

Ответ на пост «Ископаемости»
Этот парень выглядит немного напряжённым.

Этот парень выглядит немного напряжённым.
Здравствуйте, камень был найден на камском устье лет 10 назад, все это время лежал дома, хотелось бы узнать что это за окаменелость, примерный возраст и возможно ценность такого камушка. Заранее спасибо.



Рис. 1. В конце раннего триаса рептилии начали завоевывать моря и быстро дали линию ихтиозавров — облигатных морских животных с рыбообразным телом, которые почти сразу превратились в страшных хищников. Здесь художник изобразил часть разнообразия ихтиозавров, отметив, что никакие мезозойские мозазавры и плиозавры не сравнятся с ихтиозаврами по свирепости и величине (аквалангистка, естественно, нарисована для масштаба). Рисунок с сайта just9art.tumblr.com
Китайские палеонтологи десять лет препарировали гигантского ихтиозавра Guizhouichthyosaurus и, наконец, завершили свою работу. Их долгая работа дала неожиданный и яркий результат: в желудке этого ихтиозавра сохранилась почти целиком его жертва, которую он проглотил незадолго до своей гибели. Жертвой 5-метрового хищника оказалась другая морская рептилия, длина которой, согласно реконструкции по костям, составляла 4 метра! Это означает, что населявшие океаны 240 млн лет назад Guizhouichthyosaurus — таков возраст находки — были сверххищниками, охотившимися на крупную добычу. Прежде считали, что их рацион составляли головоногие и рыбы, а ниша сверххищников была в то время не занята. Палеонтологам удалось воссоздать картину нападения ихтиозавра на свою жертву — он схватил и зажал ее зубами, резкими движениями головы оторвал голову и хвост, а затем целиком проглотил.
На рисунке 1 изображены некоторые представители отряда ихтиозаврид — морских рептилий, обитавших на нашей планете с конца раннего триаса (примерно 250 млн лет назад) до второй трети мела (примерно 90 млн лет назад). На сегодня в этот отряд входит около 80 родов. Среди них — гуйчжоуихтиозавр (Guizhouichthyosaurus). Этот крупный ихтиозавр, описанный 20 лет назад, мог достигать длины 7 м, обладал крупными треугольными зубами с притупленными остриями без острых краев. На основании такого строения зубов предполагалось, что гуйчжоуихтиозавры охотились на мягких головоногих и рыб среднего размера, захватывая и всасывая добычу. Но факты оказались гораздо любопытнее логичных предположений.
Китайские палеонтологи нашли в отложениях среднего триаса (242–237 млн лет назад) почти полный скелет 5-метровой особи гуйчжоуихтиозавра. Он примечателен тем, что не только прекрасно сохранился сам, но и в нем не менее прекрасно сохранились остатки трапезы гуйчжоуихтиозавра. Его добычей оказалось позвоночное животное, заглоченное незадолго до смерти самого хищника. Об этом говорит хорошая сохранность костей жертвы, не успевших раствориться в кислой среде желудка.
Скелет был найден 10 лет назад, в 2010 году, и с тех пор им занималась группа ученых из нескольких научных учреждений Китая, а также их коллеги из Калифорнии, Милана, Чикаго. Эти 10 лет ушли на препарирование скелета самого ихтиозавра и разбор плотно упакованных в желудке остатков его жертвы. Долгая работа того стоила.

Рис. 2. Скелет гуйчжоуихтиозавра (A) и его желудок (B), в котором ученые нашли остатки проглоченной жертвы. Рисунок из обсуждаемой статьи в iScience
По костям жертвы удалось определить ее видовую принадлежность (сравнив с голотипом, для которого имеется почти полный скелет) — это Xinpusaurus xingyiensis, морская рептилия из группы талаттозавров. Более того, по размеру костей смогли реконструировать размер этого талаттозавра: он имел длину около 4 метров.
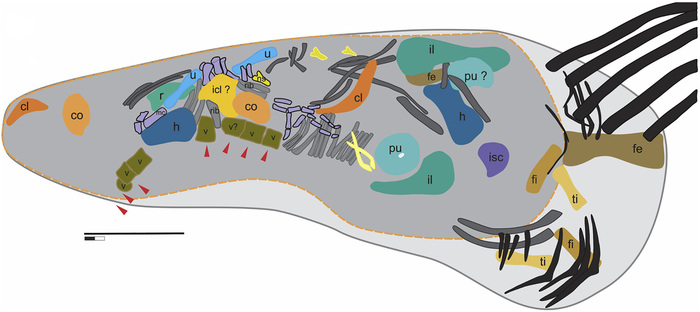
Получается, что 5-метровый Guizhouichthyosaurus напал на рептилию почти такого же размера, как и он сам. Так что рацион этих ихтиозавров оказался совсем не похож на мягкую, податливую кашу из головоногих и рыб. Монстр набросился на длинную узкую рептилию, ухватил ее за туловище и, зажав длинными, похожими на крокодильи, зубами, резкими движениями оторвал голову и хвост и проглотил оставшееся туловище целиком. Хвост жертва могла отбросить и сама, но вот голову она могла потерять только метафорически. Хвост жертвы, кстати, нашелся неподалеку, в 13 метрах, от места сражения и, очевидно, от места гибели самого хищника.
Авторы работы, рисуя красочную картину этого нападения, все же задаются естественным вопросом — возможно, этот ихтиозавр был падальщиком, а не хищником, и ему просто повезло найти такой роскошный обед. Против гипотезы «падальщика» авторы приводят несколько доводов. Главный из них — присутствие костей конечностей жертвы, а это первое, что отваливается при разложении тела рептилии, голова отваливается в последнюю очередь. Второе возражение касается приповерхностного обитания ихтиозавров. Они все же дышат воздухом, так что вряд ли приспособлены к поиску отмерших на дне остатков.
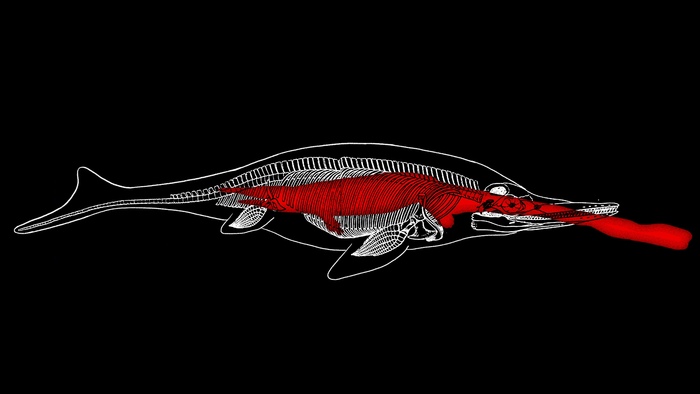
Таким образом, перед нами фактический материал, наглядно демонстрирующий, что в среднем триасе уже существовали сверххищники, которые активно охотились на самую крупную добычу. В те времена ими были ихтиозавры: кроме гуижоуихтиозавра, судя по строению зубов, эту нишу могли занимать Besanosaurus и Cymbospondylus. В наши дни ее занимают акулы и касатки. Прежде считалось, что в среднем триасе эта ниша была еще не занята.













Тушка колибри рядом с отпечатком гигантского вымершего муравья титаномирма (Titanomyrma lubei) из нижеэоценовых (~50 млн лет) отложений Вайоминга.

125-миллионолетняя окаменелость, сохранившая мельчайшие детали. Образец рыбы Rhacolepis buccalis из меловой формации Сантана, Бразилия.

Отпечаток мягких тканей ихтиозавра, сохранённый вместе с окаменелыми остатками переднего плавника.

50-миллионолетняя плита с отпечатками стаи рыбок, плывущих в одном направлении. Окаменелость проливает свет на наличие стайного движения у различных групп живых существ.

Отпечаток юрской рыбы под названием гиродус (Gyrodus), которая имела полный рот плоских зубов, идеально приспособленных для дробления раковин аммонитов и ракообразных.

Пчёлы – ровесники динозавров. Они производят мёд ещё с Мелового периода (100 миллионов лет назад). Древние пчёлки, застывшие в янтаре, имеют очень небольшие размеры. Судя по всему, цветы, на которых кормились эти насекомые, тоже были очень маленькими. Палеонтологи полагают, что именно пчёлы, приспособившиеся к питанию нектаром цветов, вызвали бурный расцвет цветковых растений в Меловом периоде. До появления пчёл опылением цветов могли заниматься только мухи и жуки, которые, конечно, играли определённую роль в размножении цветковых растений, но пчёлы, если можно так выразиться, вывели процесс опыления на новый уровень.

Отпечаток кальмара Plesioteuthis sp. из знаменитых золенгофенских сланцев позднеюрского возраста (~150 млн лет), Золенгофен, Германия.


В 2012 году в Белоруссии в шахте по добычи соли на глубине 620 рабочими был обнаружен отпечаток ракоскорпиона. Остатки членистоногого были извлечены из массива и доставлены на поверхность. По словам главного геолога предприятия Д. Барбикова, это уже шестая подобная находка за всю историю деятельности предприятия. Первая была в 1976 году, вторая — в начале 80-х, третья — в 2002-м, четвертая — в 2006-м (тогда была обнаружена только одна конечность), а пятая — в 2008 году. Найденный ракоскорпион — рекордсмен: его длина составляет 34 см. Предыдущий его "собрат" был короче на 9 см. В вырезанном куске породы он занял свое место среди экспонатов Музея трудовой славы "Беларуськалия". А в прошлом году исследователями Д. Плаксом, Дж. Ламсделлом, М. Вразо и Д. Барбиковым этот экземпляр был описан под новым родовым и видовым названием Soligorskopterus tchepeliensis (смотреть статью к посту). Эти древние морские отложения в которых были обнаружены ракоскорпионы морского происхождения и относятся солигорской формации среднего фамена (верхний девон) и имеют возраст около 363 млн лет. Ну и этот год уже стал урожайным на ракоскорипонов в этой шахте (четвертая фотография). Он, конечно, меньше по размерам, чем найденный в 2012 году и относится к описанному виду, но дополняет сведения о разнообразии этих древних животных.

Отпечаток Marrella splendens, вымершего членистоногого из класса Marrellomorpha. Миниатюрные придонные организмы, чьи останки доминируют в отложениях среднего кембрия сланцев Бёрджес (Британская Колумбия, Канада).

Окаменелый череп Ludodactylus с листом юкки, застрявшим в нижней челюсти. Вероятно, этот лист стал причиной гибели животного от голода.

Туллимонстр (Tullimonstrum) — род вымерших водных хордовых. Удлинённое тело без конечностей отличалось широко расставленными глазами на стебельках, длинным хоботом на голове, который заканчивался челюстями, и ромбовидным хвостовым плавником. Размер представителей колеблется в пределах 8—35 см.
Отпечаток туллимонстра в камне
Первая часть родового названия — фамилия первооткрывателя Francis Tully, вторая часть — лат. monstrum — «чудо, диво».

Найден в среднекаменноугольных отложениях (311—307 млн лет назад) дельты реки, существовавшей в то время на месте современной реки Мазон-Крик (Иллинойс). Там же встречаются остатки других животных, сохраняющихся лишь в исключительных условиях, в том числе медуз, мальков рыб и икры.
В 2016 году туллимонстр, имевший клешнееобразные челюсти на конце длинного хоботка, был отнесён к миногообразным бесчелюстным.

Следовая дорожка дипротодона (Diprotodon optatum). Поздний плейстоцен (около 100 тыс. л. н.) штата Виктория, Австралия. Дипротодон (лат. Diprotodon) — крупнейшее известное сумчатое, когда-либо обитавшее на Земле. Дипротодон принадлежит к так называемой австралийской мегафауне — группе необычных видов, живших в Австралии в период приблизительно от 1,6 миллионов до 40 тысяч лет назад. В 2019 году исследователи из Ирландии, США и Японии обнаружили, что «монстра Талли», названного в честь своего первооткрывателя, могли напрасно отнести к позвоночным. Статью об этом авторы выпустили в издании Proceedings of the Royal Society B. Исследователи проанализировали ископаемый образец глаза «монстра Талли», найденный в Мэйзон-Крик, штат Иллинойс. Ученые также сравнили металлы меланосом Tullimonstrum с металлами меланосом его современников-позвоночных, найденных в тех же породах и в той же местности, чтобы исключить какие-то случайные аномалии. Оказалось, что ископаемые меланосомы из глаз Tullimonstrum содержат мало цинка и много меди по сравнению с меланосомами этих окаменелостей позвоночных. Сведенные воедино, результаты показывают, что «монстр Талли» с высокой вероятностью мог быть беспозвоночным.

Редчайший отпечаток юрского белемнита Passaloteuthis bisulcata.

Отпечаток кожных покровов трицератопса

В Аравийской пустыне в 2012 году были обнаружены следы древних хоботных, прошедших по грязи около 7 млн лет назад. Судя по отпечаткам, стадо вело себя вполне современным образом: взрослые мужские особи шли своей собственной тропой, а остальные следовали за лидером женского пола. Иллюстратор исходил из предположения, что следы оставили представители вида Stegotetrabelodon syrticus. На переднем плане — самка-вождь. (Изображение Mauricio Antón.).


Раннеюрские морские плезиозавры Rhomaleosaurus. Их ареал охватывал оба полушария и простирался до полярных морей на территории современной Якутии.

Выполненное российскими палеонтологами тщательное исследование остатков морских рептилий, собранных около полувека назад по берегам якутской реки Вилюй, выявило их большое таксономическое разнообразие. Оказалось, что приполярные моря юрского периода населяли несколько родов ихтиозавров, плиозавров и плезиозавров. Среди находок было много костей молодых рептилий, в том числе, возможно, принадлежащие новорожденным особям. Это позволяет предположить, что морские рептилии юрского периода использовали сибирские моря как ясли и приплывали туда летом для размножения.
Остатки морских рептилий первой половины юрского периода (201–174 млн лет назад) очень неравномерно представлены в разных регионах планеты. Почти все наши знания о них ограничены находками из Великобритании и Германии. Другие регионы дали лишь несколько экземпляров, причем в основном это всего лишь разрозненные и плохо сохранившиеся кости.
Такая ситуация объясняется прежде всего человеческим фактором и географическими особенностями. Карьеры Хольцмадена (Holzmaden) в Германии и знаменитые береговые обрывы британского курорта Лайм-Риджис (Lyme Regis) привлекают внимание геологов, палеонтологов и любителей уже две сотни лет. Они легко доступны, в них ведется постоянный мониторинг, который регулярно приносит новые интересные находки. Таких условий нет в других местах, где вскрываются раннеюрские морские отложения, — в том числе в безлюдной тайге Якутии. Тем не менее даже при несопоставимом числе находок эти регионы могут дать важную информацию для понимания исчезнувших экосистем.
В журнале Geological Magazine недавно вышла статья российских палеонтологов с описанием остатков морских рептилий из Якутии. Ископаемые кости были собраны еще полвека назад в отложениях ранней юры, которые вскрываются по обрывам реки Вилюй и охватывают интервал продолжительностью в 25 миллионов лет (190–175 млн лет назад).

Обнажения раннеюрских пород на обрывистых берегах Вилюя.
Коллекция состоит из 150 костных остатков. Почти все они — отдельные позвонки и кости конечностей. Наиболее целые находки: фрагмент позвоночного столба и остатки челюстей двух ихтиозавров.
Большинство остатков сложно определить даже до семейств, но некоторые получилось идентифицировать до родов. К примеру, на обломке морды крупного ихтиозавра сохранились отпечатки нескольких коронок зубов (сами они разрушились), на одном отпечатке удалось заметить режущую кромку. В юрском периоде такие кромки были только у ихтиозавров Temnodontosaurus. Представители этого рода были главенствующими хищниками раннеюрских морей, достигая длины десяти метров. Судя по фрагментам челюстей, якутский ихтиозавр был примерно семиметровым.
По схожим мелким деталям удалось определить и некоторые другие остатки: небольших ихтиозавров Stenopterygius, длинношеих плезиозавров Microcleidus, а также плезиозавров из семейства ромалеозаврид (Rhomaleosauridae) и примитивных плиозаврид.
Все эти животные давно известны по находкам в раннеюрских отложениях Западной Европы, а также в некоторых других регионах. Например, Temnodontosaurus недавно был обнаружен в пустыне Атакама в Южной Америке. Подобным космополитизмом отличаются многие морские животные, в том числе вымершие, но не все.
В Восточной Якутии не оказалось морских крокодиломорф талаттозухий, широко представленных в Западной Европе. Можно предположить, что им, подобно другим крокодиломорфам, не удалось заселить полярные регионы из-за климатических ограничений.
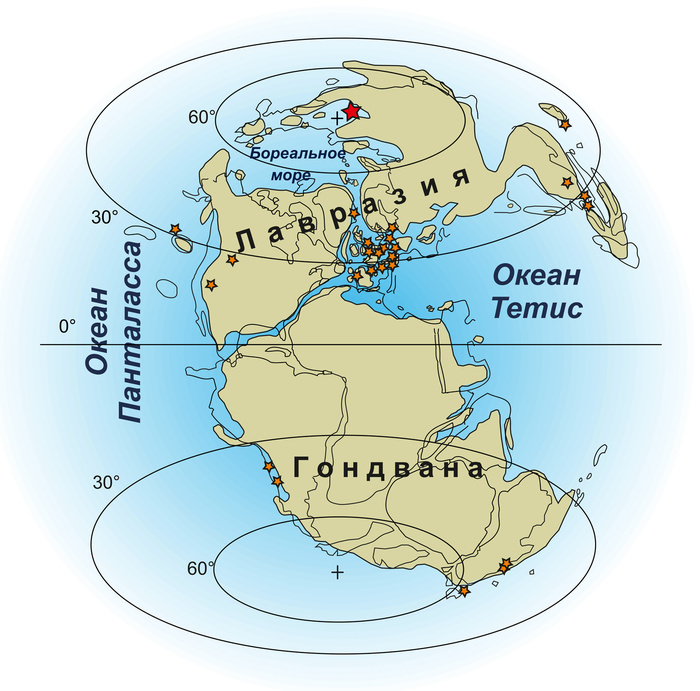
Палеогеографическая карта начала юрского периода. Звездочками отмечены местонахождения остатков морских рептилий. Полюса Земли обозначены крестиками. Красная звездочка возле северного полюса — якутские местонахождения. Климат этого времени для высоких широт пока изучен недостаточно хорошо, но считается, что он был достаточно прохладным (см. новость Мезозойские морские рептилии Сибири жили и в теплых , и в холодных водах, «Элементы», 16.07.2019). Рисунок из обсуждаемой статьи в Geological Magazine
Вилюйские местонахождения сформировались в мелководных морях к северу от восьмидесятой параллели, то есть даже ближе к северному полюсу, чем они расположены сейчас. Это самые высокие широты для находок ранних морских рептилий.
Изобилие ихтиозавров и плезиозавров в прохладных водах подтверждает мнение об их теплокровности (см. новость Мезозойские морские рептилии Сибири жили и в теплых, и в холодных водах, «Элементы», 16.07.2019). Кроме того, для обитания в полярных водах у них могли быть и другие адаптации, например, толстый слой жировой ткани, недавно описанный для раннеюрского ихтиозавра Stenopterygius (J. Lindgren et al., 2018. Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur).
В коллекции из Якутии оказались и остатки молодых плезиозавров. Судя по мелким размерам и плохому окостенению, возможно, что это кости даже младенческих особей. По словам одного из авторов статьи, Николая Зверькова, молодым особям принадлежит четверть найденных позвонков и примерно половина костей конечностей (шесть от молодых особей и семь от взрослых). Самая маленькая кость сохранилась почти целиком: в длину она достигает 10 сантиметров, ее обладатель даже с учетом длинной шеи едва ли превышал в длину 1–1,5 метра, в то время как крупные кости принадлежали более чем пятиметровым плезиозаврам.
Остатки молодых особей показывают, что морские рептилии (по крайней мере плезиозавры) использовали высокие широты как место для размножения. Подобное поведение описано для морских рептилий мелового периода, которые поднимались для размножения в высокие широты.
Летом в высоких широтах световой день длится долго (а во время полярного дня Солнце не заходит вообще). Сейчас это вызывает бурный рост фитопланктона и — далее по пищевой цепочке — увеличение численности беспозвоночных, привлекающих рыб и китообразных, которые приплывают в северные воды покормиться и вывести потомство. В юрских морях ситуация могла быть похожей, только вместо китообразных на вершине пищевой пирамиды находились морские рептилии.
Пока не удалось выяснить, обитали ли юрские морские рептилии в полярных морях круглый год или приплывали туда только на летний сезон. Авторы обсуждаемой статьи склоняются ко второму варианту, приводя следующие два аргумента. Во-первых, мигрируют почти все крупные морские позвоночные — в поисках пищи и к местам размножения. Черепахи, например, мигрируют на низких широтах, а киты мигрируют по всей планете (в том числе — и в высокие широты). Во-вторых, у морских рептилий в случае постоянного обитания в полярных морях должны были появиться адаптации к многомесячному существованию в условиях полной темноты (например, эхолокация, ночное зрение или какие-либо сенсорные органы). Такие адаптации сложно уловить на ископаемом материале, однако представляется маловероятным, что эти адаптации массово появились у нескольких родов ихтиозавров и плезиозавров, которым было проще не приспосабливаться к суровым условиям полярной ночи, а уплывать в более южные моря.
Кроме того, в отложениях зафиксирован как минимум один серьезный эпизод похолодания с вероятным оледенением. А покрытые льдом моря не могли подходить для морских рептилий, дышащих атмосферным воздухом.
Более правдоподобным выглядит сценарий, по которому ихтиозавры и плезиозавры на лето отправлялись в полярные регионы, размножались там, а с наступлением осени плыли обратно на юг. Так сейчас ведут себя многие перелетные птицы. Ну а в юрском периоде на территории нынешней Якутии плавали «перелетные» плезиозавры.
https://elementy.ru/novosti_nauki/433753/V_yurskom_periode_p...
Справились? Тогда попробуйте пройти нашу новую игру на внимательность. Приз — награда в профиль на Пикабу: https://pikabu.ru/link/-oD8sjtmAi

Креодонты (Creodonta Cope, 1875) — вымерший отряд высших млекопитающих (Eutheria), имеющих общее происхождение с настоящими хищными (Carnivora). Эти животные обладали архаичной морфологией и питались сугубо животной пищей, о чем свидетельствует и их название — «мясозубы» («creo» в переводе с древнегреческого означает мясо, а «dens» на латинском — зуб). Оформившись к концу палеоцена и пережив два крупных этапа адаптивной радиации (на рубеже палеоцена и раннего эоцена и в конце среднего – начале позднего эоцена), группа просуществовала до конца миоцена; ее представители встречались по всему Северному полушарию, а также в Африке. На сегодняшний день в отряд включается 2 семейства с приблизительно 80 родами и более чем 180 видами.

История открытия и изучения
Впервые ископаемые остатки креодонтов были обнаружены в 20-е годы XIX в. в позднеэоценовых отложениях Парижского бассейна. Во второй половине XIX и в начале XX в. последовали многочисленные находки из стран Западной Европы и США, были описаны десятки новых родов и видов. Первые находки креодонтов в Азии были сделаны в миоценовых отложениях Сивалика и формации Нагри. Основную массу наиболее информативных находок удалось получить в ходе работ Центрально-Азиатской Американской экспедиции (1920-30 гг.).
Длительное время ранние находки черепных фрагментов из Франции, принадлежащих Hyaenodon и Pterodon, относили к сумчатым (Marsupialia) или к хищным (Carnivora). Мнение о принадлежности к Eutheria утвердилось благодаря работам Г.Фийоля (1874). Э.Коп (1875) предложил название Creodonta, присвоив им подотрядный ранг в составе насекомоядных (Insectivora). М.Шлоссер (1887) повысил ранг Creodonta до отрядного, включив в его состав, кроме креодонтов (Oxyaenidae, Proviverridae), еще 3 семейства (Arctocyonidae, Mesonychia, Amblictonidae). Вортман (1901) выделил Creodonta в качестве подотряда в составе Carnivora. У.Мэтью (1902, 1909) понимал Creodonta более широко. В современном понимании объема таксона, Creodonta соответствовали выделенному им семейству Pseudocreodi. В дальнейшем объем и ранг креодонтов неоднократно пересматривался (Осборн (1910), Грегори (1939), Симпсон (1945), Ромер (1966) и др.). Большинство исследователей в разработке систематики плотоядных млекопитающих использовали в качестве диагностических признаки строения зубного аппарата. Однако, как относительно недавно было показано, они приобретались конвергентно, в процессе специализации челюстного аппарата. Современный ранг и объем креодонгам придал М. Мак Кенна (1975), выделив их как отряд в составе cyперотряда Creofaga.

Саркастодон монгольский.
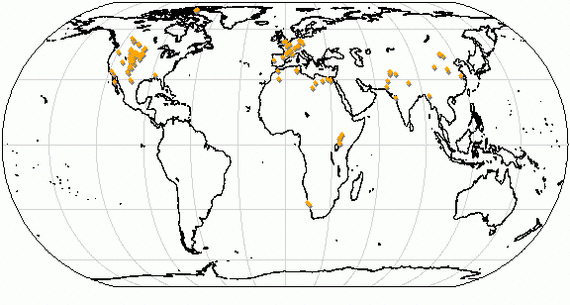
Карта основных находок ископаемых остатков Creodonta.
К сожалению, большинство креодонтов известны только по отдельным костям и фрагментам черепов. Их скелеты исключительно редки. Однако до нас все же дошли полные остатки американских оксиенид (Oxyaena, Patriofelis) и гиенодонтид (Sinopa, Tritemnodon, Hyaenodon). Особенно многочисленны остатки Hyaenodon, что свидетельствует о этом роде как об одной из наиболее массовых форм палеогеновых хищников.
Происхождение
Сегодня большинство специалистов рассматривает таксоны Creodonta и Carnivora в качестве сестринских и определяет их в надотряд Ferae (впрочем, согласно иной точке зрения они могут принадлежать к Lypothiphla). Так или иначе, не вызывает сомнения, что предками Creodonta и Carnivora являются пока неустановленные примитивные Eutheria. Однако насколько близко друг к другу находятся эти отряды, до конца неясно. Из предполагаемых синапоморфий Creodonta и Carnivora можно упомянуть наличие костного тенториума (перегородки, отделяющей мозжечок) и некоторых общих черт в морфологии основания черепа и предплюсны; указывалось также на присутствие синапоморфий в строении посткраниального скелета Creodonta и примитивных хищных Miacidae. Все же, убедительно объяснить наличие общих морфологических признаков у представителей обоих отрядов посредством синапоморфий пока еще не удалось.
Предполагается, что Creodonta дивергировали от эволюционной ветви Carnivora еще в позднем мелу, в дальнейшем обе группы хищников эволюционировали параллельно. Современные Carnivora морфологически более удалены от общей предковой предковой группы, чем Creodonta. По комплексу признаков линия Creodonta и Carnivora противопоставляется Didelphodonta. Появившись приблизительно в одно время с Carnivora, Creodonta длительное время конкурировали с ними, зачастую добиваясь большего успеха благодаря быстрой дифференциации, но, в конце концов, полностью исчезли.
Таксономия
В состав Creodonta входит 2 семейства: Oxyaenidae (11 родов) и Hyaenodontidae (69 родов). Семейства отличаются друг от друга по строению зубного и челюстного аппаратов, стопы, голеностопного сустава и когтевых фаланг. Hyaenodontidae были более адаптивны и многочисленны, особенно в Старом Свете; как правило, представители этой группы имели собакоподобный или гиеноподобный облик и были лучше приспособлены к активному передвижению. Oxyaenidae, получившие распространение преимущественно в Северной Америке, в большинстве своем были относительно менее крупными и обладали удлиненным телом на коротких стопоходящих конечностях, напоминая по своему сложению енотов, медведей и кошек.
Необходимо заметить, что на сегодняшний день вопросы о становлении двух ветвей креодонтов и ранних этапах их эволюции окончательно не выяснены и являются предметом дискуссий. Ряд исследователей ставит под сомнение монофилитическое происхождение Hyaenodontidae и Oxyaenidae и объединение их в один отряд. По их мнению, наличие общих морфологических черт у представителей обоих семейств может объясняться их независимым происхождением о цимолестоподобных предков. Все же имеющиеся данные скорее подтверждают, чем опровергают общность происхождения креодонтов.
Один из современных вариантов классификации креодонтов (Лавров, 1999) имеет следующий вид:
Отряд Creodonta Соре, 1875
Семейство Oxyaenidae Соре, 1877.
Подсемейство Tytthaeninae Gunnell, Gingerich, 1991.
Подсемейство Palaeonictinae Denison, 1938
Подсемейство Oxyaeninae Cope, 1877
Семейство Hyaenodontidae Leidy, 1869
Подсемейство Proviverrinae Schlosser, 1886
Триба Proviverrini Schlosser, 1886
Триба Teratodontini Lavrov, 1999
Подсемейство Limnocyoninae, Wortman, 1902
Подсемейство Machaeroidinae Matthew, 1909
Подсемейство Pterodontinae Polly, 1996
Триба Apterodontini Szalay, 1967
Триба Francotheriini Lavrov, 1999
Триба Metapterodontini Lavrov, 1999
Триба Pterodontini Lavrov, 1999
Подсемейство Hyaenodontinae Leidy, 1869
Триба Epipterodontini Lavrov, 1999
Триба Hyaenodontini Leidy, 1869
Ранние представители креодонтов объединяли в своей морфологии архаичные признаки как посткраниального скелета Oxyaenidae, так и зубного аппарата Proviverrinae (Hyaenodontidae). Не позже первой половины палеоцена в Азии сформировалась предковая форма Oxyaenidae и Hyaenodontidae, сочетавшая в своей морфологии зубную систему Proviverrinae (Hyaenodontidae) и архаичное плантиградное строение автоподия Oxyaenidae. Oxyaenidae дивергировали в палеоцене от предковой группы Hyaenodontidae, архаичное строение автоподия которых они унаследовали и сохранили на протяжении всей истории семейства. Уже у самых ранних Oxyaenidae зубной аппарат был достаточно специализирован. У Hyaenodontidae, напротив, в отличие от Oxyaenidae, исходное строение зубного аппарата было примитивное, и в процессе эволюции происходили прогрессивные изменения как зубного, так и локомоторного аппаратов. Такое комплексное развитие обусловило большую экологическую пластичность Hyaenodontidae и позволило успешно освоить новые территории и ландшафты.
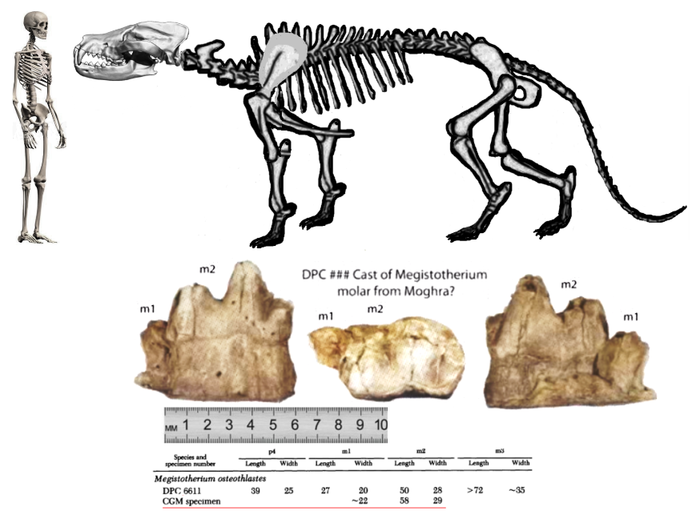
Megistotherium (Hyaenodontidae) из формации Moghara, Египет.

Скелет Hyaenodon (Neohyaenodon) macrocephalus. Найден в верхнеоэценовых отложениях на юго-востоке Монголии. Возраст находки – около 35 миллионов лет. Неогиенодон – представитель группы древних плотоядных млекопитающих из вымершего отряда креодонтов. Фото сделал я в Палеонтологический музей им Ю. А Орлова. Кстати кто не был советую.
Распространение и эволюционная история
Установлено, что центром происхождения креодонтов была Азия. Вероятно, они появляются здесь не позднее середины палеоцена, однако древнейшие известные на сегодняшний день находки — Oxyaenidae gen. et sp. indet. (Oxyaenidae) и Proviverrinae gen. et sp. indet. (Hyaenodontidae) — происходят только из позднего палеоцена Монголии (свита Наран-Булак). Возможно, им синхронна древнейшая находка североамериканского Oxyaenidae — Tytthaena (средний тиффаний, зона Ti5). Proviverrinae gen. et sp. indet. обладает специализированными туберкуло-секториальными зубами и не может быть принят в качестве неспециализированного предка Oxyaenidae. В позднем палеоцене Oxyaenidae проникли в Северную Америку и распространились здесь. Они, одновременно с Proviverrinae (Hyaenodontidae), в раннем эоцене достигли Европы (Palaeonictis gigantea и Prototomus sp. соответственно, зона МР-7), но в Африку не проникли.
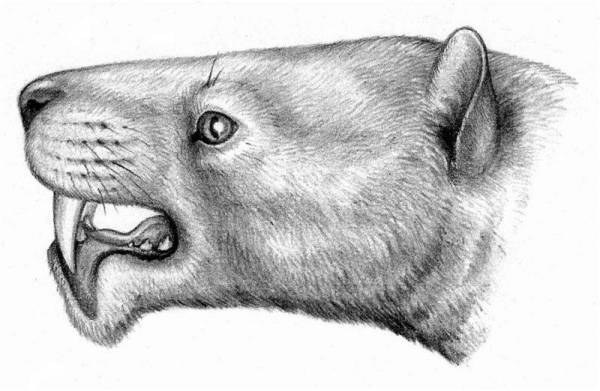
Портретная реконструкция Oxyaena lupina (семейство Oxyaenidae). Ранний эоцен, Северная Америка.
Появление креодонтов в Европе и Северной Америке происходило синхронно с появлением млекопитающих других отрядов. Так, с креодонтами в палеоцене и раннем эоцене в Северную Америку из Азии вселяются представители Mesonychia, Rodentia, Primates, Artiodactyla. Вселение в Азию из Северной Америки происходило в меньших масштабах, что, вероятно, объясняется меньшей конкурентоспособностью североамериканской фауны в раннем палеогене.
Oxyaenidae в Азии представлены палеоценовым "Охуаепа" sp. и специализированным гигаетским родом Sarkastodon (средний эоцен). Архаичное плантиградное строение автоподия Oxyaenidae, по-видимому, предопределило их меньшую конкурентноспособность по сравнению с дигитиградными Hyaenodontidae и Mesonychia. Можно полагать, что, сформировавшись в Азии, Oxyaenidae не были здесь многочисленны в палеоцене. Только после вселения в Северную Америку семейство смогло освоить широкий спектр экологических адаптаций.
В раннем эоцене в Европе Oxyaenidae вымерли (зона МР-9). Вселение Hyaenodontidae в Северную Америку произошло в раннем уосатче (Wa0), на рубеже палеоцена и эоцена (появление Proviverrinae и дивергировавших от них уже в Америке Limnocyoninae). Первый пик разнообразия креодонтов отмечен в раннем и среднем эоцене (уосатч и бриджерий) и связан с переседение из Азии в Северную Америку Oxyaenidaeи части Hyaenodontidae (Limnocyoninae и Proviverrinae). В начале среднего эоцена от Limocyonionae обособляются Machaerodinae (эндемичны для Северной Америки). К середине бриджерия исчезают Limnocyoninae. К концу века вымирают азиатские и американские Oxyaenidae, что приводит к мощному всплеску разнообразия Hyaenodontidae. В среднем эоцене Hyaenodontidae достигают Африки — Koholia (Proviverrinae).
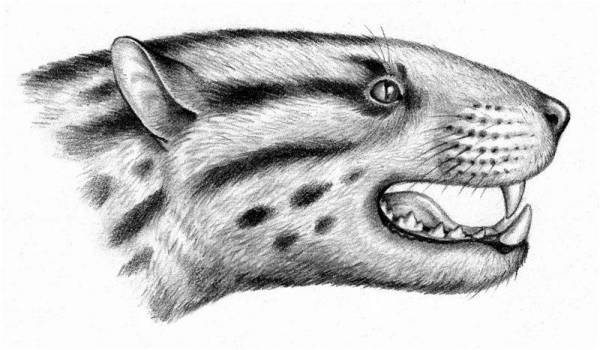
Портретная реконструкция Apataelurus kayi (семейство Hyaenodontidae). Средний эоцен, Северная Америка.
Появление Pterodontinae произошло в начале среднего эоцена в Азии (Isphanatherium, Neoparapterodon sp., нижнеалайская подсвита). Pterodontinae вселились в Северную Америку из Азии в начале среднего эоцена, предположительно не позднее средней уинты (начала среднего эоцена), до момента прекращения существования берингийского моста. Из Америки подсемейство по северо-атлантическому мосту проникло в Европу, где его появление фиксируется в зоне MP-11 Гейзейталя (Oxyaenoides). В Африке Pterodontinae известны с позднего эоцена (Apterodon).
В Азии Hyaenodontinae появляются во второй половине среднего эоцена (формация Шара-Мурун): в Северной Америке — с раннего дюшезния (конец среднего - начало позднего эоцена); в Европе Hyaenodontinae известны с зоны MP-17 (поздний эоцен), куда они вселяются из Азии. В Африке находки подсемейства не известны. В конце среднего - начале позднего эоцена происходит бурная адаптивная радиация Hyaenodontinae. Только в раннем чадроне Северной Америки известно 8 видов Hyaenodontini: от сверхкрупного Neohyaenodon gigas до карликового Microhyaenodon venturae. В Европе обитало 11 видов. В позднем эоцене наблюдается вспышка разнообразия Pterodontinae, из них описано 8 родов в Европе, 6 родовв Азии и 5 родов в Африке (в позднем олигоцене-миоцене отсюда в общей сложности известно 7 родов), в Северной Америке — 2 рода.
С середины эоцена среди Hyaenodontidae появляются животные, способные охотиться на достаточно большую добычу, не уступавшую в размерах им самим. Не исключено, что это имело непосредственную связаь с повсеместным сокращением численности и разнообразия древних копытных Mesonychia, до этого прочно занимавших ниши крупных наземных хищников. К концу эоцена некоторые Hyaenodontidae уже достигают гигантских размеров (Hemipsalodon, Neohyaenodon).

Портретная реконструкция Megistotherium osteothlastes (семейство Hyaenodontidae). Ранний и средний миоцен, Африка.
В позднем эоцене происходит угасание Proviverrinae. Они доживают до конца олигоцена - начала миоцена только в Африке и Южной Индии. Но в Африке в это время обособляется триба Teratodontini, представляющая собой уклоняющееся направление эволюции креодонтов — специализацию в склерофагии. В раннем олигоцене вымирают все Pterodontinae Северной Америки и Евразии (помимо наиболее крупного Hyainailouros), несколько уменьшается разнообразие Hyaenodontinae.
В эоцене и на рубеже эоцена и олигоцена отмечается похолодание, в Центральной Азии нарастает аридность климата. В олигоцене изменяется спектр потенциальных пищевых объектов креодонтов: сокращается численность и разнообразие Perissodactyla, происходит радиация Bovidae (Artiodactyla). В этих условиях в Северной Америке в конце олигоцена креодонты полностью вымирают. В Европе до начала миоцена (зона MN-2) доживает только Hyainailouros (Pterodontinae), а в Азии — крупные гиенодоны (Neohyaenodon gigas, N. macrocephalus, Hyaenodon pervagus — горизонт соответствует европейским зонам MN-3-4) и гигантский Hyainailouros. Дольше всего они удерживаются в Индии и Восточной Африке (датировки К-Аr методом дают 11-14 млн. лет), откуда происходят находки последних, наиболее крупных представителей родов Megistotherium, Hyainailouros и Dissopsalis.
Морфология
В ходе эволюции креодонтов реализовались различные стратегии достижения высокого уровня специализации и сформировались специализированные хищники всех размерных классов. Среди них встречались как карлики величиной с куницу, так и гиганты, способные соперничать своими размерами с самыми большими медведями (длина черепа — от 50 мм у Lesmesodon до 670 мм у Megistotherium). С течением времени представители обоих семейств становились крупнее. Морфологическая организация креодонтов обладала рядом прогрессивных особенностей, но в общих чертах оставалась достаточно примитивной, сохраняя много архаических черт, сближающих с насекомоядными мезозойскими предками.
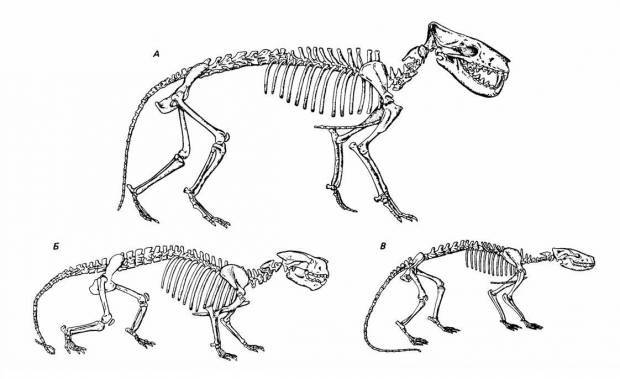
Реконструкция скелета Creodonta: А — Hyaenodon (Hyaenodontidae); Б — Patriofelis (Oxyaenidae); В — Sinopa (Hyaenodontidae).
Тело креодонтов было приземистым и удлиненным, а конечности относительно короткими и мощными, практически всегда пятипалыми. Ключица как правило хорошо развита. Плечевая кость в большинстве случаев с внутренним, а иногда и со средним надмыщелковыми отверстиями; локтевая свободная, немногим тоньше лучевой, лучевая — с нерасчлененной верхней поверхностью. Ладьевидная, полулунная и центральная кости запястья почти всегда раздельны. Бедренная кость большей частью с третьим вертелом. Астрагал имеет заднее отверстие; его блок с неглубокой бороздой; имеется ясная косо направленная шейка, выпуклая головка сочленяется только с ладьевидной костью заплюсны, изредка также и с кубовидной. Ладьевидная и кубовидная кости заплюсны раздельны.
Стопа креодонтов (особенно у Oxyaenidae) устроена крайне архаично. Oxyaenidae и ранние Hyaenodontidae (Proviverrinae, Limnocyoninae) изначально были стопоходящими и обладали достаточно широко расставленными плюсневыми костями и пальцами. У более поздних форм (Pterodontinae, Hyaenodontinae) параллельно с некоторым удлинением конечностей пальцы собрираются «в комок», как и у большинства наземных хищников, приспособленных к бегу. У креодонтов присутствует уклон в непарнопалость — третий палец длиннее других. Последние фаланги несколько сжаты с боков (преимущественно у Hyaenodontidae) или расширены и расщепленные на конце сагиттальной щелью (чаще у Oxyaenidae), когти без резкого заострения на конце. У Oxyaenidae когтевые фаланги были плоскими, а когти практически копытцеобразными. У Hyaenodontidae когти являлись маленькими и узкими, словно собачьи, только еще мельче. В целом когти креодонтов были невтяжными, тупыми и не годились для захвата жертвы.
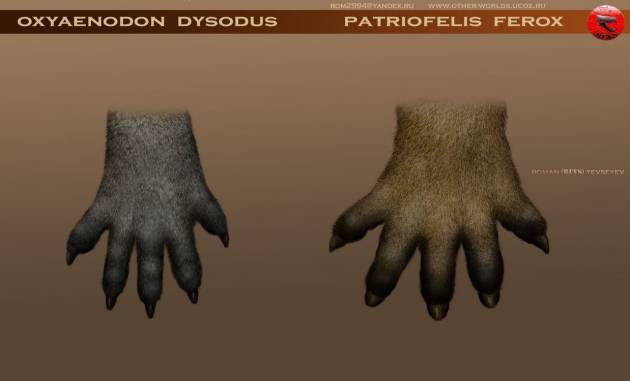
Слева — правая передняя лапа Oxyaenodon dysodus (Hyaenodontidae, Limnocyoninae), справа — правая передняя лапа Patriofelis ferox (Oxyaenidae, Oxyaeninae).
Череп креодонтов большой относительно их тела, лицевой отдел как правило вытянут и почти равен по длине мозговому. Со временем черепа Oxyaenidae приобретают ярко выраженный хищный облик — сильно укорачивается морда и раздвигаются в стороны скуловые дуги, у Hyeanodontinae, напротив, несмотря на резко хищный зубной аппарат, до конца сохраняют вытянутый узкий череп с относительно длинным и низким профилем. Особенно большими, почти непропорциональными головами обладали крупные представители группы, для которых длина головы соотносится с общей длиной тела в пропорции примерно 1 : 4. Для креодонтов, как и для всех плотоядных млекопитающих, была характерна частичная редукция и специфическая дифференциация мышц предглазничного комплекса лицевой мускулатуры, очевидно, связанная со специализацией к захвату и удержанию крупной жертвы.
Примечательно расположение глазниц некоторых высокоспециализированных Hyaenodontidae, в частности представителей рода Hyaenodon. Как и у копытных, у них глаза находятся на боковых поверхностях черепа, что обеспечивает чрезвычайно широкий угол обзора. Очевидно, такая особенность была связана со сверххищным образом жизни этих креодонтов и как-то помагала им на охоте.
Морфология слуховой области наделена существенным сходством с Carnivora. Барабанная кость кольцевидная, костного слухового канала нет. Строение слухового пузыря архаично, частично он был построен из каменистой и чешуйчатой костей, однако при этом большая передне-латеральная часть оставалась хрящевой. Пока не известно, был ли второй центр окостенения слуховой капсулы, подобно Feliformia. У Creodonta имеется алисфеноидный канал, что позволяет сближать их с Caniformia. Заглазничное сужение значительное; верхние заглазничные отростки имеются почти всегда, нижние — отсутствуют; позади глазницы нет костной дуги.
Благодаря особенностям строения, челюстной сустав креодонтов позволял очень широко открывать пасть, но допускал только вертикальные движения нижней челюсти. Именно в связи с этим обстоятельством черепа большинства креодонтов обладают сильно развитым сагиттальным гребнем и относительно слабыми скуловыми дугами. Нижняя челюсть массивна, ее венечный отросток высокий. Параллельно плотоядной специализации щечных зубов в развитии челюстного аппарата креодонтов существовала тенденция к изгибу профиля щечного зубного ряда в дорзо-вентральном направлении. Это обусловлено механическим выигрышем в разрезании пищевого объекта при поочередной окклюзии пар режущих зубов от P4/ml к М2/mЗ.
Небольшая толстостенная мозговая полость содержала соответствующий ее размерам примитивный головной мозг. Длительно время он считался сравнительно маленьким, но позже было убедительно продемонстрировано, что относительные размеры головного мозга креодонтов увеличивались так же быстро, как и у ранних хищных, параллельно расширению его новой коры (неокортекса). От самых первых креодонтов до более поздних представителей группы довольно четко прослеживается постепенное усложнение структур головного мозга. Однако в целом у креодонтов сохранялись почти гладкие и относительно крупные полушария большого мозга, не покрывающие мозжечка и крупных обонятельных луковиц, у многих были хорошо развиты височная и грушевидная доля, отвечающая за обоняние. Однако почти не вызывает сомнений, что даже при относительно гладком мозге у этих животных были неплохо развиты ассоциативные зоны, обеспечивавшие достаточно высокий уровень интеллекта, необходимый при их ярко выраженном хищном образе жизни. Мозжечек отделялся от остального мозга тенториумом.
Исследования эндокранов позволили установить, что некоторые креодонты характеризовались хорошо развитыми органами чувств (особенно зрением и обонянием, что проявляется в существенном увеличении соответствующих долей мозга и отделов новой коры) и обладали совершенной моторикой движений. Топологическая организация борозд и извилин неокортекса креодонтов относится к генерализованному продольному типу — они ориентированы параллельно сагиттальной оси (архаичная черта, присущая также ранним псовым). Специализация неокортекса сопровождалась морфологическими изменениями, характерными для всего отряда с сохранением продольной ориентации борозд и извилин. Отличия исследованных таксонов в специализации неокортеса кореллируют с различиями в строении челюстного аппарата. Гиперспециализация нервной системы, а не челюстного аппарата, сделала часть креодонтов уязвимой для изменений условий обитания и добывания пищи. Формирование адаптивных зон, связанное с глубокими изменениями в организации поведения и строении неокортекса креодонтов, привело к сужению возможностей представителей отряда, что, видимо, и явилось одной из причин его вымирания.
Зубная система Creodonta очень своеобразна и по некоторым признакам конвергентно сходна с таковой Miacoidea, а также Borhyaenidae и Dasyuridae. Для нее присуща сильно выраженная мелкозубость (особенно по сравнению с современными хищниками). Зубная эмаль обладает специфической косой слоистостью, что делало ее более устойчивой. Судя по строению зубов, Creodonta (в отличие от Didymoconida, Arctocyonia и Mesonychia) являлись высокоспециализированными хищниками.
Любопытно, что только у них наблюдается сокращение числа резцов. У ранних представителей отряда еще присутствует их полное число, они небольшие, мало различающиеся по величине и расположены в поперечном ряду. У высокоспециализированных форм, с ярко выраженным плотоядным типом коренных зубов, резцы претерпевают существенные изменения и достигают гораздо большей дифференциации, нежели у Carnivora. Из нижних i1 и i 3 уменьшаются в размерах, становятся впереди или позади i2, иногда исчезают. Из верхних редуцируются или исчезают I1 (у Pterodon) и I2, а I3 иногда сильно увеличивается и приобретает функцию дополнительного клыка (у Megistotherium), что свидетельствует о значительной хищнической специализации. У Carnivora все иначе: резцов всегда по три пары сверху и снизу (единственное исключение — медведь губач (Melursus ursinus), питающийся преимущественно муравьями). Клыки креодонтов большие, заостренные, с одним корнем, зачастую массивные. Именно увеличенные клыки занимают освободившееся от резцов пространство. Получается, что эволюционный процесс креодонтов был направлен на сьлижение расстояния между клыками. Зачем — до конца непонятно. Возможно, это увеличивало силу укуса: сокращение челюстной мускулатуры более эффективно вгоняло клыки в плоть жертвы.
Предкоренные бугорчатые и относительно просто устроенные, задние верхние иногда с низким протоконом. Коренные зубы креодонтов изначально были построены по примитивному трибосфеническому типу, но в процессе эволюции они в большинстве случаев претерпели существенные изменения. Нижние коренные обладают высокими тригонидами и узкими талонидами; тригониды зачастую шире, нежели талониды; талониды нередко уменьшены, особенно на задних коренных. Нижний хищнический зуб обычно имеет уменьшенный метаконид и секториальный паракристид. Верхние коренные треугольной формы, с соединяющимися параконом и метаконом, нобольшим конулем и хорошо заметным, но зачастую уменьшенным протоконом. Метастиль верхних хищнических зубов большой и соединенный с метаконом. Последний верхний коренной обычно уменьшен или полностью отсутствует, размеры последнего нижнего коренного варьируют от очень крупных до совсем небольших, вплоть до полной его редукции.
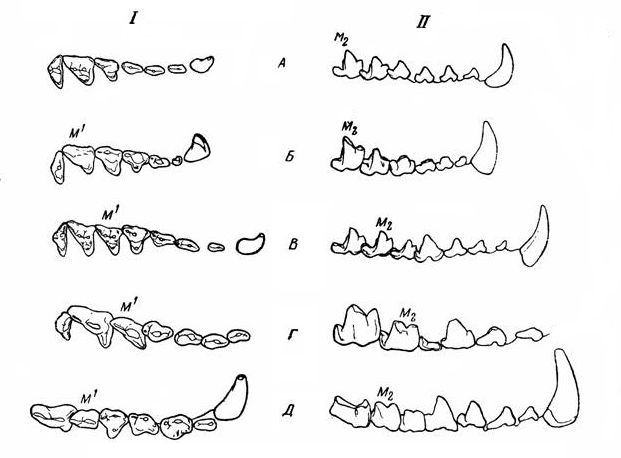
Различные типы строения щечных зубов Creodonta (схема): I верхние, II— нижние; А — Thinocyon (Hyaenodontidae), Б — Oxyaea (Oxyaenidae), В — Sinopa (Hyaenodontidae), Г — Pterodon (Hyaenodontidae), Д — Hyaenodon (Hyaenodontidae).
Для более эффективного разделывания животного корма щечной зубной ряд креодонтов эволюционировал в превосходный режущий аппарат, по действию напоминающий ножницы. Путем секториальной специализации некоторые зубы трансформировались в хищнические, обладающие острыми режущими поверхностями. Преобразованию примитивного бугорчатого зуба в режущий хищнический подверглись разные щечные зубы: у Oxyaenidae сильнее всего — М1 и m2, хотя на ранних стадиях их эволюции этим процессом частично захвачены также Р4 и m1. m1 иногда даже больше, чем m2 (подсемейство Palaeonictinae); эти зубы в дальнейшем снова упрощаются; М3, а позже и М2, так же как и m3, исчезают. У Hyaenodontidae трансформируются М1-2 и m2-3; все эти зубы на высших стадиях имеют режущий характер, но сильнее всего он выражен большей частью на М2 и m3, которые у поздних форм к тому же делаются более крупными, чем предшествующие; М3 уменьшается и исчезает. В обоих случаях наиболее специализирована задняя пара коренных зубов. Специализация хищнических зубов привела у Creodonta к редукции давящих и перетирающих поверхностей, сохранившихся у Carnivora. Кроме того, у Carnivora хищническими являются Р4 и m1, что кажется эволюционно более выгодным. Близость хищнических зубов Creodonta к заднему концу челюстей препятствовала такому значительному увеличению их в длину, как у наиболее хищных представителей Carnivora — кошек и гиен. Однако наиболее важным для дальнейшей эволюции креодонтов оказалось то, что их далеко расположенные хищнические зубы сделали затруднительным потребление какой-либо пищи, помимо мясной. Таким образом, в силу наличных морфологических предпосылок креодонты могли специализироваться исключительно в указанном направлении, в то время как зубной аппарат карнивор изначально является более многофункциональным и в ряде случаев позволяет разнообразить рацион растительным кормом.
Образ жизни
Креодонты занимали экологические ниши наземных хищников всех размерных классов и играли ключевую роль в биоценозах как консументы второго порядка, некоторые из них могли быть падальщиками. Многочисленные представители отряда предвосхитили большинство экологических групп настоящих хищных (Carnivora). Среди них были звери, аналогичные крупным и мелким куньим, псовым, виверрам, мангустам, енотам, медведям и каланам. Однако многие креодонты настолько своеобразны, что не похожи ни на одного другого хищника (например, гигантский Sarcastodon с очень широким и коротким лицевым отделом).
В большинстве своем группа включала наземных животных, хотя некоторые её ранние представители неплохо лазали по деревьям или даже вели вполне древесный образ жизни (например, куницеобразный Lesmesodon). Быстро бегать, да еще и на приличные расстояния, ранние креодонты (Oxyaenidae, Proviverrinae, Limnocyoninae) не могли. По сути, они передвигались не быстрее человека и только на короткие дистанции. Охотилось большинство ранних креодонтов преимущественно из засады, подкарауливая зазевавшуюся добычу. Однако со временем ряд креодонтов (Hyaenodontinae) становится активными бегающими хищниками открытых пространств. Значительную скорость эти животные развивать не могли, но, вероятно, были очень активными и выносливыми охотниками. Кроме того, из числа креодонтов выдвинулись и саблезубые формы (Machaeroidinae), специализированные к охоте на какую-то вполне определенную добычу. Примечательно также, что по меньшей мере один представитель отряда (склерофаг Teratodon) освоил водную среду обитания.
Причины исчезновения
Видимо, основными причинами исчезновения креодонтов являлись консервативность их морфологии, ее приверженность к архаичным чертам, которые представители отряда пронесли на протяжении всей истории его существования, и их крайняя специализация как хищников, жестко привязанных к определенным условиям и добыче. Все это способствовало сужению адаптивных возможностей в постоянно меняющемся мире, а значит, ставило под вопрос само существование таксона.
В частности, одной из возможных причин вымирания мог стать проглядывающийся эволюционный тупик в развитии архаичного мозга креодонтов. У них, в отличие от эоценовых хищных (Carnivora), имелся рисунок кольцевых структур из борозд и извилин, спереди от структуры, называемой "медвежьим" ромбом. Такое расположение могло блокировать дальнейшее усложнение коры мозга. Возможно, именно это завело креодонтов в тупик и не дало на равных соперничать с настоящими хищными.
Зачастую неверно утверждается, что креодонты были вытеснены более адаптивными карниворами. Однако согласно данным палеонтологической летописи, почти все группы настоящих хищных (за исключением мелких, древесных и некоторых узко специализированных форм) получают широкое распространение только некоторое время спустя после исчезновения креодонтов. Очевидно, различные креодонты раньше настоящих хищных заняли большинство ниш наземных хищников и вплоть до своего вымирания просто не допускали в них потенциальных конкурентов.





