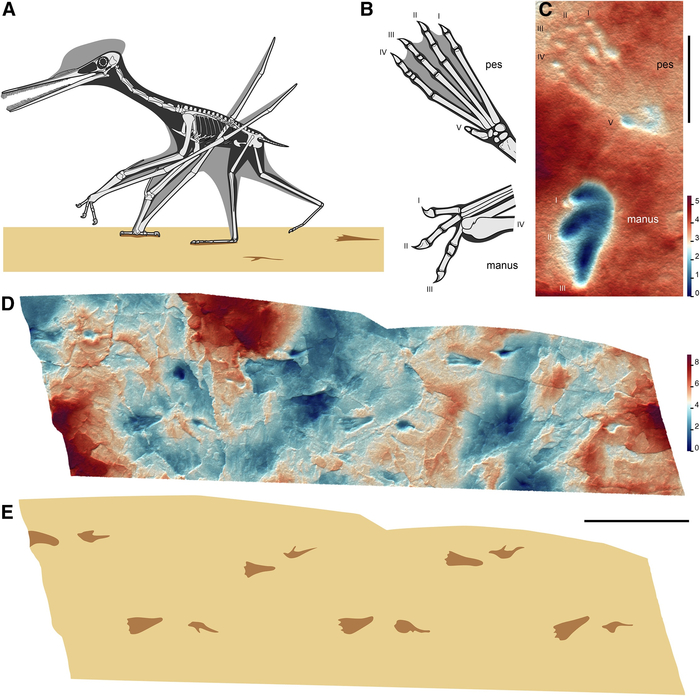
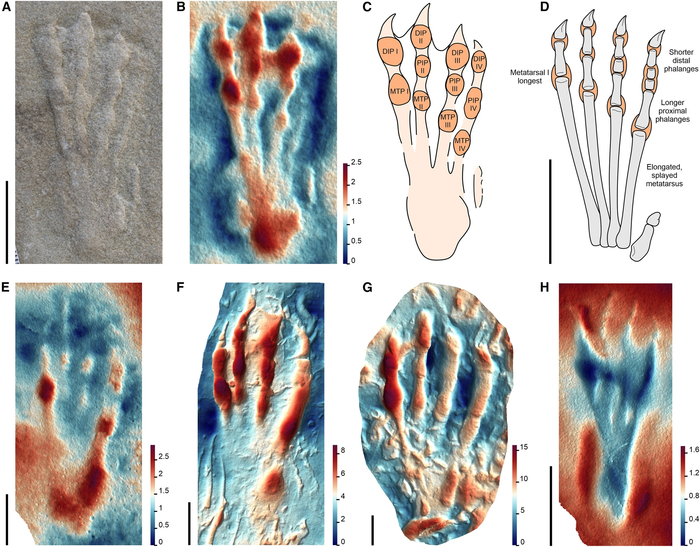
Птерозавров часто представляют парящими над головами динозавров, но новый анализ их окаменелых следов показывает, что некоторые из этих летающих рептилий чувствовали себя столь же комфортно, гуляя по земле.
Когда мы представляем экосистемы мезозоя, птерозавры неизменно возникают в воображении как величественные летуны, парящие над древними ландшафтами. Однако новое исследование несколько меняет этот стереотип: оказывается, многие из этих крылатых рептилий чувствовали себя на земле так же уверенно, как и в воздухе. Ученые обнаружили, что уже 160 миллионов лет назад несколько групп птерозавров освоили наземный образ жизни, о чем красноречиво свидетельствуют их окаменелые следы.
Ископаемые следы птерозавров стали ключом к разгадке их повседневной жизни. «Они показывают не только где жили эти существа, но и как они передвигались, охотились и взаимодействовали со средой», — объясняет палеонтолог Роберт Смит, ведущий автор исследования. Ученые выделили три типа следов, каждый из которых соответствует определенной группе птерозавров, раскрывая их уникальные экологические ниши.
Одной из самых удивительных находок стали следы неоаждархий (Neoazhdarchia ) — группы, включавшей кетцалькоатля, гиганта с размахом крыльев до 10 метров. Ранее считалось, что такие гиганты редко спускались на землю из-за их размеров. Однако следы, обнаруженные в прибрежных и внутренних регионах, доказывают обратное: эти рептилии активно передвигались по суше, возможно, конкурируя за ресурсы с динозаврами.
Другая группа, ктенохазматоиды (Ctenochasmatoidea), оставила следы в илистых прибрежных отложениях. Эти птерозавры с игольчатыми зубами и длинными челюстями, вероятно, «патрулировали» мелководные лагуны, ловя рыбу и беспозвоночных. Интересно, что их передние конечности при ходьбе складывали четвертый палец, поддерживающий крыло, превращая «руки» в функциональные трехпалые лапы. Такая адаптация позволяла им эффективно перемещаться по мягкому грунту, а обилие следов говорит о том, что прибрежные зоны были для них критически важными местообитаниями.
Третий тип следов связан с джунгариптеридами (Dsungaripteridae) — птерозаврами, чьи мощные челюсти были идеальны для дробления раковин. Их следы часто находят рядом с окаменелостями скелетов, что позволило точно установить связь между отпечатками и анатомией. Задние зубы этих рептилий, напоминающие дробилки, и беззубые изогнутые клювы указывают на специализацию в питании моллюсками, что подтверждается расположением следов в районах древних побережий.
Исследование также проливает свет на эволюцию птерозавров. Ранние виды, судя по отсутствию следов до средней юры, вели древесный образ жизни, цепляясь за ветви. Однако 160 миллионов лет назад произошел перелом: птеродактилоиды — продвинутая группа — начали осваивать наземные экосистемы. Это совпало с развитием стопоходящих конечностей (когда ступня полностью касается земли) и ипсилатеральной походки, при которой конечности с одной стороны движутся синхронно, как у современных ящериц.
В этой публикации я хочу рассказать о двух семействах меловых аммонитов - Deshayesitidae (Stoyanow) и Parahoplitidae (Spath). На нашем сайте «Аммонит.ру» представители этих семейств очень хорошо известны благодаря находкам с Кавказа, в первую очередь из Дагестана.
Семейства Deshayesitidae и Parahoplitidae жили в аптском веке раннего мела. Общее время их существования превышало 10 миллионов лет, но жили они не одновременно – первая половина аптского века была временем Deshayesitidae, затем их очень быстро, почти мгновенно по геологическим меркам, сменили Parahoplitidae, существовавшие до конца аптского века (и даже в самом начале следующего альбского века).
Но объединяет эти семейства не только время их существования. Оба они произошли от гетероморфных предков! Представители этих семейств, кроме самых первых Deshayesitidae, относящихся к роду Turkmeniceras, были вполне нормальными спирально-свернутыми аммонитами. Однако, строение их лопастной линии свидетельствует о том, что в их эволюции была гетероморфная стадия. Интересно, что одновременно с дегезитами и парагоплитами существовало еще одно семейство - Douvilleiceratidae (Parona et Воnarelli), также произошедшее от гетероморфов.
Подобная эволюция от гетероморфов к мономорфам долго время казалась палеонтологам невозможной. Однако, изучение лопастной линии и находки промежуточных форм между гетероморфными аммонитами и ранними Deshayesitidae и Douvilleiceratidae показали, что подобная ситуация была вполне реальной и даже повторялась в нескольких эволюционных линиях аммонитов. То есть, аммониты, становившиеся гетероморфными (переходившими к относительно пассивному образу жизни), не лишались навсегда тех органов и особенностей строения, которые были необходимы для активного плавания и вполне могли вернуться к типичному мономорфному плану строения, если им это было нужно. Гетероморфность вовсе не было каким-то редким тупиковым путем эволюции, «уродством», за которым неизбежно следовало вымирание. Гетероморфные аммониты вполне могли снова стать мономорфными.
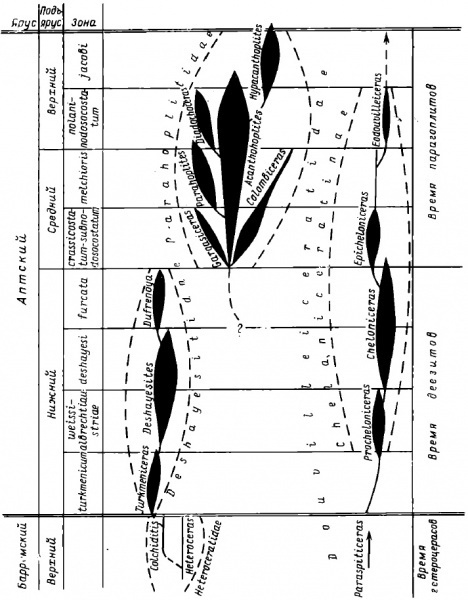
Барремский век, предшествующий аптскому, был временем расцвета гетероморфных аммонитов. Среди них были и «полугетероморфы», приобретавшие раскрученную раковину только на взрослой стадии развития, и аммониты, бывшие гетероморфными всю жизнь, с раннего детства. Но в конце баррема и в самом начале апта что-то изменилось в морях и океанах, и многие аммониты были вынуждены вернуться к классическому плану строения раковин. Большинство гетероморфных аммонитов исчезли в раннем апте, в самом начале этого века, однако, не все они вымерли, просто их потомки перестали быть гетероморфами. Родственные связи Deshayesitidae и Parahoplitidae точно не ясны. С этим связано и разночтение в выделении надсемейств – большинство российских палеонтологов выделяет для каждого семейства свое надсемейство - Deshayesitaceae и Parahoplitaceae соответственно. На Западе чаще объединяют их в надсемействе Deshayesitaceae, предполагая близкое родство этих семейств.
Deshayesitidae
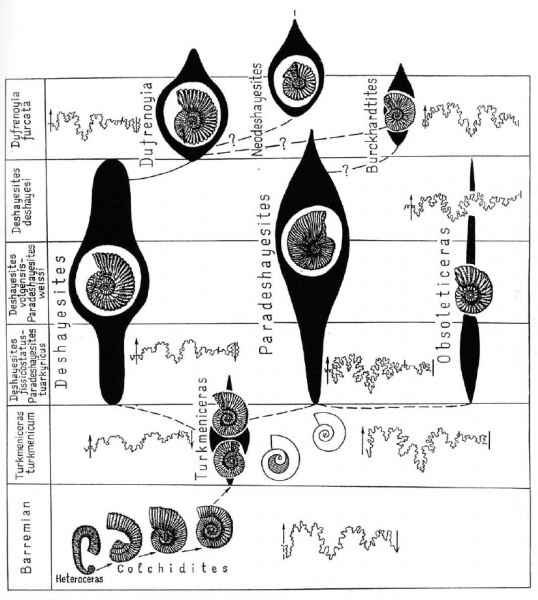
Происхождение семейства Deshayesitidae изучено очень хорошо. Российские палеонтологи Т.Н. Богданова и И.А. Михайлова составили детальную схему ранней эволюции этого семейства, которая даже вошла в знаменитый учебник палеонтологии Михайловой и Бондаренко.
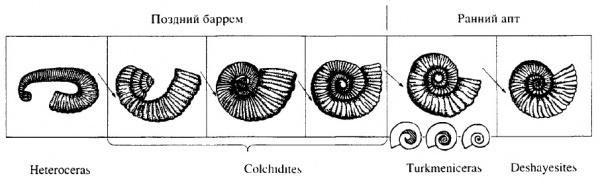
Схема происхождения Deshayesitidae от гетероморфных предков 1999 года выпуска из статьи Богданова Т.Н., Михайлова И.А. 1999 Происхождение и развитие семейства Deshayesitidae
На схеме показано происхождение от гетероморфного рода Heteroceras через Colchidites рода Turkmeniceras, который являлся непосредственным предком собственно рода Deshayesites (вслух читается как «дегезитес» или «дегезит», но писать название лучше всегда на латыни). Первым представителем семейства Deshayesitidae считается Turkmeniceras, который уже имел спирально свернутую раковину с соприкасающимися оборотами на взрослой стадии развития и лишь в детстве был гетероморфным. У него сохранялось отверстие в центральной части раковины – «пупочное зияние», быстро исчезнувшее в ходе эволюции.
Схема семейства Deshayesitidae 2004 года выпуска из книги Bogdanova, T.N. & Mikhailova, I.A., 2004. - Origin, evolution and stratigraphic significance of the superfamily Deshayesitaceae Stoyanow, 1949
Представители рода Deshayesites – основного рода семейства, были распространены на окраине океана Тетис в Северном Полушарии, от Франции до Туркменистана. В России они известны в первую очередь из Поволжья и Кавказа. Выделено множество видов Deshayesites, однако, отличия между большинством из них не очень велики и определение их далеко не всегда представляется возможным.
В конце раннего апта от Deshayesites произошел род Dufrenoya, отличающийся плоской вентральной стороной раковины и мощными бугорками-шипами по ее краям.
Некоторые авторы выделяют в этом семействе еще несколько родов, самые крупные из которых – Paradeshayesites и их потомки Neodeshayesites. С вымиранием родов Dufrenoya и Neodeshayesites исчезло и само семейство Deshayesitidae.
Parahoplitidae
Аммониты семейства Parahoplitidae жили в среднем и позднем апте. Самыми известными и многочисленными родами этого семейства являются Acanthohoplites, Parahoplites и Hypacanthohoplites.
Прежде чем рассказывать о Parahoplitidae более подробно, следует сказать о названии этого семейства. А название это, прямо скажем, очень неудачное. Во-первых, окончание –hoplitidae в названии этих аммонитов появилось в те далекие времена, когда палеонтологи не обращали внимание на лопастную линию, не задумывались об эволюции и родственных связях аммонитов, а смотрели только на внешнее сходство. Тогда всех шипастых аммонитов записывали в космоцерасы, а всех аммонитов с килем – в амальтеусы. Тогда же и Acanthohoplites с Parahoplites записали в родственники к настоящим Hoplites. Но уже в начале ХХ века стало понятно, что эти аммониты родственниками друг другу вовсе не являются. Но устоявшиеся названия решили не менять, в результате до сих пор новички в палеонтологии путаются в этих –hoplites и иногда относят Acanthohoplites или Parahoplites к семейству Hoplitidae. Это грубая ошибка! Эти рода не то что не относятся к гоплитидам, семейства Parahoplitidae и Hoplitidae даже не являются близкими родственниками!
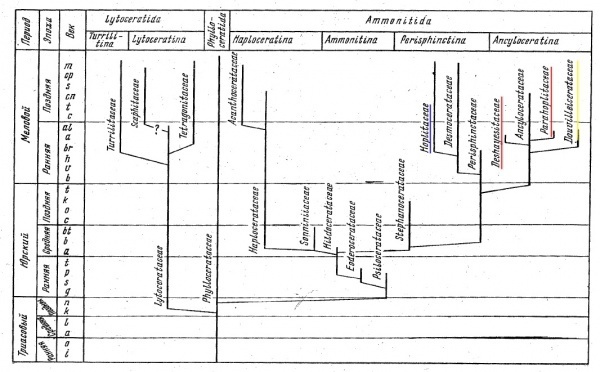
Схема из книги Безносов Н.В., Михайлова И.А. 1991 Высшие таксоны юрских и меловых Ammonitida.
На схеме красным подчеркнуты те семейства, о которых идет речь, желтым – упоминавшееся здесь Douvilleiceratidae (вообще-то на схеме указаны надсемейства, но в такой интерпретации все они включают в себя лишь по одному семейству), синим – надсемейство Hoplitaceae, которое ни в коем случае не следует путать с Parahoplitidae. Второй недостаток названия Parahoplitidae связан с тем, что род Parahoplites вовсе не был самым многочисленным или долгоживущим из родов этого семейства. Центральным, стержневым родом был Acanthohoplites, именно от него ответвлялись все остальные представители этого семейства. Так что логичнее было бы назвать семейство Acanthohoplitidae. Но увы, в свое время палеонтологи решили иначе.
В отличие от Deshayesitidae, происхождение парагоплитид покрыто мраком. Они появляются на границе нижнего и среднего апта, когда угасает род Dufrenoya, но предки их до сих пор не известны. Строение лопастной линии Parahoplitidae однозначно свидетельствует о наличии в их эволюции гетероморфной стадии, а внешний вид раковин – о родстве с Deshayesitidae. Однако, есть в их лопастной линии небольшие различия, которые, по мнению некоторых палеонтологов, опровергают предположения о родстве парагоплитид с дегезитами.
Филогенетические взаимоотношения внутри семейств Deshayesitidae, Parahoplitidae и Douvilleiceratidae из статьи И.А. Михайлова 1979 "Эволюция аптских аммоноидей"
Получается парадоксальная ситуация. Вымирают Deshayesitidae, имевшие гетероморфных предков, им на смену приходят Parahoplitidae, похожие внешне и тоже имевшие гетероморфных предков, но только вот никого из их предков никто из палеонтологов никогда не видел. Ни гетероморфных, ни мономорфных, никаких. Да и гетероморфов, способных дать начало таким аммонитам, к концу раннего апта уже не осталось.
Этому есть два вероятных объяснения. Первое – предки Parahoplitidae были крайне малочисленным видом-эндемиком, существовавшим где-то в тех далеких краях, куда еще не добрались палеонтологи. После вымирания дегезитов эти загадочные предки вышли из своего убежища и дали начало роду Acanthohoplites, который занял освободившуюся экологическую нишу, дал вспышку численности, а затем и вспышку видообразования.
Вторая версия, на мой взгляд более рациональная – все-таки, предками Parahoplitidae были какие-то дегезиты и род Acanthohoplites произошел именно от них. В этом случае нам не нужно придумывать гипотетических предков – вот они, на виду у всех. Сейчас эта версия постепенно набирает популярность, к ней склоняется все больше специалистов, так как уж очень странным кажется полное отсутствие каких-либо вероятных предков Parahoplitidae в том случае, если мы отрицаем их родство с Deshayesitidae.
Как я уже говорил, основным родом семейства Parahoplitidae был Acanthohoplites. Одновременно с ним существовали Parahoplites, Colombiceras и некоторые другие. Правда, род Colombiceras некоторые авторы относят к семейству Douvilleiceratidae, но аргументы сторонников «парагоплитовой гипотезы» выглядят вполне солидно.
Интересно, что эволюционная линия Acanthohoplites - Hypacanthohoplites напоминает линии Deshayesites - Dufrenoya и Deshayesites - Neodeshayesites – у обоих последних родов дегезид формируется плоская вентральная сторона, а Neodeshayesites так и вовсе можно спутать с Hypacanthohoplites . Именно Hypacanthohoplites оказался последним в своем семействе – история парагоплитид на этом заканчивается.
Аулакостефаниды – одни из самых интересных и в то же время мало изученных верхнеюрских аммонитов Русской платформы. Само это семейство и своей историей, и обликом некоторых своих представителей очень напоминает уже обсуждавшихся здесь космоцератид.
Аулакостефаниды – тоже настоящее суббореальное семейство и, как и остальные группы аммонитов, обитавших в экотонной зоне между двумя биогеографическими надобластями (Панбореальной и Тетис-Панталасса), существовали они недолго. Появившись в начале позднего оксфорда, аулакостефаниды полностью исчезли в конце кимериджского века. Но за полтора «века» существования в этом семействе успело появиться чуть меньше десятка родов (если считать макроконхов) и что-то около 50 видов, заметная часть которых служит видами-индексами зон, подзон и биогоризонтов суббореального и бореального оксфорда и кимериджа. Именно к этому семейству принадлежат все виды-индексы «стандартной» (английской) шкалы кимериджского яруса. Как и у большинства других позднеюрских аммонитов, у аулакостефанид был хорошо развит диморфизм. Микроконхи отличались по своей более грубой и сохраняющейся до конечной жилой камеры скульптуре, а также по более мелким размерам и присутствию ушек, а макроконхи, соответственно, в основном обладали более слабо скульптурованной раковиной с простым устьем. Диаметр раковины макроконхов мог превышать таковой микроконхов в 10 и более раз. Таксономический статус микро- и макроконхов Aulacostephanidae (опять же, как и у многих других перисфинктид) не совсем устоялся: их относили к разным родам, подродам или видам. Аулакостефаниды – первые из подотряда перисфинктин, у которых появилась лопастная линия «краспедитового» типа – с ростом раковины её усложнение шло в первую очередь за счет увеличения числа элементов, а не их сложности (как это происходит у обычных пери сфинктид).
Древнейшие аулакостефаниды (Decipia), как предполагал Дж. Кэлломон, а вслед за ним и другие аммонитчики, произошли от перисфинктид рода Liosphinctes. От своих предков Decipia отличались несколько меньшими размерами раковины, ранним появлением многоветвистых пучков ребер, меньшим углом, под которым лопастная линия подходила ко шву, а также большим количеством боковых лопастей (которые у них образовывались за счет деления внутренней боковой лопасти I, как и у остальных представителей подотряда Perisphinctina). Эти аммониты сразу же расселились по всей Бореально-Атлантической области (хотя Северной Атлантики ещё не существовало, название это широко используется) – от Восточной Гренландии до Русской платформы. Севернее и восточнее находки деципий не известны. Японские палеонтологи, правда, отнесли к Decipia несколько обломков аммонитов из Японии, но, скорее всего, эти экземпляры принадлежат к роду Dichotomosphinctes. К роду Decipia, судя по всему, принадлежит один «забытый» вид – Perisphinctes tenuifurcatus Nikitin, описанный в сборнике посмертно изданных трудов С.Н. Никитина и с тех пор никем никогда не упоминавшийся (Никитин, 1916, табл. II, фиг. 15-16). Никитин, правда, указывал этот вид из «кордатовых слоев», но эта привязка, видимо, ошибочна. Другие деципии стали известны у нас совсем недавно – статья с их описанием вышла только в прошлом году (Glowniak et al., 2010).
Потомки деципий Rinsteadia более далеко распространились на юг, их находки встречаются вместе с субтетическими аммонитами в Польше, Южной Германии и Франции; присутствуют они также в разрезах Англии, Шотландии и, конечно, Русской платформы. А вот в Гренландии рингстедии пока не найдены. Рингстедии были весьма разнообразными, их слабо скульптированные раковины были от полуэволютных до почти почти совершенно инволютных, а лопастная линия приобрела типичный «краспедитовый» облик. Если микроконхи деципий известны плохо, то с микроконхами рингстедий дело обстоит совершенно иначе. Это – небольшие хорошо скульптированные аммониты, относящиеся к роду Microbiplites. На Русской платформе рингстедии не слишком хорошо известны. Хотя их первые описания относятся к XIX веку – именно тогда Г. Траутшольд описал Ammonites cuneatus - в дальнейшем их описывали и/или изображали всего несколько раз. Микроконхи не изображались до самого последнего времени (первые изображены здесь – Glowniak et al., 2010, вместе с макроконхами), макроконхов изобразили Давиташвили (1926), Иловайский и Флоренский (1941 – под названием Ammonia kurmani). Не совсем понятно, были ли рингстедии на Приполярном Урале, откуда они изображались М.С. Месежниковым – это достаточно эволютные формы, которые могут относиться и к Pictonia. В любом случае, в Западной Сибири и восточнее этих аммонитов нет.
С границей оксфорда и кимериджа у аулакостефанид связаны важные изменения. Сама эта граница проводится по смене Ringsteadia на Pictonia, Vineta и Pomerania у макроконхов и Microbiplites на Prorasenia (=Desmosphinctes) у микроконхов. Столько родовых названий для аулакостефанид из основания кимериджа предложено не случайно – уж больно они разнообразные. Pictonia – почти эволютные, с хорошо скульптированными внутренними и гладкими внешними оборотами. Vineta, чьи раковины достигали в диаметре более полуметра - полуинволютные, с гладкой раковиной (только на ранних оборотах у этих аммонитов видны ребра). Pomerania были близки к Pictonia, но отличались появлением грубых одиночных ребер-складок на внешних оборотах. Все эти аммониты, как и оксфордские представители семейства, до последнего времени оставались у нас почти неизвестными. Пока известно только два района на Русской платформе, где пограничный интервал оксфордского и кимериджского ярусов представлен достаточно полно – это бассейн р. Унжи (Макарьев, Михаленино) и Соль-Илецкий район Оренбургской области (Ханская гора). Судя по присутствию в Москве и окрестностях Desmosphinctes, этот интервал должен присутствовать и здесь, но, насколько я могу понять, в разрезах он сейчас не обнажается. К началу кимериджа приурочено расселение аулакостефанид как на север и восток, так и на юг. В Арктике, вплоть до Таймыра и Хатангской впадины распространяются своеобразные Pictonia из группы P. involuta Mesezhnikov. В субсредиземноморских разрезах в это же время появляются «средиземноморские рингстедии», недавно выделенные А. Вежбовским в отдельный род Vielunia.
В начале фазы Cymodoce всё это разнообразие вновь свелось к одному единственному роду Rasenia. Макроконхи разений, у которых появились боковые бугорки, удивительно напоминают макроконхов нижнекелловейских космоцератид Kepplerites (Gowericeras), а вот микроконхи у них заметно различаются. Как и пиктонии, разении были широко распространены в Северном полушарии. Достигли они и Арктических островов – есть разении на Шпицбергене и на Земле Франца-Иосифа. Кстати, у микроконхов аулакостефанид бугорки в месте ветвления ребер появились ещё в самом начале кимериджа. Многие исследователи по этой причине считали Prorasenia предками Rasenia.
В конце раннего-начале позднего кимериджа разнообразие аулакостефанид вновь увеличилось. В это время и у микро- и у макроконхов появляется перерыв ребер на вентральной стороне. Сначала, видимо, такой перерыв появляется у Zonovia, которые в остальном очень похожи на разений. Потом – у эволютных, грубо скульптированных Zenostephanus и у первых, в основном слабо скульптированных Aulacostephanus (Aulacostephanoides [M], Aulacostephanites [m]). Эти аммониты несколько отличались по своему географическому распространению. Зоновии в основном характерны для довольно узкой полосы, охватывающей северную часть Суббореальной области и южную – Арктической. Аулакостефанусы были распространены более широко. Они встречались по всей Суббореальной области, заходили на север Субтетической области, а местами достигали и Арктики (Хатангской впадины). Zenostephanus [M] – Xenostephanoides [m] встречались в Суббореальной области, но наиболее широко расселились в Арктике. Это – единственные верхнекимериджские аулакостефаниды, известные на Шпицбергене и на Земле Франца-Иосифа. И это – единственные аулакостефаниды, которые встречаются в Северной Америке (на севере Британской Колумбии, в Канаде), хотя они там исключительно редки: за последние 50 лет их было найдено всего несколько экземпляров. В самом начале позднего кимериджа существовали также «южные» разении (Eurasenia), распространенные в как в Суббореальной (Бореально-Атлантической), так и в Субсредиземноморской области. У этих аммонитов (порой достигавших весьма немалых размеров – до 60 см) перерыва ребер на вентральной стороне не было. Интересно, что у нижнекимериджских микроконхов (Prorasenia) ослабление ребристости на вентральной стороне есть, но оно имеет иную природу, чем у верхнекимериджских аулакостефанид, образуясь за счет утолщение ребер при переходе на вентральную сторону (Месежников, Алексеев, 1974)
В начале фазы Eudoxus верхнего кимериджа появились достаточно грубо скульптированные аулакостефанусы, обычно рассматриваемые в составе подрода Pararasenia, но в действительности являющиеся гомеоморфами Центральноамериканских параразений (см. ниже). Их нужно относить к подроду Aulacostephanus (Sarygulia) [M] (Khudyaev in Sasonov, 1960).
Скорее всего, эти аммониты являлись потомками подрода Aulacostephanoides. Чуть позже появились настоящие Aulacostephanus (Aulacostephanus) [M] – полуинволютные, часто очень крупные и обычно не слишком сильно скульптированные аммониты, просуществовавшие до конца кимериджского века. Микроконхи аулакостефанусов из зон Eudoxus и Autissiodorensis объединятся в подрод Aulacostephanus (Aulacostephanoceras).
В самом конце кимериджа аулакостефаниды вымерли. Скорее всего, это событие произошло одновременно в Суббореальной и Субтетической областях, немного раньше аулакостефаниды исчезли в Арктике (в Хатангской впадине их уже нет в верхней зоне кимериджа, а в Восточной Гренландии – в верхней подзоне).
Кроме настоящих аулакостефанид в кимериджском веке существовали также другие аммониты, очень на них похожие. В первую очередь это Pararasenia – аммониты, описанные из слоев с Idoceras Мексики К. Бурхардтом как Aulacostephanus zakatekanus. Потом Л. Спэт (1925) сделал этот вид типовым для рода Pararasenia. Он указал, что эти аммониты очень похожи на Aulacostephanus, но у них отсутствует перерыв скульптуры на вентральной стороне, чем они напоминают Rasenia. Спэт предполагал, что этот род связывает Aulacostephanus (для которого он в этой работе предложил типовой вид A. eudoxus, d'Orbigny) и Rasenia. Кроме мексиканского вида, к параразениям Спэт отнес также европейские виды A. pseudomutabilis (de Loriol) и A. circumplicatum (Quenstedt). Однако мексиканские аммониты обладают несколько отличающейся лопастной линией и меньшими размерами по сравнению с европейскими аммонитами, а также четко изолированным ареалом. Скорее всего, они не относятся к аулакостефанидам. В кимеридже Мексике встречаются и другие очень похожие на аулакостефанид аммониты. Это известный по единственной находке аммонит, описанный А. Канту-Чапой как Pictonia (Colladites) с гладкой жилой камерой и покрытыми одиночными ребрами внутренними оборотами. Скорее всего, он близок к идоцератинам – подсемейству, широко распространенному в кимеридже Мексики. Ещё в верхнем кимеридже Мексики и Восточной Африки встречаются аммониты, очень похожие на Craspedites или небольших Ringsteadia – Procraspedites. Как было установлено Ф. Олоризом с соавторами (Oloriz et al., 1998) по присутствию характерных пережимов на внутренних оборотах эти аммониты должны относиться к семейству атаксиоцератид.
3. Номенклатурные сложности
Многим кимериджским аулакостефанидам не повезло с типовыми видами. Так, вид Ammonites cymodoce d’Orbigny стал типовым видом и Rasenia и Pictonia. А. д'Орбиньи под названием Ammonites cymodoce изобразил различных аммонитов (хотя, видимо, и принадлежащих одному виду), и в качестве типового для обоих родов был выбран экземпляр, изображенный на табл.202, фиг.3-4. В. Аркелл сначала написал статью, где среди прочего обрисовал всю непростую ситуацию (Arkell, 1935), а затем направил предложения в Международную комиссию по зоологической номенклатуре (МКЗН) – высший орган, принимающий решения в спорных случаях. В результате в 1951 году МКЗН приняла решение, зафиксировавшее типовые виды рассматриваемых родов (соответственно, Pictonia baylei и Rasenia involuta), а название cymodoce d'Orbigny, 1850 поместившее в список отклоненных названий, которые не должны в дальнейшем использоваться (Hemming, 1951). Позднее, после обсуждения проблемы с В. Дж. Аркеллом, было решено ещё раз поставить вопрос на голосование в МКЗН: по мнению Аркелла название Cymodoce, которое к этому времени уже давно стало широко используемым названием для зоны, лучше всё же было сохранить в качестве используемого, с типовым экземпляром, выбранным из коллекции д’Орбиньи в 1904 г. В результате члены комиссии большинством голосов восстановили в правах вид Cymodoce (Hemming, 1956). Ну а вместо R. involuta Spath 1935 в дальнейшем было предложено название Rasenia anglica Geyer, 1961 (т.к. вид Ammonites involutus Quenstedt угодил в род Rasenia, и название, предложенное Спэтом стало его младшим омонимом). В. Аркелл с Дж. Кэлломоном (Arkell, Callomon, 1961) считали, что это микроконх, но впоследствие Т. Биркелунд (Birkelund et al., 1978) показала, что это может быть и молодой макроконх.
Столь же непростая ситуация была и с типовым видом рода Aulacostephanus. Оно было первоначально опубликовано Торнквистом (Tornquist, 1896) без диагноза, со ссылкой на то, что название предложено Сутнером и Помпецким. Единственным видом, отнесенным к данному роду, стал Ammonites mutabulis Sowerby – однако, судя по всему, Торнквист имел в виду аммонита, изображенного под этим названием д’Орбиньи (и который в свою очередь был отнесен де Лориолем к новому виду pseudomutabilis); по правилам МКЗН этот вид и должен являться типовым для Aulacostephanus, хотя Торнквист про это ничего и не писал. Тем не менее, в 1925м году типовой вид для рода Aulacostephanus был предложен сразу двумя выдающимися аммонитчиками – Леонардом Спэтом (Spath, 1925) и Отто Шиндевольфом (Schindewolf, 1925). Спэт предложи в качестве такового A. eudoxus, а Шинденвольф – A. phorcus. Однако оба предложенных названия не были в числе видов, изначально отнесенных к роду Aulacostephanus Торнквистом, и такое обозначение типового вида не является валидным. Позднее В.Дж. Аркелл (Arkell, 1935) предложил считать типовым видом рода Aulacostephanus A. pseudomutabilis de Loriol, и это было закреплено специальным решением МКЗН в 1951 году.
Название Sarygylia было опубликовано И. Худяевым (1932) только в виде подписи к одному из рисунков, тогда как в тексте тот же самый вид был отнесен к Aulacostephanus. Такое выделение нового рода без диагноза и ссылки на типовой вид недействительно. Позднее единственным, кто использовал предложенное Худяевым название, был Н.Т. Сазонов (1960), который привел диагноз данного рода и перечислил другие относящиеся к нему виды. Сазоновым в той же работе был установлен новый род Tobolia, который оказался младшим омонимом установленного ранее рода фораминифер и заодно синонимом Aulacostephanoides.
Род Xenostephanus, установленный В. Аркеллом и Дж. Кэлломоном в 1963 году, оказался омонимом рода млекопитающих, установленных годом раньше, и для аммонитов в 2010м году было предложено новое название Zenostephanus.
Не очень понятной получилась ситуация с Ammonites uralensis d’Orbigny in Murchison et al.,1845 – типовым видом рода Zonovia. Орбиньи изобразил под этим названием двух аммонитов. Позднее Р. Зальфельд (Salfeld, 1913) посчитал, что к данному виду относится только более крупный экземпляр (Тове Биркелунд указывала, что Зальфельд таким образом выбрал лектотип – но в действительности в статье Зальфельда лишь было указано, какого из изображенных Орбиньи аммонитов он относит к данному виду). Вскоре Р. Дювие (Douville, 1911) изобразил двух аммонитов из коллекции Орбиньи, один из которых, происходящий из окрестности Кинешмы, ранее не изображался. Сам Дювийе решил, что это – нижнекелловейские Kepplerites. Более крупный из изображенных Дювийе аммонитов был впоследствии выбран в качестве лектотипа Б. Циглером (Ziegler, 1962). Поскольку более крупный аммонит с Урала, изображенный Орбиньи, был утерян, М.С. Месежников (1984) после консультаций с Дж. Кэлломон предложил одного из аммонитов из своей коллекции в качестве неотипа вида Z. uralensis – это, однако, не является валидным обозначением, поскольку лектотип был выбран Циглером корректно.
Первая статья из цикла история семейства аммонитов посвящена очень многочисленному, крупному и широко известному семейству кардиоцератид (Cardioceratidae). Это семейство аммонитов захватило в юрском периоде практически всю бореальную область Северного полушария. Их раковины находят в России, в Восточной и Западной Европе, в Канаде. Можно сказать, что кардиоцератиды определяли облик аммонитовой фауны Центральной России на протяжении большей части юрского периода. Это были бореальные аммониты – их родиной были приполярные моря. В одной из посвященных им статей автор назвал кардиоцератид «суровыми северными аммонитами», хотя что он увидел в них сурового?
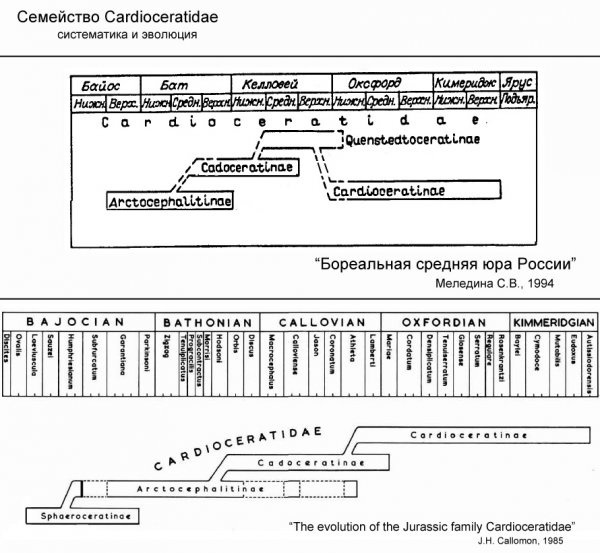
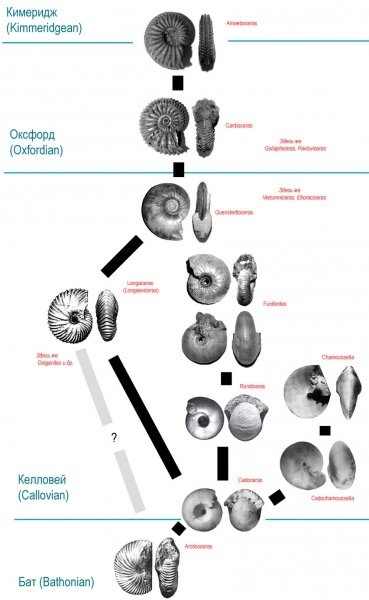
Кардиоцератиды появились на границе байоского и батского веков средней юры и вымерли в конце кимериджского века поздней юры. Они существовали почти 10 миллионов лет, в течение четырех геологических веков (бат, келловей, оксфорд, кимеридж), пережив за это время многие другие семейства аммонитов.В семействе Cardioceratidae разные палеонтологи выделяют три или четыре подсемейства. Два варианта систематики кардиоцератид пок азаны на схеме:
Добавьте описание
Верхняя схема дана по книге С.В. Мелединой [2], нижняя, по книге Дж. Кэлломона [1]. По моему мнению, правильно выделять 4 подсемейства – Arctocephalitinae, Cadoceratinae, Quenstedtoceratinae и Cardioceratinae. Нет никаких сомнений, что аммониты родов Quenstedtoceras, Vertumniceras и Eboraciceras заметно отличаются как от своих потомков кардиоцерасов (Cardioceratinae), так и от предков кадоцерасов (Cadoceratinae) и заслуживают выделения в отдельное подсемейство. Хотя, как это всегда бывает в природе, возникает вопрос с «пограничными» родами и видами, так как «последние» представители одного подсемейства часто мало отличаются от «первых» представителей другого подсемейства и где именно проводить границу совершенно непонятно. Но это частности, сейчас речь не о них. Предками кардиоцератид были аммониты семейства Sphaeroceratidae. Эти необычные аммониты с раковиной почти сферической формы и очень узким взрослым устьем не встречаются на Русской Платформе, так как во время их существования тут была суша, а не море.
Первые кардиоцератиды ответвились от них где-то на территории нынешних Сибири или Аляски. Дж. Кэлломон в своей книге, посвященной эволюции кардиоцератид [1], называет первыми представителями этого семейства аммонитов рода Cranocephalites, затем идут потомки этого рода – Arctocephalites и Arcticoceras. Меледина первым представителем Cardioceratidae считает род Boreiocephalites, а Cranocephalites ставит на второе место по старшинству в этом семействе [2]. Но это не суть важно – самое главное, что кардиоцератиды появились в позднем байосе где-то между Сибирью и Канадой.
На территории Подмосковья, да и вообще Центральной России в то время была суша, бродили динозавры, и этот, ранний, этап эволюции кардиоцератид здесь в палеонтологической летописи отсутствует
Добавьте описание
На этой иллюстрации показана примерная схема эволюции семейства Cardioceratidae. Обратите внимание, что почти все представленные здесь аммониты - макроконхи, кроме двух верхних (Cardioceras и Amoeboceras представлены микроконхами).
Несколько слов о том, что кардиоцератиды из себя представляли, как выглядели. Cardioceratidae - аммониты не очень крупные, гигантов среди них не было. Диаметр их макроконхов не превышал 20-ти сантиметров, то есть они были не крупнее современных наутилусов. Раковины микроконхов были еще меньше – от 1,5-2 до 10-12 сантиметров у разных видов.
Макроконх (самка) и два микроконха (самцы) рода Rondiceras. Возможно, что такие крупные раковины были нужны самкам не только для того, чтобы сформировать огромный объем икры, но и чтобы выносить эти икринки до рождения маленьких аммонитиков.
Кардиоцератиды были довольно необычными аммонитами – как и большинство аммонитид того времени, они были диморфными, то есть раковины самок (макроконхи) заметно отличались от раковин самцов (микроконхов), но у микроконхов кардиоцератид не было выростов-ушек на взрослом устье. Все соседи кардиоцератид, жившие в морях Центральной России в то время, были в той или иной степени «ушастыми», то есть у взрослых микроконхов были выросты-ушки по бокам устья. Ушки были и у приплывших с юга Oppeliidae, и у шипастных Kosmoceratidae, и у крайне многочисленных и эволюционно-успешных Perisphinctidae. Но не у кардиоцератид. У Cardioceratidae устье было украшено ростром – выступом на вентральной стороне. У самок-макроконхов тоже был выступ на устье, но, как правило, закругленный, не такой острый, как у самцов.
Этот ростр вызывает у палеонтологов много вопросов. На самом деле он был не только у кардиоцератид, многие семейства аммонитов имели вентральные выступы на устье. При этом не совсем понятно, имели ли эти ростры какое-то значение для размножения аммонитов, или они были необходимы им для чего-то другого – для быстрого плавания или для поддержки тела. Ушки аммонитов наверняка использовались при размножении, но для чего были нужны ростры – непонятно. Интересно также, что у самок аммонитов - кардиоцератид оксфордского рода Amoeboceras тоже вырастал полноценный ростр.
На протяжении всей истории семейства Cardioceratidae у этих аммонитов был очень заметный половой диморфизм. Самки были крупнее самцов, жили дольше и строили больше оборотов раковины. Вероятно, что и зрелости они достигали на год-полтора позже, чем родившиеся одновременно с ними самцы. В молодости представители обоих полов были ребристыми. На взрослых жилых камерах большинства макроконхов кардиоцератид ребра исчезают. Вообще, микроконхи и макроконхи кардиоцератид в детстве очень похожи, но у многих видов раковины самок даже на ранних оборотах заметно толще раковин самцов. У кардиоцератид, как и у многих других аммонитов, часто наблюдается наличие нескольких размерных классов внутри микро- и макроконхов. Особенно вариабельны в этом отношении микроконхи. Например микроконхи Quenstedtoceras взрослели с разной скоростью и достигали зрелости при разном размере. Макроконхи кардиоцератид обычно более консервативные в отношении размера взрослой раковины, но несколько размерных групп можно разглядеть и у них. Именно на кардиоцератидах (род Quenstedtoceras) польский палеонтолог Матый описал так называемые «миниконхи», которые противники теории полового диморфизма аммонитов поспешили объявить третьей морфой, отличной и от микроконхов и от макроконхов. [9] Но по фотографиям и описаниям, сделанным самим Матыем очевидно, что миниконхи – вовсе не «третий пол», а самые настоящие микроконхи – только маленькие. У них типично микроконховая ребристость, микроконховые ростры, учащение перегородок фрагмокона перед жилой камерой происходит так же, как и у более крупных микроконхов и т.д. Интересно еще и то, что у многих кардиоцератид очень сильно различается длина жилой камеры микро- и макроконхов. Например, у микроконхов Pseudocadoceras длина жилой камеры равняется половине оборота, а у соответствующих им макроконхов Rondiceras или Funiferites она превышает целый оборот. Например, у Rondiceras на жилая камера больше оборота:
А у Pseudocadoceras (правда, более поздних, соответствовавших Funiferites-ам) жилая камера (ее длина видна по следу на предыдущем обороте) всего лишь пол оборота:
Правда у поздних кардиоцератид – Quenstedtoceras, Cardioceras, Amoeboceras такой большой разницы в длине жилой камеры нет, хотя у самок объем жилой камеры все равно значительно превышает объем жилой камеры самцов. У кардиоцератид, причем у представителей разных подсемейств, известна полосчатая окраска раковин, которая, скорее всего, является прижизненной:
Но вернемся к истории семейства. Впервые в Центральной России и Поволжье кардиоцератиды появились в батском веке средней юры. Это были аммониты рода Arcticoceras, они приплыли в район нынешнего Саратова, где скорее всего впервые встретили аммонитов из тетической области – представителей семейства Parkinsoniidae. В.В. Митта нашел этих аммонитов в карьере у Сокурского тракта [7] Следом появились и потомки Arcticoceras-ов – аммониты рода Cadoceras - их обнаружили в знаменитом карьере у села Просек в нижегородской области в слоях батского возраста Д.Н.Киселев и М.А. Рогов [4]
Спустя недолгое время, когда море затопило территорию нынешних Москвы, Рязани, Ярославля – в общем, центральную часть России, кардиоцератиды – именно Cadoceras-ы, сразу начали активно осваивать новую территорию.
Тут нужно уточнить, что Cadoceras – это самки, макроконхи. Конечно, вместе с ними были и самцы – их значительно меньшие по размеру раковины условно относят к роду Pseudocadoceras. Многочисленные хорошо сохранившиеся раковины Cadoceras и Pseudocadoceras находят на реке Унже, а также в Саратовской и Нижегородской областях.
У кадоцерасов было два потомка – Cadochamoussetia и Rondiceras. Эти два макроконховых рода в плане формы раковины выбрали диаметрально противоположные пути развития. Кадошамоуссетии стали приобретать уплощенную, дисковидную форму с почти закрытым пупком, а рондицерасы наоборот, стали почти шаровидными с более или менее широким пупком. Пути постепенной эволюции обеих родов сейчас прослежены детально.
В итоге от рода Cadochamoussetia произошел новый род – Chamoussetia, раковины которого во взрослом состоянии стали толстыми оксиконами – то есть обороты раковины были довольно объемными, но вентральной стороны как таковой не было – боковые стороны сходились в острый ребристый киль. Chamoussetia, как и Cadoceras, были широко распространены, но существовали очень недолго – они вымерли еще в нижнем келловее. Исчезли и сами Cadoceras, хотя в разных местах они исчезали в разное время. В России они фактически «превратились» в Rondiceras, но в Великобритании какое-то время сосуществовали оба рода.
В итоге границу нижнего и среднего келловея на Русской Платформе пересекли лишь рондицерасы (это пересечение границы мы можем наблюдать, например, в Никитино). Правда, в среднем келловее они отказались от шарообразности и дали начало новому роду – Funiferites, который, фактически, пошел по пути кадошамоуссетий – стал уменьшать ширину раковины.
Любопытно, что микроконхи всех этих аммонитов менялись значительно меньше. Часто микроконхов трех родов - Cadoceras, Rondiceras и Funiferites относят к одному и тому же роду Pseudocadoceras. Некоторые авторы, правда, выделяют и другие микроконховые роды - Costacadoceras, Novocadoceras, но внешне их далеко не всегда возможно различить. Все они – маленькие аммониты с ребристыми, близкими к дисковидным раковинами, с заметным, но не очень длинным ростром на взрослом устье. Конечно, отличия между ними есть, например, микроконхи, которые соответствуют рондицерасам, чуть толще и ребристость у них грубее, а микроконхи, соответствующие фуниферитесам в среднем заметно мельче своих предков. Но конечно разница между ними куда меньше, чем между макроконхами. Это наводит на мысль, что изменялись не в целом условиях существования этих аммонитов, а лишь условия, в которых в некий период своей жизни оказывались самки. И скорее всего это был период вынашивания яиц в жилой камере.
В чем именно была причина изменения формы раковины, мы вряд ли когда-нибудь узнаем точно. Например, могла меняться скорость течений – при сильном течении самкам с дисковидными раковинами проще оставаться на одном месте. Самкам с икрой в жилых камерах вряд ли было легко и приятно бороться с течениями или куда-то плыть. Может быть дисковидные раковины действительно позволяли самкам Cadochamoussetia и Chamoussetia с меньшими затратами энергии сопротивляться сносу течением из удобных для них мест. А может быть, что в изменении формы раковины виноваты какие-то хищники – от одних хищников проще уплыть и тут потребуется дисковидная раковина, другим хищникам проще «подсунуть» гладкий шарик кадиконической раковины и пусть они кусают почти гладкую сферическую поверхность – все прокусить не сумеют. В общем, вариантов тут может быть много, но очевидно, что у самок кардиоцератид было больше причин менять форму своей раковины, чем у самцов.
Но давайте снова вернемся к эволюции кардиоцератид. До середины позднего келловея в Центральной России жили кардиоцератиды из подсемейства Cadoceratinae. Выше речь шла именно о них. Самцы у них были мелкие, самки очень крупные, с толстыми раковинами. Но в позднем келловее климат начал меняться, изменился режим осадконакопления, возможно, изменилась и глубина моря. Не даром в Московской и Рязанских областях – в регионах, где море всегда было не таким глубоким, как, например, в Поволжье, слои самого верхнего келловея представлены перемежающимися прослоями мергелей и глин.
В общем, фуниферитесы этого не вынесли и исчезли. Хотя история здесь темная и немного непонятная.
За Уралом, в Сибири, в районе нынешней реки Анабар в келловейском веке появился новый род – Longaeviceras. Предками его были, по видимому, ранние Cadoceras, а может быть даже и Arcticoceras. Эти лонгаевицерасы быстро распространились по Северному полушарию, доплыли до Англии (где и были впервые описаны). Впрочем, тут есть одна терминологическая трудность – палеонтолог Ю.С. Репин пришел к выводу, что английские Longaeviceras – это не совсем те аммониты, которые возникли в Сибири, а уже их потомки [8]. Сам исходных род он предложил назвать Longoceras. Тогда и приплывших в конце келловея на территорию Центральной России кардиоцератид из этой, сибирской ветви, нужно называть Longoceras. Но, в общем, этот вопрос чисто терминологический – отличия между Longaeviceras и Longoceras минимальны.
Интереснее другое – лонгаевицерасы-лонгоцерасы приплыли в нынешнюю Рязанскую область не после исчезновения Funiferites и Pseudocadoceras, а до него, изредка они встречаются в окрестностях Михайлова вместе с фуниферитами и псевдокадоцерасами. Причем, в это время среди микроконхов кардиоцератид значительно повышается разнообразие форм – некоторые из них выглядят как обычные Pseudocadoceras, но попадаются почти типичные Longoceras и даже экземпляры, очень похожие на Quenstedtoceras, о которых пойдет речь дальше. В то же время появляются странные аммониты, условно относимые к роду Eichwaldiceras. Вероятно, это боковая ветвь рода Funiferites.
Связано ли такое видообразование и смешение разных аммонитов просто с изменением климата и переселением, или они могли как-то скрещиваться друг с другом? Может быть Longoceras просто вытеснили своих родственников Funiferites, воспользовавшись изменением климата, а может быть их потомки – Quenstedtoceras, возникли от смешения родов? Интересно, что Longaeviceras встречаются и в более молодом слое – уже вместе с типичными Quenstedtoceras.
Так или иначе, от рода Longaeviceras/Longoceras произошел следующий род – Quenstedtoceras. Несмотря на широкую известность (даже в музеях США счет Quenstedtoceras из карьера «Дубки» идет на тысячи) и широкое распространение (их находят даже во Франции), Quenstedtoceras-ы существовали очень недолго. Весь келловейский век продолжался всего лишь 3,5 млн. лет, а Quenstedtoceras-ы жили в самом его конце, 500 тысяч, от силы миллион лет, если не меньше.
Но именно за ними было будущее. На рубеже келловея и оксфорда, когда некоторые семейства аммонитов вымерли, а другие были вынуждены покинуть Центральную Россию, от квенштедтоцерасов произошел новый род – Cardioceras. Он-то и дал название всему семейству. Как часто бывает в таких случаях, толком непонятно, произошло ли образование нового рода в «одной точке», или разные Cardioceras произошли от разных Quenstedtoceras. Но в любом случае именно представители этого рода и его потомки стали доминирующей группой аммонитов в северных морях на протяжении оксфорда и части кимериджа.
У кардиоцерасов был красивый рельефный киль на вентральной стороне, ростр на устье и такой же экстремальный половой диморфизм, как и у его предков. Правда, самки-макроконхи Cardioceras были не толстыми, они имели крупную раковину дисковидной формы. Кстати, макроконхи некоторых Cardioceras очень похожи на макроконхов вымерших к тому времени Chamoussetia, но прямой связи предок-потомок между ними нет, есть лишь общие предки и, по видимому, сходные условия окружающей среды.
В оксфордском веке кардиоцерасы составляли большинство в аммонитовых фаунах северных морей. Кроме них здесь жили также аммониты семейства Perisphinctidae и ближайшие родственники и потомки перисфинктид - Aspidoceratidae. Но Cardioceras явно диминировали по численности. Они далеко продвинулись на юг, достигнув современного Средиземноморья в Европе и границы США и Канады в Северной Америке.
В середине оксфордского века от рода Cardioceras произошел род Amoeboceras, отличающийся в общем-то незначительными деталями строения раковины. У Amoeboceras также был заметно выделяющийся киль на вентральной стороне. Этот род процветал и перешел в следующий век – кимериджский, который оказался последним для семейства Cardioceratidae.
В начале кимериджского века амоебоцерасы все еще были многочисленны и разнообразны, но к концу этого времени они стали исчезать. Что-то такое происходило в северных морях, к чему не могли приспособиться даже Cardioceratidae, пережившие стольких других аммонитов. Амоебоцерасы измельчали – последних из них даже выделили в отдельный род (или подрод) – Nannocardioceras. Но «наноразмеры» не помогли им, незадолго до конца кимериджа они вымерли. Может быть, где-то они могли дотянуть и до самого конца кимериджа, кто знает, но в следующем, волжском веке, кардиоцератид уже не было. На этом история семейства Cardioceratidae заканчивается.
Моя небольшая коллекция юрских аммонитов Подмосковья.
Этой небольшой публикацией я хочу открыть серию статей, посвященных различным семействам аммонитов, обитавших на территории Центральной России.
Как известно, аммониты, жители палеозойских и мезозойских морей, были очень разнообразны. Каких только раковин у них не было – были и дисковидные, и шаровидные, и похожие на колесо (иногда не только по форме, но и по размеру). Причина такого разнообразия в очень быстрой эволюции аммонитов. Они сменяли друг друга, приспосабливаясь к изменчивым условиям окружающей среды. Это только нам, «со стороны» кажется, что тот же юрский период был временем равномерно теплого климата и полного спокойствия в биосфере – на самом деле и в юре, и в мелу формировались горы, наступали или наоборот отступали и становились мелкими моря, температура воды в них падала или повышалась. Конечно, не было таких перепадов как сейчас, не было оледенений, но все равно было множество факторов, которые не давали аммонитам жить спокойно и заставляли постоянно приспосабливаться и изменяться.
Фото Михайловцемент. Стратиграфия в картинках., автор Dimka
На этой схеме, которую нарисовал уважаемый Dimka, хорошо видно разнообразие аммонитов в одном единственном карьере.
Иногда, в разговоре об аммонитах от начинающих любителей палеонтологии можно услышать сожаление – «эх, не вымерли бы аммониты, ловили бы мы сейчас в морях виргатитов и космоцерасов!». Увы, даже если бы аммониты не вымерли, ни аммонитов рода Kosmoceras, ни рода Virgatites мы бы не встретили. Они и друг с другом-то не встречались никогда, их разделяли несколько миллионов лет, оба эти рода исчезли навсегда, когда до окончания мелового периода и заката «эры аммонитов» оставалось больше 80-ти миллионов лет. Аммониты появлялись и вымирали постоянно. То же самое можно сказать и о других животных, хотя некоторые группы – например мезозойские наутилусы двустворчатые моллюски эволюционировали значительно медленнее.
Если бы аммониты не вымерли, то даже сложно себе представить, какими бы были их потомки через 65 миллионов лет эволюции. Но как они (вернее, их раковины) выглядели в прошедшие эпохи, мы знаем и о них, об их эволюции я и хочу рассказать в этом цикле статей.
Это будет серия небольших статей, посвященных разным семействам аммонитов. Я буду писать только о тех аммонитах, с которыми знаком лично, поэтому начну рассказ с юрских семейств, а потом, может быть, дойду и до мела.
Героями моих статей будут в основном аммониты, обитавшие в морях Русской платформы, точнее, в Центральной России и Поволжье, аммониты бореальные и суббореальные. Здесь необходимо пояснить, что море, занимавшее в юрском и меловом периодах большую часть современной России, было относительно мелководным и относительно холодным. На севере оно соединялось с Бореальным (полярным) океаном, на юге и востоке – с заливами океана Тетис.
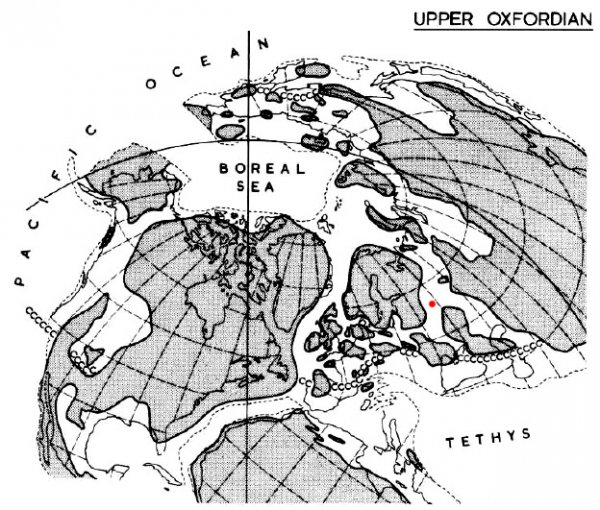
Северное полушарие в оксфордском веке юрского периода. Иллюстрация из книги Callomon J.H. (1985) "The evolution of the Jurassic ammonite family Cardioceratidae". Красной точкой отмечена Москва. Рядом букв "с" - южная граница распространения аммонитов семейства Cardioceratidae в то время.
Конечно, это описание – довольно приблизительная реконструкция, так как ситуация постоянно менялась, например в волжском веке практически пересохли проливы, соединяющие наше море с Тетисом. Но главное тут вот в чем – в океане Тетис были значительно большие глубины и широкие просторы. Там жили аммониты, которые не встречались в мелководных морях Центральной России. В первую очередь это представители подотрядов Phylloceratina и Lytoceratina. Именно поэтому их много в Крыму и на Кавказе (они располагались у края океана Тетис), но совсем нет в Подмосковье или на Приполярном Урале. Бореальные аммониты могли довольно далеко заходить на юг, практически до самого современного Средиземноморья, за которым уже начинался открытый океан. В обратном направлении – на север, аммониты тоже расселялись, например представители тетического семейства Oppeliidae. Но все эти мигранты с юга относились к подотряду Ammonitida, в то время как литоцератины и филлоцератины не хотели покидать материковые склоны океана, мелководные просторы Центральной России их не привлекали.
Поэтому мои истории пока будут посвящены только аммонитидам. Выбор в качестве главных объектов именно семейств аммонитов, а не подсемейств или родов, связан с тем, что чем меньшего уровня таксон мы рассматриваем, тем меньше получается однозначности и ясности. Если на уровне семейств аммонитов у специалистов не так уж много разногласий, то в вопросах, касающихся родов и видов, этих самых разногласий не просто много, а очень много. Кроме того, именно семейства аммонитов можно считать достоверными эволюционными рядами – в их выделении гораздо меньше субъективности, чем в выделении родов и видов, аммониты, отнесенные к тому или иному семейству действительно обладают определенными наборами хорошо заметных признаков и по изменениям этих признаков можно прослеживать ход эволюции аммонитов.
Моё фото. Аммонит Ammonitoceras. Ранний мел: Северный Кавказ. Палеонтологический музей. им Ю.А Орлова
Моё фото. Аммонит Ammonitoceras. Ранний мел: Северный Кавказ. Палеонтологический музей. им Ю.А Орлова
Моё фото. Аммонит Speetoniceras Versicolor. Ранний мел: Ульяновская область. Палеонтологический музей. им Ю.А Орлова
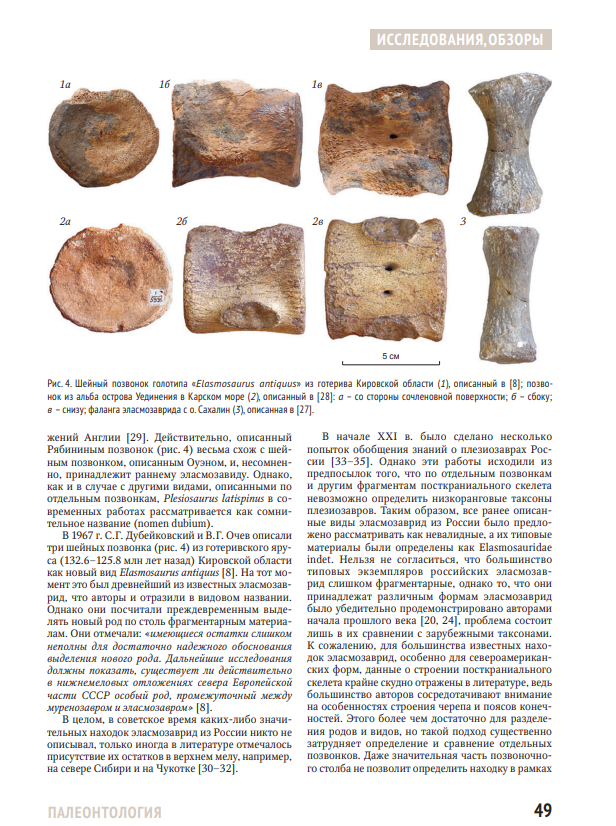
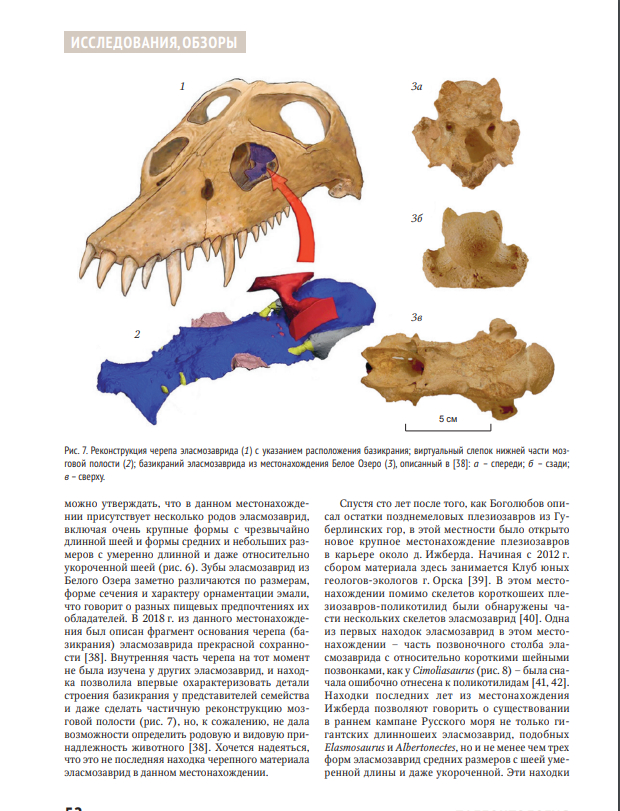
Обзорная статья российских палеонтологов Н.Г. Зверькова и М.С. Архангельского, посвященная меловым плезиозаврам-эласмозавридам России.Плезиозавры семейства Elasmosauridae получили широкую известность благодаря своим длинным шеям с рекордным количеством позвонков. В настоящее время известно множество родов и видов эласмозаврид: от небольших до гигантских, от сверхдлинношеих до вторично короткошеих. Остатки эласмозаврид описаны из всех регионов мира и часто встречаются в меловых отложениях России. Некоторые исторические находки из России были описаны как новые виды, но впоследствии не получили признания в научном сообществе. Новые находки эласмозаврид из России проливают свет на таксономическое разнообразие и эволюционную историю этого семейства в Евразии.