


Ископаемые остатки горгозавра (Gorgosaurus libratus)
Из верхнего мела (~80 млн лет) Альберты в лаборатории Королевского Тиррелловского палеонтологического музея, Канада.




Показать полностью
4
Из верхнего мела (~80 млн лет) Альберты в лаборатории Королевского Тиррелловского палеонтологического музея, Канада.

Французские палеонтологи описали древнейшего представителя отряда титаноптер, гигантских хищных насекомых, которые жили на Земле с перми до позднего триаса. Новый вид, получивший название Theiatitan azari, намного старше: отпечаток его надкрылья датирован 310 миллионами лет назад, то есть концом каменноугольного периода. Как отмечается в статье для журнала Communications Biology, T. azari также является самым древним известным насекомым, которое использовало крылья для производства звуков: судя по расположению жилок на крыльях этого вида, он трещал ими подобно некоторым современным саранчовым. А другие титаноптеры могли использовать специализированные структуры на крыльях, чтобы общаться с помощью вспышек отраженного света.
Насекомые нередко привлекают брачных партнеров и отпугивают конкурентов с помощью звуков. Например, многие виды из отряда прямокрылых (Orthoptera), включая широко известных сверчков и кузнечиков, громко стрекочут (точнее, стридулируют), используя в качестве музыкальных инструментов видоизмененные надкрылья, подкрылья и задние конечности. Другие насекомые издают треск, резко раскрывая крылья или стуча ими друг о друга. Судя по всему, впервые насекомые освоили акустическую коммуникацию еще несколько сотен миллионов лет назад. Древнейшие находки прямокрылых и их вымерших родственников с анатомическими структурами, которые могли использоваться для производства звуков, относятся к средней перми. А уже в триасовых отложениях такие виды встречаются регулярно.
К числу насекомых, которые, по мнению специалистов, одними из первых начали общаться или отпугивать хищников с помощью звуков, относятся титаноптеры (Titanoptera). Эта отдаленно родственная современным прямокрылым группа существовала с перми до границы триаса и юры и включала очень крупные, с размахом крыльев до тридцати с лишним сантиметров, хищные виды, отдаленно напоминающие богомолов. В средней части надкрыльев многих титаноптер ученые обнаружили расширенные области между жилками, которые, согласно популярной гипотезе, служили резонаторами и усиливали стрекот этих насекомых. Интересно, что по крайней мере у одного вида эти структуры были характерны как для самцов, так и для самок.
Команде палеонтологов, которую возглавил Андре Нэль (André Nel) из Национального музея естествознания в Париже, удалось получить новые ценные сведения об эволюции и биологии титаноптер. В руки исследователей попал добытый на севере Франции и датированный каменноугольным периодом образец скальной породы с частичным отпечатком единственного надкрылья насекомого. Анализ показал, что оно принадлежало виду из отряда титаноптер, причем древнейшему из когда-либо найденных. Обладатель надкрылья жил около 310 миллионов лет назад, на 50 миллионов лет раньше всех ранее известных представителей группы. На основе отпечатка ученые описали новый вид, который получил название Theiatitan azari. Родовое имя отсылает к титаниде Тейе из древнегреческой мифологии, матери Гелиоса, Селены и Эос.
По мнению Нэля и его соавторов, жилкование надкрылья T. azari свидетельствует, что данный вид использовал крылья для коммуникации с сородичами. На данный момент это самый древний вид насекомых, освоивший подобный способ общения. Предыдущий рекорд принадлежал Permostridulus brongniarti, насекомому с не до конца ясным систематическим положением, которое жило на территории современной Франции примерно на 50 миллионов лет позже T. azari и, вероятно, стридулировало крыльями.
Впрочем, авторы не уверены, как именно T. azari и другие титаноптеры общались с помощью крыльев. Традиционную точку зрения, согласно которой эти насекомые стрекотали надкрыльями подобно сверчкам, они считают сомнительной. Дело в том, что, несмотря на наличие «резонаторов» разного размера на надкрыльях титаноптер, никому до сих пор не удавалось обнаружить на их крыльях, конечностях или теле анатомические структуры, необходимые для производства стрекота. Кроме того, строение крыльев титаноптер исключает, что они могли стучать крыльями. При этом расположение жилок на надкрыльях некоторых титаноптер, включая T. azari, напоминает таковое у насекомых из надсемейства саранчовых (Acridoidea), которые умеют производить треск крыльями во время полета или резко раскрывая их (этот процесс называют крепитацией). Большинство титаноптер, судя по соотношению размаха крыльев и длины тела, были плохими летунами, однако они могли трещать крыльями, сидя на ветке дерева.
Нэль и его коллеги также допускают, что крылья служили отдельным видам титаноптер не для акустической, а для визуальной коммуникации. Некоторые современные насекомые, например, ряд видов прямокрылых, бабочек, стрекоз и двукрылых, общаются, производя вспышки света с помощью отражательных поверхностей на особых участках крыльев. Возможно, что аналогичное поведение было характерно и для титаноптер, например, для триасового Clatrotitan andersoni, чьи надкрылья кажутся особенно специализированными.
«Резонаторы» на крыльях титаноптерид были очень разнообразными по форме и размеру, так что одни виды этих насекомых, например, C. andersoni, вероятно, общались с помощью световых вспышек, в то время как другие, включая T. azari, трещали крыльями. Не исключено, что целый ряд видов использовал сразу оба способа коммуникации — к таким авторы относят триасового Gigatitan vulgaris. Причем способность производить звуки могла служить титаноптерам не только для привлечения партнеров и отпугивания конкурентов, но и в качестве защитного механизма против хищников.
Современные технологии позволяют больше узнать о биологических особенностях давно исчезнувших насекомых. Например, в прошлом году китайские палеонтологи определили структурную окраску некоторых видов ос, мух и жуков из бирманского янтаря, которые жили около ста миллионов лет назад. Судя по всему, эти существа были зеленовато-синими.

Реконструкция Гигатитана.
Гигатитан — род титаноптер из триаса республики Кыргызстан. Гигатитана нашли в 1968 году в Киргизсской ССР в мадыгенской свите. Тело длинное, размах крыльев превышает 30 см. Гигатитан был одним из самых крупных представителей титаноптер.
Охотились на мелких насекомых и ящериц.
Из разных мест Подмосковья притащил два камня, словно бы погрызенных червями. Камни довольно твердые, на обычный известняк не похожи. Размер отверстий - примерно 2-3 мм, кроме дырок имеются отпечатки мелких раковин, скелетики всякой мелкой морской нечисти, т.е. каменюка достаточно древняя и морская. Поиск по картинкам забит всякой трипофобской дрянью, сам определить не смог. Что это такое?

Пояснительная надпись: "Эти костяные шарики росли в коже стегозавра, защищая горло от хищников".


Мумифицированная нога эдмонтозавра; образец имеет собственное имя "Дакота". Хорошо видны отпечатки чешуйчатых кожных покровов и копыто на среднем пальце.

Образец пситтакозавра (Psittacosaurus, SMF R 4970), сохранивший наружные покровные структуры в виде длинных трубкообразных образований, а также пигментацию.

Редчайший случай: во время препарации челюстной кости тираннозавра один из техников Королевского тирреловского музея (Канада) обнаружил ископаемую сосновую шишку, застрявшую в верхнечелюстном отверстии образца.


Гнездо птицы, прекрасно сохранившееся в травертине. Плейстоцен Германии.


Образец птерозавра рамфоринха с сохранившейся полостью мозговой коробки.

Синомегацерос (Sinomegaceros pachyosteus) - плейстоценовый олень из Северного Китая.



Этот слепок скелета тираннозавра, известного под именем Вайрекс (Wyrex), установлен в Музее округа Картер в Монтане. Образец известен отсутствием кончика хвоста, который, как считается, откусил другой тираннозавр.

Интересный факт: голотип Allosaurus jimmadseni (вид аллозавров, описанный в 2020 году) содержит слепок кератиновой оболочки, которая сохранилась на когтях большого пальца!
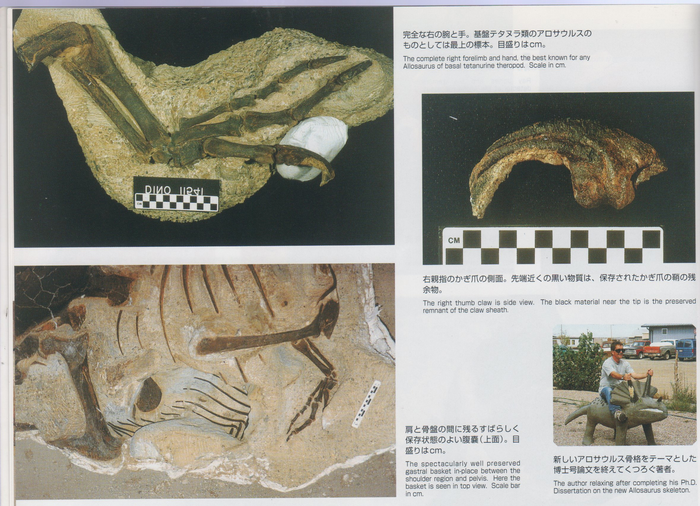
Раковина юрского аммонита Lytoceras fimbriatum, Франция. Размеры 21,6 см.

Окаменевшая раковина мезозойской улитки.

В Музее геологии горной школы Южной Дакоты имеется одна из самых крутых окаменелостей! Это ореодонт, #Merycoidodon culbertsoni, мать с детёнышами-близнецами, сохранившимися в утробе! Ореодонты представляют собой вымершее надсемейство доисторических жвачных парнокопытных, с короткими черепами и похожими на клыки зубами. Большинство ореодонтов были размером с овцу, хотя некоторые роды выросли до размеров крупного рогатого скота. Они были массивными, с короткими четырехпалыми копытами и сравнительно длинными хвостами.

Череп и передняя часть скелета завроктона (Sauroctonus progressus) из местонахождения Семин Овраг.
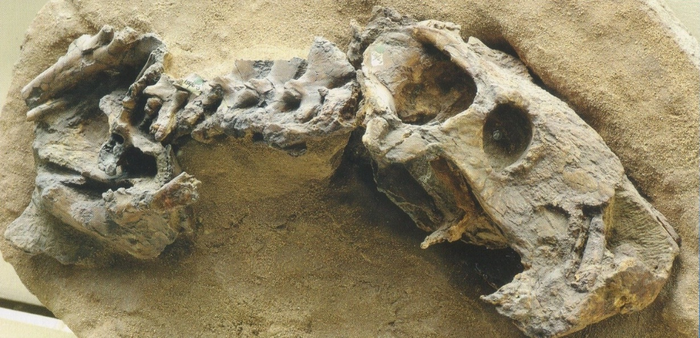
Завроктон (лат. Sauroctonus, от др.-греч. σαυροκτόνος — убивающий ящериц) — некрупный горгонопс из поздней перми Среднего Поволжья и Танзании. Входит в состав фауны Ильинского субкомплекса, предшествующего по времени знаменитым Северодвинским отложениям.
Впервые окаменелости, принадлежащие завроктону, были описаны в 1938 году Александрой Паулиновной Гартман-Вейнберг, крупным специалистом по пермо-триасовой фауне, которая отнесла обнаруженный в Татарстане неполный скелет к южноафриканским горгонопсам рода арктогнат (Arctognathus), вдобавок дав не слишком удачное видовое название progressus («прогрессивный») из-за предполагаемой близости завроктона к цинодонтам, прямым предкам современных млекопитающих. На деле же оказалось, что этим самым цинодонтам завроктон ничуть не более родственен, чем прочие саблезубые горгонопсы, и всего через пару лет после открытия один из сотрудников Гартман-Вейнберг, выдающийся палеонтолог и писатель Иван Антонович Ефремов, отнес этого звероящера уже к европейскому роду иностранцевий (Inostrancevia), а еще через полтора десятка лет, в 1955 году, друг Ефремова, Алексей Петрович Быстров, наконец-то выделил многострадального завроктона в отдельный род, где он пребывает и поныне.
Реконструкция внешнего облика завроктона (Sauroctonus progressus). Автор — Д. Богданов.
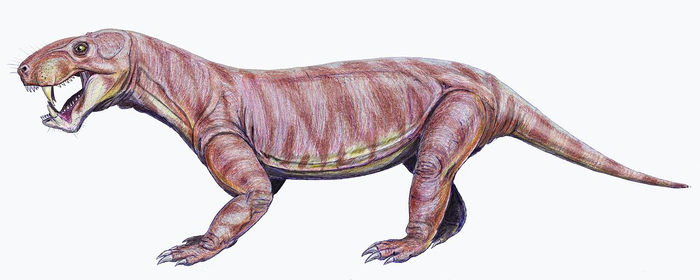
Длина черепа 22 см, общая длина до 1,2—1,5 м. Сохранял достаточно большое количество (5 пар на верхней и 6 на нижней челюсти) заклыковых зубов. Глазницы небольшие, верхний профиль черепа слегка изогнут, понижается кпереди. Максилла и носовые кости сильно скульптированы.
Виды.
Неопределённый Sauroctonus sp. был обнаружен в зоне скутозавров в месторождении Гороховец. Этот образец (нижняя челюсть) идентичен останкам типового вида.
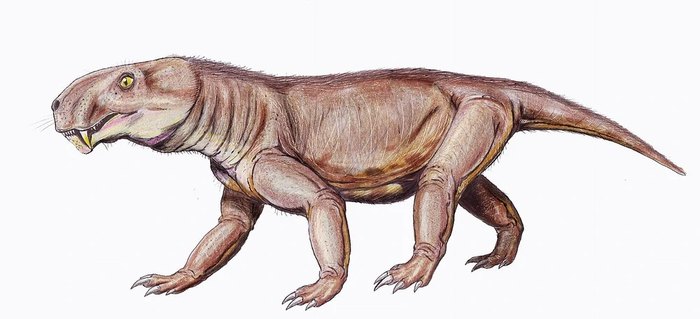
Sauroctonus parringtoni (von Huene, 1950) Gebauer, 2007 — вид из Танзании, обитавший в начале поздней перми. Первоначально описан Хюне как вид сциномогната, однако потом Сигонье определил его к элурогнату, а Гебауэр в 2007 году переописал его как завроктона. Длина черепа — 25 см. Череп относительно выше и шире, с более выгнутым верхним профилем и более массивным нижнечелюстным симфизом.
Sauroctonus parringtoni
Скелет завроктона (Sauroctonus parringtoni) из отложений грабена Рухуху.
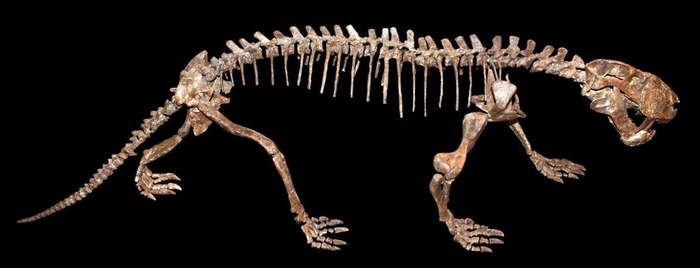
Таким образом, представители этого рода обитали как в Северном, так и в Южном полушарии, занимая нишу хищников среднего размера: у обоих видов длина черепа составляет около 25 сантиметров, а общая длина тела оценивается в 1,5–1,7 метра, это были животные размером с волка. К слову, современного волка завроктон напоминал не только габаритами, но и относительной подвижностью: в то время как у большинства крупных горгонопсов только задние конечности были полувыпрямленными, а локти передних торчали в стороны, как у ящериц, завроктон при необходимости мог полувыпрямлять все четыре лапы и даже переходить на галоп! (О такой возможности говорит строение суставов, в частности относительно подвижного плечевого сустава.)
Завроктон был одним из крупнейших хищников своей местности. Конкурировать с ним могли только подобные ему горгонопсы, вроде диногоргона или иностранцевии, а все крупные пеликозавры и диноцефалы вымерли к тому времени. Возможно, что для завроктона могли представлять опасность стаи мелких горгонопсов, вроде лиценопсов, в то время как крупные члены семейства охотились по одиночке. Вместе с завроктоном обитали ранние цинодонты процинозухи, тероцефалы, вроде мосхорина, протерозухид архозавр и темноспондильный ринезух. Ни один из них не мог представлять угрозу для крупнейших горгонопосов, а некоторые из них вообще могли служить пищей завроктону наряду с парейазаврами. В его рацион также могли входить проэлгинии, дицинодоны, иделезавры, а также тропидостомы.
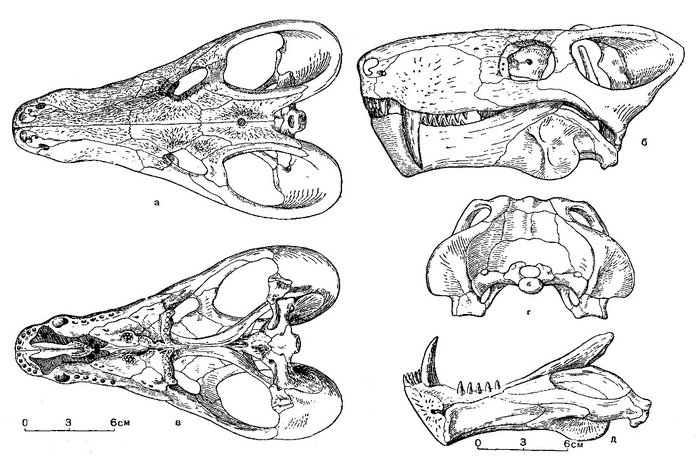
Прорисовки реконструированного черепа завроктона (Sauroctonus progressus) из местонахождения Семин Овраг: сверху (а), сбоку (б), снизу (в), сзади (г), нижняя челюсть изнутри (д).
Скелет завроктона (Sauroctonus parringtoni) из отложений грабена Рухуху.

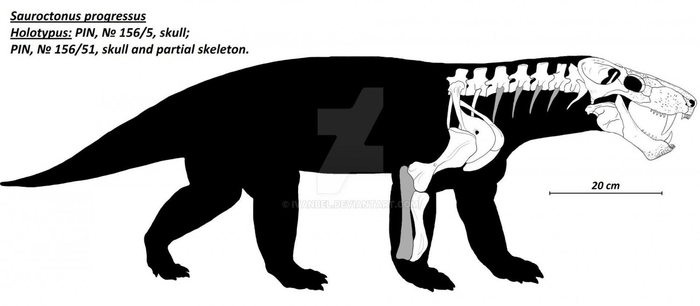
Прорисовка частично реконструированного скелета завроктона (Sauroctonus progressus) из местонахождения Семин Овраг. Автор — Ivanbel.
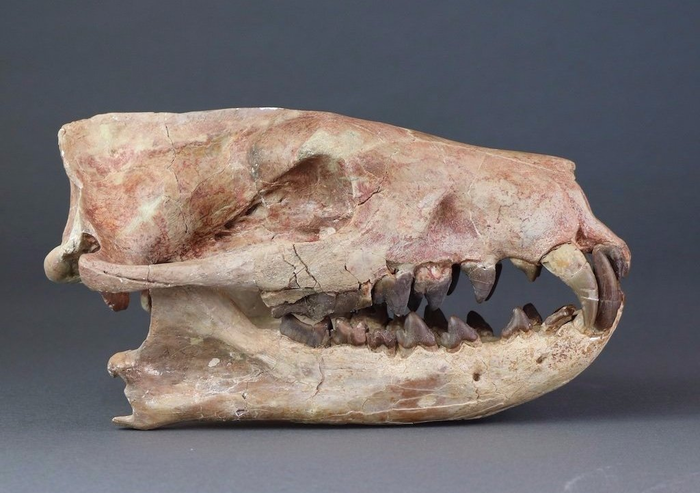
Череп завроктона (Sauroctonus progressus) из местонахождения Семин Овраг.

Что это?
Нашел на отмели реки. Похоже на голову рыбы или змеи. Внутри видно пустые полости. Текстура похожа не чешуйки змеи или ящерицы.





Ископаемый морской еж Balanocidaris glandifera из верхней юры (киммериджский ярус; ~155 млн лет) Испании.



Ископаемый пресноводный краб Potamon sp., окаменевший в известковом туфе (травертине) на реке Денизли в Турции. Возраст около 400 тыс лет

Необычный гетероморфный аммонит Hyphantoceras reussianum из верхнего мела Германии.

Ископаемый череп гиенодона Hyaenodon horridus из олигоцена (~30 млн лет) Южной Дакоты. Длина около 28 см.
Раковина гетероморфного аммонита Didymoceras stevensoni из верхнего мела Северной Америки.

Отпечатки рыбки Bobasatrania sp. в конкреции триасового возраста из Мадагаскара.

Окаменелые свернутые трилобиты Paciphacops birdsongensis из нижнего девона округа Бентона (слева) и два Flexicalymene retrorsa из ордовика Огайо (справа).

Одна вакансия, два кандидата. Сможете выбрать лучшего? И так пять раз.
Начало позднего эоцена Азии. Mongolonyx robustus убил гиракодонта форстеркуперию (Forstercooperia).

Поздняя волна развития мезонихиевых (Mesonychidae) произвела в мир по-настоящему грозных чудовищ, одними из которых были монголониксы (Mongolonyx), обитавшие в позднем эоцене Азии. На сегодня известно два вида: монголоникс массивный (Mongolonyx robustus) и более поздний монголоникс длинночелюстной (Mongolonyx dolichognathus). Возможными предтечами для монголониксов выступают среднеэоценовые мезониксы (Mesonyx), являвшие собой среднеразмерных лёгких стремительных хищников. В дайльнейшем, с распространением редколесий, саванн и степей, потомки мезониксов укрупнились, и на открытые пространства вышли на охоту плотоядные громадины, сопоставимые в размерах с крупнейшими львами.
Реконструкция черепа и головы Mongolonyx dolichognathus.

Родовое наименование Mongolonyx переводится как «монгольский коготь».
Род четко диагностируется и обладает целым рядом характерных морфологических признаков. Так, от Mesonyx его отличают более значительные общие размеры, моляризованные Р4/4, а также относительно более тупые и большие коренные зубы. Его зубы также относительно крупнее и массивнее, нежели у Dissacus, Harpagolestes и большинства других представителей семейства. В отличие от Dissacus и Pachyaena, у Mongolonyx отсутствует цингулюм на верхних зубах и не отмечаются какие-либо следы метаконида на тригонидах нижних коренных. От Synoplotherium, Mongolestes и Harpagolestes данный род отличает более прямая нижняя челюсть с удлиненным и сравнительно узким симфизом.
Двое Mongolonyx robustus схватили бронтотериида протитана (Protitan). Вдалеке в страхе разбегаются пантодонты эудиноцерасы (Eudinoceras).

Важное отличие от большинства мезонихид (вероятно, за исключением Pachyaena) состоит в том, что нижние клыки Mongolonyx располагаются очень близко друг к другу. В отличие от Harpagolestes, коронки нижних предкоренных загнуты назад не столь выражено, а их эмаль-дентиновая линия не является дугообразной. Кроме того, на р4 крупные элементы параконида параллельны протокониду, в то время как у Harpagolestes и Pachyaena параконид образует угол с протоконидом. Расположенный антеропостериорно параконид m3 также четко отличает Mongolonyx от Harpagolestes. От Mongolestes представители рода отличаются наличием m3 и редукцией р1.

Монголониксы представляли собой крупных плотоядных животных. Длина их черепа превышала 50 см, что вполне сопоставимо с черепом аляскинского гризли. Исходя из пропорций Mesonyx, известного по более-менее полному скелету, можно предположить, что эти мезонихиды достигали примерно 2,3 м в длину (не учитывая хвоста) и более 1 м в холке. Однако гораздо больше вопросов вызывает не внешний облик, а предположительный образ жизни монголониксов.
Mongolonyx robustus, более ранний вид, имел череп 52 см в кондилобазальной длине (от затылочных мыщелков до переднего края межчелюстной кости). Верхняя часть черепа до глазниц не сохранилась, но с восстановленным затылком и резцами его полная длина вполне могла составлять до 60 см. Если монголоникс имел пропорции, подобные своему лесному предку мезониксу, его высота в холке могла быть примерно 1,3 – 1,4 метра. А вес взрослого хищника мог достигать 300 - 400 кг. В общем, это был громадный хищник с мощными челюстями. В развитии зубочелюстного аппарата монголоникс хоть и уступал настоящим сверхспециалистам гарпаголестам (Harpagolestes) и монголестам (Mongolestes), но силы его челюстей вполне хватало, чтоб съедать добычу полностью, разгрызая даже крупные кости. В составе одной с Mongolonyx robustus фауны начала позднего эоцена обитали бронтотериевые (например, Protitan), гиракодонты (например, Forstercooperia) тапироиды семейства лофиалетовых, примитивные свинообразные и пантодонты Eudinoceras. Эти растительноядные служили пищей для монголониксов. Примерно в одно время с ними в Азии обитало довольно немалое количество не менее (а то и более) громадных плотоядов – заболоченных мест придерживались эндрюсархи (Andrewsarchus), хонанодоны (Honanodon macrodontus) и паратриисодоны (Paratriisodon), в лесах разбойничали саркастодоны (Sarkastodon). Такого плотоядного мегазверинца, наверное, не собиралось нигде на одном участке суши вплоть до плейстоценовой Северной Америки.
К концу позднего эоцена монголониксы заметно измельчали - Mongolonyx dolichognathus уже на 15 – 20% уступал в размерах своему предшественнику. Наиболее целая находка – нижняя челюсть достигает примерно 35 см, тогда как длина нижней челюсти M. robustus составляла 49 см.
Судя по имеющемуся материалу, коронки их мощных щечных зубов не подвержены значительному износу, нижние предкоренные мало загнуты назад, эмаль-дентиновая линия не дугообразная (загнутые коронки премоляров млекопитающих хищников препятствуют выскальзыванию раскусываемых костей, а дугообразность эмаль-дентиновой линии указывает на повышенную прочность эмали их зубов). На первый взгляд, подобная зубная система кажется не слишком хорошо приспособленой к разгрызанию костей и сухожилий. Это довольно неожиданно для поздних мезонихид, в большинстве своем демонстрирующих выраженную костедробящую специализацию. Само собой разумеется, Mongolonyx существенно превосходит в этом отношении ранних представителей группы типа Mesonyx, однако на порядок уступает таким «специалистам», как Harpagolestes и Mongolestes.
В то же время, тупобугорчатые щечные зубы монголониксов все-таки достаточно массивны (особенно у M. dolichognathus). Установлено, что их корни сливались в своей верхней части, образуя под коронкой дентиновую платформу, что могло способствовать продлению функционирования зуба после стирания коронки. Весьма вероятно, что подобное строение зубов было направлено на сопротивление их изнашиванию и может свидетельствовать о питании очень твердой пищей, в частности о дроблении костей трупов. У других мезонихид, питавшихся более мягким кормом, такое приспособление отсутствовало — их корни не сливались, а разделялись непосредственно под коронкой зуба.
С другой стороны, близко расположенные нижние клыки монголониксов свидетельствуют в пользу адаптации к мощному укусу-отщипыванию, что наблюдается также у некоторых креодонтов (Sarkastodon, Megistotherium). Такие сближенные клыки существенно увеличивали силу укуса, по принципу аккумулирования давления на меньшей площади. Можно предположить, что эти хищники вкладывали всю свою мощь в один укус, вцепляясь в жертву намертво, а не наносили ей множественные раны.
Следующая примечательная особенность морфологии монголониксов — округлая головка их суставного отростка. При этом, судя по сохранившимся образцам, левая и правая мандибулярные ветви полностью срастались в симфизной области. В результате нижняя челюсть получала возможность достаточно свободно двигаться из стороны в сторону, избегая при этом вывиха в симфизе (интересно, что подобная способность развилась также у других выраженных мясоедов — креодонтов рода Hyaenodon). Не менее интересно наличие на сильно вытянутом небе специальных костных трубок, служивших для защиты дыхательных каналов. Они позволяли хищникам не задохнуться, когда те, на охоте или в процессе питания, захватывали в пасть большие куски плоти. По сходным причинам подобные образования вполне независимо развились и у ряда других хищников (к примеру, все у тех же гиенодонов).
Учитывая вышесказанное и принимая во внимание неповоротливость, вполне вероятную для столь массивных животных, можно предположить, что монголониксы являлись преимущественно охотниками на крупную добычу. Очевидно, их жертвами становились в основном громоздкие травоядные, например бронтотерии (особенно молодые), вполне обычные в то время. Очевидно, монголониксы были способны не только справиться с большой жертвой, но и полноценно ее утилизировать. При этом также более чем вероятно, что монголониксы при случае поедали найденные трупы или отнимали добычу у более слабых хищников (или оспаривали у равных по силе — примерно тогда же существовали крупные азиатские гиенодоны).


