


Хищники
Предлагаю создать сообщество "Хищники". Эта увлекательная тема нравится многим читателям. Есть очень интересные статьи, которые расширят кругозор заинтересованных лиц. Со своей стороны я буду регулярно выкладывать авторские публикации и отвечать на комментарии читателей. Это разнообразит контент на pikabu и привлечет новых читателей, что немаловажно для любого сайта. А в нынешних условиях, когда из-за борьбы с эротикой, многие подписчики покинули pikabu, это сообщество поможет приобрести новых.



Тилозавры.

Тилозавры (лат. Tylosaurus от др.-греч. τύλος «деревянный гвоздь, шип» и σαῦρος «ящерица») — род гигантских мозазавров мелового периода. Одни из самых известных и крупных мозазавров. Род описан Г. Ч. Маршем в 1872 году; название выбрано, вероятно, из-за вытянутого рыла. Интересно, что типовой вид был описан в 1869 году Э. Д. Коупом, соперником Марша, из отложений Канзасского Мелового моря. Но Коуп отнес животное к роду Macrosaurus, а это название было уже «занято». Позднее он же отнес типовой образец к малоизвестному европейскому роду Liodon. Остатки тилозавров широко распространены в отложениях позднего мела США, поэтому их описывали под самыми разными названиями (Rhamphosaurus, Rhinosaurus).
История находки
Ранняя история этого мозазавра запутана и связана с соперничеством между двумя американскими палеонтологами, Эдвардом Дринкером Коупом и Отниэлем Чарльзом Маршем. Первоначально название «Macrosaurus» proriger было предложено Коупом для фрагментарного черепа и тринадцати позвонков, найденных у близлежащих скал в Западном Канзасе в 1868 году. Образец был размещен в коллекции Гарвардского музея сравнительной зоологии. Год спустя, Коуп более подробно описал тот же материал и причислил его к английскому роду мозазавра Liodon. Затем, в 1872 году, Отниел Марш называет более полный экземпляр под новым родом Rhinosaurus («носатая ящерица»), но вскоре обнаружил, что это имя уже использовалось для описания другого животного. Коуп предложил заменить Rhinoaurus еще одним новым именем, Rhamposaurus, которое также оказалось дублирующим. Наконец позднее, в том же 1872 году, Марш установил название Tylosaurus, включив в него оригинальный материал из Гарварда, а также дополнительные, более полные экземпляры, которые также были доставлены из Канзаса.[1] В 1899 году Генри Осборн описал полный скелет мозазавра Tylosaurus dyspelor (образец AMNH FR221).
То что является поистине уникальным, это то, что профессор Самуэль Уиллистон, в 1898 году, впервые опубликовал подробные фотографии этого полного экземпляра мозазавра, включаящих фотографии 1,2-метрового черепа и отдельных частей мозазавра, в качестве фигур в своем отчете для геологической службы Университета. Образец добрался до Канзасского музея в серии из больших плит, извлеченных под руководством г-на Германа и Томпсона. За одиночными исключениями, все контуры оригинальных плит были сохранены и установлены вместе, в исходных позициях, поэтому животное находится в таком же положении, как оно было найдено в скалах. Оригинальная матрица окружает практически все кости, и ее можно отличить от окрашенной по блеску штукатурки даже на фотографиях, по её несколько более темному оттенку. Окаменелости были установлены на панели длиной 25 футов (7,5 м) и размещены в коридоре, посвященному морским рептилиям. Как отметил Уиллистон, кончик хвоста экземпляра обрывается и не имеет плавного завершения, вероятно, полная длина этой особи составляла около 8,8 метра.
Летом 1918 года Чарльз Х. Штернберг и его сыновья (Джордж Ф. и Леви) обнаружил частично переваренные кости плезиозавра в содержимом желудка у взрослого (9 м) скелета Tylosaurus proriger в отложениях Смоки-Хилла из образования Ниобрары вблизи Дона Битт-Крик в графстве Логан, штат Канзас. Штернберг сообщил о своем открытии на ежегодном собрании Канзасской академии наук в 1919 году и указал, что материал был отправлен в Национальный музей Соединенных Штатов. Однако из-за необычных обстоятельств, связанных с публикацией его краткой статьи в 1922 году, ассоциация двух экземпляров оставалась в основном незамеченной до 2001 года. Этот образец был вновь найден и описан Майклом Эверхартом в 2004 году. Эта история легла в основу сюжетной линии в фильме National Geographic «Морские монстры» и книги под тем же названием (Sea Monsters, 2007). Эта необычная ассоциация убедительно доказывает, что тилозавр питался плезиозаврами и предоставляет дополнительные данные о экология западного внутреннего моря. Этот образец дает первое доказательство того, что более крупные особи Тилозавра, по крайней мере, иногда питаются морскими рептилиями, кроме черепах и мелких мозазавров. То, что 9-ти метровый мозазавр с длиной черепа в 1,2 м смог проглотить большую часть 3-х метрового плезиозавра , дает дополнительные данные о хищнических привычках и возможностях кормления этих крупных морских животных. Реконструированный образец тилозавра (USNM 8898) в настоящее время установлен в Национальном музее Смитсоновского института, а остатки плезиозавра хранятся в хранилище коллекции. Фотография черепа тилозавра была сделана Джорджем Ф. Штернбергом примерно в 1926 году после того, как он нашел и подготовил образец, который был обнаружен у Смоки-Хилл округа Логан, штат Канзас.[3] Штернберг предложил образец Смитсоновскому институту и включил фотографию в своем письме Чарльзу Гилмору. Копии оригинальных фотографий находятся в архивах Музея естественной истории Штернберга (FHSM).
В 1974 году был обнаружен экземпляр по прозвищу «Брюс». Он был найден к северу от Торнхилла, в долине Пембины канадского штата Манитоба. Ископаемые были обнаружены в 80-млн летних морских отложениях формации Пьерр Шэйл, которое образовано Западным внутренним морем, разделяющим Северную Америку на две части. Для раскопок скелета потребовалось около двух полевых сезонов, скелет был достаточно полным с 65-70% исходных костей. Размер оценивается приблизительно в 13 метров (43 фута) от рыла до кончика хвоста.[4] В 1988 году канадский исследователь Бэтси Николлс описала этого мозазавра под видом Hainosaurus pembinensis (хайнозавр), однако последующее изучение позволило установить, что этот экземпляр принадлежит роду Tylosaurus (J. Lindgren 2005, Timon Bullard & Michael Caldwell 2010). Экземпляр демонстрируется в Канадском центре обнаружения окаменелостей (CFDC) в Модерне, штат Манитоба. 15 марта 2015 года сотрудники CFDC с гордостью представили реконструкцию нового образца из коллекции морских окаменелостей рептилий. «Сьюзи» - скелет мозазавра длиной более 9 метров, присоединится к «Брюсу» и будет экспонироваться на выставке, которая раньше называлось «Галерея Брюса», а теперь будет называться Мозазавр-Холл.
На протяжении многих лет были названо множество видов тилозавра[5], однако многие из них, основаны на несовершеннолетних экземплярах и только некоторые из них теперь признаны учеными. Исследования опубликованные в 2016 году, указывают на то, что европейский хайнозавр (Hainosaurus bernardi), вероятно может принадлежать роду Tylosaurus.
Описание
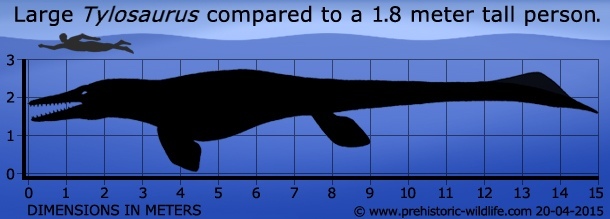
Наряду с плезиозаврами, акулами, и другими мозазаврами, тилозавр был доминирующим хищником западного внутреннего моря, во время позднего мелового периода. Этот род был одним из самых крупных мозазавров (наряду с Мозазавром Гоффмана), например вид T. pembinensis достигал сопоставимых размеров с Hainosaurus bernardi, длиной около 12,2 метров (40 футов), а T. proriger, самый крупный вид тилозавров, достигал длины более 14 метров (46 футов). Содержание желудка, связанное с образцами Tylosaurus proriger, указывает на то, что этот свирепый мозазавр имел разнообразную диету, включая рыбу, акул, более мелких мозазавров (клидаст и платекарп), плезиозавров и нелетающих птиц, таких как гесперорнис (Hesperornis). Так, например, в желудке сравнительно некрупного вида Tylosaurus nepaeolicus были найдены зубы огромного гибодонта Ptychodus mortoni и останки платекарпа Platecarpus planifrons. Относительно недавно в области желудка 8,8 м тилозавра обнаружили скелет небольшого короткошеего плезиозавра долихоринхопса длиной около 2,5 м и позвонок крупного ксифактина, первые упоминания о таких находках относятся к 1918 году.
Череп легкий и кинетический, позволяющий заглатывать крупную добычу. Зубы уплощены изнутри, наружная их поверхность закруглена, эмаль на внутренней поверхности исчерчена. Кончик ростра лишен зубов, именно эта особенность и дала название ящеру. По одной из версий, ростр являлся центром чувствительных нервных окончаний, позволявших охотиться на больших глубинах в темноте. Известны находки черепов со сломанным при жизни ростром (вместо него развилась костная грибовидная мозоль). Скелет был довольно легкий и имел 29—30 предкрестцовых позвонков, 6-7 крестцовых, 33-34 хвостовых с «шевронами», 56-78 терминальных хвостовых. В толще рёбер обнаружены участки аваскулярных некрозов (признак перенесённых при жизни глубоких погружений, при том, что тилозавр не был к ним приспособлен) и остеопороза. Ласты относительно узкие. Отпечатки кожи показывают наличие ромбических мелких чешуек с килями и гребнями, причем чешуи у 8-9 метрового тилозавра были мельче, чем у средних размеров гремучей змеи (у тилозавра 90 чешуек на квадратный дюйм, а у змеи — всего 80).

Отпечатки кожи
Публикация «О кожном покрытии мозазавровых рептилий» была издана профессором Канзасского университета Фрацисом Сноу в 1878 году, где он описал сохранившиеся отпечатки кожи Tylosaurus proriger у образца KUVP-1075, который в настоящее время находится в коллекции Музея естественной истории Университета Канзаса. Однако в публикации профессора Сноу он упоминается под видом Liodon.
Научная экспедиция Канзасского университета 1878 года, под руководством профессора Бенджамина Франклина Маджа и Франциса Хантингтона Сноу, исследовала геологический участок в графстве Гоув, штат Канзас. Эти породы относятся к группе позднемелового образования Ниобрара. Местность ранее уже посещали несколько геологов-исследователей и вряд ли ожидалось, что находки будут содержать что-то необычное, однако менее чем за три недели было обнаружено 41 ящерица, 117 рыб и кости 6-ти птеродактилей. Наиболее ценным образцом стал ящер, найденный в желтом известняке. Его первая кость была обнаружена 11 июля, на второй день, при отчистки других костей от породы, на срезе была обнаружена своеобразная сетчатая поверхность, имеющая площадь в несколько квадратных дюймов- это было нечто, что раньше никогда не представлялось человеческому глазу. Профессор Сноу сразу же отметил, что сетчатая поверхность была ископаемым отпечатком кожи ящера, настолько прекрасно сохранившаяся, что каждая чешуйка была отчетливо видна, с четко обозначенным контуром и выделявшийся центральной линией или «килем». Согласно записям, уникальная текстура была настолько прекрасно сохранена, как если бы это животное умерло только вчера.
Отпечаток кожи тилозавра (KUVP-1075).

Профессор Марш из Йельского университета, несколько лет назад уже описывал большие кожные пластинки или щитки, принадлежащие мозазаврам, которые, как он считал «по-видимому, в основном ограничены шеей и горлом». Предполагалось, что кожное покрытие мозазавров будет состоять из более крупных пластин, таких как у аллигатора или крокодила. Однако как выяснилось, оно состоит из небольших чешуек, очень напоминающих по размеру, форме и расположению, покровы современных чешуйчатых, в частности гремучих змей. Все отпечатки были сохранены на одном блоке известняка, который также содержит несколько позвонков, фаланг и другие кости. Сравнение с текстурами живых гремучих змей (Crotalus confluentus) указывает на то, что обнаруженные чешуйки были несколько меньше, чем у змеи, которая содержит в среднем восемьдесят сегментов на квадратный дюйм, а не девяносто.
Череп мозазавра и семья Штернберга (1920-1929 годы)

Образец тилозавра, содержавший короткошеего плезиозавра в области желудка.

Т. proriger Чарльз Р. Найт, 1899.

Окаменелость тилозавра, найденная на острове Джеймса Росса (Антарктида)

Крупный тилозавр по прозвищу «Брюс», ранее классифицируемый как гайнозавр.

Диорама Г. Осборна с реконструкцией скелета Tylosaurus proriger, 1899 г.

Показать полностью
10
Не успел убрать палец
Кормил кайманову черепашку. Мне пипец?

Апдейт, фото боевого черепана

Показать полностью
2

Монголоникс
Начало позднего эоцена Азии. Mongolonyx robustus убил гиракодонта форстеркуперию (Forstercooperia).

Поздняя волна развития мезонихиевых (Mesonychidae) произвела в мир по-настоящему грозных чудовищ, одними из которых были монголониксы (Mongolonyx), обитавшие в позднем эоцене Азии. На сегодня известно два вида: монголоникс массивный (Mongolonyx robustus) и более поздний монголоникс длинночелюстной (Mongolonyx dolichognathus). Возможными предтечами для монголониксов выступают среднеэоценовые мезониксы (Mesonyx), являвшие собой среднеразмерных лёгких стремительных хищников. В дайльнейшем, с распространением редколесий, саванн и степей, потомки мезониксов укрупнились, и на открытые пространства вышли на охоту плотоядные громадины, сопоставимые в размерах с крупнейшими львами.
Реконструкция черепа и головы Mongolonyx dolichognathus.

Родовое наименование Mongolonyx переводится как «монгольский коготь».
Род четко диагностируется и обладает целым рядом характерных морфологических признаков. Так, от Mesonyx его отличают более значительные общие размеры, моляризованные Р4/4, а также относительно более тупые и большие коренные зубы. Его зубы также относительно крупнее и массивнее, нежели у Dissacus, Harpagolestes и большинства других представителей семейства. В отличие от Dissacus и Pachyaena, у Mongolonyx отсутствует цингулюм на верхних зубах и не отмечаются какие-либо следы метаконида на тригонидах нижних коренных. От Synoplotherium, Mongolestes и Harpagolestes данный род отличает более прямая нижняя челюсть с удлиненным и сравнительно узким симфизом.
Двое Mongolonyx robustus схватили бронтотериида протитана (Protitan). Вдалеке в страхе разбегаются пантодонты эудиноцерасы (Eudinoceras).

Важное отличие от большинства мезонихид (вероятно, за исключением Pachyaena) состоит в том, что нижние клыки Mongolonyx располагаются очень близко друг к другу. В отличие от Harpagolestes, коронки нижних предкоренных загнуты назад не столь выражено, а их эмаль-дентиновая линия не является дугообразной. Кроме того, на р4 крупные элементы параконида параллельны протокониду, в то время как у Harpagolestes и Pachyaena параконид образует угол с протоконидом. Расположенный антеропостериорно параконид m3 также четко отличает Mongolonyx от Harpagolestes. От Mongolestes представители рода отличаются наличием m3 и редукцией р1.

Монголониксы представляли собой крупных плотоядных животных. Длина их черепа превышала 50 см, что вполне сопоставимо с черепом аляскинского гризли. Исходя из пропорций Mesonyx, известного по более-менее полному скелету, можно предположить, что эти мезонихиды достигали примерно 2,3 м в длину (не учитывая хвоста) и более 1 м в холке. Однако гораздо больше вопросов вызывает не внешний облик, а предположительный образ жизни монголониксов.
Mongolonyx robustus, более ранний вид, имел череп 52 см в кондилобазальной длине (от затылочных мыщелков до переднего края межчелюстной кости). Верхняя часть черепа до глазниц не сохранилась, но с восстановленным затылком и резцами его полная длина вполне могла составлять до 60 см. Если монголоникс имел пропорции, подобные своему лесному предку мезониксу, его высота в холке могла быть примерно 1,3 – 1,4 метра. А вес взрослого хищника мог достигать 300 - 400 кг. В общем, это был громадный хищник с мощными челюстями. В развитии зубочелюстного аппарата монголоникс хоть и уступал настоящим сверхспециалистам гарпаголестам (Harpagolestes) и монголестам (Mongolestes), но силы его челюстей вполне хватало, чтоб съедать добычу полностью, разгрызая даже крупные кости. В составе одной с Mongolonyx robustus фауны начала позднего эоцена обитали бронтотериевые (например, Protitan), гиракодонты (например, Forstercooperia) тапироиды семейства лофиалетовых, примитивные свинообразные и пантодонты Eudinoceras. Эти растительноядные служили пищей для монголониксов. Примерно в одно время с ними в Азии обитало довольно немалое количество не менее (а то и более) громадных плотоядов – заболоченных мест придерживались эндрюсархи (Andrewsarchus), хонанодоны (Honanodon macrodontus) и паратриисодоны (Paratriisodon), в лесах разбойничали саркастодоны (Sarkastodon). Такого плотоядного мегазверинца, наверное, не собиралось нигде на одном участке суши вплоть до плейстоценовой Северной Америки.
К концу позднего эоцена монголониксы заметно измельчали - Mongolonyx dolichognathus уже на 15 – 20% уступал в размерах своему предшественнику. Наиболее целая находка – нижняя челюсть достигает примерно 35 см, тогда как длина нижней челюсти M. robustus составляла 49 см.
Судя по имеющемуся материалу, коронки их мощных щечных зубов не подвержены значительному износу, нижние предкоренные мало загнуты назад, эмаль-дентиновая линия не дугообразная (загнутые коронки премоляров млекопитающих хищников препятствуют выскальзыванию раскусываемых костей, а дугообразность эмаль-дентиновой линии указывает на повышенную прочность эмали их зубов). На первый взгляд, подобная зубная система кажется не слишком хорошо приспособленой к разгрызанию костей и сухожилий. Это довольно неожиданно для поздних мезонихид, в большинстве своем демонстрирующих выраженную костедробящую специализацию. Само собой разумеется, Mongolonyx существенно превосходит в этом отношении ранних представителей группы типа Mesonyx, однако на порядок уступает таким «специалистам», как Harpagolestes и Mongolestes.
В то же время, тупобугорчатые щечные зубы монголониксов все-таки достаточно массивны (особенно у M. dolichognathus). Установлено, что их корни сливались в своей верхней части, образуя под коронкой дентиновую платформу, что могло способствовать продлению функционирования зуба после стирания коронки. Весьма вероятно, что подобное строение зубов было направлено на сопротивление их изнашиванию и может свидетельствовать о питании очень твердой пищей, в частности о дроблении костей трупов. У других мезонихид, питавшихся более мягким кормом, такое приспособление отсутствовало — их корни не сливались, а разделялись непосредственно под коронкой зуба.
С другой стороны, близко расположенные нижние клыки монголониксов свидетельствуют в пользу адаптации к мощному укусу-отщипыванию, что наблюдается также у некоторых креодонтов (Sarkastodon, Megistotherium). Такие сближенные клыки существенно увеличивали силу укуса, по принципу аккумулирования давления на меньшей площади. Можно предположить, что эти хищники вкладывали всю свою мощь в один укус, вцепляясь в жертву намертво, а не наносили ей множественные раны.
Следующая примечательная особенность морфологии монголониксов — округлая головка их суставного отростка. При этом, судя по сохранившимся образцам, левая и правая мандибулярные ветви полностью срастались в симфизной области. В результате нижняя челюсть получала возможность достаточно свободно двигаться из стороны в сторону, избегая при этом вывиха в симфизе (интересно, что подобная способность развилась также у других выраженных мясоедов — креодонтов рода Hyaenodon). Не менее интересно наличие на сильно вытянутом небе специальных костных трубок, служивших для защиты дыхательных каналов. Они позволяли хищникам не задохнуться, когда те, на охоте или в процессе питания, захватывали в пасть большие куски плоти. По сходным причинам подобные образования вполне независимо развились и у ряда других хищников (к примеру, все у тех же гиенодонов).
Учитывая вышесказанное и принимая во внимание неповоротливость, вполне вероятную для столь массивных животных, можно предположить, что монголониксы являлись преимущественно охотниками на крупную добычу. Очевидно, их жертвами становились в основном громоздкие травоядные, например бронтотерии (особенно молодые), вполне обычные в то время. Очевидно, монголониксы были способны не только справиться с большой жертвой, но и полноценно ее утилизировать. При этом также более чем вероятно, что монголониксы при случае поедали найденные трупы или отнимали добычу у более слабых хищников (или оспаривали у равных по силе — примерно тогда же существовали крупные азиатские гиенодоны).

Показать полностью
5



