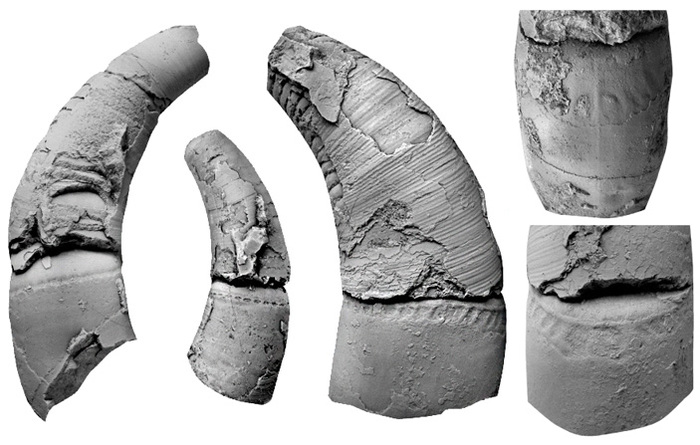
Рис. 1. Жаберные щели у хрящевых рыб (китовой акулы — а и манты — б), у современного представителя бесчелюстных — речной миноги (в; справа — схема фронтального среза миноги, на котором видны энтодермальные жаберные мешки) и жабры у низших хордовых: ланцетника (г), асцидии (д), бочоночника (е). Фото с сайтов: funniestmemes.com, discoverymaldives.com, tumblr.com, faculty.baruch.cuny.edu
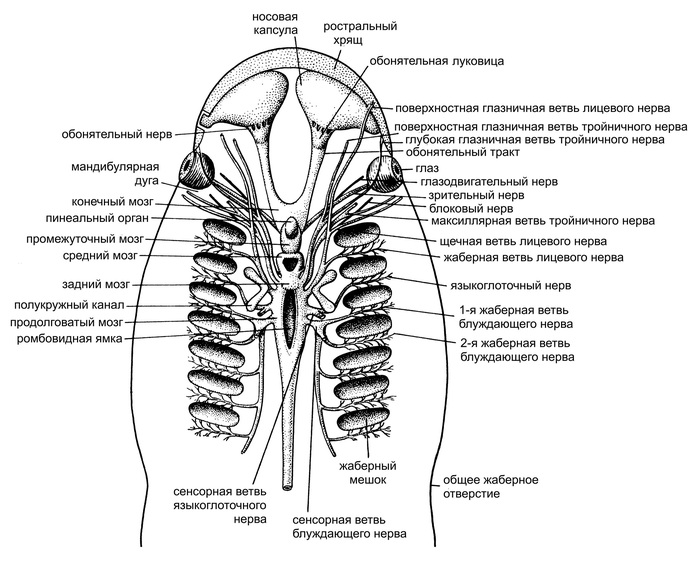
Все хорошо представляют, как выглядят у костных рыб (например, у щуки или окуня) жабры — они скрыты костяными крышками. У хрящевых рыб (акул и скатов) жаберные щели видны снаружи, они напрямую соединяют передний отдел кишечника с внешней средой (рис. 1, а, б). У более примитивных бесчелюстных позвоночных (например, у миног) это жаберные мешки — метамерные выросты кишечника, открывающиеся во внешнюю среду серией округлых отверстий по бокам тела (рис. 1, в).
Жаберные щели есть не только у позвоночных, но и у других подтипов типа хордовых. У ланцетников (небольших донных животных, похожих на маленьких рыбок, которые входят в подтип бесчерепных) жаберные щели представляют собой серию из более чем сотни метамерных отверстий, проникающих в кишечную трубку в передней половине тела (рис. 1, г). Есть жаберные щели и у еще одного подтипа хордовых животных — оболочников, которые так называются потому, что их тело заключено в оболочку из особого материала туницина, похожего на целлюлозу. Среди оболочников есть сидячие организмы — асцидии (рис. 1, д), а также плавающие в толще воды животные — бочоночники, сальпы, огнетелки и аппендикулярии (рис. 1, е). Все ископаемые хордовые тоже имели жаберные щели (рис. 2).
Рис. 2. Ископаемые хордовые с жаберными щелями: а — Didazoon haoae (ранний кембрий), б — Haikouichthys ercaicunensis (ранний кембрий), в — Metaspriggina walcotti (средний кембрий), г — Astraspis desiderata (поздний ордовик), д — конодонты (ордовик-девон), е — Jamoytius (ордовик-силур). Фото с сайтов:, anthropology-news.org, tumblr.com, dinopedia.wikia.com, pvsm.ru
У хордовых животных жаберные щели служат не только для дыхания, но и для фильтрации, что позволяет извлекать из воды мелкие пищевые объекты. Так питаются не только оболочники и ланцетники, но и многие рыбы, в том числе и самая крупная из них — китовая акула (см. рис. 1, а).
Эволюция жаберных щелей представляет собой одну из самых увлекательных глав сравнительной анатомии позвоночных животных. По мнению К. Гегенбауэра, А. Н. Северцова, И. И. Шмальгаузена и других выдающихся сравнительных анатомов XIX–XX вв., с жаберными щелями связано развитие важнейших структур в челюстном аппарате, органах чувств и кровеносной системе. Но несмотря на это, до сих пор в зоологии не существует внятной гипотезы, объясняющей, как возникли столь таинственные, своеобразные органы — метамерные жаберные щели, напрямую соединяющие полость энтодермального кишечника с внешней средой.
Эволюция жаберных щелей у позвоночных
Рис. 3. Эволюция жаберных дуг у позвоночных животных: желтым показаны губные хрящи, красным — челюстная дуга, фиолетовым — подчелюстная дуга и ее производные, голубым — жаберные дуги задних жаберных щелей и подчелюстной аппарат ([1; 2], с изм.)
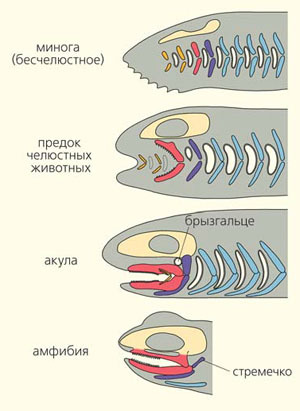
У позвоночных жаберные щели имеют хрящевой или костный скелет — жаберные дуги (рис. 3). Одна из передних дуг у большинства позвоночных изменила свою функцию — превратилась в челюсти, благодаря которым все современные челюстноротые позвоночные (кроме миног и миксин) имеют возможность хватать, откусывать и пережевывать добычу. За челюстной жаберной дугой следует так называемая подчелюстная дуга. Между двумя этими дугами у хрящевых рыб (например, акулы) даже сохраняются рудиментарные жаберные щели — брызгальца. У большинства рыб подчелюстная дуга обеспечивает подвижную связь челюстей с черепной коробкой. Челюсти как бы подвешены снизу к черепной коробке с помощью парных «подвесок» (по-латыни эти хрящи или косточки называются hyomandibulare).
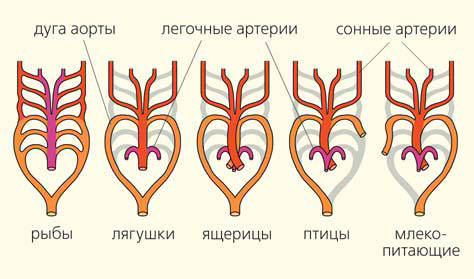
У позвоночных жаберные щели и кровеносные дуги связаны между собой (рис. 4), ведь жабры — основной орган газообмена: к ним притекает бедная кислородом кровь, в них она обогащается кислородом и далее несет его ко всем органам животного.
Рис. 4. Эволюция жаберных кровеносных дуг у водных и наземных позвоночных ([2], с изм.)
У наземных позвоночных во взрослом состоянии жаберных щелей нет, но элементы жаберного скелета, естественно, сохраняются. У таких животных (амфибий) верхняя челюсть прирастает к черепной коробке. «Подвески» освобождаются от функции прикрепления челюстей к черепу и приобретают новую функцию: они превращаются в слуховые косточки — стремечко, а полость брызгальца становится полостью среднего уха. Скелет последующих жаберных щелей превращается в подъязычный аппарат, щитовидные хрящи, хрящи гортани и трахеи. Жаберные кровеносные дуги водных позвоночных преобразуются у наземных в главные сосуды кровеносной системы — сонные артерии, дуги аорты и легочные артерии (рис. 4).
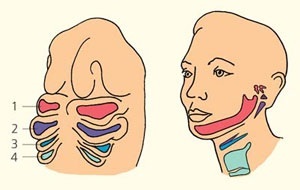
Даже в развитии человеческого зародыша закладываются жаберные щели. Хрящевые зачатки жаберных дуг дают важнейшие элементы скелета человека — челюсти, слуховые косточки, шиловидный отросток височной кости и хрящи гортани. Любой дефект в развитии жаберных щелей приводит к необратимым нарушениям и гибели зародыша человека (рис. 5). Так что можно сказать, что без них и человек не был бы человеком.
Рис. 5. Преобразование жаберных дуг зародыша человека; цифрами обозначены номера жаберных дуг ([3], с изм.)
Все изложенное показывает, как много элементов строения тела высших позвоночных, включая человека, происходит от жаберных щелей. Между тем само их происхождение остается неразгаданной тайной эволюционной морфологии животных. В самом деле, как можно представить себе происхождение многочисленных дырок, связывающих кишку с внешней средой? По какой причине могли возникнуть такие дырки? Какие органы могли дать начало жаберным щелям? Какова их первичная функция? На эти вопросы не только нет ответа, но они, по существу, даже не ставятся — по-видимому, мы слишком привыкли к тому, что жаберные щели существуют как характерный признак типа хордовых, и нечего об этом спрашивать. Чтобы попытаться ответить на вопрос о происхождении жаберных щелей, придется выйти за пределы типа хордовых.
Жаберные щели вторичноротых животных
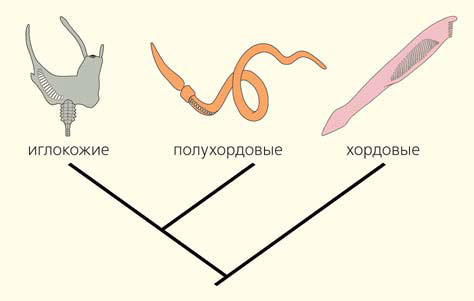
Хордовые входят в состав вторичноротых животных — одной из главных филогенетических ветвей билатерально симметричных животных, которая очень рано обособилась от общего корня билатерий [4]. Два других типа в составе вторичноротых — полухордовые и иглокожие. Согласно современным представлениям, основанным на данных молекулярной филогенетики, иглокожие и полухордовые — это родственные типы, образующие кладу Ambulacralia. Заметим, что впервые на филогенетическую близость иглокожих и полухордовых указал выдающийся российский биолог И. И. Мечников [5], который и ввел в научный обиход название Ambulacralia.
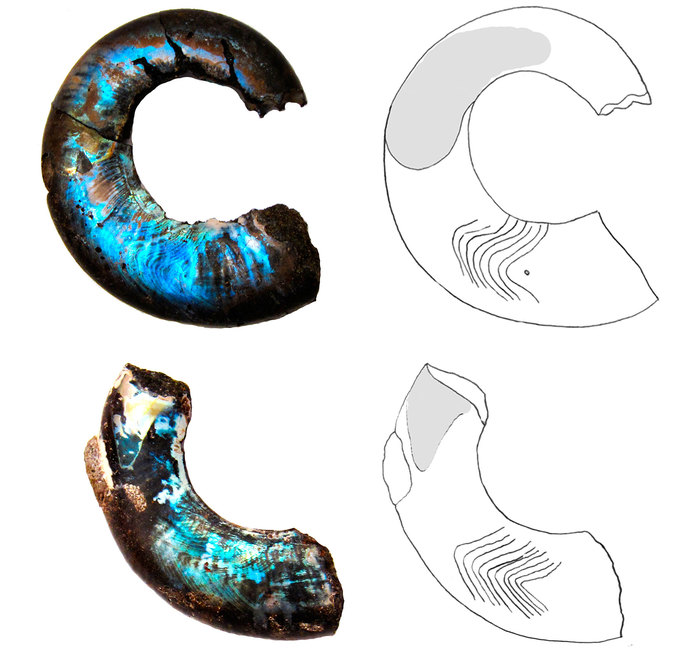
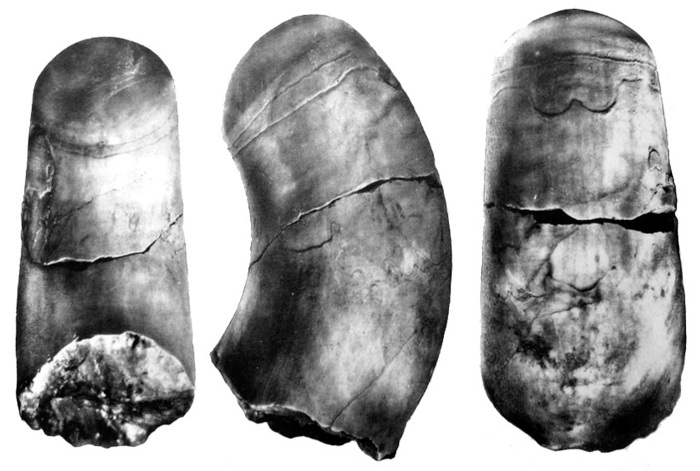
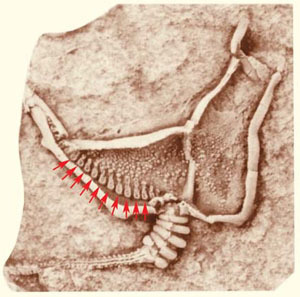
Рис. 6. Ископаемое иглокожее Cothurnocystis elizae (Calcichordata) [6]. Стрелками показаны метамерные жаберные щели
Иглокожие широко распространены в морях и океанах планеты. Это морские звезды, морские ежи, офиуры-змеехвостки, голотурии и морские лилии. Ни у кого из современных иглокожих жаберных щелей нет. Однако, у этих животных есть известковый скелет, был он и у их предков, поэтому палеонтологическая история иглокожих хорошо известна. Оказывается, самые древние кембрийские иглокожие, выделяемые в подтип Calcichordata, или Carpozoa, обладали серией отверстий [6], которые рассматриваются как гомологи жаберных щелей позвоночных (рис. 6).
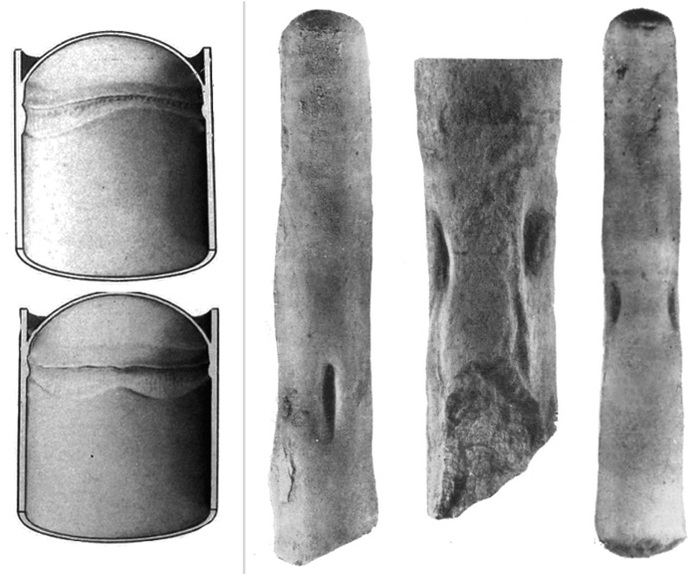
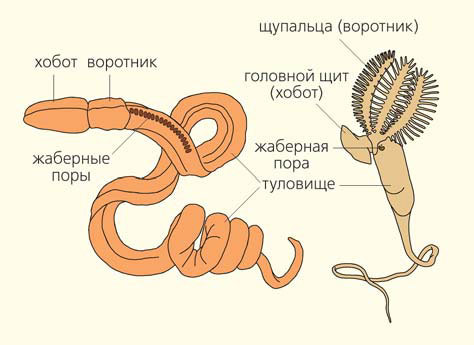
Полухордовые — тоже морские животные. В состав этого типа входят две группы, очень непохожие друг на друга, — кишечнодышащие и крыложаберные. Кишечнодышащие — довольно крупные морские черви, обитающие в толще грунта. На пляжах Бразилии в приливно-отливной полосе обитает Balanoglossus gigas, который достигает в длину 2 м. Исследования последних лет показали, что на больших глубинах (более 2 км) Мирового океана скрывается богатая и разнообразная фауна кишечнодышащих [7, 8]. Тело взрослых особей подразделяется на три отдела: мускулистый хобот, воротник, где располагается рот, и длинное туловище, в передней части которого открываются многочисленные (до 200 пар) метамерные жаберные щели (рис. 7). Крыложаберные — это мелкие колониальные организмы. На воротниковом отделе они несут щупальца, с помощью которых собирают пищу — мелкую органическую взвесь. Крыложаберные тоже имеют жаберные щели, но их немного — одна или две пары (см. рис. 7).
Рис. 7. Строение кишечнодышащих (слева) и крыложаберных животных
Таким образом, жаберные щели есть у животных всех трех типов, входящих в состав вторичноротых (рис. 8). Это означает, что, скорее всего, жаберные щели были унаследованы от общего предка вторичноротых животных. Биологи называют такие унаследованные от общего предка признаки синапоморфиями. Мы можем с полным основанием утверждать, что жаберные щели — это важнейшая синапоморфия вторичноротых животных.
Рис. 8. Три типа вторичноротых животных. В каждом типе есть представители с многочисленными жаберными щелями
Происхождение жаберных щелей
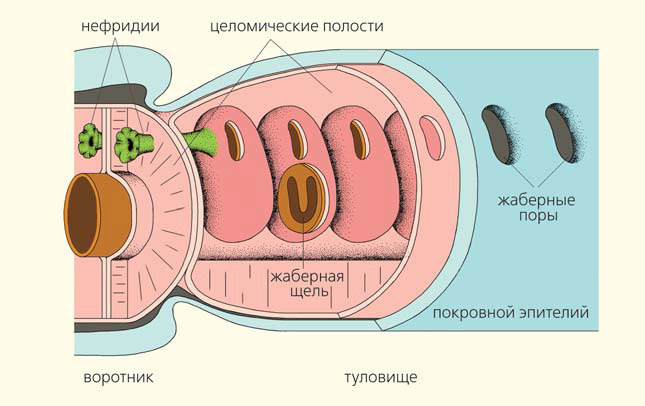
Чтобы попытаться понять происхождение жаберных щелей, надо разобраться в том, как они устроены у низших вторичноротых животных, а именно у полухордовых. Рассмотрим организацию жаберного аппарата на примере типичных кишечнодышащих. В передней части туловищного отдела кишечник с двух сторон пронизан жаберными щелями, которые имеют подковообразную форму. Они открываются не прямо во внешнюю среду, а в жаберные мешки, которые представляют собой выросты кишечника. А уже жаберные мешки соединяются с внешней средой метамерными порами, располагающимися по бокам туловища (рис. 9).
Рис. 9. Устройство жаберного аппарата кишечнодышащих (по [9], с изм.). Второй жаберный мешок частично вскрыт, и видна жаберная щель кишечника
Первая пара жаберных мешков имеет особое строение. В них открывается первая пара жаберных щелей, и одновременно в них же открываются выделительные органы (нефридии), выводящие продукты обмена из воротникового отдела. Таким образом, первая пара жаберных мешков выполняет двойную функцию — дыхательную и выделительную. Такое слияние нефридиев и первых жаберных щелей у кишечнодышащих позволило нам предложить гипотезу происхождения метамерных жаберных щелей от метамерных выделительных органов, первоначально располагавшихся в каждом сегменте тела предков вторичноротых [10].
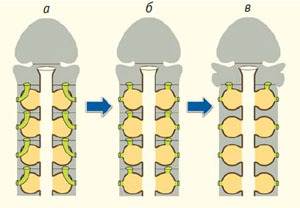
Рис. 10. Гипотеза происхождения жаберных щелей в результате слияния метамерных выделительных органов с метамерными карманами кишечника. Пояснения в тексте
Согласно нашей гипотезе, у предков вторичноротых каналы выделительных органов соединились с кишечными карманами (рис. 10, б). В результате в каждом сегменте сформировалась пара сложных органов, в состав которых вошли карманы кишечника и выделительные органы. Каждый такой орган имел три отверстия: воронку выделительного органа, щель, ведущую в кишечник, и отверстие во внешнюю среду (рис. 10, б). Такое соединение оказывается выгодным для животных, ведущих роющий образ жизни (как современные кишечнодышащие) или обитающих в трубках (как современные крыложаберные). Когда животное закапывается в толщу осадка, продукты обмена выводятся через кишечник, а когда высовывается из грунта — через наружные отверстия в воду.
Заметим, что у большинства роющих животных происходит редукция перегородок между сегментами. Причины этого кроются в том, что роющие червеобразные беспозвоночные, как правило, используют для передвижения в толще грунта гидравлический способ локомоции, при этом полостная жидкость перекачивается вдоль оси животного. Перегородки между сегментами препятствуют этому и потому редуцируются, нередко вместе с поддерживаемыми ими нефридиями.
У современных кишечнодышащих только первая пара жаберных щелей соответствует по своей организации тому сложному органу, который имелся у предков вторичноротых во всех сегментах (рис. 9, 10, в). Здесь имеется все, что нужно: воронки выделительных органов, открывающиеся в предыдущий сегмент (воротниковый отдел), дивертикул кишечника (первый жаберный мешок) и отверстие во внешнюю среду. Кишечные дивертикулы последующих сегментов утратили воронки выделительных органов, сохранив только два отверстия — в кишечник и во внешнюю среду (рис. 10, в).
Появление отверстий, которые соединяют дивертикулы кишечника с внешней средой, позволило гипотетическим сложным органам осуществлять две функции — и выделительную, и дыхательную. Это вполне очевидно для первой пары жаберных мешков современных кишечнодышащих. Однако, как оказалось, у этих животных стенки других жаберных мешков тоже сохраняют выделительную функцию. Со стороны целома они окружены специализированными выделительными клетками — подоцитами, которые обеспечивают ультрафильтрацию из целома в полость жаберных мешков [11].
Наша гипотеза позволяет проследить связь специфических органов вторичноротых животных — жаберных щелей — с метамерными органами, имеющимися у других билатерально симметричных животных, а именно с метамерными целомодуктами и метамерными дивертикулами кишечника. Гипотеза объясняет энтодермальную природу жаберных мешков и у полухордовых, и у низших хордовых вплоть до современных бесчелюстных — миног. Вряд ли можно считать случайным и то, что жаберные мешки и жаберные щели сохраняют выделительную функцию — и не только у полухордовых, но и у позвоночных животных. Хорошо известно, что жаберные мешки миног и жабры рыб выполняют не только дыхательную, но и экскреторную функцию [12].
В заключение мы считаем нужным отметить, что предлагаемая гипотеза, как бы авторам этого ни хотелось, не является абсолютно новой. Очень близкие взгляды высказывал еще в 1875 г. выдающийся немецкий биолог, основатель всемирно известной Неаполитанской зоологической станции, Антон Дорн. Он предполагал, что у метамерного предка хордовых животных произошло слияние метамерных нефридиев и метамерных выростов кишечника, которые соединились с внешней средой и дали начало жаберным щелям: «Если мы теперь представим себе, что в различных точках внутренние отверстия сегментальных органов сольются со стенкой кишечника, то установится постулированная нами связь кишечника с внешней средой, помимо ротового и заднепроходного отверстия. <...> Мы уже предположили выше, что между сегментальными органами и кишкой установилась связь, — возможно, что связь эта была вызвана или по крайней мере поддержана образованием выростов кишечника» [13]. Современники не обратили внимания на интересную гипотезу Дорна, вероятно, потому, что ее автор высказал эту идею, так сказать, походя, при обсуждении других вопросов, и не снабдил ее иллюстрациями. Гипотеза Дорна была полностью забыта почти на полтора столетия. Но «время — честный человек», как сказал Пьер Огюстен Бомарше в своей бессмертной «Женитьбе Фигаро», и время, как мы видим, подарило гипотезе Дорна вторую жизнь в XXI в.
Работа выполнена при поддержке Российского научного фонда. Проект 14-50-00034.
Об авторах
Владимир Васильевич Малахов («Природа» №7, 2016)
Владимир Васильевич Малахов — член-корреспондент РАН, заведующий кафедрой зоологии беспозвоночных биологического факультета МГУ им. М. В. Ломоносова, руководитель лаборатории биологии морских беспозвоночных Дальневосточного федерального университета (Владивосток). Область научных интересов — сравнительная анатомия, эмбриология и филогения беспозвоночных.
Ольга Владимировна Ежова («Природа» №7, 2016)
Ольга Владимировна Ежова — кандидат биологических наук, доцент той же кафедры, сотрудник той же лаборатории. Специалист в области морфологии и тонкой организации кишечнодышащих, морских звезд, офиур и голотурий; занимается изучением эволюции амбулакралий, морфологии, микроскопической анатомии и ультраструктуры полухордовых, иглокожих и хордовых.
Литература
1. Северцов А. Н. Морфологические закономерности эволюции. М.; Л., 1939.
2. Шмальгаузен И. И. Основы сравнительной анатомии позвоночных животных. М., 1947.
3. Shubin N. Your Inner Fish: A Journey into the 3,5-Billion-Year History of the Human Body. Pantheon. 2008. [Шубин Н. Внутренняя рыба. История человеческого тела с древнейших времен до наших дней / Пер. с англ. П. Петрова. М., 2010.]
4. Малахов В. В. Революция в зоологии: новая система билатерий // Природа. 2009. № 3. С. 40–54.
5. Metschnikoff E. E. Über die Systematische Stellung von Balanoglossus // Zool. Anziger. 1869. Bd. 4. S. 139–143, 153–157.
6. Jefferies R. P. S., Brown N. A., Daley P. E. J. The Early Phylogeny of Chordates and Echinoderms and the Origin of Chordate Left-Right Asymmetry and Bilateral Symmetry // Acta Zool. (Stockh.). 1996. V. 77. № 2. Р. 101–122.
7. Holland N. D., Clague D. A., Gordon D. P. et al. ‘Lophenteropneust’ hypothesis refuted by collection and photos of new deep-sea hemichordates // Nature. 2005. V. 434. P. 374–376. DOI: 10.1038/nature03382.
8. Priede I. G., Osborn K. J., Gebruk A. V. et al. Observations on torquaratorid acorn worms (Hemichordata, Enteropneusta) from the North Atlantic with descriptions of a new genus and three new species // Invertebrate Biology. 2012. V. 131. № 3. Р. 244–257. DOI: 10.1111/j.1744-7410.2012.00266.x.
9. Balser E. J., Ruppert E. E. Structure, ultrastrusture, and function of the preoral heart-kidney in Saccoglossus kowalevskii (Hemichordata, Enteropneusta) including new data on the stomochord // Acta Zool. (Stockh.). 1990. V. 71. P. 235–249.
10. Ezhova O. V., Malakhov V. V. The nephridial hypothesis of the gill slit origin // J. Exp. Zool: (Mol. Dev. Evol.). 2015. V. 324B. Р. 647–652. DOI: 10.1002/jez.b.22645.
11. Pardos F., Benito J. Fine structure of the branchial sacs of Glossobalanus minutus (Enteropneusta) with special reference to podocytes // Arch. Biol. 1988. № 99. Р. 351–363.
12. Наумов Н. П., Карташев Н. Н. Зоология позвоночных. Часть 1: Низшие хордовые, бесчелюстные, рыбы, земноводные. М., 1979.
13. Dohrn A. Der ursprung der wirbelthiere und das princip des functionswechsels. Genealogische skizzen. 1875. Leipzig: Verlag von Wilhelm Engelmann. Р. 10–17. [Дорн А. Происхождение позвоночных животных и принцип смены функции / Пер с нем. Б. И. Балинского. М.; Л., 1937. С. 104–105, 112.]