


Палеонтологи уточнили время появления первых членистоногих Часть первая.

Международная группа палеонтологов выпустила свежий обзор данных, касающихся появления в палеонтологической летописи первых представителей самого крупного типа животных — членистоногих. Сразу несколько дисциплин, включая молекулярную филогенетику и тафономию (науку о закономерностях захоронения ископаемых остатков), приводят к выводу, что реальным временем возникновения членистоногих был рубеж эдиакарского и кембрийского периодов, когда произошел так называемый «кембрийский взрыв». Авторы обзора отвергают конкурирующую гипотезу «долгой скрытой докембрийской эволюции», согласно которой главные ветви животных, в том числе и членистоногих, возникли гораздо раньше.
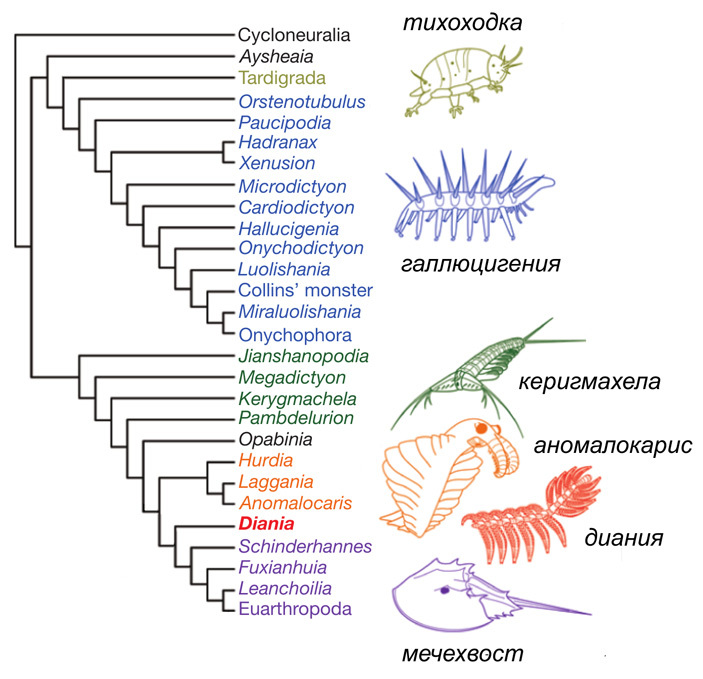
Рис. 1. Эволюционное древо панартропод, сопровожденное изображениями некоторых членов этой группы. Подробные объяснения — в тексте. Детальная структура приведенного древа в данном случае неважна, это просто одна из версий. Среди изображенных животных — два современных (тихоходка и мечехвост) и три ископаемых, из которых галлюцигения (Hallucigenia) близка к онихофорам, а керигмахела (Kerygmachela), аномалокарис (Anomalocaris) и диания (Diania) относятся к обсуждаемым ниже «стволовым эуартроподам». Стоит упомянуть, что керигмахела получила видовое название в честь великого датского философа Сёрена Кьеркегора — Kerygmachela kierkegaardi. Иллюстрация из статьи D. A Legg et al., 2013. Lobopodian phylogeny reanalised, с изменениями
Хозяева Земли
Говорят, что знаменитый английский биолог Джон Холдейн (John Burdon Sanderson Haldane) однажды оказался в компании богословов, и они задали ему вопрос: какие черты Творца обнаруживаются при изучении природы творения? Холдейн, известный своими материалистическими взглядами, проворчал: «Неумеренная увлеченность жуками» (“An inordinate fondness for beetles”; цитируется по статье: G. E. Hutchinson, 1959. Homage to Santa Rosalia or why are there so many kinds of animals?). Этот шуточный ответ был, однако, основан на биологических фактах. Широко известно, что жуки, или жесткокрылые, — это самый многочисленный отряд во всем животном царстве. Если, например, сравнить жуков и млекопитающих, окажется, что число известных на данный момент видов жуков превосходит число видов млекопитающих в 72 раза. И это несмотря на то, что млекопитающие — не отряд, а класс, т. е. группа значительно более высокого уровня.
Сказанное о жуках можно распространить и на их эволюционных родственников — от близких до дальних. Жуки относятся к насекомым, которые, в свою очередь, входят в тип членистоногих. Разнообразие этого типа поистине грандиозно. По современным подсчетам, больше 80% всех видов многоклеточных животных — это членистоногие. Другой настолько успешной эволюционной ветви просто нет в природе. Замечательная книга Эдварда Уилсона (Edward Osborne Wilson), значительная часть которой рассказывает о возникновении социальных систем у членистоногих, недаром называется «Хозяева Земли».
Неудивительно, что эволюция членистоногих занимает многих исследователей. Дело облегчается тем, что палеонтологическая летопись членистоногих сравнительно богата. Более того, ее «чтение» — работа достаточно благодарная. Согласно принципу актуализма, ключом к пониманию прошлого служит настоящее. В частности, любые выводы, касающиеся ископаемых животных, должны быть в конечном счете обязательно основаны на данных о животных современных — иначе исследователю будет просто не на что опереться. В случае с членистоногими этот принцип работает прекрасно, потому что в распоряжение ученых попадает огромное количество как современных, так и ископаемых форм, очень разнообразных, но в то же время устроенных более-менее по одному плану. Это — идеальная ситуация для палеонтолога, и естественно, что исследователи ископаемых членистоногих пользуются ей сполна (см., например: Палеоэнтомология в России).
Членистоногие — группа организмов, очень подходящая для проверки всевозможных гипотез, касающихся хода эволюции. Например, действительно ли большинство современных крупных групп животных возникло во время кембрийского взрыва, или их внезапное появление в начале кембрийского периода связано с образованием твердых скелетов, а истинные эволюционные «корни» лежат гораздо глубже? Попытки выяснить это, основываясь на материале по членистоногим, уже предпринимались (см. Членистоногие подтверждают реальность кембрийского взрыва, «Элементы», 17.11.2013). Но эволюция членистоногих, особенно ранняя, крайне интересна и сама по себе — как история, героями которой являются многие совершенно необычные с нашей современной точки зрения живые существа. Палеонтология XXI века успела сделать много открытий в этой области. Впрочем, и нерешенных вопросов тут, как всегда, еще хватает.
Усики, жвалы и хелицеры
По современным данным, тип членистоногих состоит из двух крупных эволюционных ветвей.
Одна ветвь — хелицеровые (Chelicerata), куда относятся паукообразные, мечехвосты и морские пауки. У них нет усиков, зато есть хелицеры — околоротовые придатки, оканчивающиеся клешнями, крючками или стилетами. Большинство хелицеровых, во всяком случае крупных, — хищники, потому что для других способов питания их ротовой аппарат подходит мало.
Вторая ветвь членистоногих называется жвалоносными (Mandibulata). В их ротовой аппарат входят челюсти — в том числе мандибулы, которые иначе называются жвалами, — представляющие собой не что иное, как сильно видоизмененные ходильные ноги. Этим жвалы принципиально отличаются от хелицер, которые не имеют с ногами ничего общего. Ротовой аппарат жвалоносных, оснащенный набором челюстей, позволяет осваивать предельно разнообразные способы питания, что мы и видим на примере членов этой группы — многоножек, ракообразных и особенно насекомых. Кроме того, для жвалоносных очень характерны усики, или антенны — одна или две пары. Как уже упоминалось, у хелицеровых усиков нет.
Строго говоря, тип, состоящий из жвалоносных и хелицеровых, называется Euarthropoda (эуартроподы, «настоящие членистоногие»). Это название, более точное, чем просто «членистоногие», предложил в 1904 году знаменитый английский зоолог Рэй Ланкестер (Sir Edwin Ray Lankester). Среди признаков эуартропод — четкое деление тела на отделы (тагмы) и присутствие членистых конечностей, внутри которых есть суставы.
Животные, у которых членистых конечностей нет и никогда не было, не относятся к эуартроподам, даже если по многим другим признакам они к ним близки. Существует две группы таких животных: онихофоры — наземные существа, которых иногда называют «бархатными червями» (velvet worms), — и тихоходки, мельчайшие водяные жители (см., например: Укорочение тела тихоходок связано с потерей Hox-генов, «Элементы», 04.03.2016). И у онихофор, и у тихоходок тело сегментировано почти так же, как у членистоногих, но конечности нечленистые. Поэтому диагнозу типа Euarthropoda, равно как и буквальному значению слова «членистоногие», они не соответствуют.
Тем не менее онихофоры и тихоходки, безусловно, являются близкими родственниками эуартропод. В конце XX века эуартропод, онихофор и тихоходок формально объединили в надтип Panarthropoda (букв. «все членистоногие»). Панартроподы — огромная эволюционная ветвь, которая, в свою очередь, входит в группу линяющих животных (Ecdysozoa; см., например: Китайские палеонтологи нашли древнейшего головохоботного червя, «Элементы», 04.06.2014). Таково положение членистоногих в системе животного мира.
«Стволовые эуартроподы»
До сих пор, говоря о членистоногих и их родственниках, мы упоминали только современные группы. Однако для полного понимания эволюционных событий, сформировавших самый многочисленный тип животного царства, просто необходимо учитывать палеонтологическую летопись, и прежде всего — летопись кембрийского периода, когда сформировались почти все крупные эволюционные ветви животных. В кембрии встречаются интереснейшие живые существа, совершенно не похожие на своих современных родственников (многих из них нельзя отнести ни к какому современному типу — по крайней мере, без явных натяжек). Трудность в том, что «мягкотелые» существа, у которых нет минеральных скелетов или раковин, сохраняются в ископаемом состоянии относительно редко, и к членистоногим это, увы, тоже относится. Для их захоронения нужны лагерштетты — глинистые осадочные породы, образующиеся в малокислородных условиях (там, где кислорода много, в грунт проникают роющие животные, которые сразу нарушают структуру формирующегося осадка; см. А. Ю. Журавлев, 2014. Ранняя история Metazoa — взгляд палеонтолога). Особой известностью в качестве источника кембрийских ископаемых, заключенных в лагерштеттах, пользуются канадские сланцы Бёрджесс (Burgess Shale), где раскопки ведутся уже больше ста лет. Фауна сланцев Бёрджесс настолько знаменита, что местонахождения с подобным типом сохранности обозначаются в научных статьях аббреавиатурой BST — Burgess Shale-type, «тип сланцев Бёрджесс». Палеонтология в наши дни развивается быстро, и местонахождений BST известно много, причем в некоторых из них степень сохранности просто поразительна. Это позволило неплохо изучить древнейших родственников членистоногих и, что называется, пролить свет на их происхождение. Правда, открывшуюся картину никак не назовешь простой. Но тем она интереснее.
Бросим взгляд на эволюционное древо (рис. 2). По молекулярным данным, которые в данном случае неплохо согласуются с палеонтологическими, ближайшие «внешние» родственники эуартропод, дожившие до наших дней — это онихофоры. В палеонтологической летописи онихофоры — или, во всяком случае, близкие к ним формы — тоже присутствуют. Правда, в кембрийском периоде они были еще не наземными, а морскими (см. Aysheaia). Это гусеницеобразные существа, которые передвигались по дну на множестве коротких ножек, оканчивающихся коготками. Вполне вероятно, что общие предки эуартропод и онихофор были на них похожи.
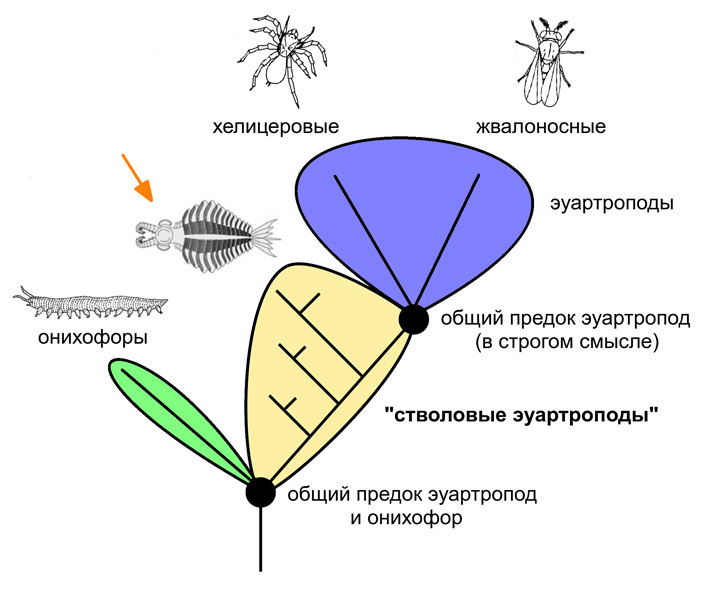
Рис. 2. Упрощенное эволюционное древо членистоногих и их родственников. Тихоходки, положение которых до сих пор остается спорным и которые в любом случае представляют собой крайне специализированную боковую ветвь, сюда не включены. Оранжевая стрелка указывает на представителя «стволовых эуартропод». У него можно видеть плавательные выросты на туловище, фасеточные глаза и длинные, мощные членистые околоротовые придатки (об этих признаках см. ниже в тексте)
Однако онихофоры вряд ли являются непосредственными предками эуартропод (во всяком случае, эуартропод в строгом и традиционном смысле этого слова). Вот тут-то и начинается самое интересное. На реальном эволюционном древе между ветвью онихофор и ветвью эуартропод находится целая совокупность весьма оригинальных полностью вымерших ветвей, члены которых обладали отдельными «эуартроподными» признаками, но еще не набрали их полного комплекса, какой мы видим у паука, сороконожки или мухи. Эти ветви можно было бы назвать переходными, но надо помнить, что для своей среды и своего времени каждое относящееся к ним животное было вполне завершенным организмом, отлично вписанным в определенную экологическую нишу (иначе бы оно не попало в палеонтологическую летопись). Хотя, конечно, с нашей современной точки зрения многие члены этих «переходных» ветвей выглядят крайне странно.
В современной научной литературе упомянутую совокупность эволюционных ветвей принято называть «стволовой группой эуартропод», или просто «стволовыми эуартроподами» (stem-Euarthropoda). Честно говоря, такое обозначение может изрядно запутать: ведь в том-то и дело, что животные, на которых оно распространяется, к эуартроподам в узком смысле этого слова не относятся (см. J. Ortega‐Hernandez, 2014. Making sense of ‘lower’and ‘upper’ stem‐group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848). Однако — тут нам поневоле придется употребить предложение, насыщенное терминами — кладистическая систематика, господствующая в современной биологии почти безраздельно, категорически запрещает вводить парафилетические таксоны. Если говорить попросту, это означает, что группа организмов, охватывающая не единственную эволюционную ветвь, а целый эволюционный уровень, не может считаться единицей классификации и иметь собственное научное название (в частности, латинское). Поскольку в данном случае такая группа налицо и как-то называть ее все же надо, исследователи используют временное обозначение, которое было сочтено корректным по формальным соображениям. Мы тоже — куда деваться — будем этим обозначением пользоваться, оставляя его в кавычках и помня, что с точки зрения классической зоологии «стволовые эуартроподы» — это не эуартроподы.
Положение «стволовых эуартропод» на эволюционном древе показано на рис. 2. Если на время оставить в стороне строгую терминологию, можно сказать, что «стволовые эуартроподы» — это эволюционный уровень, охватывающий часть древа выше онихофор и ниже эуартропод в узком смысле. Что же на этом эволюционном уровне происходило?
А происходило там много интересного. «Стволовые эуартроподы» могут поразить чье угодно воображение — настолько необычен и многообразен их облик (см. рис. 1 и 3). Их членистое туловище часто украшали ряды жаброподобных, крылоподобных или лезвиеподобных выростов, способных служить своего рода плавниками. Появлялись хитиновые головные щиты, которые в некоторых группах становились двустворчатыми. Ходильные конечности менялись во всем диапазоне от примитивных «сосочков» (или их отсутствия) до сложно устроенных длинных ног, членистых, а иногда и двуветвистых, как у ракообразных. Невероятным разнообразием отличались околоротовые придатки: членистые или нечленистые, зачаточные или очень мощные, короткие или длинные, хватательные или фильтрующие, иногда ветвящиеся, а иногда с клешнями, крючками, шипами или щупальцами. Наконец, у ряда форм возникли огромные сложные фасеточные глаза, примерно такие же, как у современных насекомых, и иногда сидящие на стебельках. И все эти признаки вступали между собой во множество сочетаний, часто неожиданных для современных зоологов.
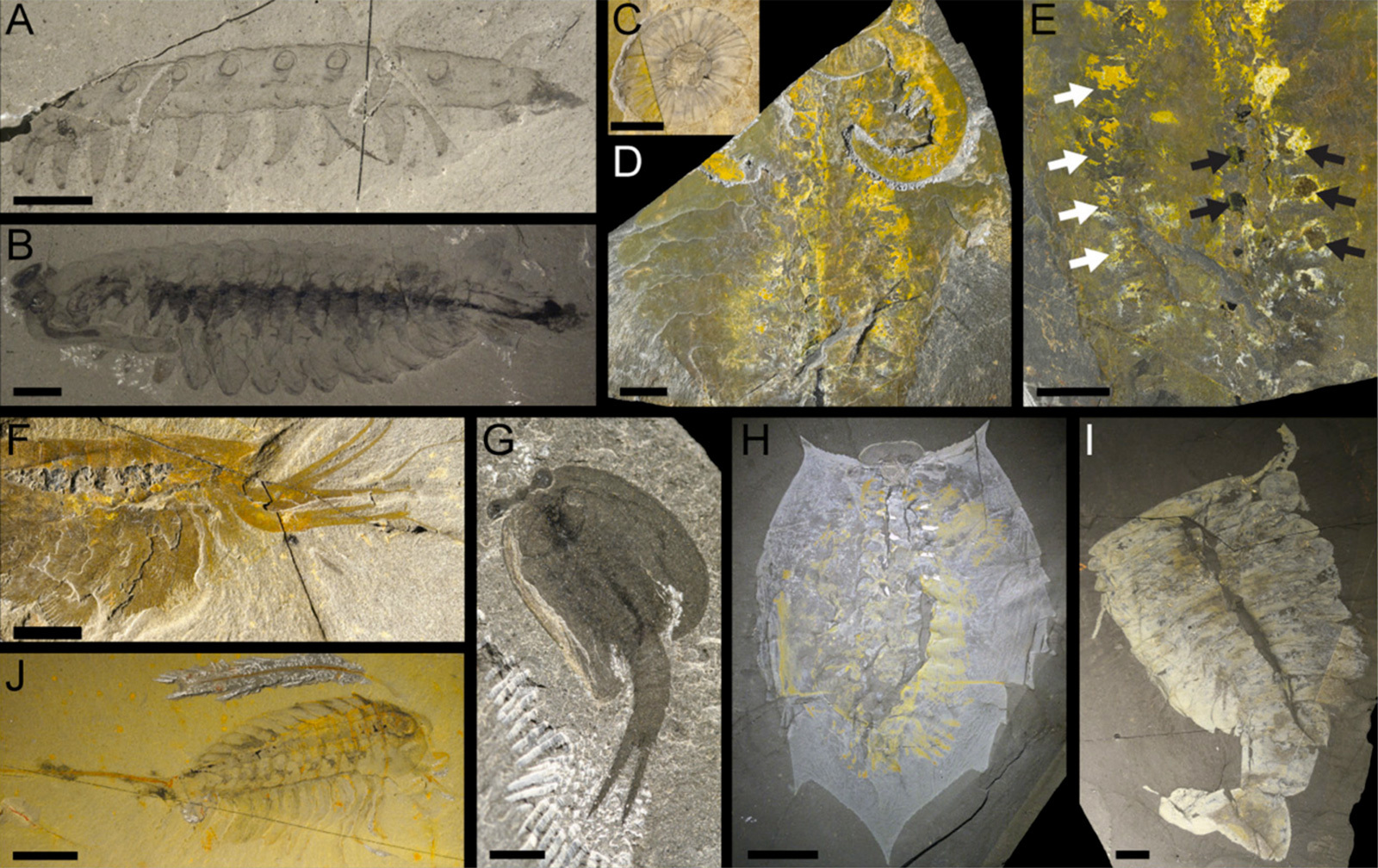
Рис. 3. Панартроподы из сланцев Бёрджесс. А — близкая к современным онихофорам Aysheaia. B — Opabinia, обладатель плавниковых выростов, фасеточных глаз и членистого хобота, в данном случае подогнутого под туловище. C — Hurdia, от которой сфотографирован только ротовой аппарат с концентрическим расположением хитиновых зубцов (у настоящих членистоногих ничего подобного не бывает). D — Anomalocaris. Хорошо видны длинные членистые околоротовые придатки. E — еще один Anomalocaris. Белые стрелки указывают на сегментарные мышцы, черные — на железистые выросты кишечника. F — Leanchoilia. Околоротовые придатки длинные и разветвленные, эти ветви служат щупами. G — Perspicaris, обладатель двустворчатого головного щита. H, I, J — Helmetia, Sidneyia и Emeraldella, вероятные родственники трилобитов. Длина масштабной линейки 5 мм на A, 10 мм на B–F и H–J и 3 мм на G. Иллюстрация из обсуждаемой статьи в PNAS
Источник: Allison C. Daley, Jonathan B. Antcliffe, Harriet B. Drage, and Stephen Pates. Early fossil record of Euarthropoda and the Cambrian Explosion // Proceedings of the National Academy of Sciences. 2018. V. 115. № 21. P. 5323–5331. DOI: 10.1073/pnas.1719962115.
Сергей Ястребов
http://elementy.ru/novosti_nauki/433315/Paleontologi_utochni...
Лига Палеонтологии
2.4K постов15.1K подписчиков
Правила сообщества
– Провокации в дискуссиях, излишнюю агрессивность, троллинг, расизм.
– Ничем не подкрепленные нелепые гипотезы, предположения, фальсификации.
– Креационизм, MLP-фэндом – им не место в научном сообществе.
– Оскорбления любой разновидности, Будьте вежливы!
– Поднимание тем политики – для неё есть отдельный раздел.
– Рекламирование чего-либо.
– Многократные нарушения критериев постинга, и игнорирование пометок для исправления.
– Активное отрицание эволюции как факта:
Вы можете признавать Синтетическую Теорию Эволюции или научную парадигму в целом, можете не признавать их, но сама эволюция есть как факт, доказанный экспериментально: опыты Шапошникова и эксперимент с E.Coli,
E.Coli и антибиотик, Методы борщевика, Карта эволюции.
Теория же описывает процесс и механизмы эволюции, и является высшей формой организации научного знания:
Почему СТЭ все ещё теория?
Критерии одобрения постов:
– Посты соответствуют тематике сообщества ⇑
– Посты не должны содержать ненаучную ересь и фальсифицированные факты (Например люди-великаны)
– Тег Копипаста, если статья не ваша, так же желательна ссылка на источник.
– Тег Юмор, если пост несет исключительно юмористическую тему.
– Видеоматериалы обязаны сопровождаться кратким описанием.
– Название постов не должно вводить читателей в заблуждение.