


Эоценовая ящерица в балтийском янтаре

Взято отсюда: https://vk.com/club.dinosaurtime2018
Показать полностью
1
Взято отсюда: https://vk.com/club.dinosaurtime2018

Рис. 1. Проартикуляты — представители эдиакарской биоты с уплощенным сегментированным телом и «симметрией скользящего отражения» (сегменты левой и правой стороны тела сдвинуты относительно друг друга; особенно хорошо это заметно у вендии). Слева направо и сверху вниз: дикинсония, вендия, андива (см. Andiva), ёргия (Yorgia), сприггина. Изображения из обсуждаемой статьи в Science, из статьи A. Yu. Ivantsov, 2002. New Proarticulata from the Vendian of the Arkhangel’sk Region и с сайтов en.wikipedia.org, en.wikipedia.org и ediacaran.org
Дикинсония и ее родня (так называемые проартикуляты) — одни из самых странных и загадочных представителей эдиакарской биоты, населявшей моря 571–541 млн лет назад, незадолго до кембрийского взрыва. Проартикуляты так сильно отличаются от всех современных организмов, что определить их место в системе живой природы долго не удавалось: трактовки варьировали от колониальных бактерий и гигантских протистов до наземных лишайников и высших животных (билатерий). Палеонтологи из России, Австралии и Германии внесли ясность в этот запутанный вопрос, изучив «молекулярные ископаемые» — остатки эукариотических мембранных стеролов, сохранившиеся в отпечатках дикинсоний из местонахождения Лямца (Онежский берег Белого моря). Полученные результаты позволяют уверенно отнести дикинсонию (а значит, и ее родню — других проартикулят) к царству животных. Открытие показало, что расцвет эдиакарской биоты действительно был прелюдией к бурной диверсификации животных в начале кембрия, а не «черновым экспериментом природы» по созданию каких-то альтернативных форм макроскопической жизни, как предполагали некоторые специалисты.
Изучение «молекулярных ископаемых» (см. Molecular fossils) — остатков органических молекул, некогда входивших в состав живых организмов, — относительно новый и весьма многообещающий метод палеонтологических исследований. Он может оказаться особенно полезным для выяснения природы ископаемых существ, настолько непохожих на современные организмы, что традиционными методами определить их место в системе живой природы не удается.
Эдиакарская биота почти целиком состоит как раз из таких загадочных существ. Как их только не интерпретировали: и как настоящих двусторонне-симметричных животных, и как колонии бактерий, и как гигантских протистов, и даже как наземные лишайники (см.: Могли ли эдиакарские ископаемые жить на суше?, «Элементы», 01.02.2013).
На сегодняшний день более или менее ясно, что эдиакарская биота неоднородна и включает представителей разных групп морских (все-таки не наземных!) организмов. Из них к царству животных до сих пор уверенно относили только кимбереллу и с чуть меньшей уверенностью — еще несколько форм (Cloudinidae, Namapoikia, Namacalathus).
Большинство других представителей эдиакарской биоты по-прежнему остаются в категории «проблематиков», то есть ископаемых организмов неясной систематической принадлежности. Это справедливо и для проартикулят (Proarticulata) — крайне своеобразной группы эдиакарских организмов (рис. 1). Изначально их интерпретировали как билатерий, но потом выяснилось, что по целому ряду признаков проартикуляты резко отличаются от всех известных двусторонне-симметричных животных. В частности, у них не удалось обнаружить ни рта, ни кишечника (возможно, у них было наружное пищеварение или они просто питались растворенной органикой, впитывая ее всей поверхностью тела). Проартикуляты оставляли на бактериально-водорослевых матах, покрывавших морское дно, очень странные следы передвижения — цепочки одинаковых, по-разному ориентированных отпечатков всего своего тела. Возможно, они переплывали с места на место при помощи ресничного эпителия, но не похоже, что у них были мышцы. Своеобразная «симметрия скользящего отражения», характерная для проартикулят (она хорошо видна у вендии на рис. 1), не встречается у животных. Не очевидно даже, были ли у проартикулят морфологически различимые брюшная и спинная стороны тела. Учитывая эти факты, наиболее правдоподобной выглядела трактовка проартикулят как неких очень примитивных, «базальных» животных, однако и другие версии нельзя было полностью исключить.
Поскольку эдиакарские организмы нередко сохраняются в виде органических пленок, логично попробовать применить к ним методы «молекулярной палеонтологии». Именно это и сделала группа палеонтологов из России, Австралии и Германии. Совсем недавно, в начале 2018 года, эта группа сообщила о первом крупном успехе в деле молекулярной идентификации загадочных эдиакарских ископаемых. Исследователям удалось показать, что круглые отпечатки, известные под названием Beltanelliformis, представляют собой колонии цианобактерий (см.: Загадочные эдиакарские организмы Beltanelliformis оказались колониями цианобактерий, «Элементы», 25.01.2018).
Новая статья, опубликованная 21 сентября в журнале Science, сообщает о еще более выдающемся достижении. Авторы изучили состав органических пленок в отпечатках двух представителей проартикулят — дикинсонии и андивы (см. Andiva). Материал был собран в России на побережье Белого моря (местонахождения Лямца и Зимние Горы). При помощи газовой хроматографии и масс-спектрометрии (см. Gas chromatography–mass spectrometry) анализировался состав насыщенных стеранов и моноароматических стероидов — веществ, в которые превращаются эукариотические стеролы (компоненты клеточных мембран эукариот) в процессе фоссилизации. Группы эукариот различаются по соотношению стеролов с углеродными «скелетами», состоящими из 27, 28 или 29 атомов углерода (их называют, соответственно, холестероидами, эргостероидами и стигмастероидами). Например, отличительной особенностью животных является абсолютное преобладание 27-углеродных холестероидов, у зеленых водорослей больше всего 29-углеродных стигмастероидов, а у многих грибов преобладают 28-углеродные эргостероиды.
Наиболее четкие результаты дал анализ крупных дикинсоний из Лямцы, сохранившихся в виде органических пленок толщиной до 3 мкм. Изученные экземпляры андивы с Зимних Гор оказались более тонкими и менее информативными, но в общих чертах результаты по андиве и дикинсонии получились похожие.
При жизни проартикуляты лежали на поверхности бактериально-водорослевой пленки, покрывавшей морское дно. Поэтому не вся органика в отпечатке дикинсонии происходит от самой дикинсонии: частично она представляет собой остатки этой пленки. В состав пленки, помимо прокариот, входили зеленые водоросли со своим характерным набором стеролов. Поэтому, чтобы понять, какие стеролы были у дикинсонии, нужно вычесть из результатов анализа «фоновый» водорослевый сигнал.
Оказалось, что в породе, прилегающей к отпечаткам дикинсоний, среди моноароматических стероидов преобладают 29-углеродные стигмастероиды (их доля составляет от 71,3 до 76,0%), на втором месте 28-углеродные эргостероиды (13,4–16,8%), на третьем — 27-углеродные холестероиды (10,6–11,9%). Примерно такое же соотношение молекул с 29, 28 и 27 атомами углерода обнаружилось и среди насыщенных стеранов. Это характерная молекулярная «подпись» зеленых водорослей, которые, по-видимому, представляли собой доминирующую группу эукариот на дне эдиакарского моря.
Совсем другая картина наблюдается в крупных отпечатках дикинсонии: здесь резко преобладают характерные для животных 27-углеродные холестероиды (93%), а эргостероидов и стигмастероидов относительно немного (1,8 и 5,2%). Выяснилось также, что чем мельче дикинсония, тем ниже в ней доля холестероидов (то есть «животного» компонента) и выше доля стигмастероидов (водорослевого компонента). Это логично, потому что дикинсонии разного размера, толстые и тонкие, при жизни лежали на водорослевой пленке одной и той же толщины. Соответственно, в ископаемом отпечатке мелкой дикинсонии должна быть повышена доля водорослевой примеси по сравнению с отпечатком крупной дикинсонии. Именно это и показал проведенный анализ (рис. 2).
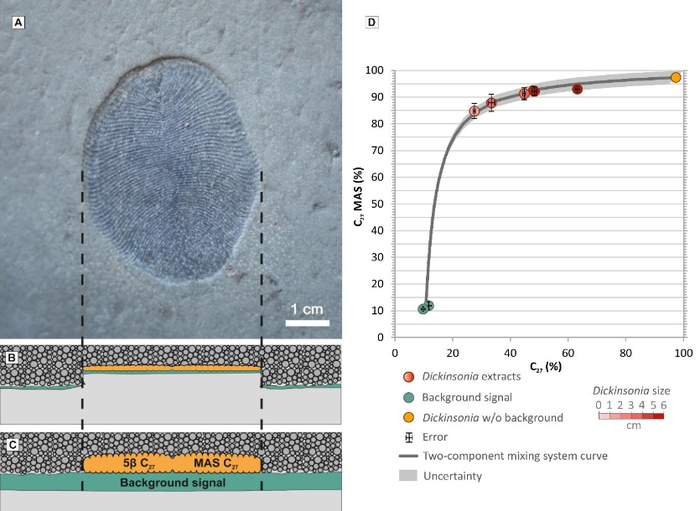
Рис. 2. Слева: отпечаток дикинсонии (A) и схема, иллюстрирующая процесс его формирования (B, C). Дикинсония (оранжевая) с повышенным содержанием 27-углеродных стеролов лежала на водорослевом мате (зеленом) — источнике «фонового сигнала» (background signal), то есть набора стеролов, характерного для зеленых водорослей (с преобладанием 29-углеродных молекул). Дикинсонию засыпало песком (C), затем органика сплющилась, и на нижней поверхности слоя песчаника сформировался отпечаток (B). Рисунок показывает, что чем тоньше была дикинсония при жизни, тем сильнее должен быть «фоновый сигнал» в отпечатке. Справа (D): график, показывающий связь между долей 27-углеродных моноароматических стероидов (по вертикальной оси), 27-углеродных стеранов (по горизонтальной оси) и размером дикинсонии (Dickinsonia size): бледно-розовые кружки соответствуют самым мелким экземплярам, темно-красные — самым крупным. Зелеными кружками показан «фоновый сигнал», оранжевый кружок — реконструированный состав стеролов дикинсонии за вычетом фонового сигнала. Изображение из дополнительных материалов к обсуждаемой статье в Science
По соотношению стереоизомеров стеранов (5б и 5в) можно понять, в каких условиях шло разложение органики — кислородных или бескислородных. Судя по этому показателю, в ходе формирования изученных отпечатков дикинсонии 27-углеродные стеролы находились в бескислородных условиях, что характерно для разлагающихся останков животных. Напротив, 28 и 29-углеродные стеролы, скорее всего, разлагались в присутствии кислорода. Этот результат еще раз подтверждает, что 27-углеродные стеролы дикинсонии — ее собственные, а 28 и 29-углеродные представляют собой водорослевую примесь.
На основе полученных данных авторы рассчитали, что при жизни у дикинсонии доля 27-углеродных стеролов составляла не менее 99,7%, а возможно, и все 100%.
Из этого можно сделать вполне определенные выводы о таксономическом положении дикинсонии (а заодно и других проартикулят).
Они точно не были лишайниками: у лишайников всегда преобладают 28-углеродные эргостероиды, которых дикинсония при жизни почти или вовсе не производила. Они также не могли быть гигантскими протистами: все изученные протисты, достигающие сколько-нибудь крупных размеров, производят смесь 27, 28 и 29-углеродных стероидов, а многие из них синтезируют вдобавок 30-углеродные, которые в отпечатках дикинсоний отсутствуют.
Характерной особенностью животных является присутствие только 27-углеродных стеролов у подавляющего большинства видов. У протистов, являющихся ближайшими родственниками животных, а именно у хоанофлагеллят (см. Choanoflagellate) и Filasterea, доля 27-углеродных стеролов варьирует от 84 до 100%. Но сближать их с проартикулятами нет оснований, потому что эти протисты всегда имеют микроскопические размеры.
Таким образом, полученные данные убедительно свидетельствуют в пользу отнесения проартикулят к животному царству. Самые древние экземпляры дикинсоний имеют возраст 558 млн лет, что делает их, по мнению авторов, самыми древними «подтвержденными макроскопическими животными» (confirmed macroscopic animals). Древнейшие кимбереллы чуть моложе (555 млн лет).
Поскольку ни рта, ни кишечника, ни мускулатуры у проартикулят так и не удалось обнаружить, они, скорее всего, представляют собой одну из базальных ветвей животного царства наряду с трихоплаксом и губками.
Источник: Ilya Bobrovskiy, Janet M. Hope, Andrey Ivantsov, Benjamin J. Nettersheim, Christian Hallmann, Jochen J. Brocks. Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals // Science. 2018. V. 361. P. 1246–1249.
Александр Марков
http://elementy.ru/novosti_nauki/433336/Podtverzhdena_prinad...
Палеонтологи обнаружили в меловом бирманском янтаре предполагаемых каллиграмматид — хоботковых сетчатокрылых насекомых, которые напоминали современных бабочек по своему внешнему облику и особенностям биологии. Находка доказывает, что каллиграмматиды, появившиеся еще в ранней юре, сумели дожить до середины мелового периода, то есть их эволюционная история насчитывает по меньшей мере 80 млн лет. Благодаря замечательной сохранности хоботков каллиграмматид, которую обеспечивает янтарь, ученым удалось разобраться в особенностях их строения. С помощью хоботков каллиграмматиды могли высасывать опылительные капли голосеменных растений, попутно занимаясь опылением. Видимо, из-за слишком тесной связи с голосеменными эти насекомые и вымерли, когда в середине мела началась экспансия цветковых растений.
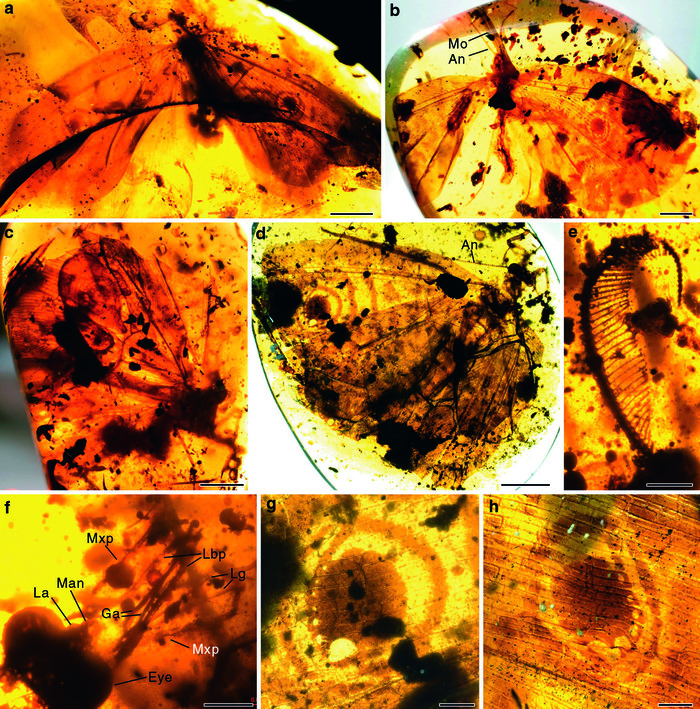
Рис. 1. Каллиграмматида Burmogramma liui из бирманского янтаря. a, b, c, d — внешний вид, e — гребенчатая антенна самца, f — детали строения ротового аппарата, g, h — глазчатые пятна на крыльях. An — антенна, Ga — галеа, La — верхняя губа, Lbp — нижнегубной щупик, Lg — лигула, Man — мандибула, Mo — ротовые части, Mxp — максиллярный щупик. Фото из обсуждаемой статьи в Nature Communications
Каллиграмматиды (Kalligrammatidae) — это вымершее семейство, относящееся к отряду сетчатокрылых насекомых (Neuroptera). В состав этого отряда помимо каллиграмматид входят всем известные златоглазки и муравьиные львы, а также более экзотичные создания вроде нитекрылок и мантиспид. Всех их находят в ископаемом состоянии, но именно каллиграмматиды благодаря своему конвергентному сходству с дневными бабочками являются одними из самых узнаваемых и запоминающихся древних насекомых. Впервые каллиграмматид нашли в начале XX века в юрских отложениях Германии, затем — на территории бывшей Казахской ССР, а в 1990–2000-е годы этих мезозойских «бабочек» стали в большом количестве находить в Китае (см. картинку дня «„Бабочки“ мезозоя»). Теперь они отыскались и в янтаре, который добывают в Мьянме (Бирме) и затем перепродают в другие страны.
Все предыдущие находки каллиграмматид (а всего к настоящему моменту описано 17 родов и почти 50 видов этого семейства) представляли собой плоские отпечатки на каменной породе. Так что находка в бирмите впервые позволяет изучить их в буквальном смысле со всех сторон, ведь для «янтарных» насекомых характерна трехмерная сохранность.
Всего в руки ученых попало 25 кусков янтаря с каллиграмматидами, относящимися к 7 видам и 5 родам. Самый впечатляющий из этих видов — Burmogramma liui, который на крыльях несет характерные глазчатые пятна, «фирменный знак» многих классических каллиграмматид. Считается, что благодаря сходству этих пятен с глазами позвоночных каллиграмматиды, подобно павлиньему глазу и другим современным дневным бабочкам, могли отпугивать хищников. А врагов у них в середине мела, как, впрочем, и всегда, было немало — достаточно вспомнить о птицах и ящерицах, найденных в том же бирманском янтаре (Птенец из бирманского янтаря помог уточнить особенности развития мезозойских птиц, «Элементы», 12.06.2017).
Другие бирманские каллиграмматиды тоже несут пятна на крыльях, но простые, без светлого ободка внутри, создающего иллюзию присутствия зрачка. Надо сказать, что три из пяти родов (Oligopsychopsis, Cretanallachius и Burmopsychops), которые авторы новой статьи причисляют к каллиграмматидам, уже находили в бирмите и ранее. Однако об их систематической принадлежности шли споры. Некоторые исследователи относили три вышеперечисленных рода к современному семейству Dilaridae. По-английски это семейство почему-то называют «приятные сетчатокрылые» (pleasing lacewings), хотя ничего особенно приятного в этих невзрачных существах нет. Единственное, чем могут похвастаться дилариды, — это гребенчатые антенны у самцов. Ни у каких других современных сетчатокрылых таких антенн нет. Зато они есть у Oligopsychopsis и его сородичей из бирмита, что и стало главным аргументом в пользу их отнесения к диларидам (Diying Huang et al., 2015. The first Mesozoic pleasing lacewing (Neuroptera: Dilaridae)).

Рис. 2. Гребенчатоусые сетчатокрылые из бирманского янтаря, которых отнесли к каллиграмматидам. a — Burmopsychops labandeirai, b, с — Cretogramma engeli и его гребенчатая антенна, d — Cretanallachius magnificus, e — Oligopsychopsis grandis, f — Cretogramma engeli. Фото из обсуждаемой статьи в Nature Communications
Другие же авторы просто помещали загадочных гребенчатоусых из бирмита в сборную группу психопсидообразных (Psychopsoidea) — эти сетчатокрылые с широкими многожилковыми крыльями были очень разнообразны и обильны на протяжении почти всего мезозоя (Xiu-Mei Lu et al, 2016. New long-proboscid lacewings of the mid-Cretaceous provide insights into ancient plant-pollinator interactions). Тем не менее систематические отношения, в которых состоят различные древние психопсидообразные, весьма запутанны, и отнесение к ним Oligopsychopsis и прочих бирманских родов вносило еще больше неясности в общую картину. Но теперь ситуация, по крайней мере по мнению авторов статьи, прояснилась. Вновь обнаруженное сетчатокрылое Burmogramma с глазчатыми крыльями и гребенчатыми антеннами похоже на каллиграмматид, а ранее найденные гребенчатоусые неясной принадлежности похожи на Burmogramma — значит, всех этих насекомых можно скопом зачислить в семейство Kalligrammatidae в качестве отдельного подсемейства.
Это подсемейство, объединяющее все пять родов бирманских каллиграмматидоподобных сетчатокрылых, носит название Cretanallachiinae. Среди прочих его отличительных признаков — наличие хоботка. Это важный аргумент в пользу родства Cretanallachiinae с каллиграмматидами, ведь, помимо этого семейства, ни у кого из современных или вымерших сетчатокрылых хоботка нет и в помине. Есть еще только одно известное исключение — это сетчатокрылое Paradoxosisyra groehni из семейства Sisyridae с коротким колющим хоботком, недавно найденное опять-таки в бирмите (вероятно, оно могло питаться кровью амфибий или гемолимфой других насекомых, см.: V. N. Makarkin, 2016. Enormously long, siphonate mouthparts of a new, oldest known spongillafly (Neuroptera, Sisyridae) from Burmese amber imply nectarivory or hematophagy). Но на сизирид представители Cretanallachiinae явно не похожи, а вот с каллиграмматидами их объединяют, помимо наличия хоботка и глазчатых пятен, также некоторые особенности жилкования, вроде сильно разветвленной жилки MP в переднем крыле.

Рис. 3. Палеонтологическая реконструкция, на которой художник изобразил юрских каллиграмматид, найденных близ китайской деревни Даохугоу (a) и меловых каллиграмматид из бирманского янтаря (b). Изображение: Phys.org
Традиционно считалось, что хоботок каллиграмматид был устроен примерно так же, как и у бабочек, представляя собой трубочку, составленную из двух створок (видоизмененных галеа). По бокам хоботка располагалась всего одна пара членистых максиллярных щупиков. Такой вывод был сделан по итогам изучения каменных отпечатков каллиграмматид с сохранившимися ротовыми частями. Однако анализ ротовых частей каллиграмматид из бирмита свидетельствует о более сложном строении. Во-первых, хоботок Cretanallachiinae «сделан» не из двух, а из четырех частей — по мнению авторов статьи, они представляют собой парные галеа и лигулы (отростки нижней губы). Во-вторых, рядом с хоботком этих сетчатокрылых находится не одна, а две пары щупиков — максиллярные и нижнегубные. Авторы статьи дают понять, что точно так же дело могло обстоять и у всех остальных каллиграмматид — просто на каменном материале детали строения хоботка видны хуже, чем на янтарном.
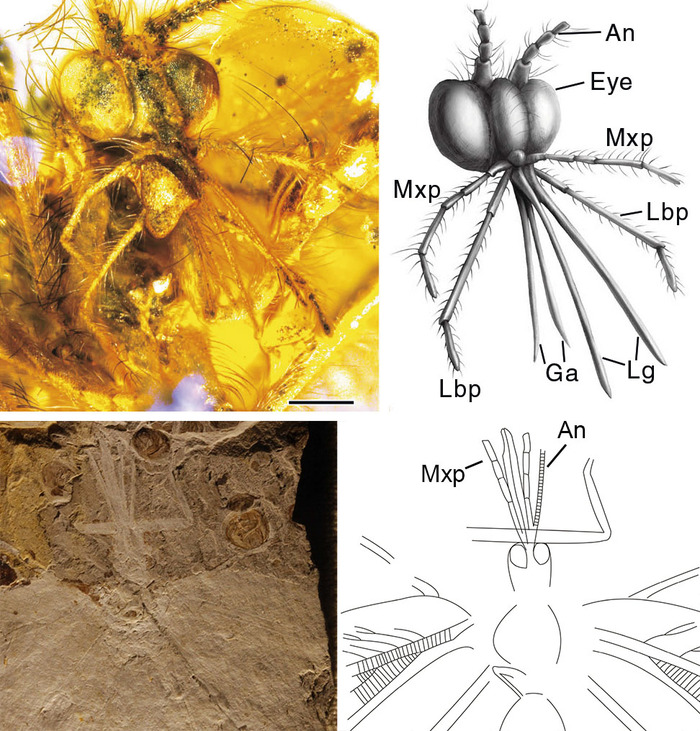
Рис. 4. Вверху — ротовые части бирманской каллиграмматиды Oligopsychopsis groehni. Изображение из обсуждаемой статьи в Nature Communications. Внизу — ротовые части классической каллиграмматиды из средней юры Китая: An — антенна, Ga — галеа, Lbp — нижнегубной щупик, Lg — лигула, Man — мандибула, Mo — ротовые части, Mxp — максиллярный щупик. Изображение из статьи: Q. Yang et al., 2014. Mesozoic lacewings from China provide phylogenetic insight into evolution of the Kalligrammatidae (Neuroptera)
У каллиграмматид из бирманского янтаря длина хоботка составляла всего 0,6–3,2 мм. Это весьма скромный размер, учитывая, что у каллиграмматид, известных по каменным отпечаткам, хоботок достигал 5–18 мм. Тем не менее даже такое короткое «орудие» всё же позволяло представителям подсемейства Cretanallachiinae посещать генеративные органы голосеменных растений. Две пары длинных волосистых щупиков помогали им снимать пробу с пыльцы и сладковатых выделений. А вот гребенчатые антенны почти наверняка использовались самцами Cretanallachiinae, чтобы находить партнерш по воздушному следу из феромонов, которые те испускали. В понимании авторов работы, каллиграмматиды, сохранившиеся в бирманском янтаре, — это «последние из могикан», самая молодая находка данного семейства. Судя по ней, даже на излете своей эволюции каллиграмматиды продолжали демонстрировать высокий уровень разнообразия.
А теперь пришло время высказать несколько критических замечаний. Дело в том, что каллиграмматиды, обнаруженные в бирмите, существенно отличаются от типичных представителей семейства. Во-первых, они гораздо мельче: максимальная длина передних крыльев Cretanallachiinae составляет всего около 3 см, тогда как классические каллиграмматиды в среднем в 2–3 раза крупнее. Во-вторых, для обычных каллиграмматид очень характерны плотно расположенные поперечные жилки, образующие как бы мелкоячеистую решетку. Эта «решетка» позволяет безошибочно идентифицировать даже небольшие фрагменты крыльев каллиграмматид, встречающиеся в осадочных породах. А вот у находок из бирмита ничего подобного нет — поперечные жилки по их крыльям разбросаны весьма скупо. В-третьих, ни у одних каллиграмматид, кроме бирманских, нет гребенчатых антенн. И наконец, в-четвертых, нельзя сбрасывать со счетов и различие в строении ротового аппарата. Факт остается фактом: на всех каменных отпечатках хоботок окружен одной парой щупиков, а у бирманских каллиграмматид — двумя парами. Сохранность каменного материала, конечно, хуже, но не настолько, чтобы четыре можно было принять за два.
Иначе говоря, нет уверенности в том, что палеонтологи нашли в янтаре именно каллиграмматид. Возможно, это какая-то другая группа сетчатокрылых, например одна из ветвей психопсидообразных, которые просто пошли по тому же пути, что и каллиграмматиды: обзавелись хоботком, глазчатыми пятнами и отправились на поиски «сладенького». Если это так, то расклад становится даже более интересным. Получается, сетчатокрылые на протяжении мезозоя дважды независимо друг от друга пытались сделать из себя «бабочек», то есть ширококрылых хоботковых нектарофагов. Но затем, ближе к началу кайнозоя, эту же модель гораздо более успешно реализовали чешуекрылые, дав начало настоящим хоботковым бабочкам, и именно они, а не сетчатокрылые, извлекли пользу из появления цветковых растений.
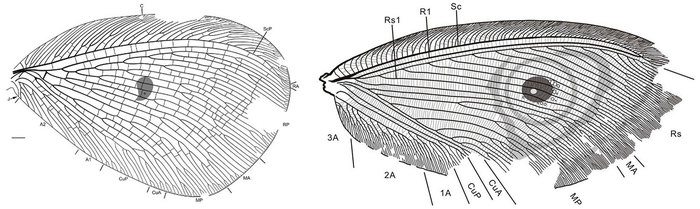
Рис. 5. Слева — крыло классической каллиграмматиды Kalligramma paradoxum из средней юры Китая. Изображение из статьи: Q. Liu et al., 2013. Two new kalligrammatids (Insecta, Neuroptera) from the Middle Jurassic of Daohugou, Inner Mongolia, China. Справа — крыло каллиграмматиды Oligopsychopsis groehni из бирманского янтаря. Обратите внимание на плотность расположения поперечных жилок. Изображение из обсуждамой статьи в Nature Communications
Это не такая уж и нетипичная ситуация для эволюции: известно, что в определенные эпохи разные группы живых существ параллельно начинают экспериментировать с одним и тем же типом организации. В этой связи академик-палеонтолог Леонид Татаринов говорил о «маммализации», то есть о независимом появлении признаков млекопитающих у разных зверообразных рептилий. Имела место также и «артроподизация», когда кембрийские беспозвоночные разными путями пытались превратиться в членистоногих. Можно говорить и об «ангиоспермизации», когда различные группы голосеменных растений эволюционировали в одном и том же направлении, обзаводясь признаками цветковых. Точно так же и у насекомых могла происходить «бабочкизация» или, если использовать латинский корень, «глоссатизация» (от лат. Glossata — ‘хоботковые чешуекрылые’).
Самая молодая находка каллиграмматид, в которой не приходится сомневаться, была сделана в меловых отложениях Бразилии аптского яруса (125–113 млн лет). Кстати, именно оттуда происходят наиболее крупные каллиграмматиды размахом крыльев до 32–33 см (Günter Bechly, Vladimir N. Makarkin, 2015. A new gigantic lacewing species (Insecta: Neuroptera) from the Lower Cretaceous of Brazil confirms the occurrence of Kalligrammatidae in the Americas). По возрасту это не так уж и далеко от бирманского янтаря, датируемого примерно 100 миллионами лет. Вполне возможно, что наряду с мелкими каллиграмматидоподобными сетчатокрылыми Cretanallachiinae, описанными в статье, в это же время существовали и каллиграмматиды в классическом смысле слова. Но только они были слишком большими, чтобы прилипнуть к смоле и сохраниться в янтаре. Впрочем, кто знает, может быть, рано или поздно и в бирмите будет найден оторванный кусочек их крыла — нашли же там кончик хвоста динозавра...
Источник: Qing Liu, Xiumei Lu, Qingqing Zhang, Jun Chen, Xiaoting Zheng, Weiwei Zhang, Xingyue Liu & Bo Wang. High niche diversity in Mesozoic pollinating lacewings // Nature Communications. 2018. V. 9. Article number: 3793. DOI: 10.1038/s41467-018-06120-5.
Александр Храмов
http://elementy.ru/novosti_nauki/433335/Dvoyniki_babochek_iz...

Скелет диплодока — представителя завропод — в Палеонтологическом музее им. Ю. А. Орлова. Целиком он, к сожалению, не помещается в кадр. Фото Г. Врублевского и Н. Ковалева
Динозавры-завроподы — самые крупные наземные животные всех времен — весили до 80-100 тонн и достигали 40-50 метров в длину. Обобщив все имеющиеся данные, палеонтологи назвали пять основных факторов, сделавших возможным развитие гигантизма у завропод: отказ от пережевывания пищи, быстрое размножение путем откладки множества небольших яиц, быстрый рост, совершенная дыхательная система, снижение интенсивности обмена веществ у взрослых животных по сравнению с молодыми.
Журнал Science опубликовал статью палеонтологов Мартина Зандера (Martin Sander) из Боннского университета и Маркуса Клаусса (Marcus Clauss) из Цюрихского университета, в которой авторы подводят итоги многолетнего изучения гигантских динозавров-завропод. Эти животные по своей массе на порядок превосходили крупнейших наземных млекопитающих, равно как и представителей других групп динозавров (теропод и птицетазовых).
Завроподы появились в конце триасового периода (около 210 млн лет назад). На сегодняшний день палеонтологи описали около 120 родов завропод. Эти растительноядные гиганты доминировали во многих наземных экосистемах начиная с середины юрского периода и до конца мела, то есть около 100 млн лет — вдвое дольше, чем длится период расцвета крупных растительноядных млекопитающих. Таким образом, завроподы были весьма успешной и процветающей группой, а вовсе не «ошибкой эволюции».
Вполне естественно, что ученых, так же как и широкую публику, волнует вопрос: почему завроподы стали такими огромными?
Объяснить гигантизм завропод какими-то внешними причинами не удается, хотя такие попытки делались неоднократно. Например, пытались найти корреляцию между эволюционной динамикой размеров завропод и такими факторами, как концентрация кислорода и углекислого газа в атмосфере, изменения климата, уровня моря и площади суши — и во всех случаях оказывалось, что значимых корреляций нет. Из этого авторы делают вывод, что ключ к тайне гигантизма завропод нужно искать в их биологии.
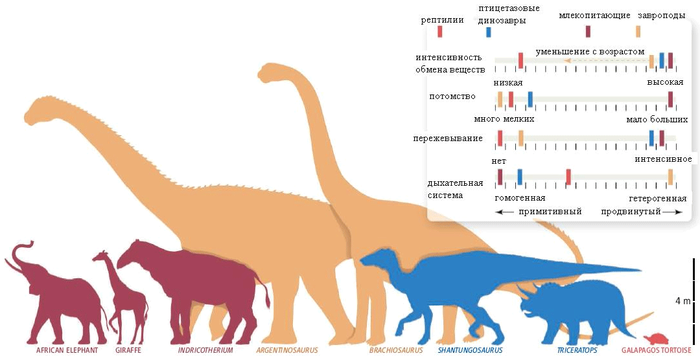
Схема, иллюстрирующая соотношение примитивных и продвинутых характеристик у крупнейших растительноядных животных: млекопитающих, завропод, птицетазовых динозавров и современных рептилий. Слева направо: млекопитающие (африканский слон, жираф, ископаемый носорог индрикотерий), завроподы (аргентинозавр, брахиозавр), птицетазовые динозавры (шантунгозавр, трицератопс), гигантская галапагосская черепаха. Рис. из обсуждаемой статьи в Science
Строение зубов, рта и шеи у завропод довольно разнообразно, что свидетельствует об отсутствии у них какой-то одной раз и навсегда установленной диеты (хотя все они, несомненно, питались только растительной пищей). Однако у всех завропод есть одна примитивная черта, которая резко отличает их от других крупных фитофагов — млекопитающих и птицетазовых динозавров. Завроподы никогда не пережевывали пищу: зубы и челюсти у них были небольшие и совершенно не приспособленные для жевания. Не было у них и привычки глотать камни, чтобы с их помощью перетирать проглоченную растительность в желудке, как это делали некоторые другие динозавры и птицы. По-видимому, отсутствие жевательного аппарата компенсировалось тем, что пищеварительный тракт завропод из-за их чудовищных размеров был очень длинным, так что в нём успевала перевариться даже непережеванная пища.

Череп жираффатитана, обратите внимание на количество отверстий, именно отсутствие жевательных зубов и как следствие отсутствие необходимости прикреплять жевательные мышцы позволило облегчить голову. И череп индрикотерия самого крупного сухопутного млекопитающего. (прим. копипастера)

Отказ от пережевывания пищи позволил голове завропод оставаться маленькой, а это, в свою очередь, сделало возможным развитие очень длинной шеи, благодаря которой завроподы могли, не затрачивая особых усилий, добираться до источников пищи, недоступных другим животным. Предполагается, что длинная шея могла использоваться и для привлечения половых партнеров; может быть, самцы завропод «боролись» своими шеями, подобно современным жирафам. В этом случае удлинению шеи должен был способствовать половой отбор.
У большинства крупных растительноядных млекопитающих и птицетазовых динозавров, напротив, развились очень мощные зубы, челюсти и жевательная мускулатура, что привело к резкому увеличению размеров головы. Это наложило существенные ограничения на допустимую длину шеи.
Крупные размеры тела порождают целый ряд физиологических проблем, важнейшей из которых является проблема перегрева. Кроме того, наличие длинной шеи затрудняет поступление свежего воздуха в легкие (нужно очень глубоко вдохнуть, чтобы в легкие попал не только тот воздух, который остался в длинном дыхательном горле после предыдущего выдоха). По-видимому, завроподы решили обе эти проблемы благодаря развитию очень сложной и совершенной системы «воздушных мешков».
Воздушные мешки, скорее всего, появились еще у общих предков всех ящеротазовых динозавров, то есть завропод и хищных теропод, а от последних были унаследованы птицами. У птиц благодаря воздушным мешкам, соединенным с легкими сложной системой трубок и клапанов, свежий воздух прогоняется через легкие как на вдохе, так и на выдохе. Помимо интенсификации дыхания, воздушные мешки обеспечивают эффективное охлаждение организма. О том, что у ящеротазовых динозавров тоже были воздушные мешки, свидетельствуют результаты изучения их костей (многие воздушные мешки проникают внутрь костей или оставляют на них характерные отпечатки). Новейшие сведения о воздушных мешках у динозавров приведены в только что вышедшей статье: Sereno P.C., Martinez R.N., Wilson J.A., Varricchio D.J., Alcober O.A., et al. (2008) Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina // PLoS ONE 3(9): e3303.
Для того чтобы гигантский размер приносил завроподам реальную пользу и поддерживался отбором, завроподы должны были расти как можно быстрее. Если уж животное сделало ставку на гигантизм — с целью защиты от хищников или для доступа к кронам высоких деревьев — значит, быть маленьким ему невыгодно, и период детства должен быть пройден в кратчайшие сроки. Между тем завроподам, чтобы достичь максимального размера, нужно было увеличить свою массу в 100 000 раз — рекордный показатель для рептилий, не говоря уж о птицах и млекопитающих (вылупившийся детеныш весил всего несколько килограммов, а взрослый матерый динозавр — многие десятки тонн). Гистологический анализ костей завропод в целом подтверждает предположение о быстром росте, хотя эти данные не совсем однозначны. По-видимому, завроподы достигали половой зрелости на втором, а максимального размера — на третьем десятке лет.
Столь быстрый рост возможен только при условии очень интенсивного обмена веществ — такого же, как у современных птиц и млекопитающих. Однако если бы у взрослых завропод интенсивность метаболизма оставалась такой же высокой, они бы неизбежно перегрелись, и никакие воздушные мешки не смогли бы их спасти. Кроме того, им потребовалось бы совершенно невообразимое количество пищи. Авторы видят только один выход из этого противоречия: по-видимому, интенсивность обмена веществ у завропод в молодости была высокой, а с возрастом значительно снижалась. К сожалению, прямых данных, позволяющих судить о скорости обмена веществ у завропод, палеонтологи пока не имеют.
Все динозавры, как известно, были яйцекладущими. Их детеныши появлялись на свет маленькими и незащищенными, зато их было много. Крупные растительноядные млекопитающие, напротив, производят на свет крупных и хорошо защищенных детенышей, но в очень небольшом количестве. Из-за этого у млекопитающих развитие гигантизма резко повышает вероятность вымирания вида: чем больше зверь, тем меньше он производит детенышей, и тем больший срок требуется для восстановления численности популяции после ее временного сокращения. Завроподы не были связаны этим ограничением: гигантизм у них не вел к снижению плодовитости, и численность популяции в благоприятных условиях могла расти очень быстро.

Яйца завропод, найденные в Индии в 2007 году. Фото с сайта news.nationalgeographic.com
Таким образом, по мнению авторов, гигантизм у завропод стал возможным благодаря особому сочетанию примитивных и продвинутых характеристик. Примитивные черты, способствовавшие гигантизму, — это неспособность пережевывать пищу и откладка яиц; продвинутые — быстрый рост, совершенная дыхательная система и меняющаяся с возрастом интенсивность обмена веществ.
Источник: P. Martin Sander, Marcus Clauss. Sauropod Gigantism // Science. 2008. V. 322. P. 200–201.
Александр Марков
http://elementy.ru/novosti_nauki/430876/Chtoby_stat_gigantom...
Мои безграмотные соображения на ту же тему https://pikabu.ru/story/pustogolovyie_zauropodyi_5210896
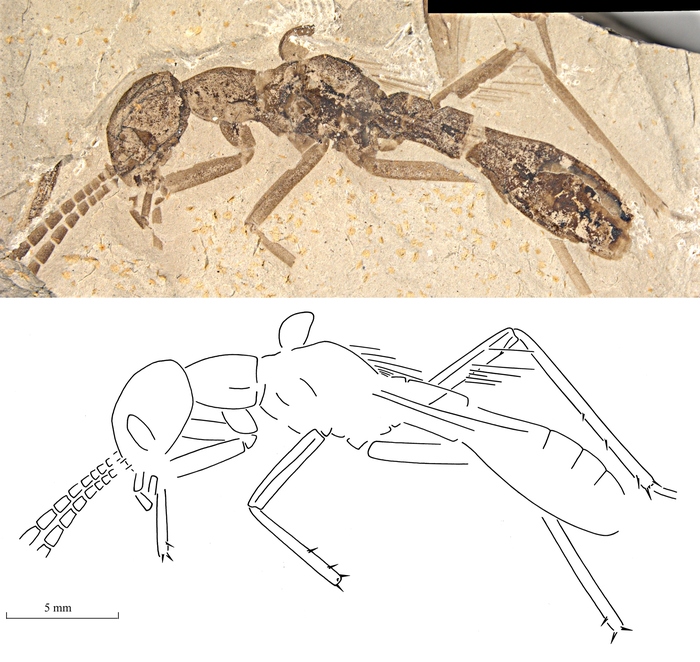
При первом взгляде на это изображение может показаться, что на ней представлен отпечаток какого-то древнего муравья. А вот и нет, перед нами — таракан Grant viridifluvius, найденный в эоценовых отложениях Северной Америки (возрастом около 47 млн лет). Он относится к причудливой группе алиеноптер, представители которой начали притворяться муравьями уже в середине мелового периода. На этом примере можно сделать вывод не только о значении мимикрии в эволюции тараканов, но и о той роли, которую в экосистемах играли муравьи и другие перепончатокрылые, кому алиеноптеры подражали.
В наши дни бейтсовская мимикрия (подражание неопасных организмов хищным или ядовитым) широко распространена среди насекомых. Чего стоят только безобидные мухи или бабочки, которые до мельчайших подробностей копируют облик ос (см. картинку дня Осиная мимикрия). Муравьи в силу своей многочисленности и малосъедобности (мало кто хочет получить порцию муравьиной кислоты или быть укушенным или ужаленным) также очень часто выступают в качестве объектов для подражания. Под муравьев мимикрируют жуки, сверчки, богомолы, палочники, клопы, мухи (есть даже особое семейство — мухи-муравьевидки) и даже пауки.

Паук-скакун Myrmarachne (слева) и жук-быстрянка (справа), подражающие муравьям. Фото © Kurt (Orionmistery) G и © Nicky Bay с сайта flickr.com
А вот среди современных тараканов мы не найдем никого, кто бы копировал муравьев. Мимикрия у тараканов вообще почему-то встречается крайне редко — отдельные виды подражают лишь малосъедобным жукам. Однако новые палеонтологические находки доказывают, что в древности тараканы предавались подражанию другим насекомым куда более охотно. Во всяком случае, это справедливо по отношению к алиеноптерам — особой вымершей группе тараканообразных насекомых. Некоторые исследователи выделяют алиеноптер в отдельный отряд Alienoptera, находящийся где-то между тараканами и богомолами, другие же рассматривают их как семейство Alienopteridae в составе отряда тараканов (Blattoidea). Отличительная особенность алиеноптер — передние крылья, превратившиеся в небольшие кожистые пластинки, которые закрывают основания нормально развитых задних крыльев.

Два года назад уже был описан найденный в бирманском меловом янтаре (возрастом около 100 млн лет) пятимиллиметровый бескрылый монстрик Aethiocarenus burmanicus, похожий на муравья и относящийся, предположительно, к алиеноптерам (см. картинку дня «Чужие» из янтаря). В этом году ученым удалось найти в бирмите еще один вид алиеноптер, подражавший муравьям. Это сантиметровое создание, получившее название Teyia branislav, обладало треугольной головой, удлиненной переднеспинкой и птеростигмами (темными плотными участками на переднем крае крыла). Все эти признаки придавали ему сходство с муравьями из вымершего подсемейства Sphecomyrminae, доминирующих в бирманском янтаре. С другой стороны, церки (членистые отростки) на конце брюшка сразу выдают в T. branislav тараканообразных.

Меловой таракан-алиеноптера Teyia branislav и древнейшие муравьи, которым он подражал. Изображение из статьи P. Vršanský et al., 2018. Batesian insect-insect mimicry-related explosive radiation of ancient alienopterid cockroaches
На бирмит приходятся одни из древнейших находок муравьев, причем их доля от общего числа насекомых там еще крайне мала — в более молодых кайнозойских янтарях муравьев в процентном соотношении гораздо больше. Но, надо полагать, несмотря на свою немногочисленность, уже на заре своей эволюции муравьи были грозной и опасной силой — иначе как объяснить, что алиеноптеры бросились копировать их внешний облик? А вот в том, что муравьям подражала эоценовая алиеноптера Grant viridifluvius, нет ничего удивительного, поскольку в это время современные подсемейства муравьев уже появились и прочно утвердили свои позиции. Как и у остальных алиеноптер, передние крылья у Grant viridifluvius представляют собой небольшие пластинки.
Авторы статьи о Grant viridifluvius и Teyia branislav, приводят описания еще нескольких видов древних алиеноптер, которые, по их утверждению, могли мимикрировать также под ос, пчел и жуков. Однако подражание муравьям, на наш взгляд, удавалось алиеноптерам лучше всего.
Фото и рисунок Даниила Аристова.
Александр Храмов
http://elementy.ru/kartinka_dnya/699/Drevniy_tarakanomuravey
Российский ученый Константин Наговицын представил убедительные доказательства существования разнообразной грибной флоры миллиард лет назад. Фотографии показывают не только грибные мицелии, но и различные стадии жизненного цикла грибов. По ископаемым остаткам удается восстановить облик предков высших грибов, тех, которые сейчас так радуют грибников.

Прототакситы (Prototaxites) — шести-девятиметровые конусовидные сооружения девонского периода, напоминающие окаменевшие хвойные деревья, на самом деле являются гигантскими грибами. Но богатая грибная флора существовала гораздо раньше, не меньше миллиарда лет назад.
Каково было население Земли миллиард лет назад, то есть на границе мезо- и неопротерозоя? Миллиард лет назад жизнь развивалась в мелководных морях, а возможно, и в первичных почвах. Именно в это время происходила так называемая неопротерозойская революция, когда разнообразные сложно устроенные водоросли сместили одноклеточных и нитчатых цианобактерий. Эукариоты внедрились в прокариотические сообщества и заняли господствующее положение среди обитателей древней Земли.
Миллиард лет назад эукариот представляли не только водоросли, но и грибы. В качестве грибов (Phycomyces) того времени сейчас рассматриваются крупные клетки с хвостами, а также нитевидные образования. Нужно заметить, что в протерозойских породах нетрудно найти клеточные ископаемые остатки, микробиоты среднего протерозоя широко распространены по всему миру. Но имея разнообразные остатки, очень трудно различать водоросли и грибы, а порой и минеральные образования, маскирующиеся по форме под микроископаемые. Поэтому основное препятствие для ученого, решившего взяться за протерозойские флоры, связано именно с этим различением.
Константин Наговицын, специалист по низшим растениям из Института нефтегазовой геологии и геофизики им. А. А. Трофимука Сибирского отделения РАН (Новосибирск), выделил грибные остатки, ориентируясь на известные циклы размножения современных грибов. Объектом его исследований стала разнообразная лахандинская биота (восточная Якутия), абсолютный возраст которой составляет 1000–1030 млн лет. Среди явных водорослей найдены и загадочные булавовидные образования на нитчатых ножках — Caudosphera (что в переводе с латыни означает «шарик с хвостом»), вот такие:
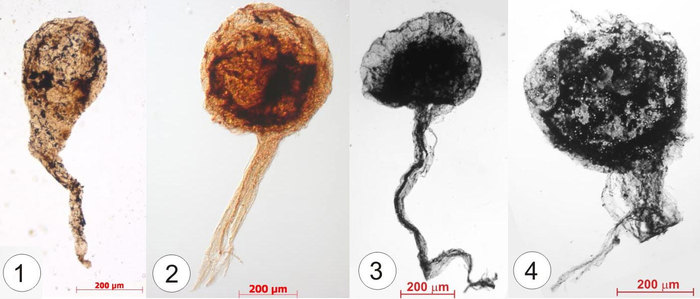
Рис. 1. Загадочные булавовидные образования на нитчатых ножках — Caudosphera. Здесь и «груши» на полых ножках (1), и светлые сферы на ножках из сплетенных нитей (2), и непрозрачные шарики на нитчатых ножках (3) и разрушенные полости (4). Все фото здесь и далее — из цитируемой статьи, публикуются с любезного разрешения К. Наговицына
Они были известны и раньше, но виртуозное владение техникой электронного микроскопирования позволило исследователю заглянуть внутрь плотных булавовидных образований (3). Оказалось, что полость заполнена ячейками с темными спорами внутри.
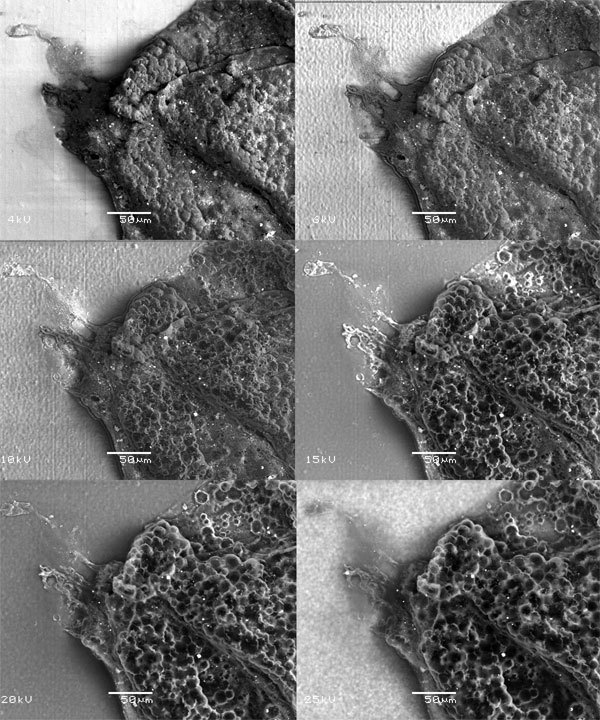
Рис. 2. В этом ряду фотографий электронный луч просвечивает оболочки сферы (3) на разную глубину, то есть слой за слоем постепенно проявляется внутреннее содержимое сферы: округлые камерки с темными спорами
Изображенные четыре формы складываются в последовательные стадии созревания грибного спорангия. Первая и вторая формы — это незрелый спорангий, 3 — спорангий со зрелыми спорами, а 4 — раскрытый спорангий, в котором остались единичные споры. Эти спорангии располагались над поверхностью осадка, как это видно по взаимному расположению ножки и спорангия в пластах окаменевшего осадка. В органических пленках рядом со спорангиями сохраняются и отдельные нити, похожие на те, из которых сложен сам спорангий. Можно представить себе этот древний гриб: крупный мицелиеподобный нитчатый каркас, существующий в органической пленке, из которого вырастали вертикальные спорангиеносцы со спорангиями.
Из современных грибов сходное строение спорангиев имеют высшие грибы, однако у них грибные нити имеют клеточное строение. У грибов из древней толщи клеток не видно. Отсутствие клеточных перегородок характерно для низших грибов. Так, на примере каудосферы почтенного миллиардолетнего возраста можно видеть признаки общего предка высших и низших грибов.
По данным сравнительной геномики предполагается, что линия грибов и животных отделилась от простейших около полутора миллиардов лет назад, а 750 миллионов лет назад от этой ветви отделилась группа амёбоидных простейших (Amoebozoa). Получается, что где-то между этими двумя событиями, но не позже, чем миллиард лет назад, и произошло разделение на высшие и низшие грибы. В это время существовало достаточно много других разнообразных грибов. Константин Наговицын описал, помимо каудосферы, еще и другие морфологические типы грибов.
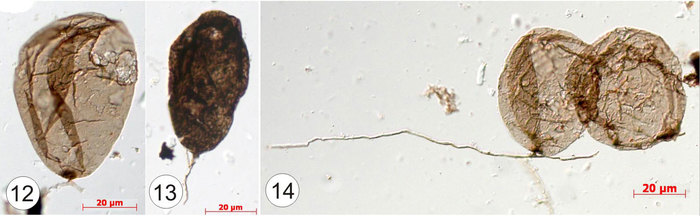
Рис. 3. Похожие на воздушные шарики на ниточках зооспорангии и гифы оомицетов, среди которых попадаются как заполненные спорами (13), так и пустые (12), при этом на одной мицелиальной нити могут располагаться несколько спорангиев (в случае 14 их два)
Мы должны при этом помнить, что оомицеты, по некоторым современным представлениям, вынесены за пределы царства грибов и относятся к полифилетичной группе простейших. Поэтому если в дальнейшем подтвердится оомицетная природа этих остатков, то лахандинская биота станет ареной изучения эволюции простейших.
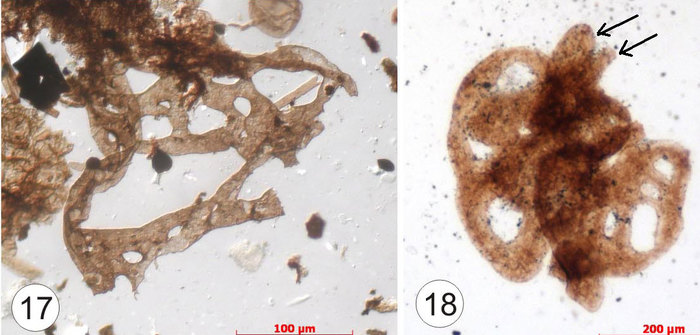
Рис. 4. Встречаются и вот такие ячеистые талломы (cм. также thallus) размером полмиллиметра, распластанные по органической пленке. Но на некоторых экземплярах видны растущие вверх столбчатые образования, как на экземпляре 18 (указаны стрелками). Найдены и начальные стадии развития этого таллома, представляющие собой что-то типа сдвоенных колец восьмерки
Эти разнообразные грибные остатки свидетельствуют о диверсификации грибов не позднее миллиарда лет назад. Позже, в неопротерозое (900 млн лет назад) уже существовали аскомицеты, относимые к высшим грибам. К такому выводу пришли Т. Н. Герман, В. Н. Подковыров, специалисты из Института геологии и геохронологии докембрия РАН (Санкт-Петербург). Им так же, как и для остатков из лахандинской биоты, удалось вычленить стадии жизненного цикла, характерного для грибов аксомицетов. В их случае это были микрофоссилии из мироедихинской биоты (Туруханский р-н, Северо-Западная Сибирь, 850–950 млн лет). Они определили, что цепочки клеток с утолщенными стенками схожи с конидиями аскомицет. Этот вывод подтверждается и разнообразием форм и размеров нитей из собранных в цепочки клеток, которые демонстрируют, вероятно, последовательный ряд созревающих аскоспор.
В заключении мне бы хотелось подчеркнуть, что отсутствие отработанных технологий в распознавании и описании того сложнейшего материала, который дают нам древнейшие осадочные отложения Земли, не останавливают исследователей. Правда, им приходится в каждом случае изобретать новые способы изучения остатков и доказательства своих гипотез. Соединение различных по морфологии форм в единый жизненный цикл вместо описания каждой формы отдельным «видом» может дать новое осмысление эволюционной последовательности и динамики разнообразия у грибов и водорослей.
Источники:
1) К. Е. Наговицин. Биоразнообразие грибов на границе мезо- и неопротерозоя (лахандинская биота, восточная Сибирь) // Новости палеонтологии и стратиграфии. 2008. Том 49. Вып. 10–11. С. 147–151.
2) Т. Н. Герман, В. Н. Подковыров. О природе докембрийских микрофоссилий arctacellularia и glomovertella // Палеонтологический журнал. № 6. Ноябрь–декабрь 2008. С. 81–88.
Елена Наймарк
Источник

На реконструкции изображен ихтиозавр мелового периода, обитавший в Поволжье и получивший название лениния звездная (Leninia stellans), которое принесло животному курьезную известность.
Неполный череп этой рептилии нашли в начале двухтысячных годов сборщики ископаемых из Ульяновска. В твердой конкреции сохранилась средняя часть с громадным склеротическим кольцом в глазнице. Скелета не было. Не было и передней части морды: череп оказался сломан на уровне ноздрей. Сборщики передали остатки в Ульяновский краеведческий музей. Ихтиозавр оказался ранее не известным родом, спустя несколько лет вышло его описание.
Отечественные ученые назвали ихтиозавра в честь В. И. Ленина: в приблизительном переводе «Leninia stellans» означает «Ленин звездный» или «Ленин блистательный». В пояснении указывалось, что череп описали накануне дня рождения Ленина, а кроме того, остатки нашли на родине революционера — в Ульяновской области. Отечественные и западные СМИ подхватили новость про «ихтиозавра-Ленина». В публикациях хватало курьезов. Одна из заметок называлась «Ленин был шестиметровым рыбоящером». В ней говорилось: «Саратовские ученые открыли, что Ленин был рыбоящером, имел очень мелкие и слабые зубы, расположенные по всей длине челюсти, вследствие чего питался некрупной рыбой и головоногими моллюсками». Кто-то из журналистов назвал ихтиозавра «Вождем мировой эволюции». А в блогах предложили выставить кости морской рептилии на Красной площади — в Мавзолее, точнее в Мавзолинии.

Фрагмент черепа ленинии со склеротическим кольцом в глазнице. Фото © П. Годефруа
Спустя пару лет после описания ленинии в Поволжье нашли более полный скелет похожего ихтиозавра. Возможно, это также лениния, и тогда она позволит уточнить морфологию, филогению и палеоэкологию этого животного. Сейчас данных об этом виде мало, хотя в целом ясно, что лениния была консервативной и архаичной рептилией, мало отличавшейся от своих предков юрского периода. Ее можно назвать старомодной.
У ленинии было несколько интересных особенностей. Во-первых, двойные ноздри, разделенные перегородкой, — по сути, четыре ноздри. Такие странные носовые отверстия требовались, чтобы нюхать воду. Дышали ихтиозавры, скорее всего, через пасть. Подобные двойные ноздри были и у некоторых других российских ихтиозавров. По четыре ноздри было у свелтонектеса (Sveltonectes) и симбирскиазавра (Simbirskiasaurus).

Фрагмент черепа ленинии. Фото © П. Годефруа
Во-вторых, очень крупные склеротические кольца в глазницах. Такие костяные образования имелись у большинства морских рептилий, а также у многих динозавров, у экзотических рептилий пермского и триасового периода. Сейчас склеротические кольца есть в глазницах птиц, в том числе сов, куриц и ястребов. Кольца выполняли, вероятнее всего, укрепляющие функции, позволяли глазу выдерживать нагрузки. Это было особенно важно для обитателей моря, которым требовалось защищать глаза от давления водяного столба, волн, плавающего мусора. Есть мнение, что склеротические кольца работали как зум фотоаппарата: пластинки сжимались и раздвигались, фокусируя зрение на дальних или ближних объектах. У ихтиозавров склеротические кольца часто сохранялись в геологической летописи, и на ископаемых останках они выглядят эффектно. Зачастую эти кольца гигантские: у ихтиозавров были одни из самых крупных глаз в истории Земли. Крупнее только глаза гигантских кальмаров. Даже у синего кита глаза меньше.
Хорошее зрение считается одной из характерных черт ихтиозавров. Судя по расчетам, многие ихтиозавры видели в темноте так же хорошо, как кошки. Обычно их крупные глаза объясняют необходимостью охоты на глубоководье. Но более вероятной причиной могла стать ночная тактика охоты.
Это справедливо и для лениний. Они жили в море, где плавало немало крупных головоногих моллюсков: аммонитов-гетероморфов и гигантских белемнитов Volgobelus (от них сохранились огромные внутренние раковины фрагмоконы). Многие современные крупные головоногие совершают суточные миграции. Возможно, меловые моллюски по ночам тоже поднимались к поверхности воды. Ленинии могли охотиться на них, а для этого требовалось хорошее зрение и, соответственно, громадные глаза.
Иллюстрация © Андрей Атучин.
Антон Нелихов
Справились? Тогда попробуйте пройти нашу новую игру на внимательность. Приз — награда в профиль на Пикабу: https://pikabu.ru/link/-oD8sjtmAi


На этой уютной зимней картинке изображен динозавр Ugrunaaluk kuukpikensis, живший на Аляске в конце мелового периода, примерно 70 миллионов лет назад. Он был описан в 2016 году по скелетам, обнаруженным на севере Аляски, близ побережья моря Бофорта. На языке местных эскимосов название рода Ugrunaaluk означает «древнее пасущееся животное».
Перед нами — типичный утконосый динозавр из семейства гадрозаврид: двуногий и травоядный. И, в данном случае, некрупный: судя по приводимым размерам костей, взрослый угрунаалук был всего раза в два крупнее среднего человека. Иные утконосые динозавры достигали и куда больших размеров.
Интересно, что территория, где обитал угрунаалук, находилась в конце мела далеко на севере — гораздо выше Полярного круга (там же она остается и до сих пор). И он не одинок: сейчас известно, что на полярной Аляске была целая динозавровая фауна. Как эти динозавры выживали в условиях полярной ночи и полярной зимы? Такой выход, как впадение в зимнюю спячку, представляется маловероятным из-за их размера (утконосые динозавры бывали и десятиметровыми). Но просто снизить уровень обмена веществ они могли. Есть и гипотеза, что северные динозавры совершали сезонные миграции, подобно современным оленям-карибу. Однако в любом случае для мелового периода был характерен ровный мягкий климат; вероятно, зимняя температура тогда даже в высоких широтах была около нуля, так что растительная пища оставалась доступной. Хотя, конечно, динозаврам всё равно приходилось терпеть дожди, слякоть и снег.
Крайне интересно, что на реконструкции угрунаалука, которую мы видим, он покрыт перьями (или, во всяком случае, какими-то пероподобными покровными образованиями). Есть ли в этом биологический смысл? То, что многие динозавры действительно обладали перьями, сейчас широко известно, и на эту тему постоянно делаются новые открытия (см., например: «Прекрасный король в перьях» — новый динозавр из Китая, «Элементы», 11.04.2012). Более того, пернатые динозавры в наше время вошли в моду (до такой степени, что на сайте DeviantArt появилось целое наглядное антируководство: «Как не надо рисовать пернатого динозавра»). Однако тут надо прежде всего учитывать, что перья были не у всех динозавров. Именно у утконосых динозавров их, как обычно считают, не было — это подтверждается некоторыми отпечатками ископаемой кожи. Автор рисунка — финская художница Майя Карала (Maija Karala) — прекрасно это знает. Но в комментариях она обращает внимание на то, что у нескольких птицетазовых динозавров (отряд, к которому относятся утконосые динозавры) все-таки обнаружены пероподобные структуры, явно образовавшиеся из чешуи. Причем они есть у представителей разных эволюционных ветвей. Тогда можно допустить, что это признак, примитивный в целом для отряда птицетазовых. Почему бы в каких-то эволюционных линиях ему и не сохраниться? Особенно если это могло послужить адаптацией к жизни в холодном климате. Никаких реальных отпечатков перьев (или их аналогов) у угрунаалука на данный момент не найдено, так что перед нами чистая версия художника — но, как видим, достаточно продуманная.
Рисунок © Maija Karala с сайта deviantart.com.
Сергей Ястребов

