они протестировали 56 подростков 16–17 лет. Часть из них училась в специализированных гимназиях, а часть — в общеобразовательных школах. Все они показали одинаковый уровень чувства числа, но при этом ученики гимназий продемонстрировали значительно более высокие результаты при прохождении математических тестов.
Профессор Стэнфорда Джо Боулер, которая много лет преподавала математику в школах, написала книгу под названием «Математическое мышление», где рассказала о нескольких экспериментах, опровергающих и доводы об объёме гиппокампа. Она привела в пример исследование, проведённое с водителями такси в Лондоне, которым во время обучения нужно было запомнить все улицы города. Им проводили МРТ в начале и в конце учёбы, и оказалось, что у тех, кто прошёл обучение успешно, гиппокамп за это время значительно увеличился в размерах. Таким образом, возможно, и у детей, которые были более успешны в математике, это было не врождённое качество, а следствие интенсивного обучения (и не обязательно математике!), предшествовавшего эксперименту.
Исследование, проведённое в 2020 году в США, показало, что какими бы ни были генетические задатки, на успехи в математике в немалой степени влияет среда, в которой растёт и учится ребёнок. Так, они провели исследование на трёх тысячах американских подростков, сопоставляя их полигенные баллы (закодированные в ДНК способности к обучению), окружение и успехи в математике. Оказалось, что если подросток живёт в неблагополучной семье и учится в плохой школе, его успехи в математике тоже будут довольно плачевны — несмотря на природные задатки. И наоборот, ученики с низкими полигенными баллами, учащиеся в хороших школах и живущие в семьях с высоким достатком, показывали довольно неплохие математические знания.
С просьбой поделиться своим мнением относительно интересующего нас вопроса мы обратились к учителю математики Константину Кнопу, члену жюри математических олимпиад и популяризатору науки. Кноп ответил: «Гены на успехи в математике влияют намного в меньшей степени, чем воспитание. Я видел приёмного ребёнка, у которого были выдающиеся математические способности, которые объяснялись просто тем, что с ним много занимались и ему самому хотелось быть похожим на папу-математика.
Что касается чувства числа, то, возможно, умение считать предметы одним взглядом действительно врождённое, но связь с математикой здесь очень опосредованная. Математика — это не умение считать ни в какой степени, это умение рассуждать. И оно не связано с возможностью быстро навскидку понять, шесть перед нами коров или семь. Трудолюбие и усидчивость — гораздо более важные качества для освоения математики, чем какие-либо ещё свойства. И конечно, очень важно, чтобы родители занимались с ребёнком с рождения, а не тогда, когда они начнут замечать у него какие-то там способности».
Таким образом, учёные пока так и не пришли к выводу, существуют ли врождённые способности к математике. Однако гораздо более распространённое мнение состоит в том, что, каковы бы ни были задатки, для развития математических навыков намного важнее благоприятная жизненная среда, доступ к образованию, а также собственное желание и интерес ребёнка.

Наш вердикт: большей частью неправда
В сообществах отсутствуют спам, реклама и пропаганда чего-либо (за исключением здравого смысла)
Аудиоверсии проверок в виде подкастов c «Коммерсантъ FM» доступны в «Яндекс.Подкасты», Apple Podcasts, «ЛитРес», Soundstream и Google.Подкаст






