

Пауки-птицееды держат в своих норах домашних лягушек

Ядовитые лягушки микроквакши из семейства узкоротых защищают гнездо паука от вредителей, а паук, в свою очередь, защищает лягушку от хищников.
Ядовитые лягушки микроквакши из семейства узкоротых защищают гнездо паука от вредителей, а паук, в свою очередь, защищает лягушку от хищников.
Молодые раки-отшельники часто забирают молодую актинию, чтобы прикрепить ее к своему домику-раковине, и становятся партнерами на всю жизнь. И растут они примерно одинаково. Когда рак-отшельник перерастает свою раковину, то он забирает актинию и пересаживает на новый домик.
Симбиоз рака-отшельник и актинии выгоден для обеих сторон, по следующим причинам:
Защита.
Рак-отшельник получает дополнительную защиту от хищников, если у него на домике находится актиния. Морской анемон распространяет свои жалящие нитки над раком, в случае опасности. Благодаря этому отшельник с меньшей вероятностью будет съеден крупными хищными рыбами. А рак-отшельник, в свою очередь, также старается защитить сожительницу, избегая или сражаясь с хищниками, которые пытаются добыть прикрепленную к раковине актинию.
Питание.
А что же получает актиния, взамен на защиту отшельника? Поскольку морские анемоны потребляют практически все, что сможет поймать в море, то они не побрезгуют и лакомыми кусочками, оставшимися от питания рака. Рак-отшельник ловит ужин, а актиния подчищает за ним остатки. Это обеспечивает стабильное питание для полипа.
Мобильность.
Актинии, в основном, ведут сидячий образ жизни и передвигаются очень мало, если вообще это делают. А вот нахождение на постоянно движущемся домике рака-отшельника дает дополнительное преимущество, благодаря которому есть больше возможности поймать пищу. Пока рак-отшельник курсирует по морскому дну, анемон ловит планктон и мальков, а также расширяет ареал своего обитания.
После того, как актиния поселяется на домике отшельника, со временем подошва ее ноги разрастается и начинает покрывать всю раковину, часто начинает выступать за устье. Если же на раковине обитают несколько морских анемон, то они располагаются таким образом, чтобы уравновешивать друг друга и не свалить раковину и ее жителя.
Очень интересно происходит смена старого домика-ракушки новым. Рак-отшельник находит новую ракушку, подходит к ней близко и забирается в новый домик. Но анемонов рак не оставляет. Отшельник своими клешнями заботливо и осторожно отцепляет актинию от раковины и переносит ее на новый дом. А актиния своей подошвой прикрепляется уже на новое место. Примечательно, что актиния отсоединится от раковины только в том случае, когда ее будет отцеплять сожитель.
Рак-отшельник постоянно потребляет пищу, сраженную стрекательными клетками актинии, в результате чего у него появляется иммунитет к яду морского анемона. Впрыскивание этого яда переносится сожителем актинии безболезненно, в то же время такая доза способна убить краба Carcinus.
Некоторые раки-отшельники дорожат своими актиниями, и если последним чем-то не понравится новый домик, и они не захотят к ним прикрепляться, отшельник будет искать другие раковины и предлагать разные варианты, пока морская анемона не одобрит одну из них и не прикрепится сверху. Тогда же и рак займет раковину и успокоится.
Некоторые виды раков усаживают морскую анемону на свой домик, другие держат актинию в клешне, которой прикрывают дверь, а морские крабы, например, Lybia edmondsoni или краб-боксер, носят актинии в обеих передних клешнях и используют их для защиты от хищников, а также для охоты.

Всем привет! Сегодня во время прогулки по реке встретил этих красавцев, не мог не поделиться :) всем приятных прогулок и удивительных встреч!

В Африке живут трубкозубы (на фото), они роют норы и кушают по ночам термитов и муравьёв. Поэтому у них мощные лапы и почти нет зубов.
А ещё в Африке растёт трубкозубов огурец — это такой огурец, который запихивает свой плод под землю на глубину 30 см. Когда трубкозубу хочется пить, он не идёт к воде, он или ест термитов, или роет землю в поисках огурца.
Побывать в пузе у трубкозуба — единственный способ огурца размножиться, потому что зверь потом какает семенами. Найти огурец — единственный способ утолить жажду в сухой сезон для трубкозуба. И вот так они живут миллионы лет — трубкозуб и огурец. И больше им никто не нужен.
А кто ты в отношениях — трубкозуб или огурец?
Не моё.
Долгожитель-паразит, продлевающий жизнь хозяина.

Тихоокеанский лосось рода Oncorhynchus (сем. Salmonidae), неизбежно умирающий после первого же в жизни нереста, является ярким примером природного феномена, названного феноптозом — запрограммированной смерти особи — как способа очистки сообщества организмов от ставших лишними индивидуумов посредством включения ими программы собственной гибели. Все без исключения виды рода Oncorhynchus — кета, горбуша, кижуч, чавыча, нерка, сима и другие — массово гибнут в результате ускоренного старения (прогерии) вскоре после окончания длительной анадромной миграции из океана в реку на нерестилище и вымета икры. Их смерть не связана с перерасходом энергии, так как даже когда кета и горбуша нерестятся в совсем коротких речках — ключах, где течение едва заметно, рыбы все равно умирают в двухнедельный или месячный срок после нереста. Гибель происходит вследствие включения особой биохимической программы, в которой ключевую роль играет продукция стероидных гормонов, в частности гормона стресса кортизола. Ускоренное старение тихоокеанских лососей можно предотвратить, если у неполовозрелых рыб удалить гонады или надпочечники. Тогда жизненный цикл чавычи продлевается в два раза: с 4 до 8 лет. Биологический смысл самоубийства родителей-лососей состоит в том, что тела рыб служат источником пищи для речных беспозвоночных, которые в свою очередь поедаются маленькими лососями. Важными сигналами к включению программы прогерии служит переход лососей из морской воды в пресную, вымет половых продуктов, стрессы от конфронтации на нерестилищах.
В природе, однако, обнаруживается удивительный пример, когда биохимическая программа пострепродуктивного самоубийства атлантического лосося (семги) Salmo salar может выключиться под воздействием симбиотического организма — тканевого паразита эпителия жабр лосося — личинки пресноводной жемчужницы Margaritifera margaritifera. Тем самым предельная продолжительность жизни лососей — хозяев жемчужницы в процессе коэволюции продлевается до 13 лет, и лососи оказываются способными нерестится многократно — от 2 до 6 раз.
В данной статье на примере системы «моллюск-рыба» мы представим доказательства того, что паразит одновременно может ингибировать старение хозяина и стимулировать неспецифическую устойчивость к стрессам, т. е. регулировать продолжительность жизни. Эти многолетние данные были получены в ходе разработки биотехнологии восстановления исчезающего вида жемчужницы и приведены в табл. 1, 2.
Пресноводная жемчужница (сем. Margaritiferidae, отряд Unionoida) — наиболее долгоживущий вид из беспозвоночных животных, достигающий максимального возраста 200 лет. Палеонтологические данные указывают, что жемчужница и лососи рода Salmo (семга и кумжа) совместно эволюционировали в Европе 8 млн. лет, с плиоцена, и современный ареал моллюска вписывается в ареалы этих видов рыб.
К XXI веку в Европе несколько десятков воспроизводящихся популяций М. margaritifera остались в России, странах Фенноскандии и Шотландии. Во время изучения в разных водоемах северо-запада России особенностей развития личинок (глохидиев) жемчужницы на жабрах молоди и взрослых рыб атлантического лосося мы обратили внимание на то, что зараженные личинками производители-лососи не умирают после нереста осенью и не скатываются в море, а продолжают жить в реке без признаков прогерии до следующего лета [12]. При этом дикие лососи вынашивают на жабрах в зимний период до 2–7 тыс. мелких (диаметром 50–70 мкм) глохидиев жемчужниц на 1 рыбу. К лету у этих похудевших, но проворных рыб обычно сохраняется нормальный агрессивный рефлекс — атаковывать блесну спиннингиста.
В экспериментах в садках обнаружилось, что паразит не только не наносит заметного ущерба, например, не нарушает формулу крови и двигательную активность сперматозоидов, но и непосредственно оздоравливает своего хозяина, повышая устойчивость к неблагоприятным факторам среды. Например, у отнерестившегося "лошалого" лосося, инфицированного жемчужницей, наблюдалась неплохая выживаемость (53% по сравнению со стопроцентной летальностью интактного лосося) при таких жестких стрессах, как асфиксия (45-60 с вне воды), или термический ожог жабр от горячих пальцев человека (13%). Выше на 14% также оказалась выживаемость лососей - носителей моллюска при ранениях тела рыбы крючками.
Таблица 1. Сравнение выживаемости двух групп производителей лосося S. salar, (экспериментально зараженных и незараженных личинками жемчужницы) в садках после воздействия трех типов стрессов

Во время изучения в разных водоемах северо-запада России особенностей развития личинок жемчужницы на жабрах молоди и взрослых рыб атлантического лосося мы обратили внимание на то, что зараженные личинками производители-лососи не умирают после нереста осенью и не скатываются в море, а продолжают жить в реке без признаков прогрессии до следующего лета. При этом дикие лососи вынашивают на жабрах в зимний период до 2-7 тыс. мелких (диаметром 50-70 мкм) личинок жемчужниц на 1 рыбу. К лету у этих похудевших, но проворных рыб обычно сохраняется нормальный агрессивный рефлекс атаковать блесну спиннингиста. В бассейне Белого моря летняя экологическая форма лосося проводит, в реках один год; (с июня по июнь следующего, года), а осенняя: форма почти два года (например с августа 2002 г до июня 2004 г.). Таким образом, осенняя форма лосося может вынашивать личинки жемчужниц дважды за один визит из моря в реку. Летом обе формы производителей лососей скатываются в море и только после этой формы миграции большинство рыб в море погибает от истощения. Наиболее сильные рыбы выживают . Доля повторно, и трех-кратно размножающихся рыб составляет 10 - 40%. Отмечаются случаи 5-ти и 6-кратного нереста.

Биологический смысл подавления программы старения рыб, зараженных личинками жемчужницы состоит в следующем. Личинки растут в жабрах рыбы и увеличиваются в размерах до 10 раз. Им требуется для завершения паразитической фазы около 1500 градусодней. В холодных речках северной Европы годовая сумма градусо-дней составляет 1750, так что личиночная фаза моллюска оказывается очень длинной 300-350 cyт. В отличие от паразитического гельминта - червя лигулы физически убивающего промежуточного хозяина-рыбу чтобы попасть в окончательного хозяина - птицу, паразитическому моллюску, критически важно, чтобы жизнь хозяина не оборвалась и не оказалась короче личиночной стадии моллюска, те 8-11 мес. Жемчужница заботится чтобы лосось-хозяин (взрослый производитель и молодь-пестрятка), принявший на жабры порцию личинок осенью, не умер от быстрого старения, а прожил в здоровом состоянии как можно дольше, - как минимум до следующего лета, с тем, чтобы маленький, моллюск успел завершить свой долгий метаморфоз в жабрах рыбы, покинуть хозяина и перейти к свободному образу жизни на дне реки. За миллионы лет коэволюции моллюск-симбионт выступал как фактор движущего отбора на увеличение приспособленности и долголетия хозяина и вполне мог внедрить в геном хозяина часть своих генов долголетия , например генов, контролирующих устойчивость к голоданию или асфиксии.
Следует отметить; что у самих жемчужниц, даже старых, до недавнего, потепления климата не было отмечено болезней, паразитов и опухолей. Скорее всего особи почтенного возраста погибают не от старческих болезней, а из-за непрерывного аллометрического роста, ведущего к чрезмерному утяжелению раковины к концу жизни.
Теперь рассмотрим возможные объяснения всех этих данных, альтернативных гипотезе выключения старения. Например, можно было бы постулировать, что лососи Salmo живут дольше в Фенноскандии, чем лососи Oncorhynchus на Сахалине, потому что на севере ниже температуры среды и замедлен обмен веществ. Но этому противоречит результат акклиматизации горбуши Сахалина в Белом море в 70-х годах: в тех же жемчужно-лососевых реках (Варзуга, Умба, Кереть и др.) горбуша погибала после нереста, т.е. жизнь горбуши от низких температур не продлевалась. Значит, признак "посленерестовая гибель" у горбуши невозможно модифицировать простым понижением температуры. Можно возразить иначе: европейская жемчужница вовсе не продлевает жизнь хозяина, а просто пользуется более длинной речной фазой жизни атлантического лосося: личинки не спеша растут на хозяине, пока он жив. Но при такой трактовке образ жизни и поведение атлантического лосося после окончания размножения представляются достаточно бессмысленными: спрашивается, зачем потерявший после нереста 50% массы лосось (в пресной воде он не питается) бесцельно плавает в реке еще 8-9 месяцев? Ведь ему несомненно лучше скатиться по течению в море и там нагуляться на богатых кормом акваториях течения Гольфстрим, поскольку впереди его ждет новый нерест. Вряд ли природа допустила бы такую ненужную расточительность. Продление жизни лосося в реке приобретает вполне понятный биологический смысл, если принять, что рыба - это "суррогатная мать" для зародышей моллюска, обеспечивающая им питание, рост, защиту и расселение. В свою очередь, личинка моллюска, по-видимому, регулярно секретирует в организм рыбы вещества, ингибирующие старение и стимулирующие стрессоустойчивость. Отметим, что взрослые моллюски обеспечивают маленьких лососей укрытиями на дне реки, обрастаниями (кормовой базой) и улучшают качество воды путем биофильтрации.
Многолетние полевые исследования выявили, что не только у взрослых лососей, но и у молоди рыб жемчужница усиливает неспецифическую сопротивляемость к таким опасным заболеваниям , как опухоли-эпителиомы и грибковые поражение сапролегнией. Так, в реке Варзуга где пока еще осталось в живых несколько десятков миллионов жемчужниц и порядка 10 млн. мальков, молодь лосося обнаруживает невиданную мире высокую плотность поселений - 100-170 рыб на 100 м2 (обычная плотность 20-40 рыб). При жизни в таких перенаселенных водоёмах, лососи имеющие территориально- оборонительное поведение, должны были бы находиться в хроническом стрессе из-за территориальных конфронтации. Однако в действительности этого нет, и лососи-пестрятки хорошо уживаются друг с другом (не теряя при этом нормальной агрессивности) на нерестово-выростных угодьях без признаков истощения нервной системы. То, что личинки жемчужницы оптимизируют нейроэндокринный контроль поведения лососей, подтверждается отсутствием заболеваемости молоди лосося в главном русле реки Варзуга где обитает 90% пестряток. Здесь мальки живут в прозрачной воде среди колоний жемчужниц и практически все являются носителями глохидиев : Из просмотренных 3200 мальков за период 1997-2003 гг. при визуальном исследовании нами не было обнаружено ни одного малька с изяъзвлениями кожи, эктопаразитами, опухолями кожи и грибковыми заболеваниями. В то же время в болотистых притоках обитает примерно 10% пестряток. Здесь имеются благоприятные кормовые условия, и мальки достигают более крупных размеров, чем в главном русле. Однако вода малопрозрачна имеет коричневый "чайный"цвет и здесь, отсутствуют колонии жемчужницы. В притоках эпидемиологическая ситуация не столь идеальная - более 50 мальков из 2400 просмотренных оказались пораженными опухолями кожи и грибком-сапролегнией ( табл. 2 )

Личинки жемчужницы замедляют рост, созревание и продлевают длительность речного периода жизни молоди лосося. Рассмотрим, как это происходит на примере крупнейшей в Европе популяции атлантического лосося реки Варзуга. Специфика гидрологического режима реки (мелководность, отсутствие озер, изобилие болот в водосборе, мелкий нерестовый грунт и др) обусловила формирование стада в основном из небольших рыб, массой 2- 5 кг. Кроме того, селективный промысел неводами в ХХ в. "выбил" крупных рыб старших возрастных групп. Тем не менее у этого стада до сих пор сохраняется неожиданно сложная возрастная структура (12 возрастных категорий). Наблюдаются: следующие возрасты взрослых рыб (число лет в реке + число лет в море): 2+1+, 2+2+, 2+3+, 3+1+, 3+2+, 3+3+, 4+1+, 4+2+, 4+3+, 5+1+, 5+2+, 5+З+.; Ихтиологи давно уже обратили внимание на то, что р. Варзуга молодь лосося скатывается из реки в море при значительно меньшей длине тела (10 см), чем в соседних реках (12-14 см). Притом высказывалось удивление, почему не наблюдается элиминации таких мелких и, казалось бы, не готовых к обитанию пелалагиали моря рыб при их резком переходе из реки и море.
Многолетние данные по возрастной структуре мигрирующих летом вниз по течению посеребрившихся мальков-покатников (смолтов) указывают, что покатники из болотистых притоков скатываются в море уже в возрасте 2-3 лет, в то время как мальки из главного русла живут на колониях моллюска до 3-5 лет и только в этом возрасте покидают реку. В целом под влиянием жемчужницы средняя продолжительность жизни молодых лососей в реке до ската в море значительно больше, чем таковая в притоках (соответственно 3,3 и 2,5 г). Именно минимальный срок в 0.8 г (около 300 сут) требуется молодому моллюску, чтобы полностью завершить свою паразитическую стадию в рыбе. На некоторых крупных колониях жемчужницы среди покатников лосося доля рыб с возрастом 4-5 лет, составляет 70% и средняя продолжительность речного периода жизни лосося достигает рекордных 3,8 г.
Источник https://arctic-plus.com/nauka/280-dolgozhitel-parazit-prodle...

Однако следующий этап эволюции является ключевым для появления сложной жизни, а значит и нас с вами это первые клетки имеющие ядро - эукариоты, первые следы клеток имеющих ядро появляются в породах возрастом 1,7 миллиарда лет. Эти организмы ни что иное как симбиоз бактерий и архей. По-видимому, митохондрии произошли от альфа-протеобактерий (аэробных эубактерий), пластиды – от цианобактерий, а основная клетка – цитоплазма – от какой-то архебактерии. Митохондрии позволили адаптироваться к ядовитому кислороду и снабдили клетки более энергоэффективной системой. Хлоропласты же снабдили клетку сахарами и кислородом, таким образом единственные кто освоил кислородный фотосинтез это цианобактерии и их потомки симбионты-хлоропласты.
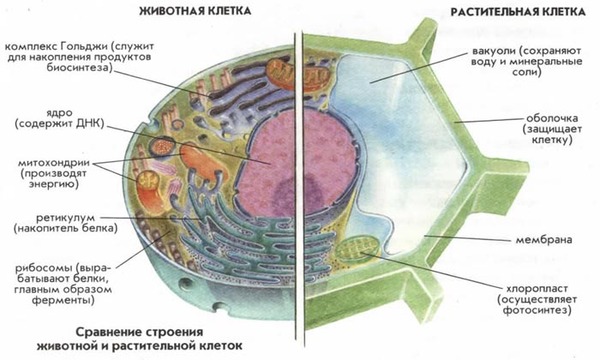
Доказательства.
Митохондрии и пластиды:
имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя — бактерий.
размножаются бинарным делением (причём делятся иногда независимо от деления клетки), никогда не синтезируются de novo.
генетический материал — кольцевая ДНК, не связанная с гистонами (По доле ГЦ ДНК митохондрий и пластид ближе к ДНК бактерий, чем к ядерной ДНК эукариот)
имеют свой аппарат синтеза белка — рибосомы и др.
рибосомы прокариотического типа — c константой седиментации 70S. По строению 16s рРНК близки к бактериальной.
некоторые белки этих органелл похожи по своей первичной структуре на аналогичные белки бактерий и не похожи на соответствующие белки цитоплазмы.
Клеточное ядро, нуклеоцитоплазма.
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предположить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. Гистоны, к примеру, обнаружены у эукариот и некоторых архей, кодирующие их гены весьма схожи. Другая гипотеза, объясняющая сочетание у эукариот молекулярных признаков архей и эубактерий, состоит в том, что на некотором этапе эволюции похожие на архей предки нуклеоцитоплазматического компонента эукариот приобрели способность к усиленному обмену генами с эубактериями путём горизонтального переноса генов.
В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза. В её основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядр
Эукариотические клетки имеют ряд важных особенностей которые позволили сделать шаг к первым многоклеточным организмам, которые будут симбионтами второй ступени.
Первое это наличие как такового ядра, которое защищает генетический материал от воздействия внешних факторов, например ультрафиолета.
Второе фагоцитоз/эндоцитоз то есть способность поглощать другие клетки, не только питаясь(фагоцитоз) ими но и превращая часть в симбионтов(эндосимбионты).
Значительная часть эукариот активные хищники, что способствует ускорению эволюции.

Существуют различные бактерии, способные развиваться в цитоплазме инфузории туфельки. Бактерии рода Caedobacter, что в переводе означает бактерия-убийца, придают инфузории-хозяину свойства киллера. Если встретятся инфузория с такими симбионтами и инфузория, их лишенная, то последняя этими симбионтами либо будет заражена, либо убита. Это зависит от состояния клеток бактерий, вышедших в среду и заглоченных незараженной инфузорией. Если это обычные вегетативные клетки бактерии, инфузория будет заражена, но она погибнет, если в этих бактериях активировались гены находящегося в них вирусного генома. Тогда бактерии перестают делиться, в них обнаруживаются вирусные частицы и крупные, преломляющие свет R-тела. Эти R-тела образованы скрученной белковой лентой, которая в вакуолях инфузории, заглотившей бактерий, может раскручиваться, разрушая окружающую вакуоль мембрану. Видимо, R-тело - одно из возможных орудий убийства лишенных симбионта туфелек. Животные, содержащие симбионтов, к R-телам нечувствительны. Очевидно, в популяции инфузорий в результате описанного эффекта должно возрастать число туфелек, содержащих симбионтов. Это скорее способствует процветанию бактерий, но едва ли полезно для инфузорий. Этот пример свидетельствует о том, что эндоцитобиоз может влиять на характер взаимоотношений между организмами.
Эндоцитобионтами инфузорий могут быть метанобразующие археи. Подобный симбиоз наблюдается у инфузорий, обитающих при отсутствии молекулярного кислорода, в кишечнике животных, илах и т.п. В таких условиях кислородное дыхание невозможно и энергетический обмен приводит к освобождению молекулярного водорода. У этих животных отсутствуют митохондрии, но в клетках имеются специальные органеллы - гидрогеносомы, которые и ответственны за образование водорода. Археи, которые могут присутствовать в клетках таких инфузорий в качестве симбионтов, окисляют водород углекислотой, накапливающейся в процессе жизнедеятельности инфузории, при этом образуется метан. Археи в результате такого сожительства получают очевидную выгоду. Для инфузории тоже полезно удаление конечного продукта энергетического метаболизма.
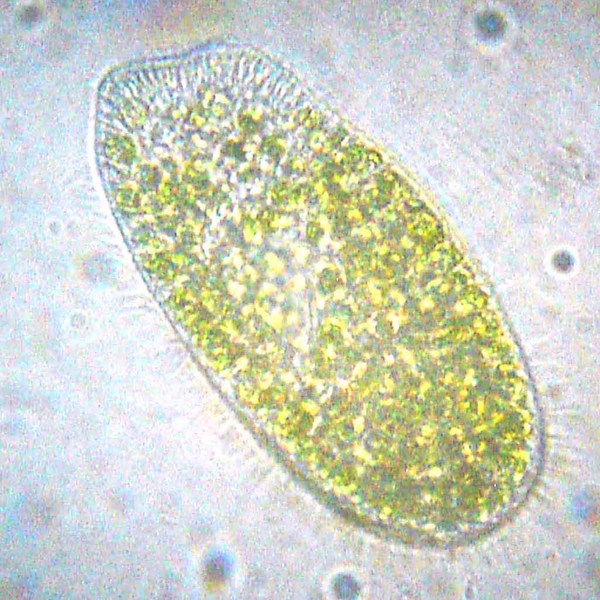
Разные группы эукариотических водорослей могут быть эндосимбионтами гетеротрофных протистов и животных. Так, в клетках инфузории-туфельки Paramecium bursaria в особых вакуолях, похожих на пищеварительные, живут симбиотические одноклеточные водоросли рода Chlorella — так называемые зоохлореллы.
Инфузория Paramecium bursaria с симбиотическими хлореллами.
Mixotricha paradoxa живёт в кишечнике термита Mastotermes darwiniensis и разлагает целлюлозу из его пищи до соединений, которые термит может усвоить.
Обладая четырьмя жгутиками, M. paradoxa для движения, тем не менее, использует 250 000 клеток бактерии Treponema spirochetes, а также 200 более крупных Canaleparolina darwiniensis, имеющих вытянутую, спиралевидную форму и прикреплённых к поверхности протиста. M. paradoxa лишена митохондрий: их функции берут на себя аэробные бактерии, живущие внутри её клетки.
Всего M. paradoxa имеет 4 эндосимбионта и являются важнейшим организмом при изучении симбиогенеза.
Третье особенности строения клеточной стенки, большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать достаточно прочную клеточную стенку из одной молекулы. Это обстоятельство могло заставить их использовать для клеточной стенки иной материал. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Прочная клеточная стенка бактерий препятствует как фагоцитозу так и появлению многоклеточных форм с общим обменом веществ, вот и остались они на стадии колоний.
Ещё одно отличие — строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15—20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотических жгутиков жгутики эукариот изгибаются или извиваются.
К так называемым заднежгутиковым относятся все многоклеточные животные и грибы, но о них следующий пост.


Строматолиты (дословно каменная прослойка) это продукт жизнедеятельности микробных матов, карбонатные постройки из минерализованных остатков их нижних слоёв.
Сами маты представляют собой симбиотические сообщества различных аэробных (использующих кислород) и анаэробных (живущих в бескислородной среде) бактерий.
Слои образованные минеральными остатками жизнедеятельности бактерий.

Верхний фотосинтезирующий аэробный слой — поверхность роста, образованная автотрофными фотосинтезирующими цианобактериями и аэробными гетеротрофами, утилизирующими кислород, выделяемый цианобактериями и органические соединения отмерших микроорганизмов.
Промежуточный слой, образованный фотосинтезирующими микроорганизмами — анаэробами, использующими прошедший через вышележащий слой свет, и гетеротрофными аэробами. В дневное время в промежуточном слое может присутствовать кислород, в ночное время при прекращении фотосинтеза условия становятся анаэробными.
Нижний слой афотическая (безсветовая) гетеротрофная анаэробная зона, составленная микроорганизмами, минерализующими органические остатки.
Срез микробного мата солевых озёр.

Микробные маты представляют собой фактически замкнутую экосистему с нулевым балансом: продукция кислорода и органических веществ равна их потреблению. Фотосинтезирующие бактерии промежуточного слоя содержат фотосинтезирующие пигменты, имеющие максимум поглощения света в спектральных областях, отличных от максимума поглощения цианобактериального хлорофилла. Такое отличие позволяет им более эффективно использовать прошедший через верхний слой свет. При изменении спектра освещённости («покраснение» утром и вечером) в мате происходят упорядоченные вертикальные миграции бактерий с разными типами пигментов.
Микробные маты в горячих источниках.

В настоящее время микробные маты обитают в самых экстремальных условиях, в которых отсутствует или слаба конкуренция со стороны более развитых форм жизни. Они способны существовать в широчайшем диапазоне температур и солёностей, и распространены от приполярных (психрофильные микробные маты Антарктики) до приэкваториальных областей (строматолиты солёных озёр и лагун Австралии) и гидротермальных источников.
Строматолиты Shark Bay Австралия.

До недавнего времени самыми древними строматолитами считались находки возрастом 3,5 миллиарда лет. В прошлом году учёные нашли древнейшие на данный момент строматолиты в формации Исуа, возрастом 3,7-3,8 миллиарда лет. До этого были только косвенные подтверждения, по изотопному составу горных пород (повышенное содержание изотопов лёгкого углерода используемого живыми организмами)
Строматолиты формации Исуа.

В течение всего архея(от 4,0 до 2,5 млрд. лет назад.) и протерозоя( от 2,5 млрд до 541 млн лет назад) они были наиболее массовыми из пород бесспорно биогенного происхождения, таким образом, являются одним из древнейших свидетельств существования и распространённости жизни на Земле.






