Краткая справочка: Синтетоцерас - род вымерших парнокопытных из семейства процератид. Обитали в штатах Вайоминг и Небраска в позднем миоцене (10-5 миллиона лет назад). У них было 3 рога. Два рога находились над глазами и напоминали рога современных рогатых млекопитающих. А третий носовой рог был в форме буквы Y и длинный. Только у самцов был такой рог и они, возможно, пользовались им для схваток за самку или за территорию.
Размеры фигурки: - высота скульптуры с подставкой по самую высокую точку - 15,2 см; - длина от рога до хвоста - 12 см; - деревянная подставка в высоту - 1,8 см и в длину - 15 см. Материалы: деревянная подставка, проволочный каркас, фольга, полимерная глина SuperSculpey (Living Doll), матовые акриловые краски, матовый и глянцевый лак, разные элементы имитации растительности.
Коллекция древних животных в моем исполнении пополнилась новым экземпляром. Окрас, кстати, был позаимствован у ныне живущей антилопы - вилорога 🤗💗)))
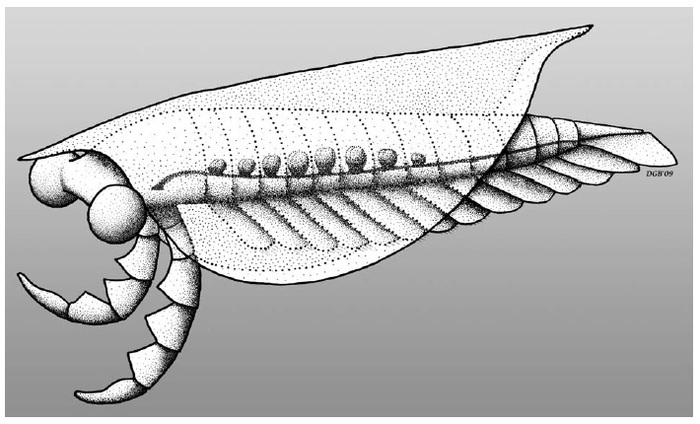
Изоксис (лат. Isoxys) — род членистоногих неопределённого положения, обитавший в середине кембрия. Отличается наличием больших глаз и двух придатков.
Валькотт описал изоксиса в 1890 году на основе образцов, добытых в формации Хилови. Позднее, в 1908 году, на основе образцов из зоны трилобитов горы Стивен Валькотт описал Anomalocaris? acutangulus (Walcott, 1908), однако в 1975 году Симонетта и Делле Кейв определили его к изоксису, заодно описав второй вид из слацев Бёрджесс — I. longissimus.
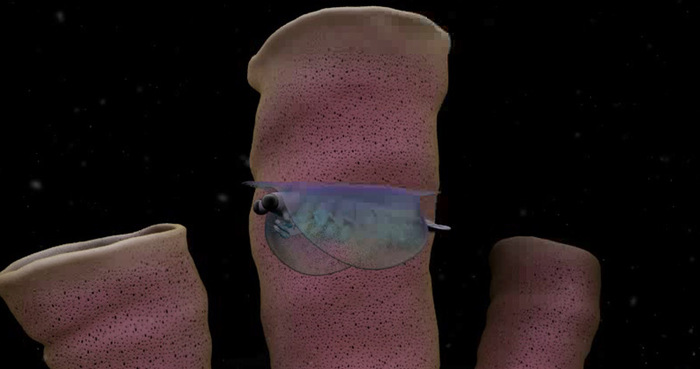
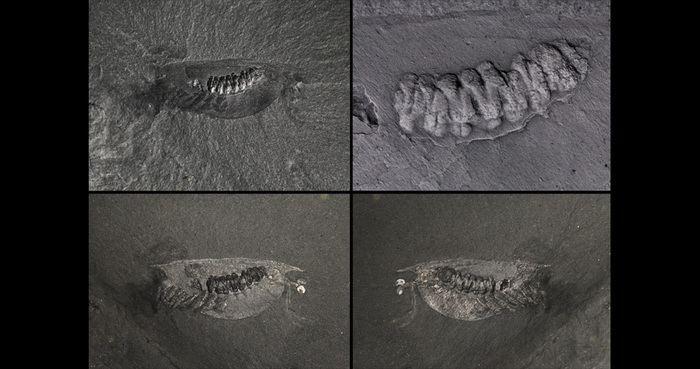
Первое время от изоксиса были известны только панцири, отпечатки мягких тканей удалось найти только в 2009 году.
Изоксис обладал неминерализированным головным щитом с гибкими боковыми плевральными складками, покрывавшими всё тело. Глаза большие, сферические, расположены на стебельках. Передняя пара конечностей большая, с толстыми шипастыми выростами на переднем крае. Данная пара состоит из пяти сегментов, пятый сегмент имеет копьевидную форму. Остальные 13 пар конечностей разделялись на две части, с тонкими несегментированными шагательными ногами и большими длинными лопастями, окаймлёнными щетинками. За конечностями следовал тельсон с боковыми отростками, выступающими за край складок. Панцирь достигал в длину от 1 до 4 см. Спереди и сзади он имел удлинения в виде шипов.
Некоторые окаменелости имеют отпечатки внутренних органов. Так, кишечник был похож на трубку, тянущуюся ото рта к тельсону. Вокруг кишечника располагалось несколько пар пищеварительных желёз, имеющих форму луковицы.
Isoxys acutangulus
Существует огромное количество видов изоксисов. I. volucris наиболее часто встречается в районе Сириус-Пассет, I. auritus был обнаружен в Китае. В районе Большого ложа филлоподов обнаружено 163 образца I. acutangulus и один образец I. longissimus. Благодаря этим находкам удалось установить, что на изоксиса приходилось 0,31 % от всего сообщества данного ложа.
I. chilhoweanus Walcott, 1890 — группа Хиловей, США;
I. acutangulus (Walcott, 1908) — зона трилобитов горы Стивен;
I. bispinatus Cui, 1991 — свита Шуцзиньтоу, Китай;
I. carbonelli Richter & Richter, 1927 — Серро-Морена, Испания;
I. communis Glaessner, 1979 — сланцы Эму в Австралии;
I. curvirostratus Vannier & Chen, 2000 — Маотяньшанские сланцы Китая;
I. glaessneri García−Bellido, Paterson, Edgecombe, Jago, Gehling & Lee, 2009 — сланцы Эму в Австралии
I. globulus Liu et al., 2018
I. jianheensis Liu et al., 2018
I. longissimus Simonetta & Delle Cave, 1975 — карьеры Вальконт, Реймонд и Коллинс на Хребте фоссилий.
I. mackenziensis Kimmig & Pratt 2017
I. minor Luo et al., 2008
I. paradoxus Hou, 1987 — Маотяньшанские сланцы Китая;
I. volucris Williams, Siveter & Peel, 1996 — формация Буэн в Сириус-Пассет, Гренландия.
I. wudingensis Luo & Hu, 2006 — Гуаньшаньская фауна Китая;
I. zhurensis Ivantsov, 1990 — зона Profallotaspis jakutensis в Западной Сибири.
Также есть несколько неописанных видов, обитавших на территории Горы Маккензи, Северо-Западных территорий Канады, формации Игер возле Крэнбурка, а также в районе формации Кайли, Гуйчжоу, и формации Кинзерис, Пенсильвания.
Долгое время систематика данного животного была не ясна из-за того, что чаще всего находили панцири. Лишь только новые находки с отпечатками мягких тканей позволили определить, что изоксис является одним из самых примитивных членистоногих. Он базальнее мегахейров, но производнее радиодонтов. Так же, как и у вышеперечисленных групп, у изоксиса имеются увеличенные придатки.
Изоксис обитал в тропических морях, хотя, возможно, он был распространён повсеместно. Данные животные обитали в разнообразных условиях: например, некоторые представители I. auritus могли вести сумеречный образ жизни на мелководье или обитать на большой глубине, около 140 м, а другие были активны днём при ярком свете.
Изоксис был активным хищником, для которого основной упор в охоте был сделан на глаза, которые обеспечивали обзор в почти 360 градусов. Изоксис мог плавать с помощью своих плавательных ног со специальными лопастями, направляя тело с помощью переднего шипа и похожего на лоскут хвоста или тельсона. Судя по всему, изоксис был придонным охотником.
BRIGGS, D. E. G., B. S. LIEBERMAN, J. R. HENDRICK, S. L. HALGEDAHL AND R. D. JARRARD. 2008. Middle Cambrian arthropods from Utah. Journal of Paleontology, 82: 238—254.
BUDD, G. E. 2002. A palaeontological solution to the arthropod head problem. Nature, 417: 271—275.
CARON, J.-B. AND D. A. JACKSON. 2008. Paleoecology of the Greater Phyllopod Bed community, Burgess Shale. Palaeogeography, Palaeoclimatology, Palaeoecology, 258: 222—256.
CHEN, J. Y., D. WALOSZEK AND A. MAAS. 2004. A new 'great-appendage' arthropod from the Lower Cambrian of China and homology of chelicerate chelicerae and raptorial antero-ventral appendages. Lethaia, 37: 3-20.
EDGECOMBE, G. D. 2010. Arthropod phylogeny: An overview from the perspectives of morphology, molecular data and the fossil record. Arthropod Structure & Development, 39: 74-87.
GARCÍA-BELLIDO, D. C., J. VANNIER AND D. COLLINS. 2009a. Soft-part preservation in two species of the arthropod Isoxys from the middle Cambrian Burgess Shale of British Columbia, Canada. Acta Palaeontologica Polonica, 54: 699—712.
GARCÍA-BELLIDO, D. C., J. R. PATERSON, G. D. EDGECOMBE, J. B. JAGO, J. G. GEHLING AND M. S. Y. LEE. 2009b. The bivavled arthropods Isoxys and Tuzoia with soft-part preservation from the lower Cambrian Emu Bay Shale Lagerstätte (Kangaroo Island, Australia). Palaeontology, 52: 1221—1241.
SIMONETTA, A.M. AND L. DELLE CAVE. 1975. The Cambrian non trilobite arthropods from the Burgess Shale of British Columbia. A study of their comparative morphology, taxonomy and evolutionary significance. Palaeontographia Italica, 69: 1-37.
STEIN, M., J. S. PEEL, D. J. SIVETER AND M. WILLIAMS. 2010. Isoxys (Arthropoda) with preserved soft anatomy from the Sirius Passet Lagerstätte, lower Cambrian of North Greenland. 2010. Lethaia, 43: 258—265.
VANNIER, J. AND J.-Y. CHEN. 2000. The Early Cambrian colonization of pelagic niches exemplified by Isoxys (Arthropoda). Lethaia, 35: 107—120.
VANNIER, J., D. C. GARCÍA-BELLIDO, S. X. HU AND A. L. CHEN. 2009. Arthropod visual predators in the early pelagic ecosystem: evidence from the Burgess Shale and Chengjiang biotas. Proceedings of the Royal Society of London Series B, 276: 2567—2574.
WALCOTT, C. D. 1890. The fauna of the Lower Cambrian or Olenellus Zone. Reports of the U.S. Geological Survey, 10: 509—763.
WALCOTT, C. D. 1908. Mount Stephen rocks and fossils. The Canadian Alpine Journal, 1: 232—248.
WILLIAM, M., D. J. SIVETER AND J. S. PEEL. 1996. Isoxys (Arthropoda) from the early Cambrian Sirius Passet Lagerstätte, North Greenland. Journal of Paleontology, 70: 947—954.
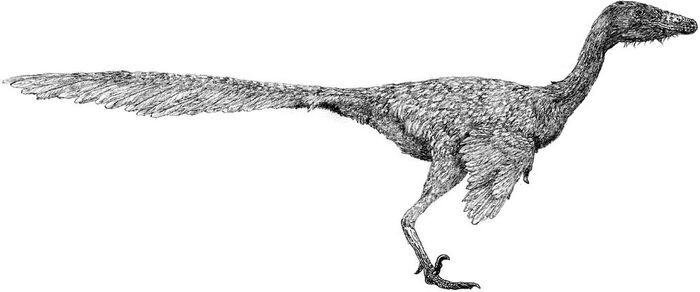
Талос - род североамериканских троодонтид, живший 70 млн лет назад
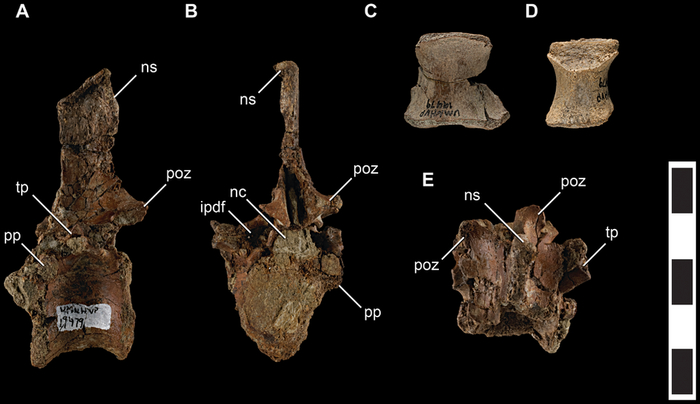
Талос известен только по образцу UMNH VP 19479, частичному посткраниальному скелету субадультной особи, включающему фрагменты передних конечностей, таз, фрагменты позвонков, шевроны и левую локтевую кость. Он был обнаружен в 2008 году М. Дж. Кнеллом во время проекта "Котловина Кайпаровиц", инициированного Университетом штата Юта в 2000 году, в формации Кайпаровиц в Национальном памятнике Гранд Лестница-Эскаланте.
Восстановленная нога
Впервые талос был назван Линдси Э. Занно, Дэвидом Дж. Варриккио, Патриком М. О'Коннором, Аланом Л. Титусом и Майклом Дж. Кнеллом в 2011 году, типовым видом является Talos sampsoni. Родовое название образовано от имени героя древнегреческой мифологии, Талоса, гигантского бронзового витязя, с включением английского слова "talon", "коготь". Видовое название дано в честь палеонтолога доктора Скотта Сэмпсон за изучение и сбор окаменелостей в рамках проекта "Котловина Кайпаровиц".
Позвонки
В длину талос достигал 2 метров, а его вес оценивается в 38 килограмм. Черты животного более элегантны, чем у Troodon, но чёткого представления о облике Talos sampsoni нет в связи с фрагментарностью находки. Вероятно, внешне он был похож на других троодонтид, обладая короткими передними конечностями и длинными ногами. Определенно точно, это был теплокровный двуногий динозавр, покрытый перьями. Особенностью известного образца является частично поврежденный серповидный коготь левой ноги, несущий следы заживления. Авторы описания предполагают, что палец ноги мог быть поврежден в результате какой-либо физической травмы или инфекции. Согласно популярной теории, он был поврежден в результате охоты, хотя это остается спекулятивной идеей.
Реконструкция
В 2011 году талос был определен в семейство троодонтид. Кладистический анализ показал, что он входил в кладу производных троодонтид вместе с Byronosaurus, Saurornithoides, Zanabazar и Troodon.
Talos sampsoni вместе с Troodon — первые представители троодонтид из верхнего мела Северной Америки. В те времена там был тёплый и влажный климат. По описанию нового динозавра учёные предположили, что, вероятно, в те времена там обитало много неописанных видов, близких к троодону. Эта гипотеза связана с очень широким пробелом (20 миллионов лет), к которому можно отнести множество остатков, представляющие собой плохой и фрагментарный материал.
Единственный известный образец талоса был обнаружен в формации Кайпаровиц в южной части штата Юта. Радиоизотопное датирование показывает, что формация Кайпаровиц была отложена между 76,1 и 74,0 млн лет назад, во время кампанского яруса мелового периода. Тогда формация Кайпаровиц была расположена недалеко от западного берега Западного внутреннего моря, большого внутреннего моря, которое разделяло Северную Америку на две части: Ларамидию на западе и Аппалачию на востоке. Местность, на которой жили динозавры, несла на себе древнюю пойму, в которой преобладали крупные каналы и торфяные болота, пруды и озера, и была окаймлена горами. Климат был влажным, и поддерживал богатый и разнообразный спектр организмов. Флора и фауна этой формация является одной из самых изученных в мире.
Космоцератопс отвлекается на проходящего рядом талоса
Талос разделял среду обитания с различными животными, например тероподами из семейства дромеозаврид, овирапторид, орнитомимидами орнитомимами, тираннозавридами тератофонеями, бронированными анкилозавридами, утконосыми гадрозавридами паразауролофами и грипозаврами, цератопсами ютацератопсами, насутоцератопсами, космоцератопсами. Также в формации Кайпаровиц обитало множество амфибий, черепах, ящериц и крокодилов, а также хрящевых рыб (акул и скатов). Было множество ранних млекопитающих, в том числе представителей групп многобугорчатых, сумчатых и насекомоядных.
Сисязавр (лат. Xixiasaurus) — род троодонтид из Китая, обитавших в меловом периоде.
Название Xixiasaurus образовано от «Xixia» (по системе пиньинь так пишется название уезда Сися в Китае) и от греч. σαῦρος «ящерица», что дословно может означать «ящерица из уезда Сися».
На основании окаменелостей ученые отнесли данный род к семейству Troodontidae. При исследовании Xixiasaurus были выявлены несколько признаков характерных для троодонтидов: более высокое количество зубов, чем у большинства других тероподов, в том числе и минирапторов, таких как дромеозавриды; сужение между зубной коронкой и корнем; прорезывание зубов около рострального кончика нижней челюсти и присутствие отличного углубления для нервно-сосудистых отверстий. Тем не менее, Xixiasaurus отличается от большинства представителей семейства, но похож на Byronosaurus jaffei, Mei long, Urbacodon itemirensis и Anchiornis huxleyi в прорезывании зубов, которое является необычным для тероподов, у которых никаких зазубренности не найдены ни на предшествующих, ни на следующих этапах эволюции. Однако это отличие зубов также присутствует у некоторых других представителей, такие как альваресзаврид, Pelecanimimus polyodon и Spinosaurus aegyptiacus имеющие отличительные особенности зубов. Это, очевидно, указывает, что зазубренности были потеряны у нескольких эволюционных ветвей тероподов.
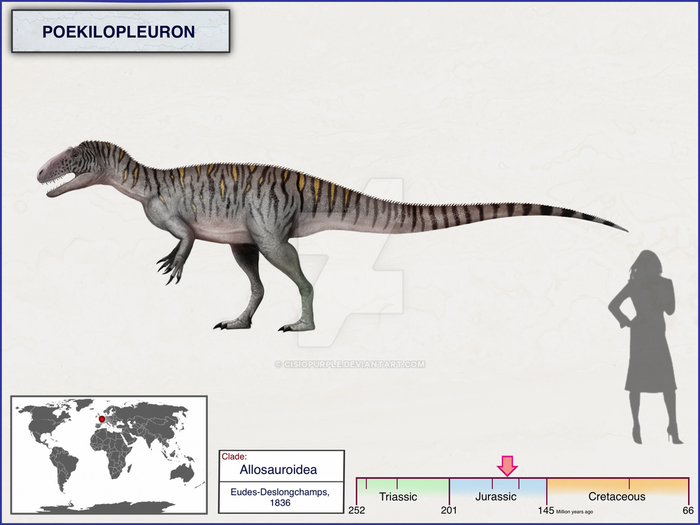
Пойкилоплеврон (лат. Poekilopleuron, что означает «разнообразные рёбра») — род тероподов-динозавров, который жил в средний юрский период (батский ярус), примерно 168–166 миллионов лет назад. Типовой вид — P. bucklandii, названный в честь Уильяма Бакленда, и многие младшие синонимы этого вида также были выделены. В настоящее время известно мало материала, так как голотип был уничтожен во время Второй мировой войны, хотя многие слепки материала всё ещё существуют.
Название Poekilopleuron происходит от греческого др.-греч. ποικίλος — «разнообразный» и πλευρόν — «ребро», из-за трёх типов рёбер. Видовое название Bucklandii дано в честь Уильяма Бакленда.
Окаменелости пойкилоплеврона происходят из среднего бата Нормандии и были описанны Эд-Делонгшаном в 1838 году, через 13 лет после мегалозавра из среднего бата Стоунзфилда в Оксфордшире. Эд-Делонгшан придал особое значение названию «Bucklandii» к его новому роду, чтобы в случае синонимии двух родов, видовое название подавляло только родовое. На основе материала голотипа, который в настоящее время уничтожен. В 1837 году Эд-Делонгшан опубликовал более подробный отчет об этом открытии в монографии, которая также была включена в следующем году в шестой том "Воспоминаний о нормандском обществе Линнея".
Слепок гастролий голотипа, Национальный музей естественной истории
Голотип, хранившийся в Музее факультета естественных наук в Кане, был уничтожен во время Второй Мировой войны, включал гастралии, фаланги, левую переднюю конечность, хвостовые позвонки, шевроны, рёбра и заднюю конечность. Из всего материала сохранилось немногое, хотя гастролиты, фаланги и передние конечности были отлиты и теперь представляют собой пластотип с отливками в Национальном музее естественной истории (образец MNHN 1897-2) и Йельском музее Пибоди (образец YPM 4938). Оригинальный материал, обнаруженный в слое Канского известняка в Нормандии, Франция, можно отнести к среднему батскому ярусу, примерно 167.7 – 166 миллионов лет назад.
В той же публикации 1836 года Эйдс-Делоншан также назвал типовой вид P. bucklandii. Эд-Деслонгшамп отметил сходство с некоторыми материалами из Megalosaurus bucklandii и P. bucklandii и выбрал видовое название bucklandii для поэкилоплеврона, так что, если бы два рода были синонимами, было бы исключено только название рода.
Кау (2024) предположил, что современный дюбрейлозавр, первоначально отнесённый к пойкилоплеврону, может представлять собой младший синоним, а различия в строении скелета объясняются позиционной и индивидуальной вариативностью, как у аллозавра. Таким образом, он представляет собой менее зрелую особь пойкилоплеврона.
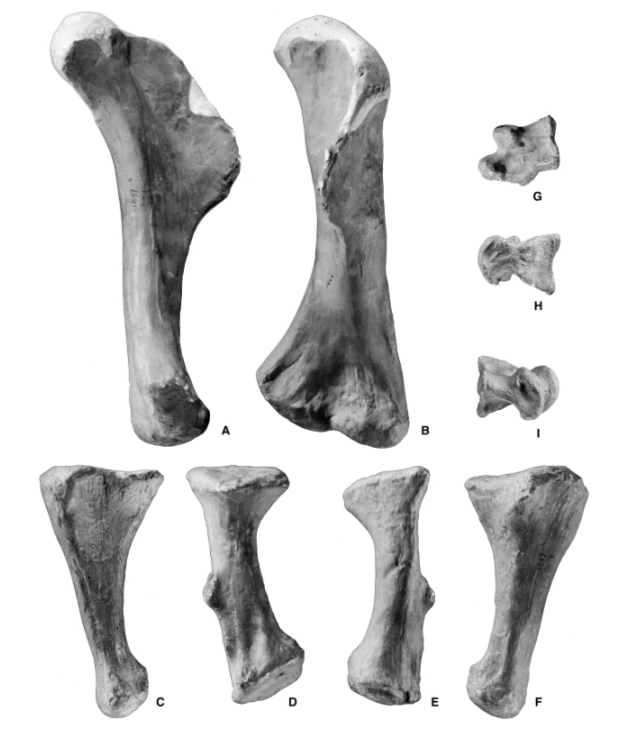
Самой отличительной особенностью пойкилоплеврона были его передние конечности. Их длина, около 60 см, что было уникально. В отличие от более поздних теропод, чьи передние конечности имели тенденцию к уменьшению в длину пропорционально размеру тела, у пойкилоплеврона были длинные, как следствие, мощные передние лапы. Длина в основном заключалась в удлиненной, но мощной мускулистой плечевой кости. Предплечья были заметно более короткими и крепкими, что характерно для немного более позднего и значительно более крупного американского родича торвозавра. Уникальной особенностью является отсутствие отростка на локтевой кости.
Кости передних конечностей
Пойкилоплеврона также сохранил редкий и полный набор гастралий: четырнадцать пар брюшных ребер, которые поддерживали тело.
Пойкилоплеврон имеет сложную историю классификации, поскольку его исходный материал утерян, и известно лишь несколько слепков. Он неоднократно переименовывался в разные виды и роды, большинство из которых теперь являются его младшими синонимами.
Эуд-Делоншан считал, что вполне возможно доказать, что образец принадлежит к ранее названному виду; если это так, то нужно будет лишь изменить родовое название. После 1879 года его часто относили пойкилоплеврона к мегалозавру. Однако выбор Эд-Делонгшана вызвал проблемы, когда Фридрих фон Хюне в 1923 году пришёл к выводу, что это был мегалозавр, но отдельный вид в рамках этого рода. Поскольку оба вида носили один и тот же видовой эпитет «bucklandii», их больше нельзя было различать. Поэтому фон Хюне переименовал вид в «Megalosaurus poekilopleuron».
Другая проблема была вызвана тем, что название было лишь частично латинизировано. На правильном греческом языке оно должно было звучать как «Poikilopleuron», на латыни — «Poecilopleurum». Это побудило более поздних авторов улучшить написание, что привело к появлению таких вариантов, как Poecilopleuron и Poikilopleuron (которые использовались до 2006 года). Однако первоначальное название имеет приоритет и является правильным.
В роде было названо ещё пять видов. В 1869 году Эдвард Дринкер Коуп переименовал «Laelaps gallicus» в «Poekilopleuron gallicum». В 1870 году Джозеф Лейди назвал «Poicilopleuron» valens на основе неполного хвостового позвонка, который, вероятно, принадлежал аллозавру. В 1876 году Ричард Оуэн назвал «Poikilopleuron pusillus», в 1879 году Коуп переименовал его в «Poekilopleuron minor»; в 1887 году Гарри Говиер Сили выделил его в отдельный род: аристозух. В 1883 году Киприянов создал «Poekilopleuron schmidti», видовое название которого дано в честь Фридриха Шмидта, на основе нескольких неопределённых рёбер, возможно ихтиозавра и плюсневой кости завропода. Эта химера является nomen dubium. Гораздо более поздний вид — «Poekilopleuron» valesdunensis, описанный Ронаном Алленом в 2002 году. В 2005 году он был переименован в дюбрейлозавра. Фрагментарные окаменелости из поздней юры и раннего мела Франции и Англии относили к пойкилоплеврону, но это является сомнительным
Реконструция черепа дюбрейлозавра, который может быть младшим синонимом пойкилоплеврона. Автор: Илья Садыков.
Поскольку оригинальная окаменелость была уничтожена, а других останков с тех пор не находили, а также из-за смены названия, вокруг его классификации возникло много споров, которые не могут быть разрешены. Традиционно его относили к мегалозавридам, но некоторые недавние анализы показали, что он относится к синрапторидам (ныне метриакантозавриды); другие результаты показали, что он является представителем мегалозавроид, занимает базальное положение или относится к эустрептоспондилинам. Бенсон и др. (2010) отнесли его и лориньянозавра к синрапторидам. Более раннее исследование показало, что он был примитивным аллозавроидом вне синрапторид. Однако недавний анализ, проведённый Каррано и др. (2012), также использованный в последующих исследованиях тетанур, показал, что он был мегалозавроидом, тесно связанным с афровенатором. Однако авторы этого исследования утверждают, что такая позиция сомнительна, поскольку «с помощью одного дополнительного шага его можно отнести к пятницкизавридам, к другим представителям мегалозаврид или к нескольким возможным представителям аллозавров».
В недавних исследованиях пойкилоплеврон был отнесён к разным группам в аллозавроидах. Кау (2024) отнёс его к метриакантозавридам, а Раугут и его коллеги отнесли его к кархародонтозаврам в 2019 и 2024 годах соответственно.
Материал, относящийся к пойкилоплеврону, включает два хвостовых позвонка, у которых шеврон одного позвонка прирос к центральной части следующего позвонка в результате развития экзостоза. Две фаланги также сохранили патологии. На одной вероятной фаланге видны три низких, неправильных выступа, похожих на экзостозы. На второй вероятной фаланге кисти имеется низкий округлый выступ, напоминающий мозоль. Ральф Молнар счёл наличие трёх патологий у одного индивида «примечательным». Образцы к сожалению могут быть исследованы для определения этиологии патологий из-за их уничтожения во время бомбардировки британцами в 1944 году.
Места нахождения останков: Африка (Лангебаанвег, Эфиопия, Танзания и долина реки Омо).
Цивета Лики является самым крупным известным на данный момент представителем семейства Виверровых. Она достигала размера маленького леопарда, имея при этом рост 59 см и вес около 40 кг.
Внешне напоминает современные азиатские виды. Считается, что этот вид тесно связан с видом Civettictis civetta (африканская цивета).
Микростоникс был крупнее любой современной дикой свиньи (его масса доходила до 300 кг, а длина черепа до 50 см). По строению он в целом напоминал современных представителей семейства свиней, но имел и характерные отличия от них. Самое заметное отличие — у микростоникса были совсем небольшие клыки. Вместо развитых клыков альвеолярные гребни над верхними клыками (особенно у самцов) были очень крупным и выглядели, как пара бугров на морде. Видимо, они были всеядными.
Предполагается, что микростониксы населяли берега рек и переувлажнённые леса.
Микростоникс появился в позднем миоцене в Азии. Он вскоре стал обычным представителем гиппарионовой фауны и за несколько миллионов лет расселился по Евразии от Испании до Китая.
К началу плиоцена ареал микростоникса начинает понемногу сокращаться. Предполагаемые причины — иссушения климата и смена экологической обстановки. Его место занял род кабаны.
Арандаспиды (лат.Arandaspida) — небольшая группа вымерших панцирных бесчелюстных из класса парноноздрёвых (Pteraspidomorpha), которой присваивают ранг от отряда до подкласса. Название образовано от названия австралийского народааранда и др.-греч. ἀσπίς — «щит» (обычное окончание для названий вымерших бесчелюстных).
По внешнему виду арандаспиды были рыбоподобными животными, но имели только один плавник — хвостовой. Достигали длины 35 см. Были покрыты тонким панцирем. Примечательны парностью отверстия для теменного глаза (хотя эти отверстия имеют и другую интерпретацию).
В составе этой группы различают от 2 до 8 видов. Арандаспид сближают с разнощитковыми, а иногда и включают в их состав.
Чаще всего к арандаспидам относят 4 рода, 2 из которых — предположительно, поскольку они известны только по мелким фрагментам. В каждом из этих родов описан только один вид:
Sacabambaspis janvieri Gagnier et al., 1986, известный из Боливии, Аргентины, Австралии и Омана. Назван в честь типового местонахождения — боливийского города Сакабамба. Жил в конце нижнего ордовика (лланвирнский и карадокский век, около 470 млн лет назад). Известен по останкам намного лучшей сохранности, чем остальные арандаспиды, в том числе по практически целым экзоскелетам.
Andinaspis suarezorum Gagnier, 1991 из ордовика или девона Боливии (недалеко от местонахождения Sacabambaspis). Известен по единственному обломку; принадлежность к арандаспидам не доказана.
Arandaspis prionotolepis Ritchie & Gilbert-Tomlinson, 1977 из центральной Австралии (Алис-Спрингс). Назван в честь живущей в этих местах этнической группы. Жил в начале верхнего ордовика (карадокский век, около 450 млн лет назад). Известен по отпечаткам передней части экзоскелета в мелкозернистом песчанике;
Porophoraspis crenulata Ritchie & Gilbert-Tomlinson, 1977 оттуда же. Кроме того, подобные ему окаменелости известны из раннеордовикских пород (аренигский ярус, около 480 млн лет назад), и это самые древние находки арандаспид). Получил название за большие поры на бугорках панциря. Известен только по мелким фрагментам; принадлежность к арандаспидам не доказана.
Кроме того, в родах Sacabambaspis, Arandaspis и Porophoraspis, вероятно, есть как минимум по одному неописанному виду. Не исключено, что к арандаспидам относится и ещё один боливийский вид — Pirchanchaspis rinconensis Erdtmann et al., 2000, но его систематическое положение очень неясное.
Почти целая окаменелость
Арандаспиды имели вытянутое каплевидное тело длиной до 35 см, покрытое хорошо развитым экзоскелетом. Брюшная сторона была более выпуклой, спинная — более плоской. Единственный плавник — хвостовой. Он хорошо изучен только у Sacabambaspis, у которого имел длинную, но узкую центральную лопасть (куда заходила хорда), более короткую спинную и ещё меньшую брюшную.
Глаза находились на самом конце головы. В них были окостенения — склеротические кольца. Между глазами расположены ноздри, а позади — пара отверстий, которые обычно интерпретируются как пинеальные.
Рот арандаспид был на нижней стороне головы. У них было до 10 пар жаберных мешков, которые открывались наружу отдельными отверстиями, а по другим данным — в общие проходы, открывавшиеся одним отверстием с каждой стороны.
Реконструкция Сакабамбасписа
Эндоскелета у арандаспид не было или почти не было, зато был хорошо развит экзоскелет. Переднюю часть тела покрывали две большие костные пластины (спинная и брюшная), а заднюю — сильно вытянутые вертикальные чешуйки. Длина передних пластин составляет около половины длины животного, а толщина очень маленькая (у Arandaspis < 0,1 мм). Они цельные (без следов слияния отдельных элементов) и не имеют признаков роста. Это означает, что они появлялись уже у взрослого животного.
Между спинной и брюшной пластиной с каждой стороны был ряд из 15—20 небольших многоугольных пластинок, между которыми находились жаберные отверстия. Кроме того, много рядов мелких костных пластинок было с нижней стороны рта. На спинном и брюшном щитке, а также на мелких чешуйках видны маленькие, но хорошо развитые каналы боковой линии.
Все элементы экзоскелета образованы аспидином — бесклеточной костью. В них различаются 3 слоя: нижний (пластинчатый), средний (сотоподобный или сетчатый) и верхний (бугорчатый).
Снаружи панцирь был орнаментирован бугорками. Для родов, известных только по обломкам (Andinaspis и Porophoraspis) их форма — главный отличительный признак. У Arandaspis они вытянутые, сужаются к концам и имеют продольный гребень, а у Sacabambaspis, Andinaspis и Porophoraspis в разной мере напоминают дубовые листья, причём у последнего пронизаны большими порами.
Формой и орнаментацией панциря арандаспиды напоминают разнощитковых и астраспид, а чешуйками хвостовой части — анаспид.
Позади глазниц арандаспид, в передней части спинного щита, расположена пара небольших отверстий. Они есть и у Arandaspis, и у Sacabambaspis. Обычно их интерпретируют как отверстия для органов пинеального комплекса — светочувствительной структуры, из которой у многих древних и некоторых современных позвоночных развивается теменной глаз. Таким образом, у арандаспид таких глаз было два: одно отверстие вмещало пинеальный орган, а другое — парапинеальный. Это очень редкая ситуация среди позвоночных (кроме арандаспид, парное или сдвоенное пинеальное отверстие известно только у некоторых ископаемых рыб — ряда плакодерм, палеонисцид, поролепообразных и ранних двоякодышащих). Кроме того, глазоподобность обоих органов пинеального комплекса сохранилась и у большинства миног (хотя пинеального отверстия у них нет).
По другой интерпретации, эти отверстия у арандаспид представляют собой выходы эндолимфатических протоков, которые есть на панцире и некоторых других остракодерм. В пользу этого говорит то, что они расположены дальше назад, чем обычно для пинеального отверстия.
Арандаспиды жили в мелком море. Поскольку они не имели стабилизирующих плавников, их движение, вероятно, было неуклюжим и напоминало движение головастика. Тем не менее они были самыми прогрессивными известными позвоночными своего времени.
Как и для других остракодерм, для арандаспид предполагается придонный образ жизни и питание детритом и микроорганизмами. На это указывает положение рта снизу головы и (как и у других остракодерм) отсутствие челюстей.
Арандаспиды — древнейшие позвоночные, которые известны по довольно полным скелетам (от более древних находили только мелкие обломки панциря или отпечатки бесскелетного тела). Самые древние фрагменты, вероятно, принадлежащие арандаспидам, имеют возраст около 480 млн лет (начало ордовика), а самые молодые — 440 млн лет (конец ордовика). Причина их вымирания неизвестна; возможно, они исчезли из-за оледенения.
В отличие от других ранних остракодерм (живших в основном в водах Лаврентии), арандаспиды известны только из Гондваны (а именно из прибрежных отложений окружавших её морей). Окаменелости этих животных встречаются в Южной Америке (Боливия и Аргентина), Австралии (центр континента) и на Аравийском полуострове (Оман). Эти места были далеки друг от друга и в ордовике, хотя все находились на краю Гондваны.
Гондвана. Находки арандаспид известны с восточного, западного и северо-восточного края.
Окаменелости арандаспид встречаются редко, но к этой группе относятся почти все ордовикские остракодермы Гондваны. В водах этого континента остракодермы были редки и в последующие времена. Кроме арандаспид, оттуда известны только телодонты (очень широко распространённая группа, возникшая, видимо, в водах Лаврентии и к концу ордовика проникшая в моря Гондваны) и питуриаспиды (маленькая эндемичная для Австралии группа из раннего — среднего девона).