Итог
Существует много общего между способами активации В-клеток и Т-клеток. У рецепторов Т- и В-клеток есть “распознающие” белки, которые распространяются за пределы клетки и которые невероятно разнообразны, потому что они производятся путем смешивания и сопоставления сегментов генов. Для рецепторов В-клеток эти белки распознавания представляют собой легкие и тяжелые цепи, из которых состоит молекула антитела. У рецепторов Т-клеток молекулами, которые распознают антиген, служат α- и β-белки. У рецепторов Т- и В-клеток есть цитоплазматические хвосты, которые слишком коротки, чтобы сигнализировать о распознавании, поэтому для этой цели требуются дополнительные молекулы. Для В-клеток эти сигнальные белки называются Iga и Igβ. Для Т-клеток передача сигналов включает комплекс белков, называемых CD3.
Чтобы активировать В- или Т-клетки, их рецепторы должны быть сгруппированы антигеном, потому что это сшивание объединяет многие из их сигнальных молекул в небольшой области клетки. Когда плотность сигнальных молекул достаточно велика, запускается ферментативная цепная реакция, которая передает сигнал “рецептор задействован” в ядро клетки. Там, в “мозговом центре” клетки, гены, участвующие в активации, выключаются или включаются в результате этого сигнала.
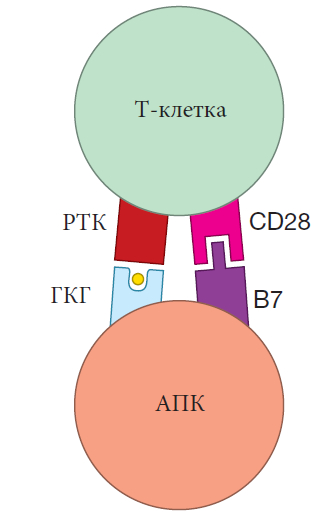
Хотя сшивание рецепторов необходимо для активации В- или Т-клеток, этого недостаточно. Наивные В- и Т-клетки также нуждаются в ко-стимулирующих сигналах, которые не антигенспецифичные. Это требование к двум сигналам для активации создает отказоустойчивую систему, которая защищает от неправильной активации В- или Т-клеток. Для активации В-клеток Т-клетка-помощник может обеспечить ко-стимуляцию с помощью поверхностных белков, называемых CD40L, которые подключаются к белкам CD40 на поверхности В-клеток. В-клетки также могут совместно стимулироваться “сигналами опасности”, включая специфические для вредителя молекулярные сигнатуры или боевые цитокины. Для Т-клеток совместная стимуляция обычно включает белки B7 на активированной дендритной клетке, которые взаимодействуют с белками CD28 на поверхности Т-клетки.
На ранних стадиях инфекции В-клетки и Т-клетки-киллеры могут активироваться без помощи Т-клеток-помощников. Беспомощные плазматические В-клетки вырабатывают антитела IgM, потому что они не переключились на класс антител, который подходит для защиты от конкретного вредителя.
Рецепторы В- и T-клеток могут связываться с молекулами корецепторов, которые служат для усиления сигнала от рецепторов. Для В-клеток этот корецептор распознает антиген, который опсонизирован комплементом. Если рецептор В-клетки распознает антиген, и если этот антиген также “украшен” фрагментами белка комплемента, антиген служит “зажимом”, который объединяет рецетор В-клетки и рецептор комплемента на поверхности В-клетки, значительно усиливая сигнал. Как следствие, В-клетки гораздо легче активируются (требуется сшивать гораздо меньше рецепторов) антигеном, который опсонизирован комплементом.
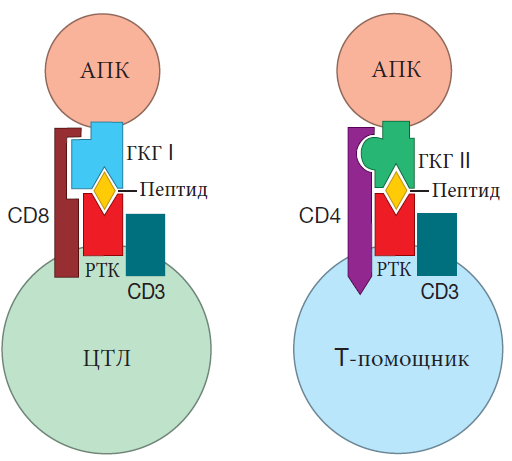
Т-клетки также оснащены корецепторами. T-клетки-помощники экспрессируют молекулы корецепторов CD4 на своей поверхности, а ЦТЛ экспрессируют корецепторы CD8. Когда рецептор Т-клетки связывается с антигеном, представленным молекулой ГКГ, корецептор на поверхности Т-клеток закрепляется на молекуле ГКГ. Это служит для усиления сигнала, который посылает рецептор в ядро, так что Т-клетка легче активируется (требуется сшивать меньше рецепторов). Эти корецепторы работают только с “правильными” типами ГКГ: класс I для ЦТЛ с корецепторами CD8 и класс II для T-клеток-помощников с корецепторами CD4. Следовательно, корецепторы действительно служат “фокусными” молекулами. Корецептор В-клеток помогает В-клеткам сосредоточиться на антигенах, которые уже идентифицированы системой комплемента как опасные (опсонизированы). Корецептор CD4 фокусирует внимание T-клеток-помощников на антигенах, проявляемых молекулами ГКГ II класса, а корецептор CD8 фокусирует ЦТЛ на антигенах, проявляемых молекулами ГКГ класса I.
Конечно, существует важное различие между тем, на что “смотрят” В-клетки и Т-клетки. Рецептор В-клетки распознает антиген в его “естественном” состоянии – то есть антиген, который не измельчен и не связан с молекулами ГКГ. Этим антигеном может быть белок или почти любая другая органическая молекула (например, углевод или жир). Напротив, αβ-рецепторы традиционных Т-клеток распознают только фрагменты белков, представленных классическими молекулами ГКГ. И в то время как рецепторы В–клетки связываются только с одним веществом – ее родственным антигеном, рецептор Т-клетки связывается как с представленным пептидом, так и с молекулой ГКГ. Поскольку набор антигенов, которые распознают рецепторы В-клетк, включает белки, углеводы и жиры, В-клетки могут реагировать на большее разнообразие вредителей, чем Т-клетки. С другой стороны, поскольку рецептор Т-клетки рассматривает небольшие фрагменты белков, он может распознавать мишени, которые скрыты от взгляда рецетора В-клетки в неповрежденном и плотно свернутом белке.
Еще одно различие между В-клетками и Т-клетками заключается в том, что во время инфекции рецпетор В-клетки может подвергаться соматической гипермутации и отбору. Таким образом, В-клетки могут “вытягивать из колоды”, чтобы попытаться получить лучшую комбинацию. Напротив, рецептор Т-клетки не гипермутирует, поэтому Т-клетки должны быть удовлетворены теми картами, которые им раздают.
Вопросы для размышления
В чем разница между ко-рецептором и ко-стимуляцией? Приведите примеры и расскажите, почему каждый из них важен для активации В- или Т-клеток.
2. Почему молекулы клеточной адгезии важны во время активации Т-клеток? Разве эти “липкие” молекулы просто не замедляют процесс?
3. Что происходит, когда дендритные клетки и Т-клетки-помощники “танцуют”?
4. По сути, все участники врожденной и приобретенной иммунной системой должны быть активированы, прежде чем они смогут “войти в игру”. Проследите этапы “активации”, которые начинаются, когда в рану попадает грамотрицательная бактерия, несущая ЛПС, и заканчиваются, когда вырабатываются антитела, способные распознать бактерию.
5. “Безотказная технология” используется для предотвращения неадекватной активации приобретенной иммунной системы. Можете ли вы привести несколько примеров?
Перевод книги LAUREN SOMPAYRAC "HOW THE IMMUNE SYSTEM WORKS", продолжение следует.