


Черепа птерозавров
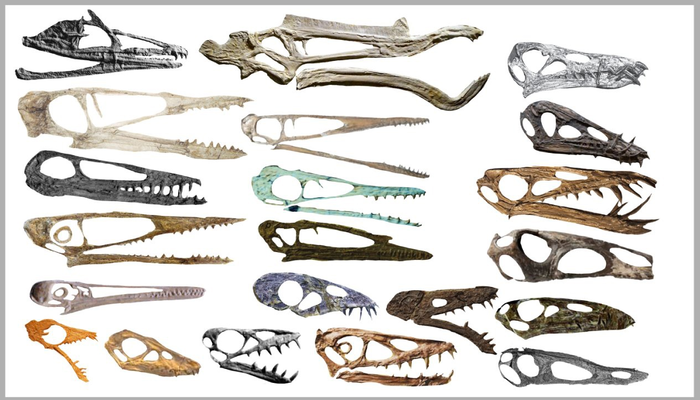

Показать полностью
1
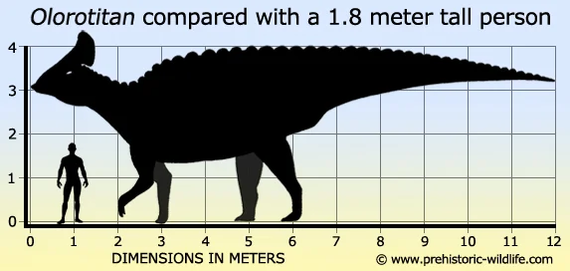
Олоротитан (лат. Olorotitan) — утконосый динозавр из России, обитал в меловом периоде 72 - 66 млн лет назад. Окаменелости этого динозавра были найдены в Кундурском костеносном месторождении, которое было обнаружено в 1990 году геологом Владимиром Нагорным в окрестностях поселка Кундур, Архаринского района Амурской области, при строительстве отрезка дороги Чита-Хабаровск федеральной трассы Москва-Владивосток. Тогда строители, работавшие на месте, замечали попадавшиеся кости, но были уверены, что это останки медведей. Масштабные научные исследования начались с 1998 года под руководством кандидата геолого-минералогических наук Юрия Болотского. В раскопках уже тогда участвовал сын ученого, Иван, который положил начало главному открытию - он обнаружил первые четыре хвостовых позвонка олоротитана. Олоротитан сосуществовал с родственными ему динозаврами харонозавром (лат. Charonosaurus) и базальным гадрозавридом амурозавром (лат. Amurosaurus). На том же участке были обнаружены окаменелости черепах, крокодилов, теропод и нодозаврид. Также на территории Амурской области в породах того же возраста были обнаружены останки керберозавра (лат. Kerberosaurus). В отличие от Северной Америки, где окаменелости ламбеозаврин практически не встречаются в отложениях маастрихтского яруса, в России и азиатских странах их останки датируемые этим времен широко распространены и разнообразны, что указывает на разницу в экологии и климате.
Юрий Болотский на раскопках Olorotitan arharensis, 1999 г

За первое лето на кундурском месторождении были найдены хвост из 75 позвонков и задняя часть туловища ящера. Раскопки в Архаринском районе велись три года. Самой большой интригой для палеонтологов оставался вопрос, отыщется ли голова динозавра. «Он действительно оказался гадом: нет чтобы вдоль трассы лечь, а он завалился перпендикулярно. Если хвост был на поверхности, то все туловище находилось в толще, а над ним четыре метра почвы. К тому же он был обложен изолированными костями, так как улегся, как плотина, на пути грязекаменного потока, и на нем были кости других тварей», — рассказывает Юрий Леонидович.
В 1999 году к раскопкам присоединился палеонтолог Паскаль Годефруа из Бельгийского Королевского института естественных наук, а в 2000 – сотрудники Палеонтологического института РАН из Москвы. Экспедиция увенчалась триумфом: под Кундуром впервые в России был обнаружен целый скелет одной особи, включая череп. Извлечение костей из породы продолжалось три полевых сезона с 1999 по 2001, а в 2003 году эта находка была описана палеонтологами Паскалем Годефруа, Болотским и Алифановым под назанием Olorotitan arharensis - «лебедь-титан».В 2012 году ими было опубликовано остеологическое исследование анатомии скелета и его отдельных костей.
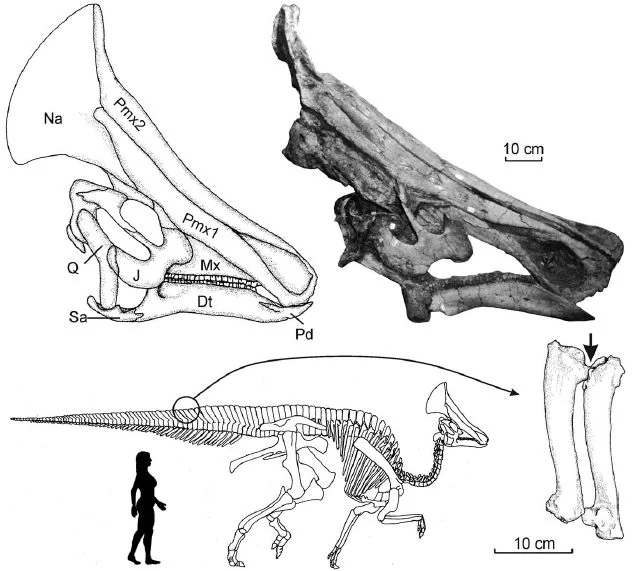
Голотип (AEHM 2/845) состоит из хорошо сохранившегося скелета с неполным черепом. Хотя большинство костей черепа были слегка смещены, а кости носа сохранились не полностью, череп удалось довольно точно реконструировать. Эта находка - первый почти полный скелет динозавра в естественном сочленении, обнаруженный на Дальнем Востоке России. Реконструированный скелет этого динозавра экспонируется в музее Королевского института естественных наук в Брюсселе, Бельгия. Найденный олоротитан при жизни имел несколько ракообразных опухолей и, похоже, хромал на одну ногу, постольку пяточная и таранная кость были сросшимися с большой и малой берцовыми костями. Олоротитан многократно подвергался нападениям теропод. Имеются повреждения позвоночного столба, пояса конечностей, нижней челюсти и дистальных позвонков хвостового отдела.
Олоротитан - это самый полный экземпляр ламбеозаврин, обнаруженный за пределами Северной Америки. Этот динозавр был одним из самых крупных представителей своего семейства, таких как Parasaurolophus, достигая в длину 8 метров (26 футов). Он характеризуется специфическими для гадрозаврид признаками, самый очевидный из которых это уникальной гребень венчающий череп. Широкий, полый гребень, образованный из расширенных костей черепа, был пронизан носовыми путями и вероятно использовался для подачи трубных звуков. Сам череп поддерживается довольно длинной шеей, имеющий 18 позвонков, другие гадрозавриды имеют максимум 15 шейных позвонков. Филологическое исследование олоротитана привело к выводу, что он состоит в очень близком родстве с североамериканскими коритозавром (лат. Corythosaurus) и гипакрозавром (лат. Hypacrosaurus). Также на территории Амурской области в породах того же возраста были обнаружены останки другого гадрозаврида — амурозавра. В 1997 году в Благовещенске был создан палеонтологический музей. Реконструированная копия скелета олоротитана, включающая череп с предполагаемой формой гребня, демонстрируется в Амурском музее естествознания, в Благовещенске.
Олоротитан в Амурском палеонтологическом музее

Филологическое исследование олоротитана привело к выводу, что он состоит в очень близком родстве с североамериканскими коритозавром и гипакрозавром. Также на территории Амурской области в породах того же возраста были обнаружены останки другого гадрозаврида - амурозавра. В 1997 году в Благовещенске был создан палеонтологический музей. Реконструированная копия скелета олоротитана, включающая череп с предполагаемой формой гребня, демонстрируется в Амурском музее естествознания, в Благовещенске.
Олоротитан в Королевском институте естествознания, Бельгия


Кархародонтозавр (лат. Carcharodontosaurus) — гигантский хищный динозавр из Африки, живший в меловом периоде 100-94 млн лет назад на территории современной Африки.
Череп кархародонтозавра в сравнении с американским палеонтологом Полом Серено.

Первые два зуба были обнаружены на территории Алжира в 1924 году. Чарлз Депере и Саворнин в 1925 году отнесли их к роду Megalosaurus saharicus, но в 1927 году переименовали в Dryptosaurus saharicus. В 1914 году, в формации Бахариджа Египта были обнаружены разрозненные кости хищного динозавра, состоящие из обломка черепа, зубов, нескольких позвонков и бедерной кости задней конечности. Эти зубы были идентичны образцам описанным Депере и сильно напоминали зубы акулы, исходя из этого, Эрнст Штромер в 1931 году описал эти останки как Carcharodontosaurus saharicus — «ящер с акульими зубами». К сожалению эти образцы были утрачены во время Второй Мировой войны, в 1944 году.

Новые окаменелости были обнаружены палеонтологом Полом Серено в 1995 году, в формации Кем-Кем, Морокко, недалеко от алжирской границы, образец состоял из неполного черепа (образец SGM-Din 1). В 2007 году Стивен Брюссат и Пол Серено обозначили его как неотип, в том же году они описали новый вид Carcharodontosaurus iguidensis, основываясь на обломке челюсти с зубами из Нигера (образец MNN IGU2).
В 2010 году Грегори Пол предложил переименовать гиганотозавра в C. carolinii. Однако это решение не поддержали.
В 1960-ых годах к кархародонтозавру отнесли огромное количество разнообразных фрагментов меловых теропод, не имея для этого достаточных признаков. Вполне возможно, что в одно время с C. saharicus обитало несколько кархародонтозаврид, поэтому многие исследователи ограничивают образцы данного вида.

Челюсть Carcharodontosaurus iguidensis
Описание
Череп известен плохо, но напоминал таковой у гиганотозавра, зубы были с зазубринами и достигали 20 см в длину. В 1996 году Пол Серено оценил возможную длину черепа в 1,6 метра. Вероятно, кархародонтозавр мог охотится на крупнейших завропод - титанозавров, чьи кости найдены в тех же отложениях. Как и для гиганотозавра, для кархародонтозавра нет никаких доказательств стайного поведения. Кархародонтозавры были доминирующими плотоядными хищниками в Африке, с предполагаемыми размерами тела 10 - 12.8 метров в длину и 7.8 т веса.
Второй вид С. iguidensis был мельче и по оценкам учёных достигал 10 - 11 м и весил 4 - 5.2 т (Paul 2010, Molina-Perez и Larramendi 2019)
Пол Серено определил отличительные черты C. saharicus, включающие в себя приподнятый нижний край выпуклости возле заглазничного отверстия, глубокую направленную вниз канавку между внутренней стенкой верхней челюсти и передним внутренним отростком, высокий направленный вверх отросток между слёзной и лобной костью. Передний внутренний угол впадины возле верхней фенестры создаёт сжатую полость в крышке черепа. Челюстные зубы имеют характерные гребни на эмали.
В 2007 году Серено также определил уникальные половые разичия. Внешняя сторона верхней челюсти рифлёная, а возле места расположения сонной артерии на черепной крышке и вокруг основания затылка присутствовала обильная пневматизация. В нижней части черепной крышки находится отверстие с подверженным эрозии феноидом.
C. saharicus и C. inguidensis различаются строением слёзной кости и верхней челюсти, однако оба вида имеют бороздчатою поверхность зубов. У C. saharicusони больше, чем у C. iguidensis, но у второго мозг имеет на 15% больший объём.

Строение мозга
В 2001 году Ханс К. Ларссон опубликовал описание внутреннего уха и эндокринной системы C. saharicus. Начиная с самого близкого к концу челюсти начинается передний мозг, за которым следует средний мозг, наклонённый к задней части черепа под углом в 45 градусов. За ним следует ромбовидный мозг, параллельный переднему и образующий с средним мозгом сорокаградусный угол. Внешне мозг C. saharicus напоминал мозг аллозавра A. fragilis. Ларссон обнаружил, что отношение головного мозга кархародонтозавра к остальным отделам было типичным для не-авиальных динозавров. У него также был хорошо развит зрительный нерв.
Три полукруглых канала внутреннего уха C. saharicus имели субтриагулярный контур, если смотреть сбоку. Такая конфигурация также есть у аллозавра, ящериц, черепах, но не у птиц. Сами эти каналы имели линейное строение, что объясняет заострённый силуэт. При жизни флокулярная доля мозга располагалась возле полукруглых каналов, как у других карнозавров и птерозавров.


Зуб кархародонтозавра в Музее Естествознания в Вене
Систематика
Первоначально, в 1931 году, Стромер определили кархародонтозавра в собственное семейство кархародонтозавриды, но в 1934 изменил своё решение и переклассифицировал его в качестве аллозаврида. В 1995 году Оливер Раухут вернул кархародонтозавридам статус семейства.В 2009 году кархародонтозавра определили к африканской ветви семейства, однако поздние исследования ставят его ближе к южноамериканским кархародонтозавридам.
Необычным можно также и отметить сходства кархародонтозавра и с гиганотозавром, и с аллозавром. Кроме того, первый обитал вместе с мапузавром, тираннотитаном и другими крупными кархародонтозавридам на территории Южной Америки, а сам кархародонтозавр - в Египте, Марокко, Тунисе, Судане, Нигере. Аллозавра, которого считают предком кархародонтозаврид, обнаружили на территории Северной Америки, поэтому его потомки могли перейти на территорию Аргентины в мелу, но не в Африку. Не исключено, что предки кархародонтозавра являются дальними родственниками южноамериканских видов, перешедшими по сухопутному мосту на другие континенты. Теория кажется правдоподобной, однако её противоречат два факт - найденный ещё в 1925 году "Allosaurus" tendagurensis из Танзании, которого сейчас выделяют в отдельный род, и Allosaurus europaeus из Португалии. Эти виды уже обитали в Африке в юрском периоде, а A. europaeus вообще нашли недалеко от Северной Африки, опровергая собой теорию моста.

Кархародонтозавр от Андрея Аутчина
Палеобиология
В 2011 году палеоиллюстратор Боб Николлс опубликовал знаменитую работу, где два кархародонтозавра несли молодого райозавра. Картина вызвала многочисленные споры и обсуждения о возможноститакой охоты. Исследования Дональда Хендерсона, куратора динозавров Королевского музея Тирелла, показали, что строение челюстей, шеи и центра массы кархародонтозавра позволяли ему поднимать животных массой до 424 кг. То есть, его добычей могли быть некрупные завроподы, вроде молодых египтозавров, а также другие хищники - некрупные мегалозавриды или крокодиломорфы элозухи. Максимальный вес добычи кархародонтозавра составлял около 850 кг, что примерно сопоставимо с длиной в 8,3 метра.
Зубы кархародонтозавра напоминают зубы большой белой акулы, кархародона. Поэтому считается, что этот динозавр совершал быстрые, резкие укусы и вскоре отступал. А жертва умирала от потери крови.
У кархародонтозавра было типичное для теропод хорошее зрение. А его мускулистые ноги позволяли передвигаться со скоростью около 8,5 м/с, хотя эта оценка довольно спорна. С весом в 4 тонны падение на большой скорости было бы для кархародонтозавра смертельным. Строение ног не позволяло бы ему вовремя остановиться.

Патология
Один из образцов кархародонтозавра, SGM-Din 1, имеет колотую рану на переднем краю целюсти и "аномальную проекцию кости на заглазничное кольцо".
Палеоэкология
Останки кархародонтозавра находят почти по всей территории северной Африки: в формации Бахария, Египет, возле посёлка Марса-Мархут, тоже Египет, в формации Тегана и Ксар-эс-Суке, Марокко, в формации Ченини, Тунис, в Гарьяне, Ливия, в формации Кем-Кем, Алжир, и в Агадасе, Нигер. Вид C. iguidensis известен только по останкам из Нигера.
Кархародонтозавр обитал на жарком побережье древней Африки. Местность, где обнаружили окаменелости, была покрыта болотами и мангровыми лесами. Учитывая размеры кархародонтозавра, он был апексхищником, однако его связи с другими местными тероподами неизвестны. Точно так же неизвестна и настоящая добыча кархародонтозавра. Формация Кем-Кем была скудна на растительноядных животных, однако в другом месте обнаружения C. saharicus, формации Бахария, обитали крупные египтозавр и паралититан. Кархародонтозавр мог охотится на них, однако не на самых крупных особей.
Находка второго вида, C. inguidensis , показывает, что все образцы из области Фундортс, ранее причисляемые крупному C. saharicus, принадлежат нигерскому виду. Рядом с ним нашли также окаменелости ругопса и спинозавра. Маловероятно, что три крупных теропода могли обитать на одной территории, поэтому весь материал из этой области отнесли к C. iguidensis. Кроме теропод, негирийский вид соседствовал с крупными крокодиломорфами, вроде саркозуха, капрозуха, лаганозуха и арарипезуха.

Раннеюрские морские плезиозавры Rhomaleosaurus. Их ареал охватывал оба полушария и простирался до полярных морей на территории современной Якутии.

Выполненное российскими палеонтологами тщательное исследование остатков морских рептилий, собранных около полувека назад по берегам якутской реки Вилюй, выявило их большое таксономическое разнообразие. Оказалось, что приполярные моря юрского периода населяли несколько родов ихтиозавров, плиозавров и плезиозавров. Среди находок было много костей молодых рептилий, в том числе, возможно, принадлежащие новорожденным особям. Это позволяет предположить, что морские рептилии юрского периода использовали сибирские моря как ясли и приплывали туда летом для размножения.
Остатки морских рептилий первой половины юрского периода (201–174 млн лет назад) очень неравномерно представлены в разных регионах планеты. Почти все наши знания о них ограничены находками из Великобритании и Германии. Другие регионы дали лишь несколько экземпляров, причем в основном это всего лишь разрозненные и плохо сохранившиеся кости.
Такая ситуация объясняется прежде всего человеческим фактором и географическими особенностями. Карьеры Хольцмадена (Holzmaden) в Германии и знаменитые береговые обрывы британского курорта Лайм-Риджис (Lyme Regis) привлекают внимание геологов, палеонтологов и любителей уже две сотни лет. Они легко доступны, в них ведется постоянный мониторинг, который регулярно приносит новые интересные находки. Таких условий нет в других местах, где вскрываются раннеюрские морские отложения, — в том числе в безлюдной тайге Якутии. Тем не менее даже при несопоставимом числе находок эти регионы могут дать важную информацию для понимания исчезнувших экосистем.
В журнале Geological Magazine недавно вышла статья российских палеонтологов с описанием остатков морских рептилий из Якутии. Ископаемые кости были собраны еще полвека назад в отложениях ранней юры, которые вскрываются по обрывам реки Вилюй и охватывают интервал продолжительностью в 25 миллионов лет (190–175 млн лет назад).

Обнажения раннеюрских пород на обрывистых берегах Вилюя.
Коллекция состоит из 150 костных остатков. Почти все они — отдельные позвонки и кости конечностей. Наиболее целые находки: фрагмент позвоночного столба и остатки челюстей двух ихтиозавров.
Большинство остатков сложно определить даже до семейств, но некоторые получилось идентифицировать до родов. К примеру, на обломке морды крупного ихтиозавра сохранились отпечатки нескольких коронок зубов (сами они разрушились), на одном отпечатке удалось заметить режущую кромку. В юрском периоде такие кромки были только у ихтиозавров Temnodontosaurus. Представители этого рода были главенствующими хищниками раннеюрских морей, достигая длины десяти метров. Судя по фрагментам челюстей, якутский ихтиозавр был примерно семиметровым.
По схожим мелким деталям удалось определить и некоторые другие остатки: небольших ихтиозавров Stenopterygius, длинношеих плезиозавров Microcleidus, а также плезиозавров из семейства ромалеозаврид (Rhomaleosauridae) и примитивных плиозаврид.
Все эти животные давно известны по находкам в раннеюрских отложениях Западной Европы, а также в некоторых других регионах. Например, Temnodontosaurus недавно был обнаружен в пустыне Атакама в Южной Америке. Подобным космополитизмом отличаются многие морские животные, в том числе вымершие, но не все.
В Восточной Якутии не оказалось морских крокодиломорф талаттозухий, широко представленных в Западной Европе. Можно предположить, что им, подобно другим крокодиломорфам, не удалось заселить полярные регионы из-за климатических ограничений.
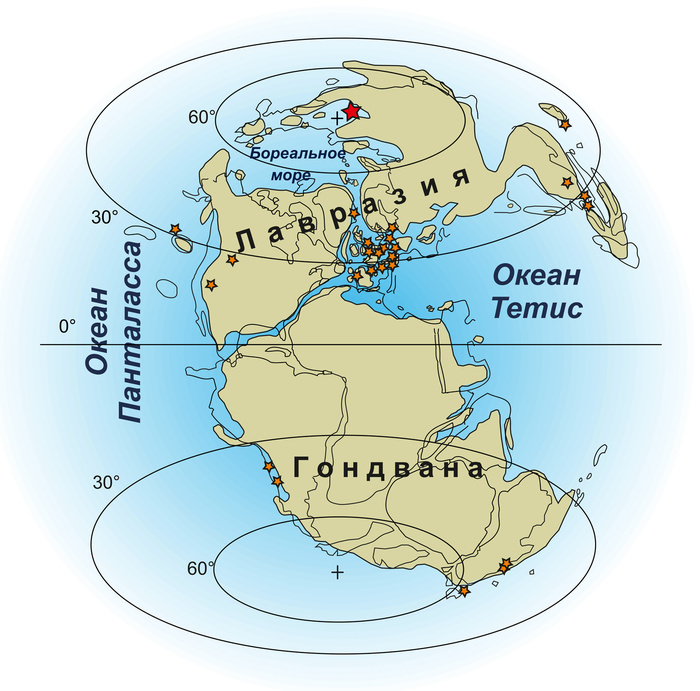
Палеогеографическая карта начала юрского периода. Звездочками отмечены местонахождения остатков морских рептилий. Полюса Земли обозначены крестиками. Красная звездочка возле северного полюса — якутские местонахождения. Климат этого времени для высоких широт пока изучен недостаточно хорошо, но считается, что он был достаточно прохладным (см. новость Мезозойские морские рептилии Сибири жили и в теплых , и в холодных водах, «Элементы», 16.07.2019). Рисунок из обсуждаемой статьи в Geological Magazine
Вилюйские местонахождения сформировались в мелководных морях к северу от восьмидесятой параллели, то есть даже ближе к северному полюсу, чем они расположены сейчас. Это самые высокие широты для находок ранних морских рептилий.
Изобилие ихтиозавров и плезиозавров в прохладных водах подтверждает мнение об их теплокровности (см. новость Мезозойские морские рептилии Сибири жили и в теплых, и в холодных водах, «Элементы», 16.07.2019). Кроме того, для обитания в полярных водах у них могли быть и другие адаптации, например, толстый слой жировой ткани, недавно описанный для раннеюрского ихтиозавра Stenopterygius (J. Lindgren et al., 2018. Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur).
В коллекции из Якутии оказались и остатки молодых плезиозавров. Судя по мелким размерам и плохому окостенению, возможно, что это кости даже младенческих особей. По словам одного из авторов статьи, Николая Зверькова, молодым особям принадлежит четверть найденных позвонков и примерно половина костей конечностей (шесть от молодых особей и семь от взрослых). Самая маленькая кость сохранилась почти целиком: в длину она достигает 10 сантиметров, ее обладатель даже с учетом длинной шеи едва ли превышал в длину 1–1,5 метра, в то время как крупные кости принадлежали более чем пятиметровым плезиозаврам.
Остатки молодых особей показывают, что морские рептилии (по крайней мере плезиозавры) использовали высокие широты как место для размножения. Подобное поведение описано для морских рептилий мелового периода, которые поднимались для размножения в высокие широты.
Летом в высоких широтах световой день длится долго (а во время полярного дня Солнце не заходит вообще). Сейчас это вызывает бурный рост фитопланктона и — далее по пищевой цепочке — увеличение численности беспозвоночных, привлекающих рыб и китообразных, которые приплывают в северные воды покормиться и вывести потомство. В юрских морях ситуация могла быть похожей, только вместо китообразных на вершине пищевой пирамиды находились морские рептилии.
Пока не удалось выяснить, обитали ли юрские морские рептилии в полярных морях круглый год или приплывали туда только на летний сезон. Авторы обсуждаемой статьи склоняются ко второму варианту, приводя следующие два аргумента. Во-первых, мигрируют почти все крупные морские позвоночные — в поисках пищи и к местам размножения. Черепахи, например, мигрируют на низких широтах, а киты мигрируют по всей планете (в том числе — и в высокие широты). Во-вторых, у морских рептилий в случае постоянного обитания в полярных морях должны были появиться адаптации к многомесячному существованию в условиях полной темноты (например, эхолокация, ночное зрение или какие-либо сенсорные органы). Такие адаптации сложно уловить на ископаемом материале, однако представляется маловероятным, что эти адаптации массово появились у нескольких родов ихтиозавров и плезиозавров, которым было проще не приспосабливаться к суровым условиям полярной ночи, а уплывать в более южные моря.
Кроме того, в отложениях зафиксирован как минимум один серьезный эпизод похолодания с вероятным оледенением. А покрытые льдом моря не могли подходить для морских рептилий, дышащих атмосферным воздухом.
Более правдоподобным выглядит сценарий, по которому ихтиозавры и плезиозавры на лето отправлялись в полярные регионы, размножались там, а с наступлением осени плыли обратно на юг. Так сейчас ведут себя многие перелетные птицы. Ну а в юрском периоде на территории нынешней Якутии плавали «перелетные» плезиозавры.
https://elementy.ru/novosti_nauki/433753/V_yurskom_periode_p...

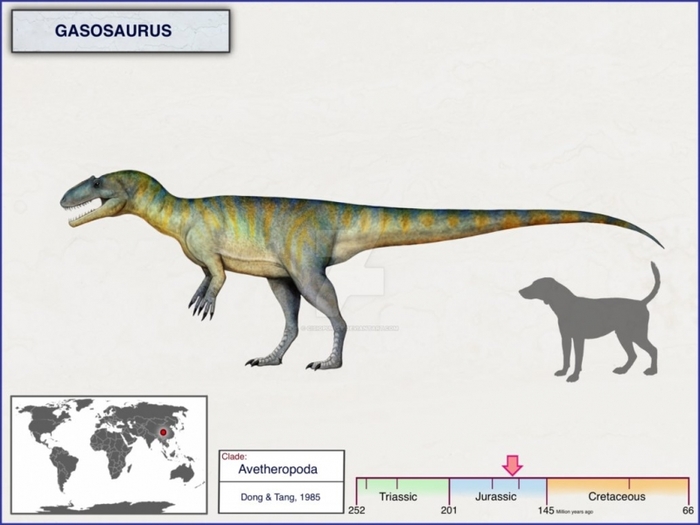
Газозавр (лат. Gasosaurus) — род хищных ящеротазовых динозавров, относящихся к группе Avetheropoda, живших в среднеюрском периоде (около 161,2—155,7 млн лет назад) на территории нынешней провинции Сычуань в Китае. Представлен единственным видом — Gasosaurus constructus. Найденный в тех же слоях теропод Xuanhanosaurus вероятно является крупной особью газозавра, а не отдельным родом.
Имел большой череп и сильные задние конечности. Длина 5 метров. Предположительно, был плотоядным. Название «газозавр» или «бензиновый ящер» было дано ящеру китайским ученым Ци-минг Донгом — в честь нефтеперерабатывающей компании Dashanpu, рабочие которой обнаружили окаменелые останки.
Gasosaurus жил в середине юрского периода, около 164 миллионов лет назад. Он сосуществовал с другими тероподами среднего размера, такими как Chuandongocoelurus и Kaijiangosaurus, двумя другими динозаврами, обнаруженными в формации Нижнего Шаксимяо (the Lower Shaximiao Formation). Эти три таксона относятся к средним и крупным тероподам и известны из соединенных посткраниальных скелетов. Китай, может похвастаться самым высоким разнообразием таксонов среднеюрских теропод всех размеров в сравнени с любой точкой мира. Большинство среднеюрских теропод из Китая являются «средними» по размеру, как и большинство среднеюрских теропод во всем мире.

Эти динозавры показывают интригующую комбинацию производных синапоморфий характерных для тетануров и примитивных черт, разделяемых с не тетанурами, это позволяет предположить, что они занимают базальное положение в Tetanurae. Понимание анатомии юрских таксонов из Китая может выявить дальнейшие подробности примитивной анатомии тетанур и помочь изучить раннюю эволюцию этой успешной клады теропод. Будущее исследование китайских среднеюрских теропод может дать новые представления о диверсификации Tetanurae, происхождении более крупных размеров тела среди более поздних теропод и палеобиогеографии средней юры.
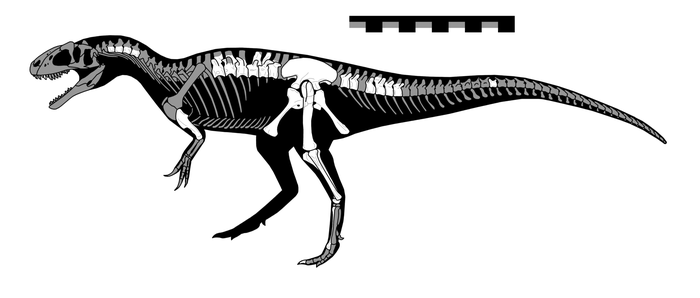
Скелетная реконструкция. Известный материал отмечен белым; остальные кости реконструированы на основе Allosaurus jimmadseni и Sinraptor dongi. Масштабная линейка равна 1 м.
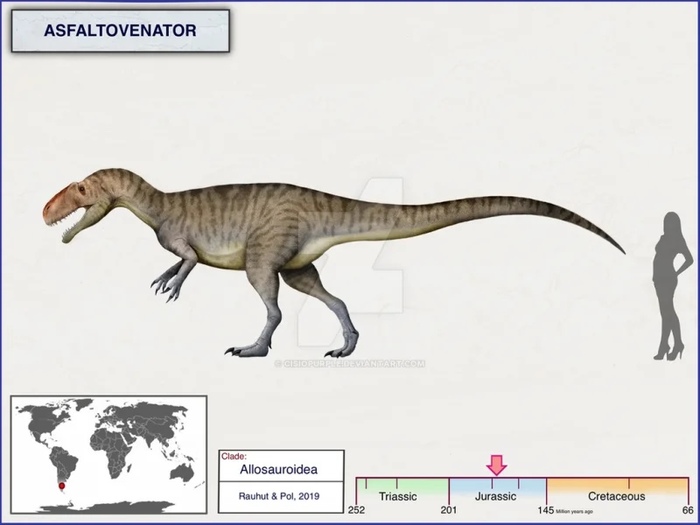
Асфальтовенатор (лат. Asfaltovenator) — ранний хищник из средней юры Южной Америки, обитал на территории Аргентины около 180–168 миллионов лет назад. Asfaltovenator — род крупных базальных тетануров. Остатки (почти полный череп и фрагментарный скелет) найдены в Аргентине в среднеюрской формации Cañadón Asfalto Formation. По состоянию на 2019 год это был самый полный известный скелет раннего тетанура. Возможно это самый примитивный представитель Allosauroidea. Назван он в честь формации Cañadón Asfalto, где был найден.
В 2002 году Леандро Канеса в 1,6 км к северу от городка Серро Кондор, в провинции Чубут на юге Аргентины, обнаружил окаменелости тероподового скелета. Раскопки продолжались в период с 2005 по 2007 годы, после чего ископаемое в виде огромного каменного блока было отправлено в лабораторию, где обрабатывалось препаратором Мариано Каффой в течении последующих пяти лет из-за чрезвычайной твердости каменной матрицы. Между позвоночником и передними конечностями теропода была обнаружена большеберцовая кость зауропода. В 2019 году немецкий палеонтолог Оливер Раухут из отделения палеонтологии позвоночных Мюнхенского университета и его аргентинский коллега Диего Пол описали типовой вид Asfaltovenator vialidadi, название образовано от формации Каньядона Асфальто, где был обнаружен экземпляр и лат. venator - «охотник». Название вида образовано от наименования аргентинского национального транспортного министерства Dirección Nacional de Vialidad, оказавшему значительный вклад и помощь Палеонтологическому музею Эгидио Ферульо в извлечении ископаемых.
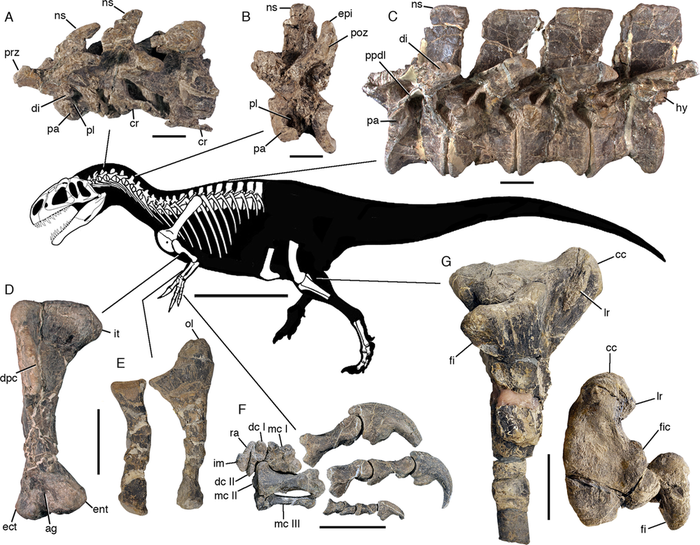
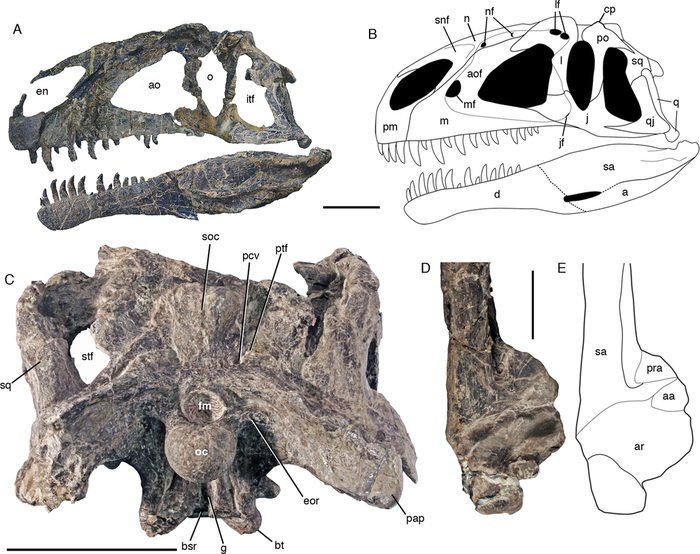
Голотип MPEF PV 3440 состоит из элементов передней части скелета и черепа с нижней челюстью, десяти шейных позвонков, тринадцати спинных позвонков, первого крестцового позвонка, полного плечевого пояса, и обеих передних конечностей; а также дистальной части лобковых костей, и частичной правой задней конечности, состоящей из дистальной части бедренной кости с элементами большеберцовой кости, малоберцовой кости, и частичной стопы.[
Асфальтовенатор был довольно крупным животным, сравнимым по размерам с известным аллозавром. Размер черепа голотипа составляет 75-80 сантиметров, реконструированный череп высокий и слегка выпуклый, аналогично другим аллозавроидам, общая длина тела оценивается в 7-8 метров. В верхней челюсти было 13 верхнечелюстных зубов, которые удерживались на месте отделенными межзубными пластинками. Нижняя челюсть содержала 14 зубов.
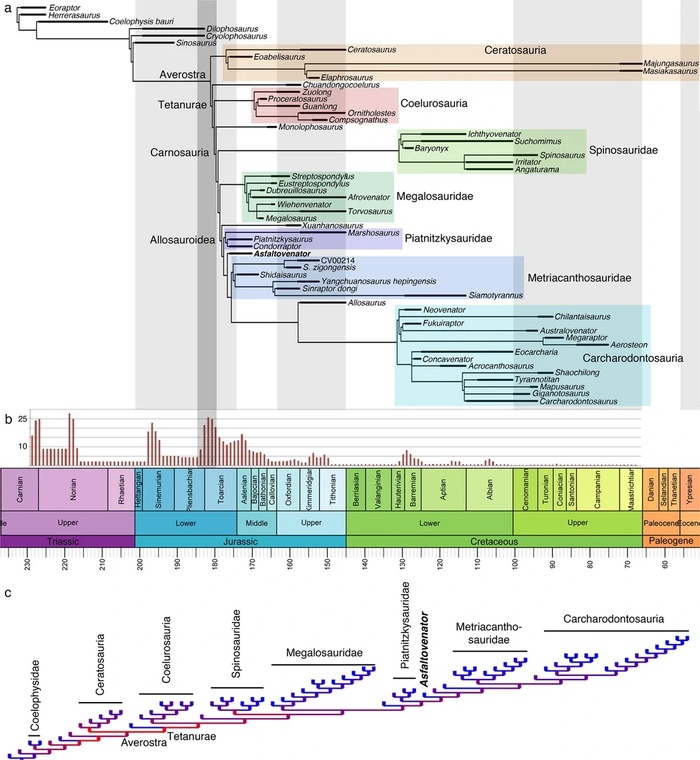
Обнаруженная мозаика примитивных и производных тетануровых символов филогенетического анализа асфальтовенатора разрушает современные представления о филогенезе тетануров и подчеркивает большое количество конвергенций в ранней эволюции этой группы. Филогенетическая позиция нового таксона была проверена с использованием филогенетической матрицы данных, включающей 66 таксонов и 355 остеологических признаков. Уникальная комбинация характеристик наблюдаемая у асфальтовенатора, указывает на то, что мегалозавроиды и аллозавроиды имеют общего предка, не разделяемого с тетанурановой группой целурозаврия (Coelurosauria). Тетануры представляют собой наиболее разнообразную кладу тероподных динозавров, которая включает в себя не только большинство известных мезозойских теропод, таких как аллозавр или тираннозавр, но и современных птиц. Считается, что в начале своей эволюционной истории эта группа разделилась на три основных клады: Megalosauroidea,Allosauroidea и Coelurosauria. Самые древние тетанураны встречаются в самом начале средней юры, но эти окаменелости очень редки. Асфальтовенатор является одним из старейших из известных и наиболее полных средне-юрских тетануранов, который имеет необычную комбинацию символов, объединяющую признаки, которые в настоящее время считаются апоморфными для различных тетанурановых линий. Филогенетический анализ приводит к монофилии группы карнозавров (Allosauroidea и Megalosauroidea).

Систематика
Найденный в Саратовской области позвонок принадлежал крупнейшему плиозавру России. В отложениях сеноманского возраста (100,5-93,9 млн лет) ранее ничего подобного не находили. Описание находки вышло в журнале Cretaceous Research.
Гигантский позвонок был найден несколько лет назад в районе поселка Нижняя Банновка в Саратовском Поволжье. Это место известно благодаря находкам одного из последних ихтиозавров (Pervushovisaurus bannovkensis) и первого ихтиорниса в России. Новая находка принадлежит к наиболее продвинутым плиозаврам подсемейства Brachaucheninae.
Исследуемый образец представляет собой шейный позвонок диаметром 19 см. Шейные позвонки такого размера известны только для трех самых крупных раннемеловых плиозавров – Kronosaurus queenslandicus из Австралии и Kronosaurus boyacensis и Sachicasaurus vitae из Колумбии. Эти крупнейшие плиозавры достигали 10-11 метров в длину, являясь самыми крупными хищниками в морях мелового периода.
Таким образом, сопоставимый по размерам позвонок из Саратовской области принадлежал крупнейшему плиозавру России. Однако наибольший интерес представляет возраст находки – сеноман (100,5-93,9 млн лет). До этого исследования считали, что к этому времени плиозавры сильно измельчали непосредственно перед вымиранием в следующем, туронском веке позднего мела. Новая находка позволяет говорить о том, что гигантские плиозавры, занимавшие вершины пищевых пирамид, существовали и в начале позднего мела. Возможно, их исчезновение как-то связано с вымиранием ихтиозавров в конце сеноманского века, но чтобы сделать однозначные выводы, необходимо больше находок из этого временного интервала.
Реконструкция сделана на основе самого полного скелета гигантского плиозавра Sachicasaurus vitae из нижнего мела Колумбии.
Публикация: Nikolay G.Zverkov, Evgeny M.Pervushov. A gigantic pliosaurid from the Cenomanian (Upper Cretaceous) of the Volga Region, Russia//Cretaceous Research, Available online 19 February 2020, https://doi.org/10.1016/j.cretres.2020.104419

Одна вакансия, два кандидата. Сможете выбрать лучшего? И так пять раз.
Многие знают, что млекопитающие мезозойской эры были мелкими и пушистыми. Они шныряли под ногами динозавров или вообще прятались в норках, ожидая ночи, чтобы выйти из укрытий, пока ужасные ящеры спят. По большому счёту так оно и было, но сейчас уже известно, что не все тогдашние звери были такими уж крохами. Конечно, слонов и носорогов в мезозое не было, но всё же некоторые млекопитающие были далеки от размеров землероек и мышей. Предлагаем вашему вниманию перечень некоторых из тех самых мезозойских млекопитающих «гигантов». Животные не расположены в строгом порядке уменьшения размеров, т.к. в большинстве случаев сделать точный расчет затруднительно.
Репеномам (Repenomamus).

Череп (вверху слева), скелет (вверху справа) и реконструкция внешнего вида (внизу) Repenomamus giganticus.
Репеномам – представитель триконодонтов (Eutriconodonta) из раннего мела Китая. Известно два вида рода Repenomamus. R. robustus, описанный в 2000 году имел череп длиной около 11 см. R . giganticus, описанный в 2005 году на сегодняшний день является крупнейшим мезозойским млекопитающим. Длина его черепа достигала 16 см, общая длина животного была около 1 метра, а весил он примерно 12-14 кг. Все триконодонты являются хищниками, питавшимися беспозвоночными и мелкими позвоночными животными. Изучив скелет R. robustus (более мелкого из двух видов), учёные обнаружили на месте желудка раздробленные кости детёныша мелкого рогатого динозавра пситтакозавра.
Винтана (Vintana sertichi).
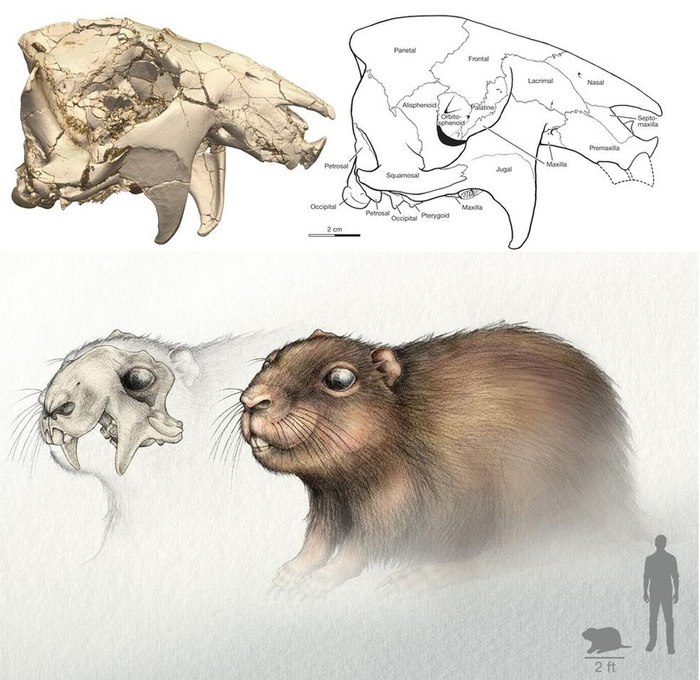
Череп (вверху) и реконструкция внешнего вида (внизу) Vintana sertichi.
Винтана, описанная в 2014 году из позднего мела Мадагаскара, относится к отряду гондванатериев (Gondwanatheria). Долгое время гондванатерии оставались весьма загадочными животными. Находка черепа винтаны позволила не только лучше изучить этих млекопитающих, но и показала, что некоторые из них были весьма крупными. Длина черепа достигает 12 см., а вес животного оценивается в 9 кг. Таким образом, винтана становится вторым по величине мезозойским млекопитающим и крупнейшим мезозойским зверем Гондваны. Винтана питалась растительной пищей, напоминая строением увеличенных резцов и высококоронковых щёчных зубов грызунов.
Дидельфодон (Didelphodon vorax).

Реконструкция внешнего вида (слева) и череп в различных проекциях (справа) Didelphodon vorax.
Дидельфодон, живший в конце мелового периода и описанный Ч. Маршем в 1889 г. , является древним родственником сумчатых (Metatheria), представителем семейства Stagadontidae. Долгое время именно дидельфодон считался крупнейшим млекопитающим мезозоя, на данный момент его можно назвать самым крупным млекопитающим позднего мела Северной Америки и, вероятно, крупнейшим териевым млекопитающим мезозоя. Размеры этого зверя сравнимы с виргинским опоссумом. Приблизительная масса тела взрослой особи оценивается в 5,2 кг. Дидельфодон обладал низким и широким черепом (длина - 122,1, ширина - 77,4 мм) и мощной челюстной мускулатурой. Вычисления показали, что дидельфодон обладал огромной силой укуса, что в совокупности с формой зубов (особенно премоляров) указывает на питание твердой пищей. Вероятно дидельфодон имел весьма разнообразную диету - он мог охотиться на мелких позвоночных, дробить раковины моллюсков и кости, а также употреблять растительную пищу.
Гобиконодон (Gobiconodon ostromi).
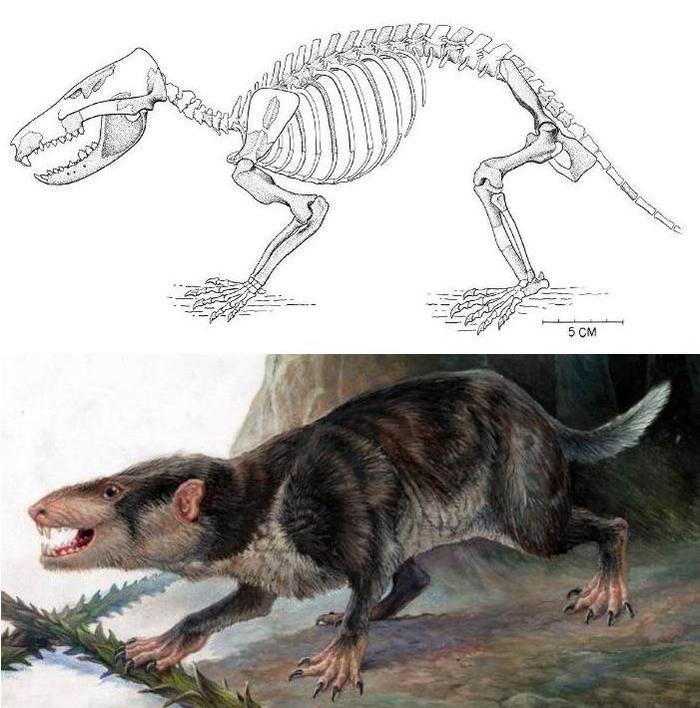
Скелет (вверху) и реконструкция внешнего вида (внизу) Gobiconodon оstromi.
Гобиоконодон является близким родственником репеномама. Известно несколько видов данного рода, которые в основном являются мелкими животными. Среди них выделяется G. ostromi, описанный в 1988 году из раннего мела Монтаны по весьма хорошо сохранившемуся скелету. Длина нижней челюсти достигает 7,8 см, а черепа – около 10 см. В целом по размерам данный вид гобиоконодона сопоставим с виргинским опоссумом, но отличался более крепким телосложением. Как и репеномам, гобиоконодон, по-видимому, питался мелкими позвоночными.
Стероподон (Steropodon galmani) и колликодон (Kollikodon ritchiei).
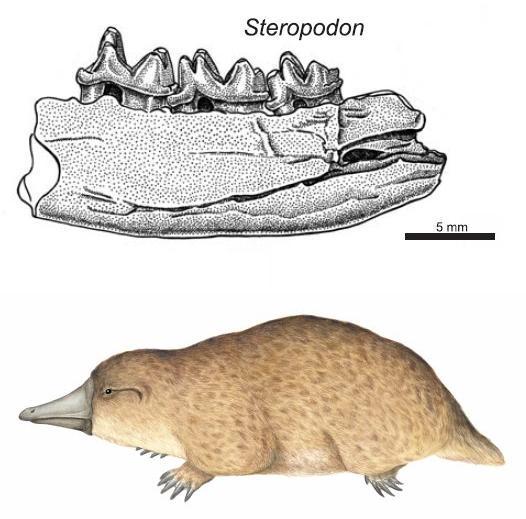
Фрагмент нижней челюсти (вверху) и гипотетическая реконструкция внешнего вида (внизу) Steropodon galmani.
Своими крупными мезозойскими млекопитающими может похвастаться и Австралия. В данном случае это были однопроходные, родственные современным утконосам. Стероподон, описанный в 1985 году из раннего мела Австралии, известен по небольшому (5 см) фрагменту нижней челюсти с зубами. Общая длина черепа могла достигать примерно 10 см, а сам зверь был около 35 см длиной, а возможно и больше. Судя по всему стероподон имел клюв, но в отличие от современного утконоса обладал зубами во взрослом состоянии, и вёл полуводный образ жизни.
Известны и другие крупные мезозойские млекопитающее Австралии, например колликодон, который также описан по фрагменту челюсти, длиной 3,5 см, с мощными широкими зубами. Он был несколько крупнее стероподона и сопоставим по размерам с современным утконосом. Хотя колликодон был описан как древний родственник утконоса, сейчас считается, что он вообще не является однопроходным.
Бубоденс (Bubodens magnus).

Зуб Bubodens magnus (вверху) и реконструкция головы Taeniolabis taoensis (внизу).
Бубоденс относится к мульттуберкулятам (Multituberculata), одним из наиболее многочисленных мезозойских млекопитающих, которые, кроме того, продолжили существовать в кайнозое, дожив до конца эоцена. Эти животные сходны с грызунами в строении зубов и возможно занимали аналогичные экологические ниши, будучи в основном растительноядными и всеядными. Мезозойские мультитуберкуляты в основном были мелкими, их масса не превышала 200 г, в палеогене они существенно подросли, достигнув размеров крупного бобра (род Taeniolabis). Однако бубоденс из позднего мела Южной Дакоты, описанный по одному зубу (левый m1) резко выделяется крупными размерами. Его масса оценивается в 5,25 кг, т.е. он был примерно с сурка. Бубоденс является родственником палеоценового тениолабиса, поэтому можно предположить, что он был сходен с ним внешне.
Шовалтерия (Schowalteria clemensi).
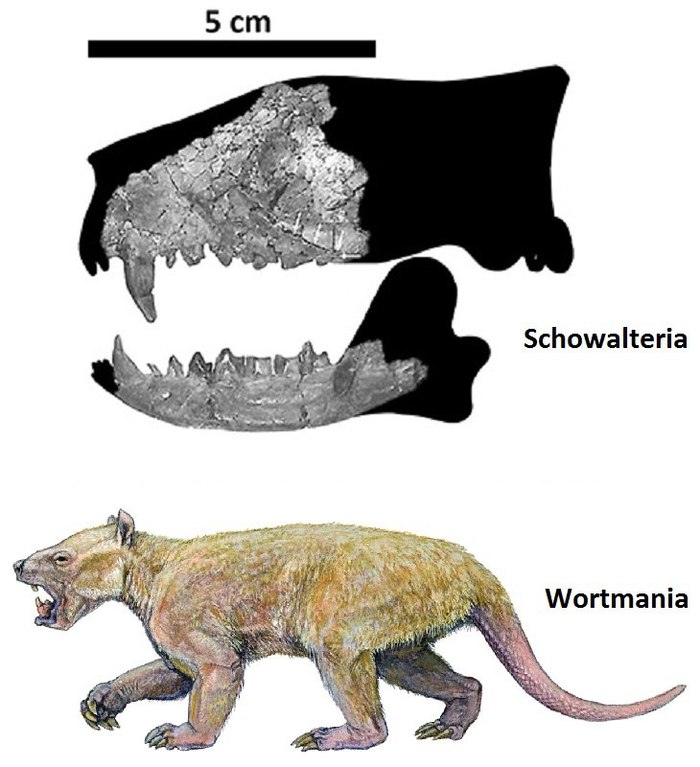
Реконструкция черепа Schowalteria clemensi (вверху) и реконструкция внешнего вида раннепалеоценового тениодонта Wortmania otariidens.
До этого речь шла о различных вымерших примитивных группах млекопитающих и о сумчатых. Но плацентарные, появившиеся ещё в юрском периоде, тоже смогли подрасти к концу мезозойской эры. Примером этого может служить шовалтерия (Schowalteria clemensi), из позднего мела Альберты (Канада), описанная в 2003 году. Она относится к тениодонтам (Taeniodonta), отряду примитивных млекопитающих, характерных для раннего палеогена Северной Америки. Поздние тениодонты были крупными растительноядными, весившими более 100 кг. От шовалтерии известна передняя часть черепа и нижняя челюсть. По размерам этот зверь был примерно сопоставим с дидельфодоном.
Альтакреод (Altacreodus magnus) .
Фрагменты челюстей и зубов Altacreodus magnus. Длина шкалы — 5,5 мм.
Данный зверь является представителем эвтериев (отряд Cimolesta, семейство Cimolestidae). Ранее считался одним из видов цимолестов (Cimolestes magnus). Известен из позднего мела США и Канады (Альберта, Монтана, Вайоминг, Северная Дакота, Саскачеван). Окаменелые останки представлены фрагментами верхних и нижних челюстей, зубами. Альтакреод является крупнейшим представителем цимолестов и может считаться одним из крупнейших позднемеловых млекопитающих Северной Америки, хотя по размерам он все же уступает дидельфодону. Моляры альтакреода были приспособленные для разрезания и характеризуются очень слабым износом вершин бугорков. Возможно этот крупный цимолест имел довольно узкоспециализированную диету и питался в основном мелкими позвоночными.
Окслест (Oxlestes grandis) и кудуклест (Khuduklestes bohlini)
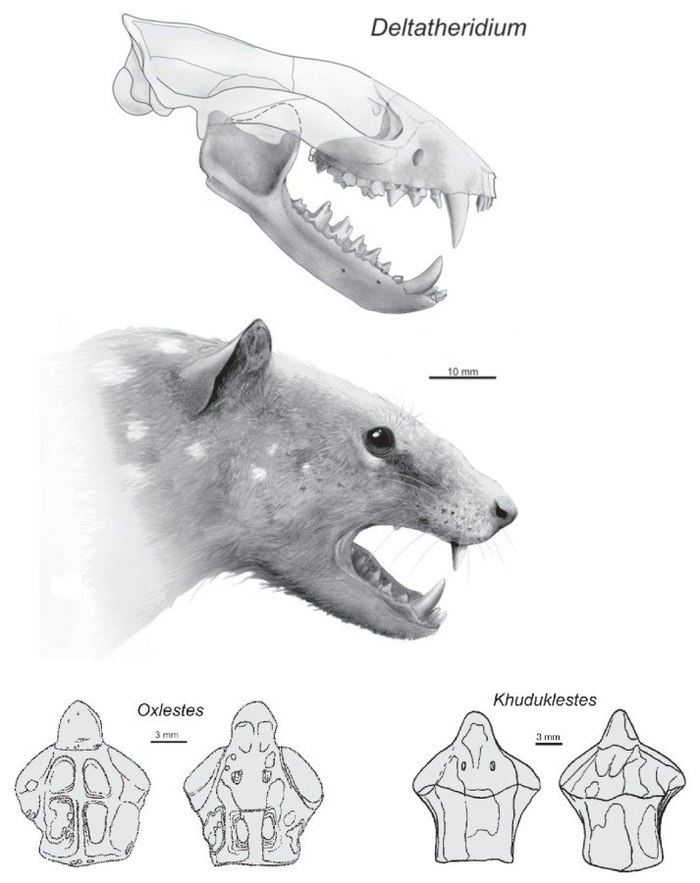
Реконструкция внешнего вида дельтатеридия (Deltatheridium), более мелкого представителя Deltatheroida из позднего мела Монголии (вверху). Шейные позвонки Oxlestes и Khuduklestes (внизу справа и слева соответственно).
Два очень плохо известных рода млекопитающих из позднего мела Азии. Останки окслеста обнаружены в Узбекистане (пустыня Кызылкум) и представлены неполным шейным позвонком (эпистофей), а также возможно фрагментом теменной кости с хорошо развитым саггитальным гребнем и крупным верхним клыком. Длина эпистрофея составляет 19,5 мм. Точные размеры установить очень сложно, но ясно, что это было весьма крупное млекопитающее для своего времени. Кудуклест также известен лишь по одному эпистрофею из Китая (провинция Гансю), достигающему длины 20,7 мм, что говорит о том, что он примерно сопоставим по размерам с окслестом. По всей видимости окслест и кудуклест были хищниками и могли охотиться на мелких позвоночных, в том числе на детенышей динозавров (например, мелких цератопсов, остатки которых обнаружены рядом с ними). По причине скудности ископаемых останков окслеста и кудуклеста сложно даже установить их систематическую принадлежность. Предположительно они относятся к дельтатероидам (Deltatheroida), группе базальных мететериев. Впрочем, это предположение основано только на размерах.
Винцелест (Vincelestes neuquenianus).
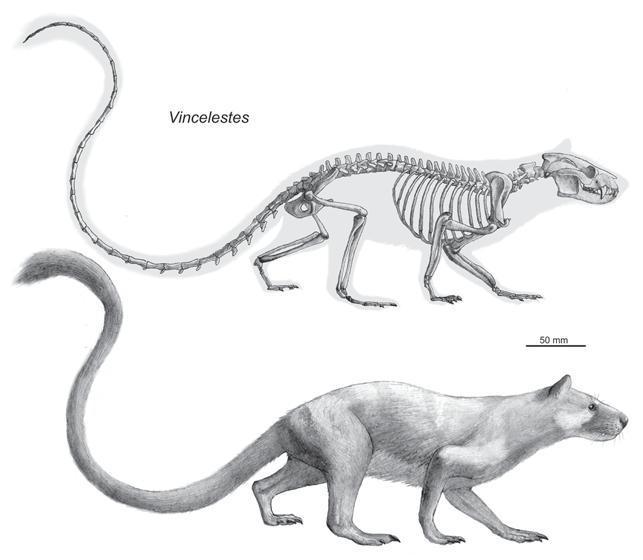
Скелет (вверху) и реконструкция внешнего вида (внизу) Vincelestes neuquenianus.
Винцелест из раннего мела Аргентины относится к группе «эупантотериев», которые близки к предкам сумчатых и плацентарных. Данное млекопитающее довольно хорошо представлено ископаемым материалом, включающим несколько целых черепов и множество элементов посткраниального скелета. Длина крупнейшего из известных черепов достигает 7,5 см. В своей экосистеме винцелест по-видимому был крупнейшим млекопитающим. Строение зубов и увеличенные клыки говорят о том, что винцелест являлся хищником, который мог питаться мелкими позвоночными.
Колониатерий (Coloniatherium cilinskii)

Фрагменты нижних челюстей Coloniatherium cilinskii.
Данное млекопитающее из позднего мела Аргентины (провинция Чубут) относится к группе млекопитающих под названием дриолестиды (Dryolestida), характерной для мезозоя Южной Америки и других континентов. Некоторые из них являлись мелкими древесными или наземными насекомоядными животными. Другие же (семейство Mesungulatidae, к которому принадлежит колониатерий) специализировались на всеядно-растительноядной диете и имели специализированные бунодонтные зубы, за что некоторых из них первоначально описывали как архаичных копытных кондиляртров. Мезунгулятиды эволюционировали на протяжении конца мезозоя, постепенно увеличиваясь в размерах. Ископаемые останки этих животных очень скудны. От колониатерия известны фрагменты нижних челюстей и зубы. По различным подсчетам длина черепа данного млекопитающего достигала от 87,5 до 161,5 мм, но первое значение больше согласуется с известными фрагментами нижней челюсти.
Касторокауда (Castorocauda lutrasimilis) и синоконодон (Sinoconodon rigneyi).
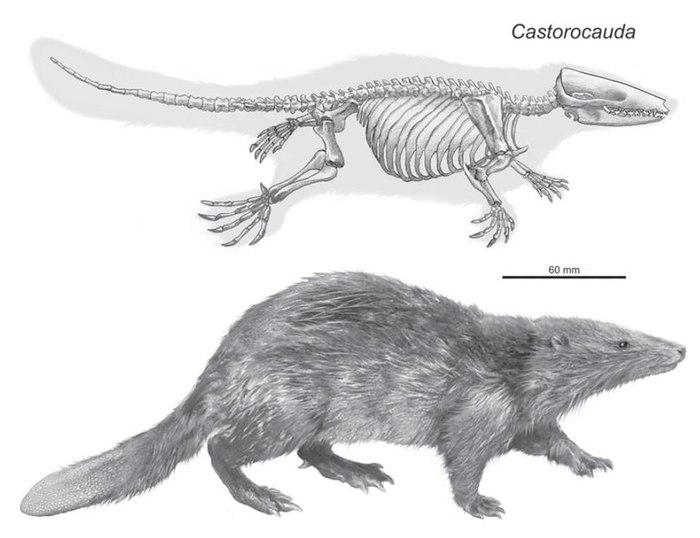
Скелет (вверху) и реконструкция внешнего вида (внизу) Castorocauda lutrasimilis.
Описанные выше животные обитали во время мелового периода, последнего из периодов мезозойской эры. Но и среди более древних протомлекопитающих (Mammaliaformes) встречаются те, кто превышал мышиные размеры. Одним из них является каторокауда, относящаяся к отряду докодонтов (Docodonta). Касторокауда описана в 2006 году из средней юры Китая по великолепно сохранившимся остаткам целого скелета. Она имела хвост, напоминающий бобровый, и вела полуводный образ жизни. Помимо этого касторокауда выделяется и своими размерами – её скелет достигает в длину 42 см, что очень немало для середины юрского периода.
Ещё раньше жил синоконодон, описанный из ранней юры Китая. Это протомлекопитающее, обладавшее рядом весьма примитивных признаков, имело череп длиной до 6,2 см, а его масса достигала около 500 г.
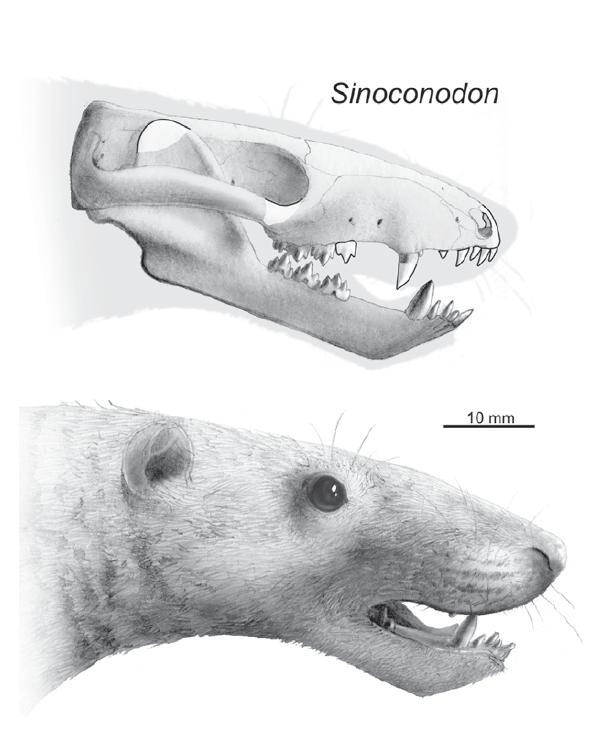
Череп (вверху) и реконструкция внешнего вида головы (внизу) Sinoconodon rigneyi.
