


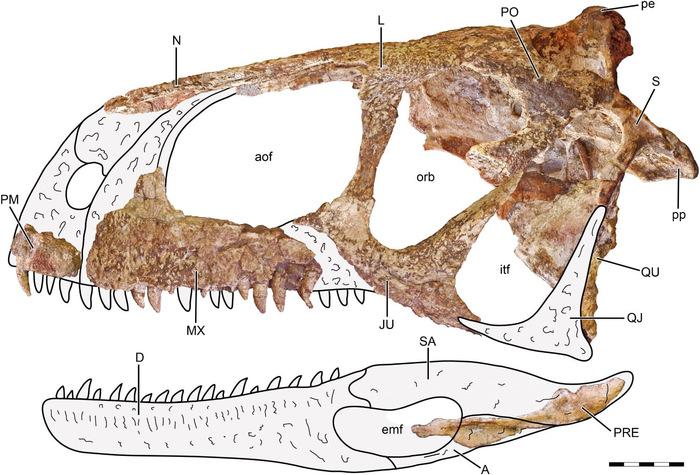
Череп аллозавра
Образец носит личное имя Эбенизер (Ebenezer). Этот череп имеет уникальную сохранность: 97 % его объёма сохранились в камне, даже подъязычные кости.

Образец носит личное имя Эбенизер (Ebenezer). Этот череп имеет уникальную сохранность: 97 % его объёма сохранились в камне, даже подъязычные кости.
Подумайте вот о чём: динозавры существовали ТАК ДОЛГО, что одни динозавры ходили по окаменелостям других динозавров! Например, тираннозавр мог ходить по окаменелостям аллозавра, поскольку их разделяли 84 миллиона лет!
Источник: https://vk.com/club175923038

Аргентинские палеонтологи обнаружили новый вид хищного динозавра, названного Llukalkan aliocranianus. Он относится к абелизавридам — группе двуногих крупных плотоядных динозавров, которые обитали в Южном полушарии, в то время как Северное населяли тираннозавры. Об открытии сообщается в статье, опубликованной в Journal of Vertebrate Paleontology.
Ллукалкан был одним из крупнейших хищников Патагонии, достигал пяти метров в длину, имел чувствительное обоняние и мощные челюсти, был вооружен острыми когтями на верхних и нижних конечностях. Череп ллукалкана имеет характерные выступы, делающие его похожим на некоторых современных рептилий, например, игуан. По оценкам исследователей, динозавр также отличался от других хищников чутким слухом.
Отличительной особенностью динозавра является небольшая задняя заполненная воздухом пазуха в зоне среднего уха, которая до сих пор не наблюдалась ни у одного другого абелизаврида. По мнению ученых, слух у него был похож на слух современных крокодилов.
Ллулкалан на языке мапуче (индейского народа в Чили и Аргентине) означает «тот, кто вызывает страх». Он населял Патагонию примерно 80 миллионов лет назад вместе с еще одним абелизавридом — Viavenator exxoni. Ископаемые останки ллукалкана и виавенатора были обнаружены всего в 700 метрах друг от друга в формации Бахо-де-ла-Карпа.
На сегодняшний день в Патагонии обнаружено почти десять видов абелизавридов. Эти динозавры обитали и в других районах древнего материка Гондваны, включая территорию современных Африки, Индии, Антарктиды и Австралии.

Пояснительная надпись: "Эти костяные шарики росли в коже стегозавра, защищая горло от хищников".


Мумифицированная нога эдмонтозавра; образец имеет собственное имя "Дакота". Хорошо видны отпечатки чешуйчатых кожных покровов и копыто на среднем пальце.

Образец пситтакозавра (Psittacosaurus, SMF R 4970), сохранивший наружные покровные структуры в виде длинных трубкообразных образований, а также пигментацию.

Редчайший случай: во время препарации челюстной кости тираннозавра один из техников Королевского тирреловского музея (Канада) обнаружил ископаемую сосновую шишку, застрявшую в верхнечелюстном отверстии образца.


Гнездо птицы, прекрасно сохранившееся в травертине. Плейстоцен Германии.


Образец птерозавра рамфоринха с сохранившейся полостью мозговой коробки.

Синомегацерос (Sinomegaceros pachyosteus) - плейстоценовый олень из Северного Китая.



Этот слепок скелета тираннозавра, известного под именем Вайрекс (Wyrex), установлен в Музее округа Картер в Монтане. Образец известен отсутствием кончика хвоста, который, как считается, откусил другой тираннозавр.

Интересный факт: голотип Allosaurus jimmadseni (вид аллозавров, описанный в 2020 году) содержит слепок кератиновой оболочки, которая сохранилась на когтях большого пальца!
Раковина юрского аммонита Lytoceras fimbriatum, Франция. Размеры 21,6 см.

Окаменевшая раковина мезозойской улитки.

В Музее геологии горной школы Южной Дакоты имеется одна из самых крутых окаменелостей! Это ореодонт, #Merycoidodon culbertsoni, мать с детёнышами-близнецами, сохранившимися в утробе! Ореодонты представляют собой вымершее надсемейство доисторических жвачных парнокопытных, с короткими черепами и похожими на клыки зубами. Большинство ореодонтов были размером с овцу, хотя некоторые роды выросли до размеров крупного рогатого скота. Они были массивными, с короткими четырехпалыми копытами и сравнительно длинными хвостами.

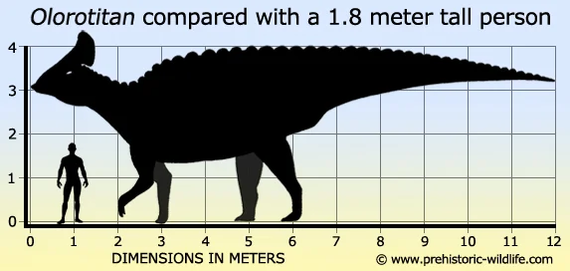
Олоротитан (лат. Olorotitan) — утконосый динозавр из России, обитал в меловом периоде 72 - 66 млн лет назад. Окаменелости этого динозавра были найдены в Кундурском костеносном месторождении, которое было обнаружено в 1990 году геологом Владимиром Нагорным в окрестностях поселка Кундур, Архаринского района Амурской области, при строительстве отрезка дороги Чита-Хабаровск федеральной трассы Москва-Владивосток. Тогда строители, работавшие на месте, замечали попадавшиеся кости, но были уверены, что это останки медведей. Масштабные научные исследования начались с 1998 года под руководством кандидата геолого-минералогических наук Юрия Болотского. В раскопках уже тогда участвовал сын ученого, Иван, который положил начало главному открытию - он обнаружил первые четыре хвостовых позвонка олоротитана. Олоротитан сосуществовал с родственными ему динозаврами харонозавром (лат. Charonosaurus) и базальным гадрозавридом амурозавром (лат. Amurosaurus). На том же участке были обнаружены окаменелости черепах, крокодилов, теропод и нодозаврид. Также на территории Амурской области в породах того же возраста были обнаружены останки керберозавра (лат. Kerberosaurus). В отличие от Северной Америки, где окаменелости ламбеозаврин практически не встречаются в отложениях маастрихтского яруса, в России и азиатских странах их останки датируемые этим времен широко распространены и разнообразны, что указывает на разницу в экологии и климате.
Юрий Болотский на раскопках Olorotitan arharensis, 1999 г

За первое лето на кундурском месторождении были найдены хвост из 75 позвонков и задняя часть туловища ящера. Раскопки в Архаринском районе велись три года. Самой большой интригой для палеонтологов оставался вопрос, отыщется ли голова динозавра. «Он действительно оказался гадом: нет чтобы вдоль трассы лечь, а он завалился перпендикулярно. Если хвост был на поверхности, то все туловище находилось в толще, а над ним четыре метра почвы. К тому же он был обложен изолированными костями, так как улегся, как плотина, на пути грязекаменного потока, и на нем были кости других тварей», — рассказывает Юрий Леонидович.
В 1999 году к раскопкам присоединился палеонтолог Паскаль Годефруа из Бельгийского Королевского института естественных наук, а в 2000 – сотрудники Палеонтологического института РАН из Москвы. Экспедиция увенчалась триумфом: под Кундуром впервые в России был обнаружен целый скелет одной особи, включая череп. Извлечение костей из породы продолжалось три полевых сезона с 1999 по 2001, а в 2003 году эта находка была описана палеонтологами Паскалем Годефруа, Болотским и Алифановым под назанием Olorotitan arharensis - «лебедь-титан».В 2012 году ими было опубликовано остеологическое исследование анатомии скелета и его отдельных костей.
Голотип (AEHM 2/845) состоит из хорошо сохранившегося скелета с неполным черепом. Хотя большинство костей черепа были слегка смещены, а кости носа сохранились не полностью, череп удалось довольно точно реконструировать. Эта находка - первый почти полный скелет динозавра в естественном сочленении, обнаруженный на Дальнем Востоке России. Реконструированный скелет этого динозавра экспонируется в музее Королевского института естественных наук в Брюсселе, Бельгия. Найденный олоротитан при жизни имел несколько ракообразных опухолей и, похоже, хромал на одну ногу, постольку пяточная и таранная кость были сросшимися с большой и малой берцовыми костями. Олоротитан многократно подвергался нападениям теропод. Имеются повреждения позвоночного столба, пояса конечностей, нижней челюсти и дистальных позвонков хвостового отдела.
Олоротитан - это самый полный экземпляр ламбеозаврин, обнаруженный за пределами Северной Америки. Этот динозавр был одним из самых крупных представителей своего семейства, таких как Parasaurolophus, достигая в длину 8 метров (26 футов). Он характеризуется специфическими для гадрозаврид признаками, самый очевидный из которых это уникальной гребень венчающий череп. Широкий, полый гребень, образованный из расширенных костей черепа, был пронизан носовыми путями и вероятно использовался для подачи трубных звуков. Сам череп поддерживается довольно длинной шеей, имеющий 18 позвонков, другие гадрозавриды имеют максимум 15 шейных позвонков. Филологическое исследование олоротитана привело к выводу, что он состоит в очень близком родстве с североамериканскими коритозавром (лат. Corythosaurus) и гипакрозавром (лат. Hypacrosaurus). Также на территории Амурской области в породах того же возраста были обнаружены останки другого гадрозаврида — амурозавра. В 1997 году в Благовещенске был создан палеонтологический музей. Реконструированная копия скелета олоротитана, включающая череп с предполагаемой формой гребня, демонстрируется в Амурском музее естествознания, в Благовещенске.
Олоротитан в Амурском палеонтологическом музее

Филологическое исследование олоротитана привело к выводу, что он состоит в очень близком родстве с североамериканскими коритозавром и гипакрозавром. Также на территории Амурской области в породах того же возраста были обнаружены останки другого гадрозаврида - амурозавра. В 1997 году в Благовещенске был создан палеонтологический музей. Реконструированная копия скелета олоротитана, включающая череп с предполагаемой формой гребня, демонстрируется в Амурском музее естествознания, в Благовещенске.
Олоротитан в Королевском институте естествознания, Бельгия

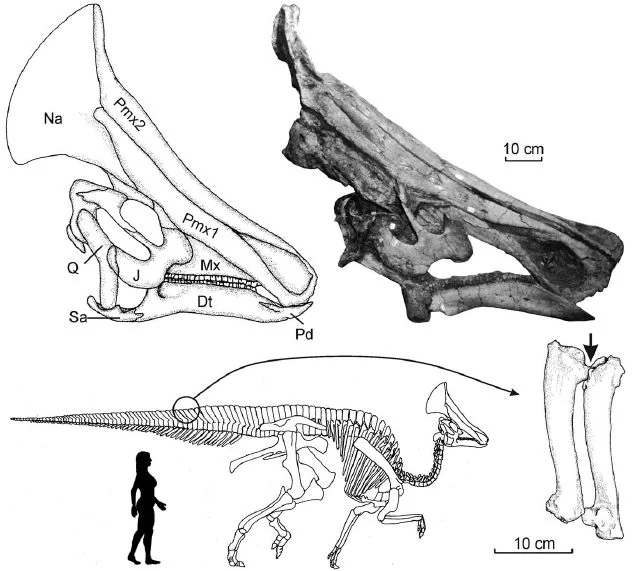


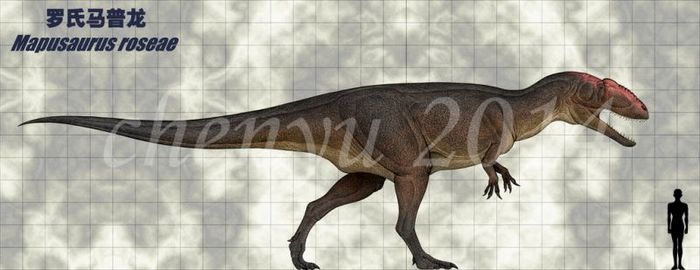
Мапузавр (лат. Mapusaurus) — род ящеротазовых динозавров из семейства Carcharodontosauridae группы тероподов, живших в меловом периоде (99,6—89,8 млн лет назад) на территории Южной Америки. Окаменелости теропода были найдены в Аргентине. Впервые описан палеонтологами Р. Кориа и Currie в 2006 году. Типовой и единственный вид — Mapusaurus roseae.
Mapusaurus roseae из позднего сеномана формации Huincul в Рио Лимай был описан 2006 году. Было обнаружено скопление костей нескольких разновозрастных особей, длиной от 5 до 10 метров. Вес динозавра мог доходить до 3 тонн. Это животное очень сходно с гиганотозавром, отличаясь более высоким и узким черепом.
Он был похож по размеру на своего близкого родственника Giganotosaurus, причем наибольшие известные особи оценивались примерно в 10,2 и более метра длиной и весом около 3 тонн. Самый длинный индивидуум, для которого Coria и Currie (2006) представили конкретную оценку, было животное, которому принадлежали бедра MCF-PVPH-208.203. Эта особь оценивается в 10,2 метра в длину.
Некоторые палеонтологи полагают, что этот двуногий плотоядный хищник мог бежать с максимальной скоростью до 30 миль в час. Если это так, то он был бы значительно быстрее, чем Tyrannosaurus Rex. Это также сделало бы его достаточно быстрым, чтобы уничтожить любого из травоядных, которые существовали в Южной Америке в то время.
Открытие семи индивидуумов Mapusaurus на одном и том же костяном ложе поставило под сомнение представление о том, что хищные динозавры всегда были одиночными существами. Когда это костяное ложе было раскопано между 1997 и 2001 годами, там были найдены только останки мапузавров разных возрастов. Это привело к тому, что животное стало считаться стайным хищником. Неизвестно, на кого охотился Mapusaurus, однако он родом из той же части мира, что и гигантский титанозавр Argentinosaurus. Взрослая особь Argentinosaurus была бы недопустной добычей для любого одиночного хищника того времени, однако стая хищников, нацеленная на ослабленных или юных особей, могла бы победить его за счет изнурения. Однако на сегодня это только гипотеза и останется ею до новых ископаемых находок.
Однако анализ места находок может поставить под сомнение потенциальное стайное поведение, посколько оно было интерпретировано как область осаждения. Это означает, что костный слой, возможно, был образован потоком или рекой, наполненными паводком. Вода могла переносить останки животных издалека и откладывать их группой. Если это так, то это была бы большая случайность, учитывая, что здесь были найдены только кости Mapusaurus.
Mapusaurus является членом группы Carcharadontosauridae, известной своими крупными членами, которые могут соперничать и даже превышать размерами более известного Tyrannosaurus rex. Зубы Mapusaurus, очень похожи на зубы родственного Giganotosaurus, они были плоскими и изогнутыми с зазубренным краем, идеально подходящими для разрезания плоти. Этим они отличаются от зубов Tyrannosaurus, которые были конусообразными и гладкими, адаптация, более подходящая для прокусывания костей.
Изучение зубов привело к дальнейшему усилению гипотезы о том, что Mapusaurus мог охотиться на динозавров, таких как Argentinosaurus, кости которых были слишком большими, чтобы перекусить. Если теория стайной охоты верна, то Mapusaurus, возможно, истощал аргентинозавра многочисленными укусами. Эти раны приводили бы к потере крови и инфекции, которые затем приводили к смерти огромных динозавров.

Mapusaurus — это комбинация слова на языке мапуче для земли и греческого слова для ящерицы. Название вида M. roseae имеет двойное значение, отсылающее к Роуз Летвин (Rose Letwin), спонсору раскопок и цвету камней, из которых был извлечен Mapusaurus.
Как упоминалось ранее, считается, что формация Huincul Formation представляет собой засушливую среду с эфемерными или сезонными потоками. Возраст этого образования оценивается в 97-93,5 млн. лет. Находки динозавров считаются здесь редкими. Mapusaurus разделял свою окружающую среду с зауроподами Argentinosaurus (один из крупнейших зауроподов, если не самый большой) и Cathartesaura. В регионе также жили абелисауроидные тероподы Skorpiovenator и Ilokelesia.

Кархародонтозавр (лат. Carcharodontosaurus) — гигантский хищный динозавр из Африки, живший в меловом периоде 100-94 млн лет назад на территории современной Африки.
Череп кархародонтозавра в сравнении с американским палеонтологом Полом Серено.

Первые два зуба были обнаружены на территории Алжира в 1924 году. Чарлз Депере и Саворнин в 1925 году отнесли их к роду Megalosaurus saharicus, но в 1927 году переименовали в Dryptosaurus saharicus. В 1914 году, в формации Бахариджа Египта были обнаружены разрозненные кости хищного динозавра, состоящие из обломка черепа, зубов, нескольких позвонков и бедерной кости задней конечности. Эти зубы были идентичны образцам описанным Депере и сильно напоминали зубы акулы, исходя из этого, Эрнст Штромер в 1931 году описал эти останки как Carcharodontosaurus saharicus — «ящер с акульими зубами». К сожалению эти образцы были утрачены во время Второй Мировой войны, в 1944 году.

Новые окаменелости были обнаружены палеонтологом Полом Серено в 1995 году, в формации Кем-Кем, Морокко, недалеко от алжирской границы, образец состоял из неполного черепа (образец SGM-Din 1). В 2007 году Стивен Брюссат и Пол Серено обозначили его как неотип, в том же году они описали новый вид Carcharodontosaurus iguidensis, основываясь на обломке челюсти с зубами из Нигера (образец MNN IGU2).
В 2010 году Грегори Пол предложил переименовать гиганотозавра в C. carolinii. Однако это решение не поддержали.
В 1960-ых годах к кархародонтозавру отнесли огромное количество разнообразных фрагментов меловых теропод, не имея для этого достаточных признаков. Вполне возможно, что в одно время с C. saharicus обитало несколько кархародонтозаврид, поэтому многие исследователи ограничивают образцы данного вида.

Челюсть Carcharodontosaurus iguidensis
Описание
Череп известен плохо, но напоминал таковой у гиганотозавра, зубы были с зазубринами и достигали 20 см в длину. В 1996 году Пол Серено оценил возможную длину черепа в 1,6 метра. Вероятно, кархародонтозавр мог охотится на крупнейших завропод - титанозавров, чьи кости найдены в тех же отложениях. Как и для гиганотозавра, для кархародонтозавра нет никаких доказательств стайного поведения. Кархародонтозавры были доминирующими плотоядными хищниками в Африке, с предполагаемыми размерами тела 10 - 12.8 метров в длину и 7.8 т веса.
Второй вид С. iguidensis был мельче и по оценкам учёных достигал 10 - 11 м и весил 4 - 5.2 т (Paul 2010, Molina-Perez и Larramendi 2019)
Пол Серено определил отличительные черты C. saharicus, включающие в себя приподнятый нижний край выпуклости возле заглазничного отверстия, глубокую направленную вниз канавку между внутренней стенкой верхней челюсти и передним внутренним отростком, высокий направленный вверх отросток между слёзной и лобной костью. Передний внутренний угол впадины возле верхней фенестры создаёт сжатую полость в крышке черепа. Челюстные зубы имеют характерные гребни на эмали.
В 2007 году Серено также определил уникальные половые разичия. Внешняя сторона верхней челюсти рифлёная, а возле места расположения сонной артерии на черепной крышке и вокруг основания затылка присутствовала обильная пневматизация. В нижней части черепной крышки находится отверстие с подверженным эрозии феноидом.
C. saharicus и C. inguidensis различаются строением слёзной кости и верхней челюсти, однако оба вида имеют бороздчатою поверхность зубов. У C. saharicusони больше, чем у C. iguidensis, но у второго мозг имеет на 15% больший объём.

Строение мозга
В 2001 году Ханс К. Ларссон опубликовал описание внутреннего уха и эндокринной системы C. saharicus. Начиная с самого близкого к концу челюсти начинается передний мозг, за которым следует средний мозг, наклонённый к задней части черепа под углом в 45 градусов. За ним следует ромбовидный мозг, параллельный переднему и образующий с средним мозгом сорокаградусный угол. Внешне мозг C. saharicus напоминал мозг аллозавра A. fragilis. Ларссон обнаружил, что отношение головного мозга кархародонтозавра к остальным отделам было типичным для не-авиальных динозавров. У него также был хорошо развит зрительный нерв.
Три полукруглых канала внутреннего уха C. saharicus имели субтриагулярный контур, если смотреть сбоку. Такая конфигурация также есть у аллозавра, ящериц, черепах, но не у птиц. Сами эти каналы имели линейное строение, что объясняет заострённый силуэт. При жизни флокулярная доля мозга располагалась возле полукруглых каналов, как у других карнозавров и птерозавров.


Зуб кархародонтозавра в Музее Естествознания в Вене
Систематика
Первоначально, в 1931 году, Стромер определили кархародонтозавра в собственное семейство кархародонтозавриды, но в 1934 изменил своё решение и переклассифицировал его в качестве аллозаврида. В 1995 году Оливер Раухут вернул кархародонтозавридам статус семейства.В 2009 году кархародонтозавра определили к африканской ветви семейства, однако поздние исследования ставят его ближе к южноамериканским кархародонтозавридам.
Необычным можно также и отметить сходства кархародонтозавра и с гиганотозавром, и с аллозавром. Кроме того, первый обитал вместе с мапузавром, тираннотитаном и другими крупными кархародонтозавридам на территории Южной Америки, а сам кархародонтозавр - в Египте, Марокко, Тунисе, Судане, Нигере. Аллозавра, которого считают предком кархародонтозаврид, обнаружили на территории Северной Америки, поэтому его потомки могли перейти на территорию Аргентины в мелу, но не в Африку. Не исключено, что предки кархародонтозавра являются дальними родственниками южноамериканских видов, перешедшими по сухопутному мосту на другие континенты. Теория кажется правдоподобной, однако её противоречат два факт - найденный ещё в 1925 году "Allosaurus" tendagurensis из Танзании, которого сейчас выделяют в отдельный род, и Allosaurus europaeus из Португалии. Эти виды уже обитали в Африке в юрском периоде, а A. europaeus вообще нашли недалеко от Северной Африки, опровергая собой теорию моста.

Кархародонтозавр от Андрея Аутчина
Палеобиология
В 2011 году палеоиллюстратор Боб Николлс опубликовал знаменитую работу, где два кархародонтозавра несли молодого райозавра. Картина вызвала многочисленные споры и обсуждения о возможноститакой охоты. Исследования Дональда Хендерсона, куратора динозавров Королевского музея Тирелла, показали, что строение челюстей, шеи и центра массы кархародонтозавра позволяли ему поднимать животных массой до 424 кг. То есть, его добычей могли быть некрупные завроподы, вроде молодых египтозавров, а также другие хищники - некрупные мегалозавриды или крокодиломорфы элозухи. Максимальный вес добычи кархародонтозавра составлял около 850 кг, что примерно сопоставимо с длиной в 8,3 метра.
Зубы кархародонтозавра напоминают зубы большой белой акулы, кархародона. Поэтому считается, что этот динозавр совершал быстрые, резкие укусы и вскоре отступал. А жертва умирала от потери крови.
У кархародонтозавра было типичное для теропод хорошее зрение. А его мускулистые ноги позволяли передвигаться со скоростью около 8,5 м/с, хотя эта оценка довольно спорна. С весом в 4 тонны падение на большой скорости было бы для кархародонтозавра смертельным. Строение ног не позволяло бы ему вовремя остановиться.

Патология
Один из образцов кархародонтозавра, SGM-Din 1, имеет колотую рану на переднем краю целюсти и "аномальную проекцию кости на заглазничное кольцо".
Палеоэкология
Останки кархародонтозавра находят почти по всей территории северной Африки: в формации Бахария, Египет, возле посёлка Марса-Мархут, тоже Египет, в формации Тегана и Ксар-эс-Суке, Марокко, в формации Ченини, Тунис, в Гарьяне, Ливия, в формации Кем-Кем, Алжир, и в Агадасе, Нигер. Вид C. iguidensis известен только по останкам из Нигера.
Кархародонтозавр обитал на жарком побережье древней Африки. Местность, где обнаружили окаменелости, была покрыта болотами и мангровыми лесами. Учитывая размеры кархародонтозавра, он был апексхищником, однако его связи с другими местными тероподами неизвестны. Точно так же неизвестна и настоящая добыча кархародонтозавра. Формация Кем-Кем была скудна на растительноядных животных, однако в другом месте обнаружения C. saharicus, формации Бахария, обитали крупные египтозавр и паралититан. Кархародонтозавр мог охотится на них, однако не на самых крупных особей.
Находка второго вида, C. inguidensis , показывает, что все образцы из области Фундортс, ранее причисляемые крупному C. saharicus, принадлежат нигерскому виду. Рядом с ним нашли также окаменелости ругопса и спинозавра. Маловероятно, что три крупных теропода могли обитать на одной территории, поэтому весь материал из этой области отнесли к C. iguidensis. Кроме теропод, негирийский вид соседствовал с крупными крокодиломорфами, вроде саркозуха, капрозуха, лаганозуха и арарипезуха.

Гигантозавр (лат. Giganotosaurus) — огромный хищник из Аргентины, обитавший в эпоху мелов ого периода 100-97 млн лет назад.

Окаменевшие останки были найдены Рубеном Каролини, охотником за ископаемыми, который 25 июля 1993 года обнаружил большую берцовую кость динозавра, выступающую из земли в месторождениях формации Рио Лимэй, в 18 км к югу от города Эль-Чокун в Патагонии, провинция Неукен, Аргентина. При извлечении окаменелостей выяснилось, что посткраниальный скелет был расчленен, а части черепа разбросаны на площади около 10 метров. В 1995 году аргентинские палеонтологи Рудольфо Кориа и Леонардо Салгадо описали эту находку под видом Giganotosaurus carolinii - «гигантский ящер» и назвали его в честь Рубена Каролини, нашедшего останки. В экземпляре (MUCPv-Ch1) сохранилось около 70 % костей, включая части черепа, таза, костей конечностей и большинство позвонков, первоначально размер особи был оценен в 12,5 метров (41 фут) в длину. Экземпляр хранится в палеонтологическом музее Эрнесто Бахманна.

Рубен Каролини и челюсть гиганотозавра (образец MUCPv-95)
В 1998 году Хорхе Кальво и Рудольфо Кориа описали обломок нижней челюсти, содержащий несколько зубов (образец MUCPv-95), признав его идентичным роду Giganotosaurus carolinii. Этот образец был найден в Сьерро Лос Кандильерос, провинция Неукен, в 1987 году и был кратко описан Кальво в 1989 году как вероятно неизвестный теропод. Этот образец был на 8% крупнее аналогичной кости голотипа, достигая 61 см в длину, и хотя эта часть челюсти является неполной, Кориа и Кальво предположили, что если череп голотипа (образец MUCPv-Ch1) достигал около 1,8 метра (6 футов) в длину, то найденный более крупный образец, вероятно, принадлежал черепу длинной 1,95 метра (6,4 фута), являясь самым крупным черепом теропода. Однако повторное исследование канадского палеонтолога Филлипа Карри и Рудольфо Кориа в 2002 году снизило максимальную длину черепа до 1,6 м. В 2004 году Джерард Маззетта с коллегами отметили, что хотя бедренная кость голотипа гиган
анотозавра была на 5 см (2 дюйма) больше, чем у тираннозавра "Сью", его голень была на 8 см (3 дюйма) короче.
В 2006 году палеонтологи Рудолф Кориа и Филлип Карри описали еще один вид - Mapusaurus roseae, образец состоял из нескольких костей, найденных в 1997 году. Гиганотозавр и мапузавр были объединены в одну группу Giganotosaurini.
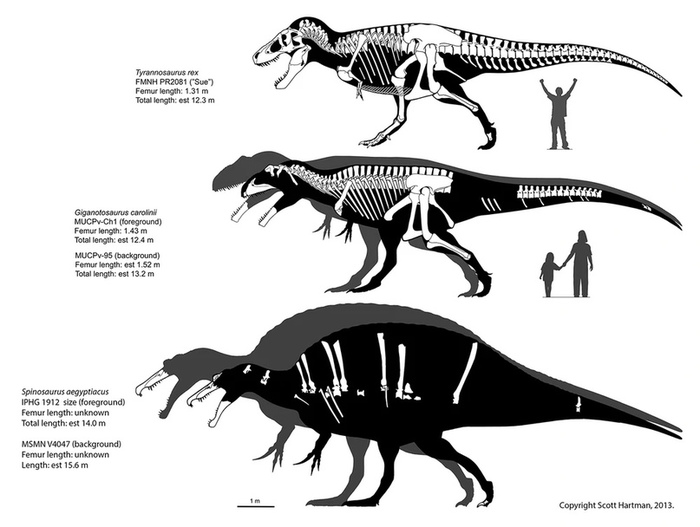
Гиганотозавр считается одним из крупнейших хищных динозавров. В 1997 году Рудольфо Кориа оценил его размер в 13,7-14,3 метра в длину и 8-10 тонн веса. В 2001 году Франк Сибахер, используя новый метод расчета размеров и массы тела, определил его размер в 12,5 метров и 6,6 тонны веса.В 2012 году Мэтью Т. Каранно и соавт. отметили, что хотя гиганотозавр и получил особое внимание из-за его огромных размеров, многие элементы его черепа не сохранились, что приводит к неопределенности и двусмысленности в определении общей длины черепа. Также они обратили внимание на то, что сами кости скелета, гиганотозавра и кархародонтозавра, были такого же размера, как и у тираннозавра. В отличие от кархародонтозавра, гиганотозавр обладал более высоким черепом с высокой закругленной передней частью, у него более развита орнаментация черепа — гребни на носовых костях, над глазами. В остальном эти животные очень схожи между собой. Следует отметить, что в отличие от тираннозавра, гиганотозавры не обладали достаточно развитым бинокулярным зрением, а их мозг был несколько меньше по объему, таким образом, гиганотозавры обладали более примитивными чертами по сравнению с тираннозавром.

Частичный череп голотипа MUCPv-Ch1(восстанавливаются белые части) с левым зубным фоном на заднем плане, Палеонтологический музей Эрнесто Бахмана
Хотя череп гиганотозавра не полностью известен, кажется, что он был низким. Верхняя челюсть имела длинный зубной ряд 92 см (36 дюймов), была глубокой сверху вниз, а ее верхний и нижний края были почти параллельны. Верхняя челюсть имела выраженный отросток (проекция) под ноздрей и небольшое эллиптическое отверстие (отверстие), как у аллозавра и тираннозавра. Носовая кость была очень морщинистой (грубой и морщинистой), и эти морщины продолжались в обратном направлении, покрывая всю верхнюю поверхность этой кости. Слезная кость перед глазом имела выпуклый морщинистый гребень (или рог), направленный вверх под задним углом. Гребень был похож на гребень и имел глубокие углубления. Посторбитальная кость за глазом имела направленный вниз и назад направленный отросток югального отростка, который проецировался на орбиту (открытие глаза), как это видно у тираннозавров, абелизавров и карнотавров. Надглазничная кость над глазом, которая контактировала между слезной и посторбитальной костями, была подобна карнизу и похожа на таковую у Ааелизавра. Квадратная кость в задней части черепа имела длину 44 см (17 дюймов) и имела две пневматические (заполненные воздухом) отверстия (отверстия) на внутренней стороне.

Частичные зубы, EBPM
Крыша черепа (образованная лобной и теменной костями) была широкой и образовывала «полку», которая нависала над короткими надвременными надколенниками в верхней задней части черепа. Челюсть сочленяется далеко позади затылочного мыщелка (где шея прикреплена к черепу) по сравнению с другими тероподами. Мыщелок был широким и низким и имел пневматические полости. У Гиганотозавра не было сагиттального гребня на верхней части черепа, и челюстные мышцы не распространялись на крышу черепа, в отличие от большинства других теропод (из-за полки над надвисковыми крыльями). Эти мышцы вместо этого были бы прикреплены к нижней боковой поверхности полки. Мышцы шеи, поднимающие голову, прикреплялись бы к выступающим надгробным костям на верхней части черепа, которые функционировали как гребень тиранозавров. Латексный эндокаст полости мозга Гиганотозавра показал, что мозг был подобен таковому из родственного рода Карахародонтозарид, но больше. Эндокаст имел длину 29 мм (1 дюйм), ширину 64 мм (3 дюйма) и объем 275 мл (9,7 импл. Унции).
Дентарий нижней челюсти расширен по высоте вперед (за счет нижнечелюстного симфиза), где он также был уплощен, и у него была выступающая вниз вершина (которая упоминалась как «подбородок»). Нижняя сторона зубного камня была вогнутой, внешняя сторона была выпуклой на виде сверху, и вдоль нее проходила канавка, которая поддерживала отверстия, которые питали зубы. Внутренняя сторона зубного ряда имела ряд межзубных пластинок, где на каждом зубе было отверстие. Мекелевская бороздка проходила вдоль нижней границы. Кривизна зубного ряда показывает, что устье Гиганотозавра было бы широким. Вполне возможно, что у каждого зубного камня было двенадцать альвеол (зубных щелей). Большая часть альвеол была длиной около 3,5 см (1,3 дюйма) спереди назад. Зубы зубной кости были одинаковой формы и размера, за исключением первого, который был меньше. Зубы были сжаты вбок, были овальными в поперечном сечении и имели зубцы на передней и задней границах, что типично для теропод. Зубы имели сигмовидную форму, если смотреть спереди и сзади. Один зуб имел от девяти до двенадцати зубцов на миллиметр (0,039 дюйма). У боковых зубов Гиганотозавра были изогнутые гребни эмали, а у самых больших зубов в предчелюстной кости (передняя часть верхней челюсти) были выраженные морщины (с их самым высоким рельефом около зубцов).
Шея Гиганотозавра была сильной, а осевая кость (шейный позвонок, который соединяется с черепом) была крепкой. Задние шейные позвонки имели короткие уплощенные центра («тела» позвонков) с почти полусферическими сочленениями (контактами) спереди и плевроцелями (полыми впадинами), разделенными пластинками (пластинами). Задние (дорсальные) позвонки имели высокие нервные дуги и глубокие плевроцели. Хвостовые позвонки имели нервные отростки, которые были вытянуты спереди назад и имели крепкую центральную часть. Поперечные отростки хвостовых позвонков были длинными спереди назад, а шевроны спереди были клинообразными. Грудной пояс был пропорционально короче тираннозавра, соотношение лопатки и бедренной кости меньше 0,5. Лезвие лопатки имело параллельные границы и сильный бугорок для введения мышцы трицепса. Коракоид был небольшим и имел форму крючка.
Подвздошная кость таза имела выпуклую верхнюю границу, низкое постацетабулярное лезвие (позади вертлужной впадины) и узкую полость грудины (выступ, где прикреплены мышцы хвоста). Лобковая нога была выражена и короче спереди, чем сзади. Седалищная кость была прямой и расширена назад, заканчиваясь в форме лепестка. Бедренная кость была сигмовидной формы и имела очень крепкую, направленную вверх голову с глубокой бороздкой (бороздкой). Меньший вертел головки бедренной кости был похож на крыло и располагался ниже большого вертела, который был коротким. Четвертый вертолет был большим и проецировался назад. Голень была расширена на верхнем конце, суставная грань (где она сочленялась с бедром) была широкой, а ее стержень был сжат спереди назад.
Кориа и Сальгадо первоначально обнаружили, что Giganotosaurus более тесно группируются с тетанурами, принадлежащими к тероподам, чем к более базальным (или «примитивным») тероподам, таким как кератозавры, из-за общих черт (синапоморфий) в ногах, черепе и тазу. Другие особенности показали, что он был за пределами более производной (или «продвинутой») клады Целурозавров. В 1996 году Серено и его коллеги обнаружили, что Гиганотозавр, Кархародонтозавр и Акрокантозавр тесно связаны с суперсемейством Алозавриди, и сгруппировали их в семью Кархородонтозарид. Особенности, общие для этих родов, включают слезные и посторбитальные кости, образующие широкую «полку» над орбитой, и квадратный передний конец нижней челюсти.
По мере того как было обнаружено больше кархародонтозавридов, их взаимосвязь прояснялась. Группа была определена как все аллозавроиды, более близкие к кархародонтозавру, чем аллозавр или синраптор, палеонтологом Томасом Р. Хольцем и его коллегами в 2004 году. В 2006 году Кориа и Керри объединили Гиганотозавра и Мапузавра в подсемейство Карахородонтозаврид Гиганотозарид, основываясь на общих чертах бедренной кости, таких как слабый четвертый трокантер и неглубокая широкая бороздка на нижнем конце. В 2008 году Серено и палеонтолог Стивен Л. Брусатте объединили Гиганотозавра, Мапузавра и Тираннотитана в племя Гиганотозавроидов. В 2010 году Павел перечислил Giganotosaurus как «Giganotosaurus (или Carcharodontosaurus) carolinii» без уточнения. Гиганотозавр является одним из наиболее полных и информативных представителей Кархородонтозарид.
Эволюция
Кориа и Сальгадо предположили, что конвергентная эволюция гигантизма у теропод могла быть связана с общими условиями в их среде или экосистемах. Серено и его коллеги обнаружили, что присутствие кархародонтозавридов в Африке (Кархародонтозавр), Северной Америке (Акрокантозавр) и Южной Америке (Гиганотозавра), показало, что группа имела трансконтинентальное распространение к раннему меловому периоду. Маршруты рассредоточения между северным и южным континентами, по-видимому, были прерваны океанскими барьерами в позднем меловом периоде, что привело к более отчетливым провинциальным фаунам путем предотвращения обмена. Ранее считалось, что меловой мир был биогеографически разделен, причем на северных континентах преобладают тираннозавриды, в Южной Америке - абелизавриды, а в Африке - кархародонтозауриды. Подсемейство Кархородонтозаврины, к которому принадлежит Гиганотозавр, по-видимому, ограничено южным континентом Гондвана (образованным Южной Америкой и Африкой), где они, вероятно, были высшими хищниками. Южноамериканское племя Гиганотозаврин, возможно, было отделено от их африканских родственников через викарий, когда Гондвана распалась в течение аптско-альбских эпох раннего мела.
Палеобиология
В 1999 году палеонтолог Риз Э. Баррик и геолог Уильям Дж. Шуерс обнаружили, что кости гигантозавра и тираннозавра имеют очень похожие структуры изотопов кислорода с одинаковым распределением тепла в организме. Эти терморегуляторные паттерны указывают на то, что у этих динозавров был промежуточный метаболизм между млекопитающими и рептилиями, и поэтому они были гомеотермическими (со стабильной внутренней температурой тела, типом «теплокровности»). Метаболизм 8-тонного Гиганотозавра был бы сопоставим с метаболизмом у 1-тонного хищника млекопитающих и поддержал бы быстрый рост.
В 2001 году физик Рудемар Эрнесто Бланко и Маззетта оценили поверхностную (бегущую) способность Гиганотозавра. Они отвергли гипотезу Джеймса Орвилла Фарлоу о том, что риск травм, связанных с падением таких крупных животных во время бега, ограничит скорость крупных теропод. Вместо этого они представляли, что дисбаланс, вызванный увеличением скорости, будет ограничивающим фактором. Подсчитав время, которое потребуется ноге для восстановления равновесия после отвода противоположной ноги, они обнаружили, что верхний кинематический предел скорости движения составляет 14 метров в секунду (50 км / ч; 31 миль / ч). Они также обнаружили, что сравнение бегущей способности гиганотозавра и птиц, таких как страус, по прочности их ног имеет ограниченную ценность, поскольку теропод, в отличие от птиц, имели тяжелые хвосты, чтобы уравновесить их вес.


Питание
В 2002 году Кориа и Керри обнаружили, что различные особенности задней части черепа (такие как передний уклон затылка и низкий и широкий затылочный мыщелок) указывают на то, что у гиганотозавра была бы хорошая способность перемещать череп вбок относительно передние шейные позвонки. Эти особенности также могут быть связаны с увеличением массы и длины мышц челюсти; артикуляция челюсти у гиганотозавра и других кархародонтозарид была перемещена назад, чтобы увеличить длину мускулатуры челюсти, позволяя быстрее закрывать челюсти, тогда как тираннозавры увеличивали массу мускулатуры нижней челюсти, чтобы увеличить силу их прикуса.
В 2005 году Терриен и его коллеги оценили относительную силу укуса теропод (оценки в абсолютных значениях, таких как ньютоны, были невозможны) и обнаружили, что Гиганотозавр и родственные таксоны имели приспособления для захвата и уничтожения добычи, доставляя мощные укусы, тогда как тираннозавры имели приспособления для сопротивления скручивающее напряжение и разрушение костей. Сила укуса у Гиганотозавра была слабее, чем у Тираннозавра, и сила уменьшалась назад вдоль зубного ряда. Нижние челюсти были приспособлены для разрезания укусов, и, вероятно, они захватывали и манипулировали добычей с передней частью челюстей. Эти авторы предположили, что Гиганотозавр и другие аллозавры возможно, были генерализованными хищниками, которые питались широким спектром добычи, меньшей их самих, такими как ювенильные зауроподы. Вентральный отросток (или «подбородок») нижней челюсти, возможно, был приспособлением к сопротивлению растягивающему напряжению, когда мощный укус был доставлен передней частью челюсти к жертве.
Первые известные окаменелости близкородственного Мапузавра были обнаружены в костном ложе, состоящем из нескольких особей на разных стадиях роста. В своем описании рода в 2006 году Кориа и Керри предположили, что, хотя это может быть связано с долгосрочным или случайным накоплением тушек, наличие разных стадий роста одного и того же таксона указывает на то, что агрегация не была случайной. В статье, опубликованной в 2006 году в National Geographic, Кориа заявил, что костяное ложе было, вероятно, результатом катастрофического события, и что присутствие животных в основном среднего размера, с очень небольшим количеством молодых или пожилых, является нормальным для животных, которые формируют стайки. Поэтому, по словам Кориа, крупные теропод могли охотиться группами, что было бы выгодно при охоте на гигантских зауропод.
Палеоэкология
Гиганотозавр был обнаружен в формации Candeleros, которая была отложена во время раннего сеноманского периода позднего мелового периода, приблизительно от 98 до 97 миллионов лет назад, хотя Гольц предоставил более раннюю нижнюю границу для своего возраста 99,6 миллионов лет. Эта формация является самой низкой единицей в группе Неукен, где она входит в подгруппу Рио Лимай. Формация состоит из грубых и среднезернистых песчаников, отложенных в речной среде (связанной с реками и ручьями) и в эоловых условиях (под воздействием ветра). Присутствуют палеозоли (погребенная почва), алевролиты и глиняные камни, некоторые из которых представляют болотные условия.
Гиганотозавр был, вероятно, главным хищником в своей экосистеме. Он разделял свою среду обитания с растительноядными динозаврами, такими как титанозавровый зауропод Андозавр и Реббахизавр зауропод Limaysaurus и Nopcsaspondylus. Другими тероподами являются абелизаврид экриксинатозавр, дромеозаврид буитрераптор и альваресзварид алнашетри. Другие рептилии включают крокодилообразных Арарипезух, змей и черепах Prochelidella. К другим позвоночным относятся млекопитающие-кладерийцы, пипоидные лягушки и кератодонтообразные рыбы. Следы указывают на присутствие крупных орнитопод и птерозавров.

Их есть у нас! Красивая карта, целых три уровня и много жителей, которых надо осчастливить быстрым интернетом. Для этого придется немножко подумать, но оно того стоит: ведь тем, кто дойдет до конца, выдадим красивую награду в профиль!

