


СМИЛОДОНЫ. Часть 1, 2
Очень интересная и подробная статья о смилодонах. Я выложу перваые 2 части. Если администрация не против выложу и остальные.
Смилодоны (Smilodon) — род саблезубых кошек (Machairodontinae), венчающий американское ответвление трибы Smilodontini. Они тяготели к областям с субтропическим и умеренным климатом, населяли полузакрытые местообитания и специализировались в охоте на крупную добычу.
Череп смилодонов значительно видоизменен в связи с сильным развитием клыков верхней челюсти. Он крупный, с коротким и широким лицевым отделом. Носовое отверстие несколько смещено назад по сравнению с обычным для кошек положением. Глазницы относительно меньше, чем у современных кошек, и сориентированы не столь фронтально. Подглазничное отверстие большое. Заглазничный отросток крупный. Лоб слегка выпуклый. Стреловидный и затылочный гребни хорошо развиты. Скуловая дуга в меру широкая и достаточно тонкая — гораздо слабее, чем у современных крупных кошек. Соответственно, менее развитой была и жевательная мускулатура смилодонов. На всем протяжении твердого неба тянется выпуклый костный гребень. Края верхней челюсти имеют хорошо выраженную рифленность. Наблюдается значительное развитие сосцевидного отростка, к которому прикреплялись мышцы, опускающие голову при нанесении удара верхними клыками, что является приспособлением, усиливающим это движение.
Нижняя челюсть невысокая и довольно легкая, с мощным прямоугольным симфизом и небольшими подбородочными выступами. Венечный отросток, на котором заканчивается височная мышца, сильно редуцирован, а ямка для прикрепления жевательной мускулатуры развита незначительно. Нижнечелюстной сустав отнесен дальше венечного отростка. Увеличенный суставный отросток близко прижат к сосцевидному отростку. Для использования сабельных клыков нижняя челюсть могла отводиться от черепа на 120°, так что расстояние между кончиками верхних и нижних клыков у смилодонов не уступало таковому у современного льва, пасть которого открывается на 65°. Нижняя челюсть двигалась строго в вертикальной плоскости.

Сравнительные размеры различных представителей рода Smilodon. Автор — Роман Учитель.
Зубная система весьма своеобразна. Формула зубов составляет I3/i3, C1/c1, P2/p1-2, M1/m1 = 26-28 (для сравнения, у льва — I3/i3, C1/c1, P3/p2, M1/m1 = 30). Как верхние, так и нижние резцы длинные, острые и сильно выдвинутые вперед. Верхние резцы расположены дугообразно, нижние образуют линию. Находящийся за достаточно широкой диастемой верхний клык очень длинный, его корень достигает верхнего края глазницы, а конец при смыкании зубов опускается далеко за край нижней челюсти. Клык сдавлен с боков и саблевидно изогнут. Приближаясь к основанию, его передний край становится округлым. Задний край уже, чем передний, особенно у основания. Покрывающая клык эмаль значительно тоньше, чем на остальных зубах. Она простирается от кончика зуба до линии десны на его заднем крае и от кончика до середины или 2/3 длины на переднем крае и по бокам зуба. Передняя и задняя кромки несут мелкие зазубрины, которые у зрелых особей стираются до гладкой поверхности. Корень клыка составляет до 40 % его общей длины. По способности выдерживать вертикальные нагрузки клык превосходит таковой современных кошек, но хуже противостоит боковым нагрузкам, в результате которых довольно легко ломается. По степени развития верхнего клыка смилодонов можно сравнить лишь с барбурофелисами (Barbourofelis) и тилакосмилами (Thylacosmilus). Нижний клык, напротив, уменьшен в размерах и по форме сходен с нижними резцами, к которым он непосредственно примыкает.
За верхним и нижним клыками расположены длинные диастемы. P2 и p2 отсутствуют. Сохранившиеся щечные зубы уплощены с боков, а их вершины истончены и формируют острую кромку. Она хорошо приспособлена для разрезания мягких тканей, но практически непригодна для дробления массивных костей. P3 небольшой, с хорошо развитыми передним и задним дополнительными выступами. P4 крупный, с редуцированным протоконом и удлиненным эктопарастилем. Маленький p3 с одним или двумя слившимися корнями наличествует у большинства древних особей, но с течением времени все больше редуцируется и к позднему плейстоцену обычно исчезает (например, он присутствует лишь у 6 % выборки S. fatalis с Ранчо Ла-Бреа).
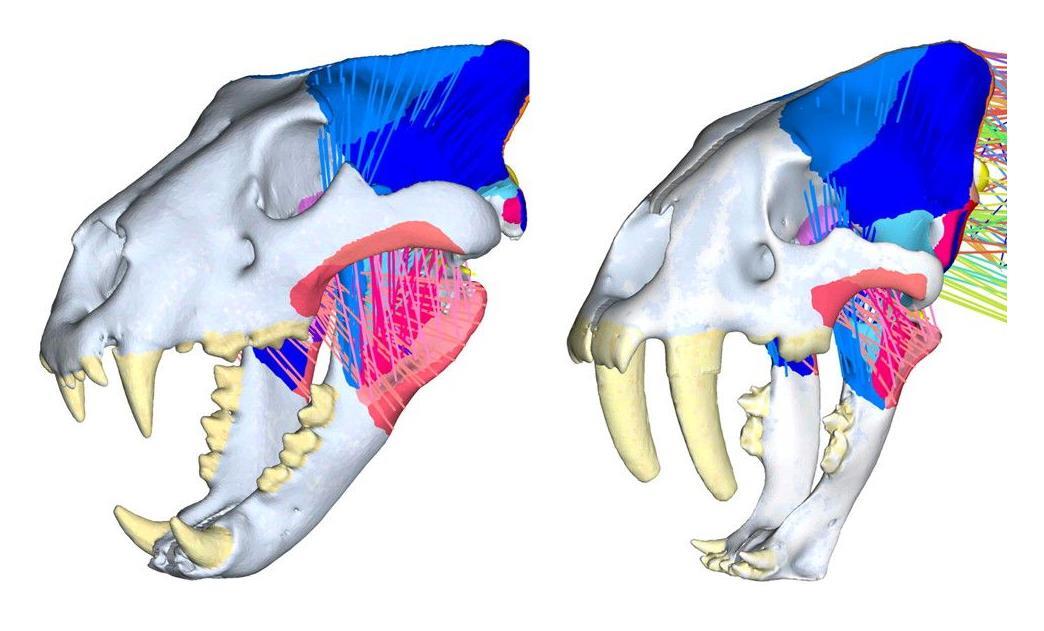
Компьютерные модели черепов и жевательной мускулатуры льва (Panthera leo) (слева) и смилодона (Smilodon fatalis) (справа).
Порядок появления и смены зубов смилодонов отличался от такового у современных кошек. Так, у S. fatalis молочный набор полностью вырастал, когда детенышу было 1-1,5 года. Вскоре после этого начинал прорезываться постоянный набор — в первую очередь, резцы и хищнические зубы, а затем клыки. Когда P4 полностью прорезывался, кончик постоянного верхнего клыка выходил из альвеолы менее чем на 1 см. Молочный клык выпадал тогда, когда идущий ему на смену постоянный клык достигал длины от 6,5 до 8 см (возможно, эта стадия затягивалась до момента, когда постоянный клык достигал 10 см в длину). К этому времени постоянные щечные зубы уже имели легкий износ. Изучение содержания стабильных изотопов кислорода в эмали показывает, что верхний клык рос со скоростью 0,5-0,7 см в месяц. Своей полной длины он достигал примерно через 18 месяцев, в возрасте 3-3,5 лет. Для сравнения, у льва постоянный верхний клык появляется в альвеоле между 9 и 11 месяцами жизни и растет со скоростью 0,13-0,33 см в месяц. Рост клыка заканчивается между 28 и 36 месяцами — примерно в то время, когда лев покидает родительский прайд. Полное окостенение задней части черепа, служащей для прикрепления мышц челюстей, у S. fatalis происходило между 1 и 1,5 годами — на 8 месяцев раньше, чем у льва. Этот возраст совпадает со временем полного формирования молочных зубов. Таким образом, челюстная мускулатура S. fatalis была вполне готова к использованию уже на ранней стадии развития зубного аппарата.
Головной мозг смилодонов соразмерен с мозгом менее крупных леопарда и ягуара. Поверхность его новой коры демонстрирует ряд характерных черт. Центры подавления агрессии выражены слабо, пониженной была и способность к обучению. Однако области мозга, отвечающие за слух, зрение и координацию движений, развиты хорошо. При этом, судя по размерам и расположению глазниц, сами глаза были меньше, а бинокулярность зрения выражалась хуже, чем у ныне живущих крупных кошек. Строение подъязычной кости предполагает возможность произведения громкого рыка, напоминающего их рев. Наличие крупного подглазничного отверстия, пропускавшего пучок нервных волокон, позволяет предположить сильное развитие вибрисс.
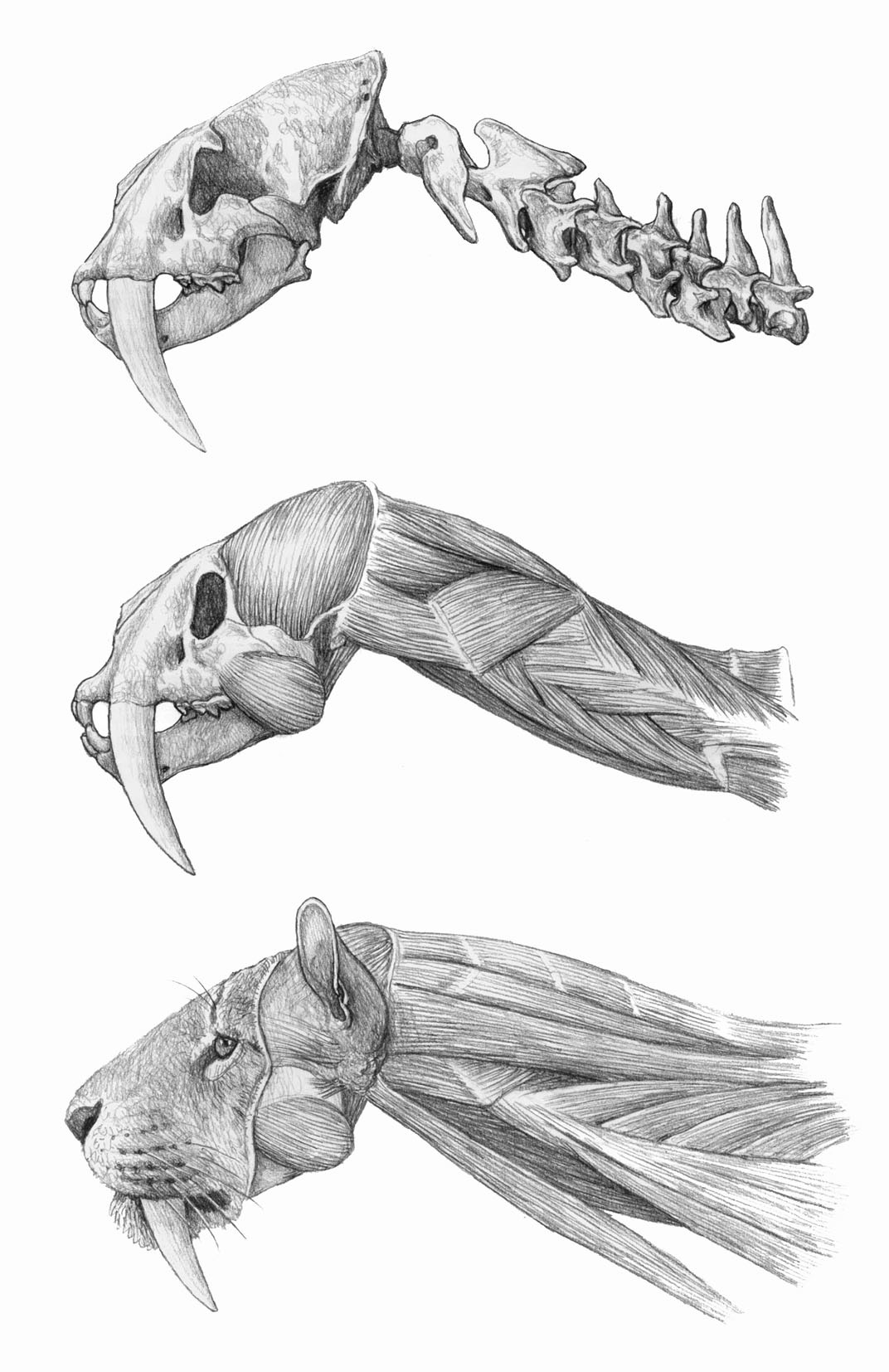
Глубокие мышцы головы и шеи смилодона (Smilodon fatalis). Автор — Маурицио Антон.
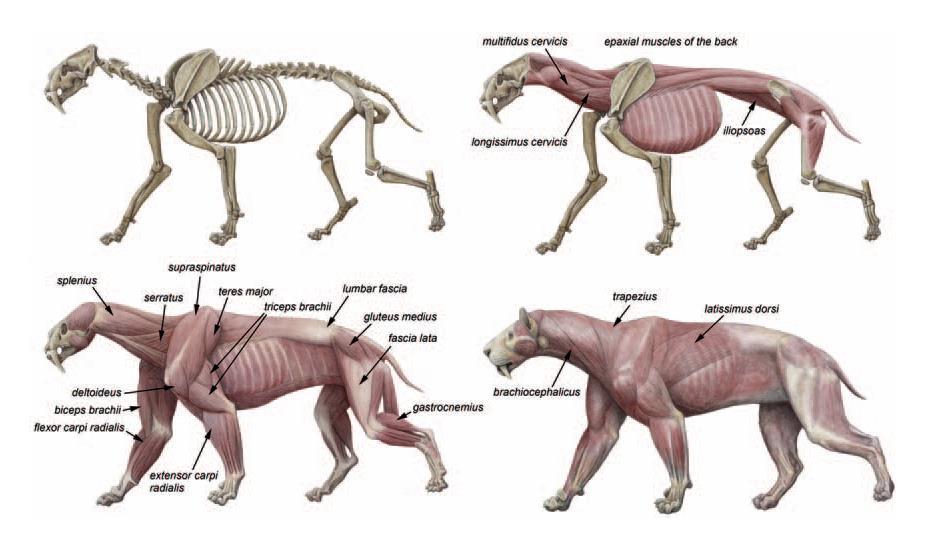
Телосложение смилодонов массивное. Шея относительно длинная, ее позвонки обладают увеличенными отростками для прикрепления хорошо развитых мышц. Эти мышцы были необходимы для совершения сильных и быстрых движений, опускающих голову вниз во время нанесения ударов верхними клыками. Грудина и ребра крепкие. Передняя конечность была развита лучше чем задняя и обладала высокой подвижностью в суставах. Судя по широкому латеральному надмыщелку плечевой кости (к нему крепятся мышцы, разгибающие предплечье), а также по хорошо развитой на этой кости шероховатости для прикрепления круглой мышцы, была усилена функция отведения конечности.
Еще одно важное движение — сгибание конечности — осуществлялось при помощи сильно развитой двуглавой мышцы плеча. Ее усилие увеличивалось за счет комбинированного воздействия двух других: большой круглой и широчайшей мышцы спины. Первая начинается на задней поверхности лопатки, а вторая с позвоночника и поясничной фасции. Они объединяются и прикрепляются к внутренней части плечевой кости. Поскольку задняя часть тела смилодонов короче, чем у современных кошек, сокращение широчайшей мышцы является более эффективным, так как это действие вовлекает объемные массы спинных мышц, лежащих под поясничной фасцией.
Значительной свободой движений отличалось соединение лучевой и локтевой костей, равно как и локтевой сустав. На это указывают форма головки лучевой кости и больший угол наклона локтевой ямки относительно длинной оси кости, уже у S. gracilis достигающий 23-27º и предоставляющий высокую степень свободы медиолатерального вращения в локтевом суставе. Развитая способность передней конечности к совершению супинационно-пронационных движений отражена в латерально расположенной крупной лучевой вырезке, обеспечивающей вращение запястья во время его сгибания. Согласно расчетам, по свободе вращения передней конечностью смилодоны превосходили всех других кошек (род Panthera — примерно на 15 %).
Локтевой отросток смилодонов крупный, с обширной латеральной кромкой. Если его относительная длина находится в середине интервального ряда современных крупных кошек (немного больше, чем у леопарда и тигра и несколько меньше, чем у льва и ягуара), то боковая бугристость выражена сильнее. Это позволяет сделать вывод, что смилодоны имели более развитую трехглавую мышцу плеча, чем ныне живущие кошки.
Плечевая кость смилодонов укреплена за счет сильного развития слоя компактной ткани, благодаря чему она не только толще плечевых костей других кошек, но и более прочная на излом (в обоих направлениях). Высокая прочность и подвижность были нужны для того, чтобы кости и связки передней пары конечностей могли противостоять большим нагрузкам при удержании крупной, сопротивляющейся добычи. Этому способствовала сильно развитая мускулатура плечевого пояса и очень большие (до 10 см в длину) втяжные когти. Кроме того, с целью лучшего противостояния нагрузкам укоротился поясничный отдел позвоночника. Хвост также короткий. В тазе лобковый симфиз укорочен и закруглен, а подвздошная ость удлинена.
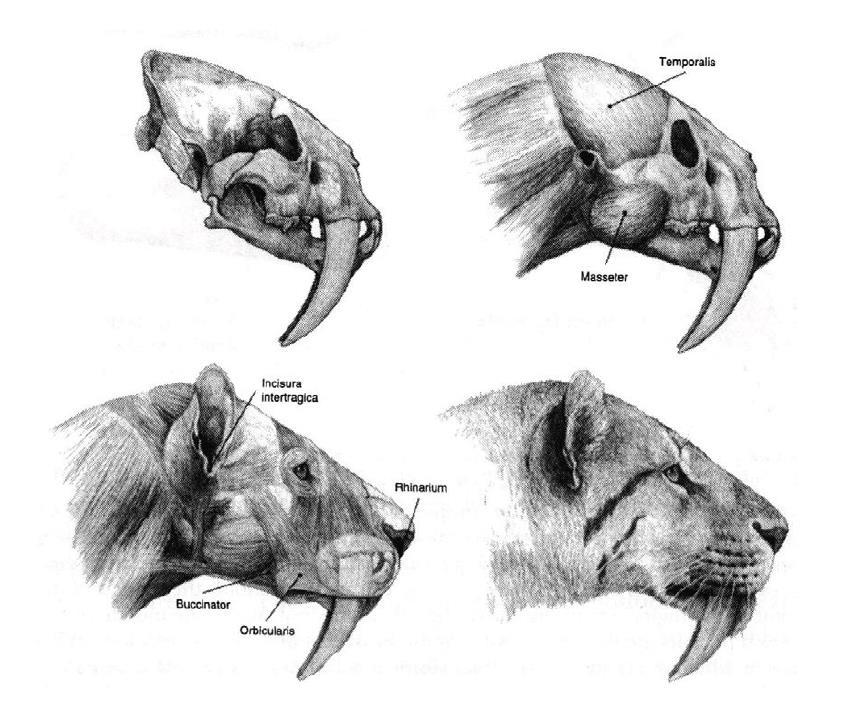
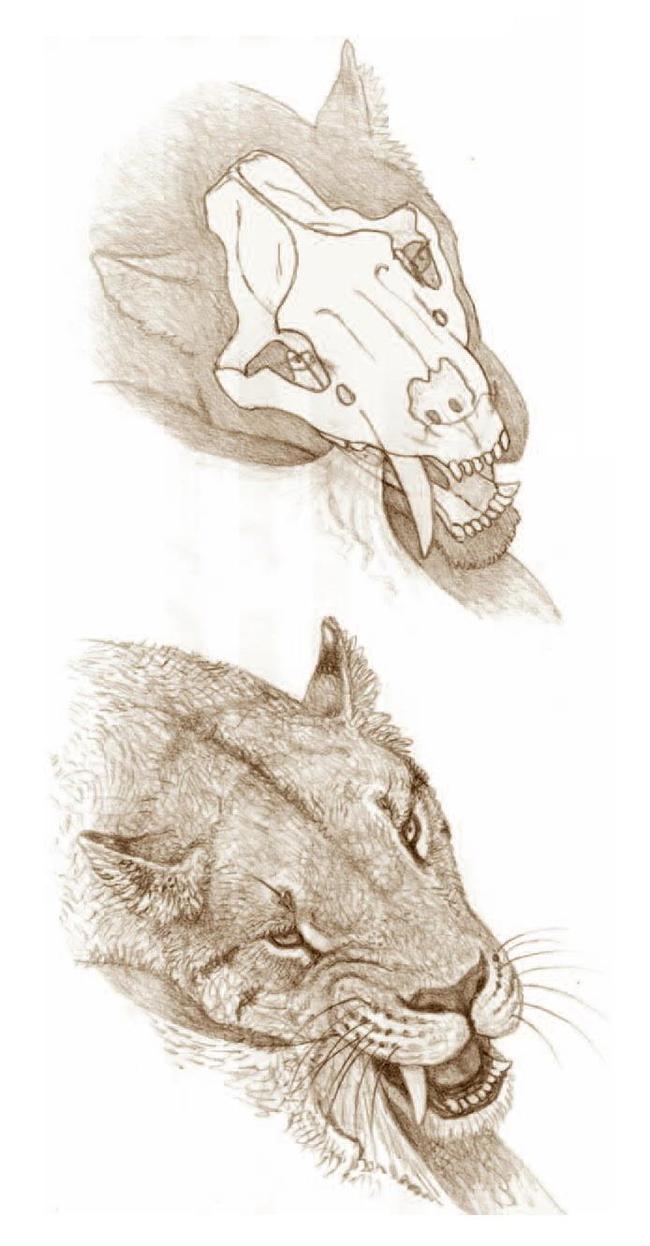
Последовательная реконструкция головы смилодона (Smilodon fatalis) на основе черепа с Ранчо Ла-Бреа. Автор — Маурицио Антон.
Задняя конечность смилодонов демонстрирует ряд «стопоходящих» черт: относительно короткие и толстые трубчатые кости, широкая площадка на большой берцовой кости для прикрепления каудальной большеберцовой мышцы, развитый крупный желобок на коротком пяточном бугре для прохода длинной малоберцовой мышцы и короткие кости плюсны, которые, будучи соединенными, имеют тенденцию развертываться веером. Подобное строение конечности улучшало стабильную позицию хищника, борющегося с добычей. В целом же характерные особенности смилодонов позволяют предположить, что у них происходило смещение акцентов со способности к быстрому бегу в сторону увеличения мощи и подвижности в суставах.
Смилодоны отличаются от мегантереонов (Megantereon) более крупными размерами тела, более массивным скелетом и более наклонной затылочной частью черепа. Нижняя челюсть смилодонов имеет не столь выраженный подбородочный выступ, но сильнее развитые суставный и сосцевидный отростки. Резцы смилодонов крупнее и больше наклонены вперед, клыки более длинные и мощные, а предкоренные зубы редуцированы сильнее. Пропорции посткраниального скелета смилодонов напоминают пропорции гомотериев (Homotherium) высокой лопаткой, мощными передними конечностями и укороченной поясницей. Однако если скелет гомотериев не массивнее скелета современного льва, то для смилодонов, наоборот, характерны очень широкие кости с сильно развитыми гребнями, а также укороченные дистальные отделы конечностей. Так, если у H. serum длина лучевой кости составляет 91 % длины плечевой, то у S. populator это соотношение равно лишь 73 %. Индекс соотношения бедренной кости с большой берцовой у S. fatalis составляет всего 0,75. Наиболее близок к нему Xenosmilus hodsonae (0,79), за которым следуют Homotherium crenatidens и Homotherium serum (0,83-0,84). Затем идут лев и тигр (у обоих 0,86), ягуар (0,87), пума (0,88) и леопард (0,93). Наконец, самый высокий среди кошек индекс отношения бедра к большой берцовой у гепарда ― почти 1,01.
По сравнению с гомотериями, смилодоны имеют более длинную пяточную кость и, несмотря на массивность, могли лучше прыгать, поскольку увеличение длины обеспечивало выигрыш в силе. У S. fatalis пяточная кость составляет 35,1 % длины большой берцовой кости — максимальный для кошек результат. Показатели льва (33,3 %), тигра (32,5 %) и ягуара (31,6 %) находятся вблизи этой границы. Далее следует леопард, пяточная кость которого составляет 29,2 % длины большеберцовой кости. С другой стороны, пума демонстрирует скорее короткую пяточную кость (30,5 %), но обладает при этом очень длинными задними конечностями. У гепарда рассматриваемый показатель равен только 27 %. Таким образом, кажется вполне вероятным, что по способности совершать прыжки смилодоны не уступали представителям рода Panthera.
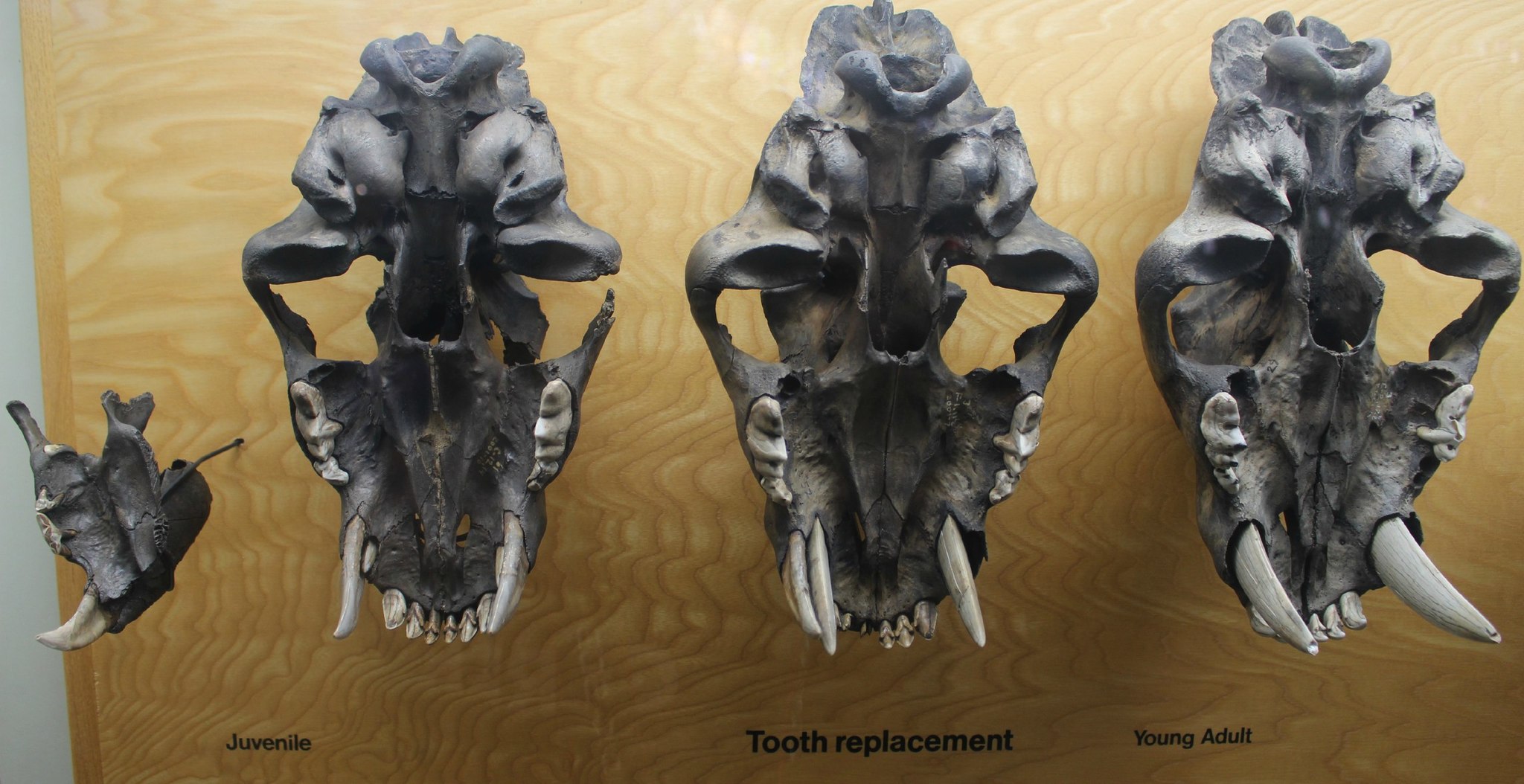
Порядок прорезания зубов у смилодона (Smilodon fatalis) на примере его остатков с Ранчо Ла-Бреа. Музей Пейджа в Лос-Анджелесе, США.

Профили голов кошкообразных хищников с контуром мозга. Вверху слева — гоплофоней (Hoplophoneus), вверху справа — проайлур (Proailurus), внизу слева — пантера (Panthera), внизу справа — смилодон (Smilodon). Автор — Маурицио Антон.
Последовательная реконструкция мускулатуры смилодона (Smilodon populator). Автор — Маурицио Антон.

Сравнение скелетов смилодона (Smilodon populator) (вверху) и льва (Panthera leo) (внизу). Автор — Маурицио Антон.
СМИЛОДОНЫ. Часть 2
Подобно другим крупным кошкам, смилодоны вполне были способны передвигаться рысью и легким галопом, но их повышенная массивность наряду с короткими большой берцовой и плюсневыми костями явно не способствовали бегу на длинные дистанции. Представляется очевидным, что в процессе охоты смилодоны совмещали тактику засады и скрадывания добычи, полагаясь главным образом на короткую стремительную атаку, а отнюдь не на долгое преследование. Приблизившись к потенциальной жертве на достаточно близкое расстояние, смилодон совершал несколько стремительных прыжков и крепко вцеплялся в ее корпус передними когтистыми лапами. Хищник старался свалить на землю и максимально обездвижить пойманное животное, чтобы оно не повредило его длинные и достаточно хрупкие верхние клыки. Затем клыками быстро наносились удары в самые уязвимые места — прежде всего, горло и нижнюю часть шеи, а также, возможно, брюхо ниже реберной дуги и промежность. Можно предположить, что вонзать клыки в другие части сопротивляющейся добычи зверь избегал. Здесь необходимо отметить, что современные крупные кошки часто кусают в горло, морду или даже в круп еще стоящую и отбивающуюся жертву.
Некоторые расчеты показывают, что клыки смилодонов могли проникать в плоть лишь сравнительно неглубоко, нанося основные повреждения за счет обширных поверхностных ран и рассечения подкожных артерий, что приводило к обильной кровопотере. Вероятнее всего, главная охотничья стратегия саблезубых хищников именно в этом и заключалась — нанесение рваных ран, ведущих к быстрому ослаблению жертвы или ее впадению в состояние болевого шока. Возможно, при укусе в горло челюсти производили тянущее движение с вырыванием кровеносных сосудов и трахеи, ускоряющее летальный исход. В то же время, ныне живущие кошки не рвут свою добычу, а убивают ее глубоким проникновением зубов или душат путем сильного сжатия челюстями горла, нередко практически бескровно.

Бегущий смилодон (Smilodon populator). Автор — Маурицио Антон.
При нападении нижняя челюсть отводилась вниз, а тяжелая голова и мощная шейная мускулатура обеспечивали сильный удар клыками по направлению сверху вниз. Длинная шея при этом обеспечивала больший замах. Клыки ныне живущих крупных кошек при определенном захвате деформируют межпозвонковые сочленения, травмируя спинной мозг. Они круглые в поперечном сечении, поэтому могут выдержать приложение силы в любом направлении. В то же время, верхние клыки смилодонов предназначались в основном для манипуляций с мягкими тканями. Их овальная форма обеспечивала минимальное сопротивление при погружении в тело жертвы, тогда как острая задняя кромка рассекала ткани. Сильно рифленые края верхней челюсти могли служить для стока крови, но более вероятной представляется их роль в укреплении черепа, необходимом для противодействия давлению, которому он подвергался во время борьбы. Высокое положение носового отверстия, вероятно, давало возможность зверю дышать, когда голова глубоко погружалась в плоть жертвы. Вероятно, дополнительную информацию об изменении положения тела сопротивляющейся добычи предоставляли хорошо развитые вибриссы.
Обычно считается, что вследствие особенностей строения черепа смилодонов сила смыкания их челюстей являлась значительно меньшей, чем у современных крупных хищников. Согласно некоторым расчетам, укус был примерно втрое слабее львиного. Однако при этом упускается из виду, что у смилодонов не столько нижняя челюсть двигалась навстречу верхней, а наоборот, верхняя двигалась навстречу нижней, выполнявшей роль своего рода ограничителя. По-видимому, в этом случае особо мощная мускулатура, приводящая в движение нижнюю челюсть, была просто не нужна. Естественно, компенсировалось все это за счет мощных шейных мышц.

Последовательная реконструкция головы смилодона (Smilodon fatalis). Автор — Роман Евсеев.
Примерно на той стадии сведения челюстей, когда кончики клыков почти достигали друг друга, из-за малого рычага плеча укороченного венечного отростка и редуцированной жевательной мускулатуры, зверь оказывался неспособным закрыть свою пасть дальше только за счет действия нижнечелюстных мышц, а сама нижняя челюсть занимала положение почти неподвижное относительно тела жертвы. Затем нижняя челюсть фиксировалась при помощи соответствующей мускулатуры и начинала давить хорошо развитой передней частью подбородочных выступов на жертву. Кроме того, нижние клыки могли проникать сквозь кожу, что позволяло лучше захватить мягкие ткани. После того, как некий упор был создан, верхние клыки двигались вниз и проходили нижние клыки благодаря опусканию головы и действию шейной мускулатуры. Согласно другой точке зрения, «якорем» служили верхние клыки, а во время укуса двигались нижняя челюсть. В этом случае достаточная сила укуса при одновременном снижении объема приводящих мышц могла быть достигнута благодаря более прямому расположению их волокон.
Высказано предположение, что после первой атаки хищник отступал и ожидал гибели жертвы, а затем возвращался к павшей добыче. Известно, что домашняя кошка может оставить только что добытое животное на некоторое время. Однако другие ныне живущие кошки редко отказываются от контакта с жертвами после их поимки и, как правило, отступают только в случае прямого нападения на них. Поэтому очень маловероятно, что смилодон рискнул бы оставить пищу другим, находящимся поблизости плотоядным. Скорее всего, зверь продолжал контролировать раненную добычу, удерживая ее лапами и навалившись массой своего тела. По-видимому, зубы для дополнительной фиксации на жертве не использовались из-за высокой опасности их повреждения.
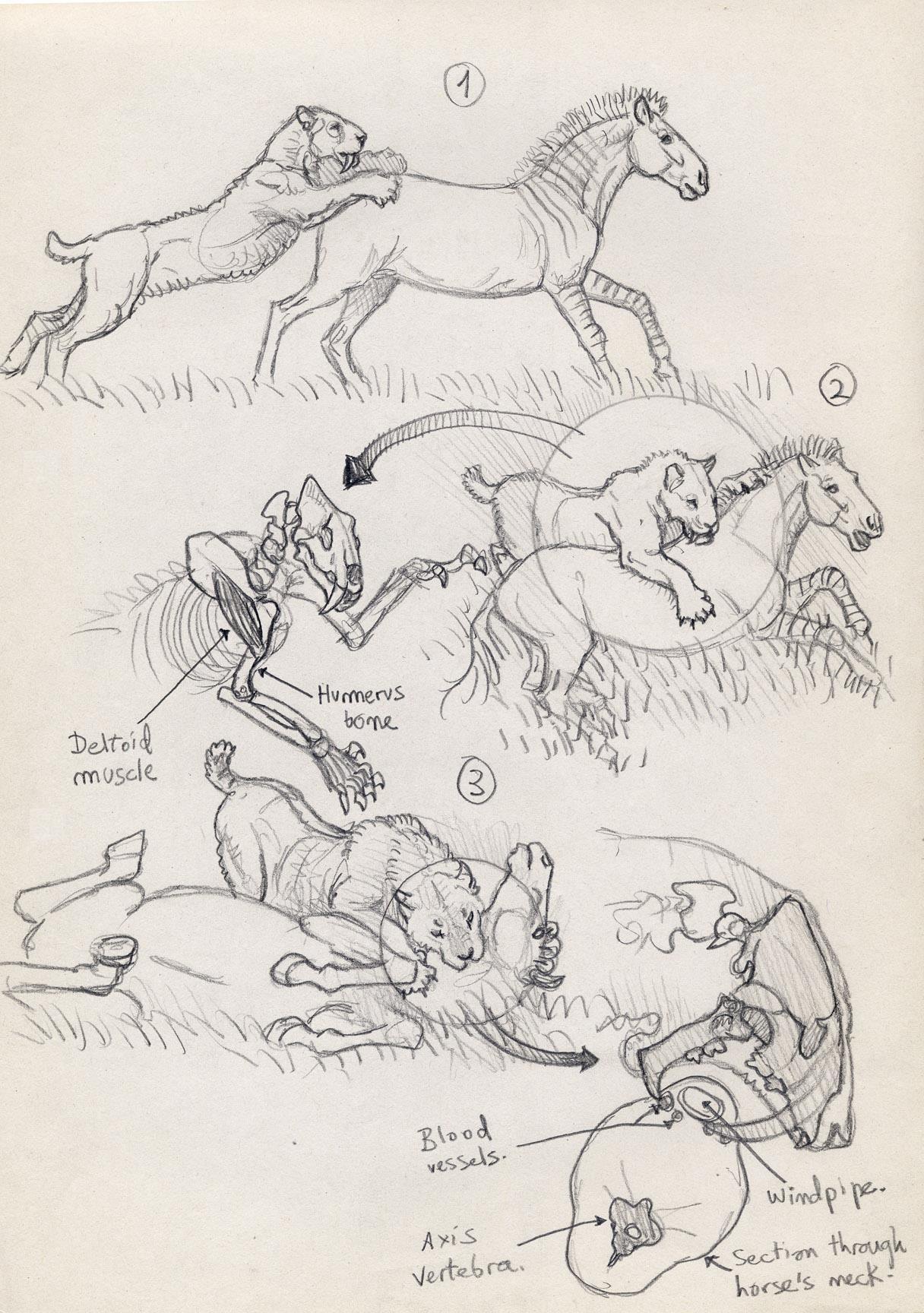
Последовательность нападения и умерщвления смилодоном добычи. Автор — Маурицио Антон.
Другие виды износа клыков, помимо стирания зазубренных граней, как правило, отсутствуют — следовательно, клыки использовались смилодонами в основном для умерщвления добычи. На это косвенно указывает покрывающий их тонкий слой эмали. В свою очередь, то обстоятельство, что клыки находятся на некотором расстоянии за резцовой дугой, означает, что для достижения съедобных частей и отделения кусков плоти применялись большие, вынесенные вперед резцы. При этом строение челюстного сустава позволяло дальше выдвигать нижнюю челюсть для лучшего захвата кусков мяса. Подобно современным крупным кошкам, смилодоны также использовали боковой укус хищнеческими зубами и лижущие движения терки-языка. Саблевидные клыки препятствий для этих действий не создавали, а силы челюстных мышц для для разрезания мяса, связок и хрящей в любом случае было достаточно. Бытует мнение, что смилодоны оставляли на туше много мягких тканей, однако характер зубного износа свидетельствует о том, что в случае необходимости они обгладывали и грызли кости. В любом случае, при обилии добычи заполучить новую мясистую жертву для этих высокспециализированных хищников не составляло особого труда.
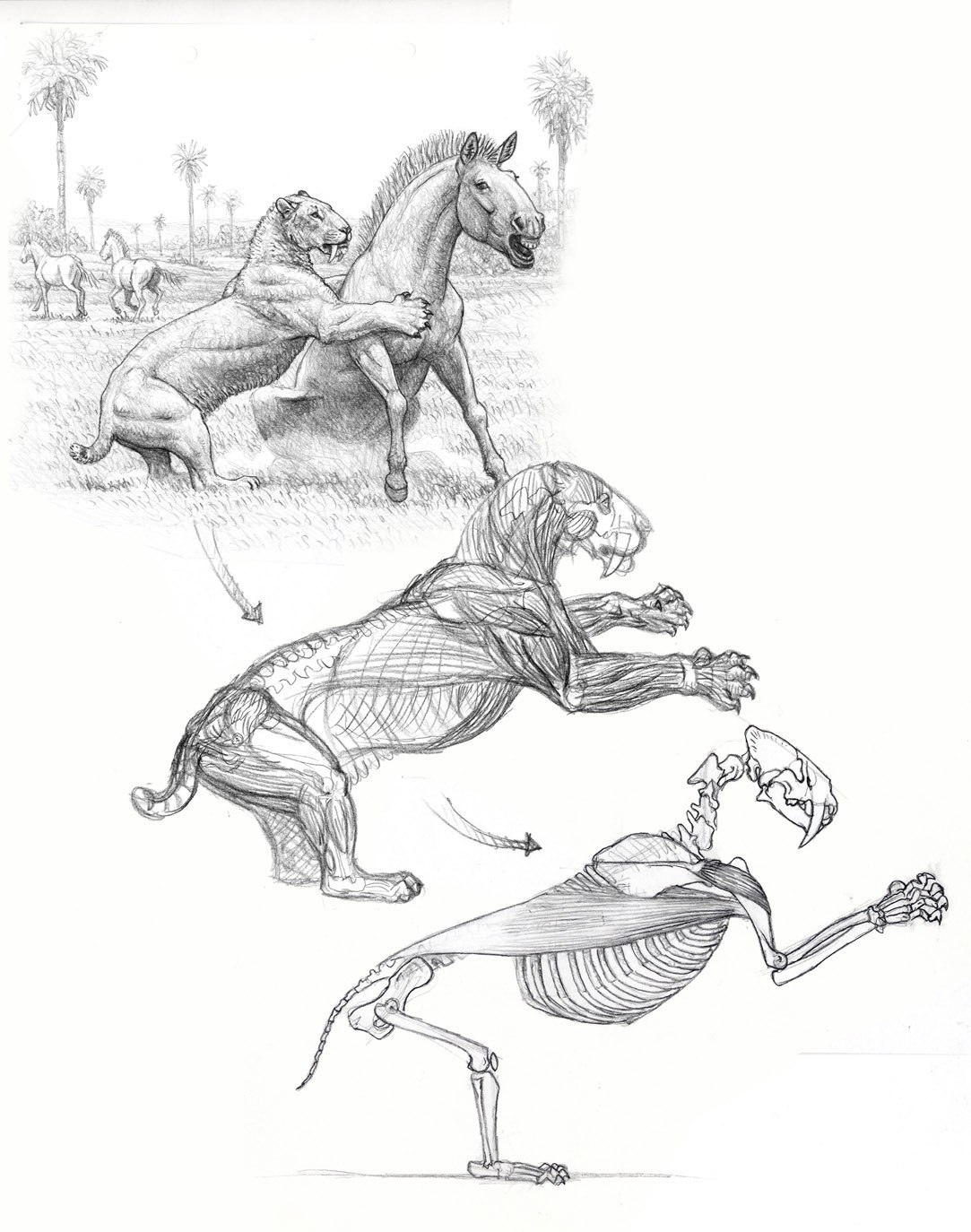
Работа мускулатуры смилодона (Smilodon populator), сваливающего лошадь. Автор — Маурицио Антон.
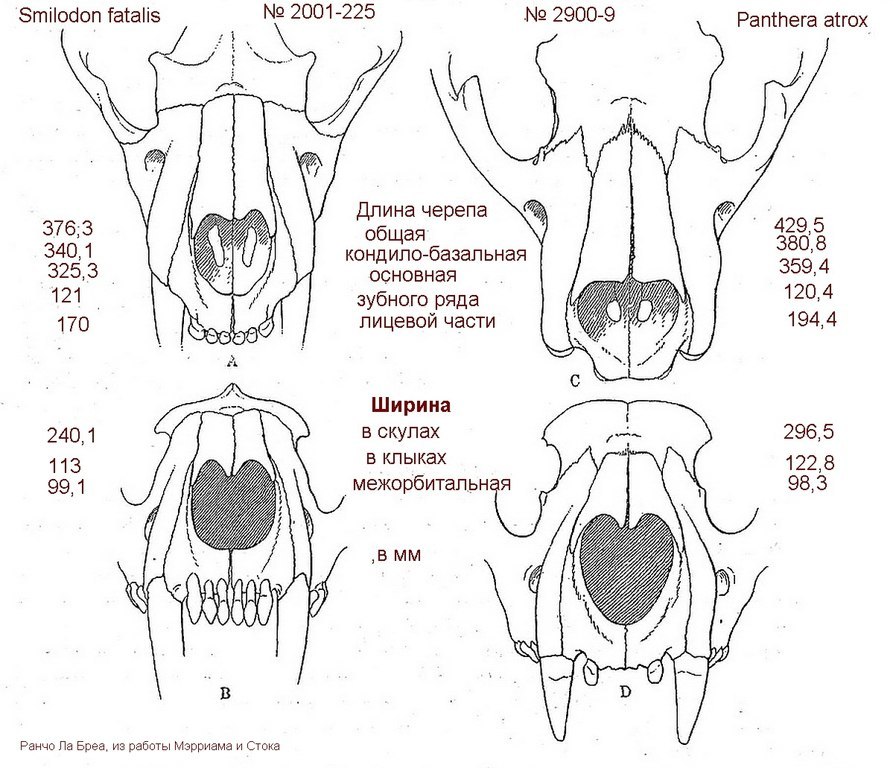
Сравнение передней части черепа смилодона (Smilodon fatalis) и американского льва (Panthera atrox).
Боковой укус смилодона хищнеческими зубами. Автор — Маурицио Антон.

Порядок прикрепления некоторых черепных мышц и строение головного мозга на примере черепов смилодона (Smilodon fatalis) с Ранчо Ла-Бреа. Музей Пейджа в Лос-Анжелесе, США.

Фрагмент когтевой фаланги смилодона (Smilodon cf. populator) и когтевая фаланга ягуара (Panthera onca).
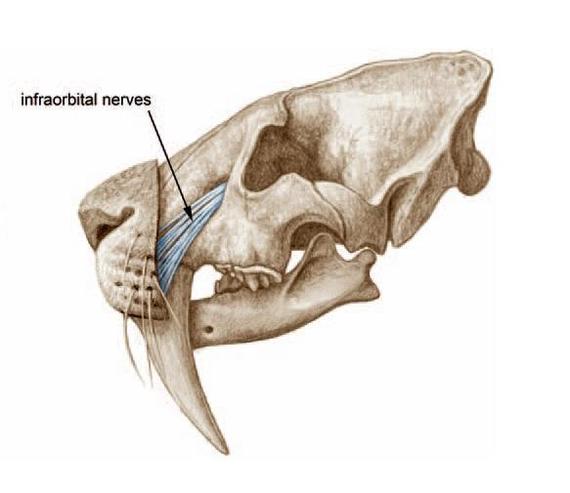
Схема прохождения нервных волокон через подглазничное отверстие к вибриссам у смилодона (Smilodon fatalis). Автор — Маурицио Антон.
Лига Палеонтологии
2.4K постов15.1K подписчиков
Правила сообщества
– Провокации в дискуссиях, излишнюю агрессивность, троллинг, расизм.
– Ничем не подкрепленные нелепые гипотезы, предположения, фальсификации.
– Креационизм, MLP-фэндом – им не место в научном сообществе.
– Оскорбления любой разновидности, Будьте вежливы!
– Поднимание тем политики – для неё есть отдельный раздел.
– Рекламирование чего-либо.
– Многократные нарушения критериев постинга, и игнорирование пометок для исправления.
– Активное отрицание эволюции как факта:
Вы можете признавать Синтетическую Теорию Эволюции или научную парадигму в целом, можете не признавать их, но сама эволюция есть как факт, доказанный экспериментально: опыты Шапошникова и эксперимент с E.Coli,
E.Coli и антибиотик, Методы борщевика, Карта эволюции.
Теория же описывает процесс и механизмы эволюции, и является высшей формой организации научного знания:
Почему СТЭ все ещё теория?
Критерии одобрения постов:
– Посты соответствуют тематике сообщества ⇑
– Посты не должны содержать ненаучную ересь и фальсифицированные факты (Например люди-великаны)
– Тег Копипаста, если статья не ваша, так же желательна ссылка на источник.
– Тег Юмор, если пост несет исключительно юмористическую тему.
– Видеоматериалы обязаны сопровождаться кратким описанием.
– Название постов не должно вводить читателей в заблуждение.