В современных морях головоногие моллюски с наружной раковиной представлены всего лишь двумя родами — это Nautilus и более редкий и потому менее известный Allonautilus, оба из семейства наутилид. Но 66 миллионов лет назад, перед великим мел-палеогеновым вымиранием, дела обстояли совсем иначе. Тогда в морях, кроме наутилусов, процветали другие головоногие моллюски с наружной раковиной — аммониты (см. рис. 1). Аммониты были значительно разнообразнее и многочисленнее, чем наутилусы, и они быстрее эволюционировали. Раковины аммонитов с их изогнутыми перегородками в целом были устроены гораздо сложнее, чем раковины наутилид. То же самое можно сказать и о строении тела этих животных: мускульная система аммонитов была значительно более сложной, чем у наутилусов.
Рис. 1. Слева — юрский аммонит Brightia, справа — современный наутилус Allonautilus. Фото Ирины Смуровой
Но многочисленные, разнообразные и сложно организованные аммониты вымерли 66 миллионов лет назад, когда катастрофа уничтожила не только широко известных динозавров, но и большую часть морского планктона и множество другой наземной и морской фауны и флоры. Причины мел-палеогенового вымирания ясны не до конца. Существуют различные гипотезы, начиная от «астероидной зимы» — падения температуры и освещенности из-за облаков пыли, поднятых падением астероида в район полуострова Юкатан, до увеличения кислотности воды в океане (возможно, вызванного тем же астероидом).
Но чем бы ни было вызвано это вымирание, аммониты вымерли, а древние и примитивные наутилусы не только пережили его, но и живут до сих пор. В чем причина такой разницы в судьбе этих головоногих? Конечно, до тех пор, пока в распоряжении палеонтологов не появится машина времени, мы не сможем сказать, что знаем единственно верный и окончательный ответ на этот вопрос. Но между наутилусами и аммонитами было одно очень существенное различие, которое, по мнению многих специалистов, могло сыграть роковую роль в судьбе подкласса аммоноидей. В чем заключалось это различие?
Существенным различием между аммонитами и наутилусами является размер их эмбриональной раковины (той части раковины, которая строилась до выхода из яйца), а следовательно, и размер новорожденных особей. Так как после рождения моллюска строительство раковины почти всегда ненадолго приостанавливается (животному нужно время для адаптации к новой для него среде обитания), эмбриональная часть раковины оказывается отделена от постэмбриональной заметной границей — ее называют «первичным пережимом». Благодаря этому палеонтологи могут судить о размерах эмбриональных раковин вымерших головоногих. Кроме того, у некоторых головоногих, в том числе и у аммонитов, структура эмбриональной части раковины отличается от постэмбриональной части — это также помогает палеонтологам судить о размерах новорожденной молоди аммонитов.
Современные наутилусы появляются из яйца с раковиной диаметром 2–2,5 сантиметра. По меркам моллюсков, это очень крупные размеры для новорожденных. Благодаря пережимам на ископаемых раковинах наутилусов мы знаем, что у мезозойских наутилид размер эмбриональных раковин был примерно таким же, как и сейчас, — от 1,5 до 2,5 см (рис. 2).
Рис. 2. Эмбриональные раковины современных наутилусов. Слева — Allonautilus scrobiculatus, справа вверху крупным планом показана его эмбриональная раковина (ее край отмечен стрелкой). Справа внизу — новорожденный наутилус, вылупившийся из яйца в японском океанариуме в городе Тоба. Вся его раковина сформирована в яйце. Сзади для масштаба рука человека, справа остатки оболочки яйца
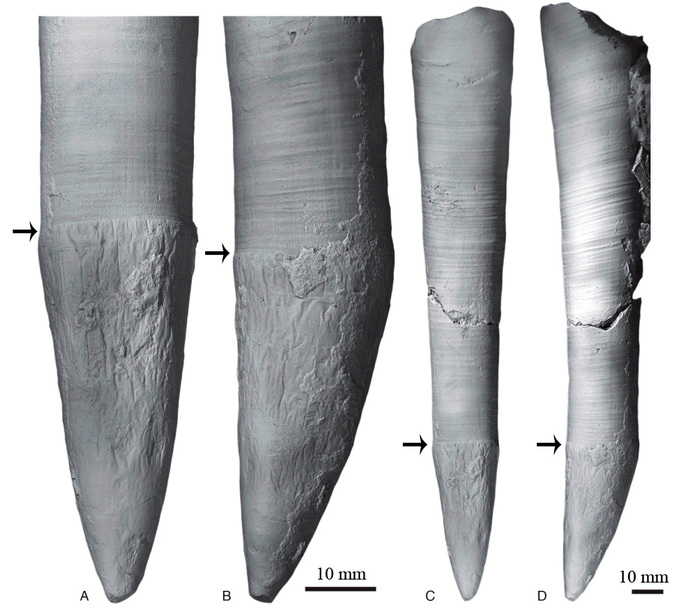
А вот молодь мезозойских аммонитов рождалась с раковинкой в 10–15 раз меньше — диаметром один, максимум два миллиметра (рис 3, 4).
Рис. 3. Раковина юрского аммонита Brightia sp. возрастом около 165 млн лет. Справа крупным планом показана эмбриональная раковина (ее граница отмечена стрелкой), диаметр эмбриональной раковины около 1 мм. Фото Ирины Смуровой
Благодаря своему крупному размеру новорожденные наутилусы могут жить в тех же условиях и питаться той же самой пищей, что и взрослые особи (рыбой, ракообразными, а чаще всего падалью). А новорожденные аммониты с их крошечными раковинами, независимо от образа жизни их родителей, жили в приповерхностных слоях воды и были, фактически, планктоном, а пищей им служили крошечные планктонные организмы.
Рис. 4. Раковина юрского аммонита Binatisphinctes sp. возрастом около 165 млн лет. Справа крупным планом показана эмбриональная раковина (ее граница отмечена стрелкой), диаметр эмбриональной раковины около 1 мм. Фото Ирины Смуровой
Кризис на рубеже мела и палеогена, который привел к вымиранию динозавров и аммонитов, очень сильно ударил по известковому планктону, который вымер практически полностью (см. Что случилось с аммонитами?, «Наука и жизнь», 1985, №6). Причины этого, как уже говорилось в условии задачи, ясны не до конца. Существуют различные гипотезы, начиная от «астероидной зимы» (см. Мел-палеогеновое вымирание) — падения температуры и освещенности из-за облаков пыли, поднятых падением астероида,— до увеличения кислотности (ацидификации) воды в океане (возможно, вызванного тем же астероидом).
Увеличение кислотности воды должно было пагубно сказаться на животных, строящих раковины из карбоната кальция, к которым относятся и головоногие моллюски. И если для крупных животных с относительно толстыми раковинами незначительное подкисление воды было не слишком опасно, то для планктона и крошечных новорожденных аммонитов с очень тонкими стенками раковин оно могло оказаться фатальным. Ведь для аммонита его раковина была и поплавком, позволявшем ему держаться в толще воды, и наружным скелетом, к которому крепились мышцы. А слишком тонкие или недостаточно прочные стенки такой раковины не позволили бы животному ни плавать, ни даже дышать, так как мышцы не имели бы нормальной опоры.
Конечно, ацидификация морской воды на границе мела и палеогена — лишь гипотеза, но вот вымирание планктона в это время — факт. Так или иначе, новорожденные аммониты относились, по сути, к планктону и погибли если не от подкисления воды, то из-за того, что исчезла их пища — тот же самый планктон. Новорожденные наутилусы же, в 10–20 раз более крупные, от планктона совершенно не зависели: они могли питаться той же пищей, что и их родители. Их раковины имели существенно более толстые стенки, и подкисление воды, если оно имело место, было для них не столь уж опасным.
Кроме того, различия в размерах эмбриональных раковин наутилусов и аммонитов привели к существенной разнице в структуре их популяций. Малые размеры эмбриональной раковины и, соответственно, малые размеры яиц позволяют существенно увеличить плодовитость. Возрастание плодовитости со временем «подталкивает» животных к тому, чтобы «выложиться по полной» ради максимально большого числа потомков, пусть даже и ценой истощения своего организма. Мы видим это на примере современных кальмаров, каракатиц и осьминогов, также обладающих миниатюрными яйцами, — они, как правило, размножаются один раз в жизни и после этого погибают от полного истощения. По-видимому, так же размножались и многие (если не все) аммониты: самец погибал сразу после нереста, а самка — выносив потомство (у аммонитов, скорее всего, были яйцеживорождение, см.: A. Mironenko, M. Rogov, 2015. First direct evidence of ammonoid ovoviviparity), зато каждая пара оставляла сотни и тысячи потомков.
Таким образом, у аммонитов, как у современных кальмаров или осьминогов, популяция очень быстро (в течение нескольких лет) полностью обновлялась. Это имело свои выгоды: быстрая смена поколений позволяет очень быстро адаптироваться к меняющимся условиям. Но в случае затяжного кризиса эта быстрая смена поколений могла оказаться «ахиллесовой пятой» аммонитов. Ведь если, к примеру, в один кризисный год зрелые аммониты размножились, но их новорожденное потомство погибло, на следующий год повзрослели и размножились представители следующего поколения, но их потомство тоже погибло, то через год-другой уже и некому будет продолжать род.
У наутилусов же самка откладывает всего по несколько крупных яиц в год, а продолжительность жизни наутилусов превышает 20 лет. Соответственно, в популяции наутилусов всегда есть молодь, есть подростки и есть зрелые особи. И всегда есть какое-то количество яиц (время их инкубации составляет почти год). Даже массовая глобальная гибель всей молоди не может подорвать такую популяцию — останутся взрослые, способные отложить новые яйца, а из ранее отложенных яиц выйдет новая молодь. Даже если бы все новорожденные наутилусы поголовно погибали в течение десяти лет, всё равно сохранились бы способные к размножению взрослые, которые дотянули бы до окончания кризиса.
Так что именно различия в размерах эмбриональных раковин и вызванные этим различия в структуре популяций аммонитов и наутилусов были, судя по всему, причиной столь разной судьбы этих головоногих моллюсков.
Две различных стратегии размножения и существенная разница в размерах эмбриональных раковин у головоногих моллюсков возникли очень давно — еще до появления отряда Nautilida и подкласса Ammonoidea. Древнейшие головоногие моллюски известны с конца кембрия (примерно 495 миллионов лет назад), но какими были эмбриональные раковины их самых первых представителей, мы пока не знаем. Известно только, что к началу следующего периода — ордовикского — у большинства головоногих эмбриональные раковины были крупные, как у современных наутилусов, поэтому считается, что этот вариант был исходным для всех цефалопод (рис. 5).
Рис. 5. Начальная часть раковины наутилоидеи Cameroceras sp. из отряда Endocerida. Длина их эмбриональных раковин (граница которых отмечена стрелочками) превышает 5 см. Фото из статьи: Björn Kröger. The cephalopods of the Boda Limestone, Late Ordovician, of Dalarna, Sweden // European Journal of Taxonomy. 2013. V. 41. P. 1–110
Однако в середине раннего ордовика (примерно 475 миллионов лет назад) появился отряд Orthocerida, у представителей которого эмбриональные раковины стали существенно меньше, и, что важно, появился протоконх — полусферическая начальная камера, служившая поплавком для новорожденного моллюска. Считается, что это позволило ортоцеридам быстро освоить толщу воды, а малый размер яиц позволил увеличить плодовитость. Именно от ортоцерид со временем произошли и аммоноидеи, и колеоидеи (двужаберные), унаследовавшие малый размер эмбриональных раковин своих предков (рис. 6).
Рис. 6. Эмбриональные раковины наутилоидей Bactrites sp. из отряда Bactritida (это потомки ортоцерид и непосредственные предки аммоноидей). Фото из статьи: Royal Mapes & Alexander Nützel. Late Palaeozoic mollusc reproduction: cephalopod egg-laying behavior and gastropod larval palaeobiology // Lethaia. 2009. V. 42. P. 341–356
Эмбриональные раковины ископаемых головоногих моллюсков привлекают внимание специалистов-палеонтологов потому, что позволяют лучше понять образ жизни вымерших цефалопод, их репродуктивные стратегии и структуру популяций. С аммонитами, наутилидами и многими другими древними головоногим исследователям повезло — у них эмбриональная часть раковины отделена от постэмбриональной заметным пережимом. Но не все головоногие легко раскрывают свои тайны. Так, у многих представителей палеозойского отряда Tarphycerida никакой видимой границы между эмбриональной и постэмбриональной частями раковины нет. Почему — непонятно, но получается, что размер эмбриональных раковин (а значит, и яиц) этих моллюсков мы пока измерить не можем.
Некоторые исследователи предлагали использовать для решения этой проблемы следы укусов хищников, которые обычно хорошо заметны на раковинах. Ведь если моллюска укусили, значит он уже не сидел в яйце, а активно плавал в толще воды. Логично? Да, но и тут всё оказалось не так-то просто. Изучение современных и мезозойских наутилусов (у которых первичный пережим есть) показало, что травмы, совершенно неотличимые от нанесенных хищниками повреждений, встречаются и на эмбриональной части раковины (A. Mironenko, 2016. Sublethal injuries and abnormalities on embryonic shells of Recent and Jurassic Nautilida)! Возникают они, видимо, из-за того, что оболочка яиц наутилусов мягкая, кожистая, и если какой-нибудь хищник решит попробовать ее на зуб или крупная рыба просто заденет яйцо, проплывая мимо, то находящаяся внутри формирующаяся раковина может получить повреждение, хотя оболочка не прорвется и молодой наутилус останется жив. Получается, что использовать эти повреждения в качестве индикатора постэмбриональной стадии нельзя.
Так что пока еще далеко не все вопросы, связанные с изучением эмбриональных раковин ископаемых головоногих, решены исследователями.
Источники:
1) Основы палеонтологии (справочник для палеонтологов и геологов СССР). Моллюски — Головоногие. I. Наутилоидеи, эндоцератоидеи, актиноцератоидеи, бактритоидеи, аммоноидеи. Отв. ред. тома В. Е. Руженцев // Издательство Академии наук СССР. Москва, 1962.
2) К. Н. Несис. Головоногие: умные и стремительные // Изд-во «Октопус». Москва, 2005.
3) Neil H. Landman. Exceptionally Well-Preserved Ammonites from the Upper Cretaceous (Turonian-Santonian) of North America: Implications for Ammonite Early Ontogeny // American Museum of Natural History. 1994. No 3086. February 16, 1994.
4) Vladimir Laptikhovsky, Alexander I. Arkhipkin. Impact of ocean acidification on plankton larvae as a cause of mass extinctions in ammonites and belemnites // Neues Jahrbuch für Geologie und Paläontologie — Abhandlungen. October 2012. V. 266(1). P. 39–50. DOI: 10.1127/0077-7749/2012/0268.
http://elementy.ru/problems/1754/Pochemu_ammonity_vymerli_a_...