

Как работает иммунная система, лекция 3 ч. 10 (ч. 37)
ИТОГ
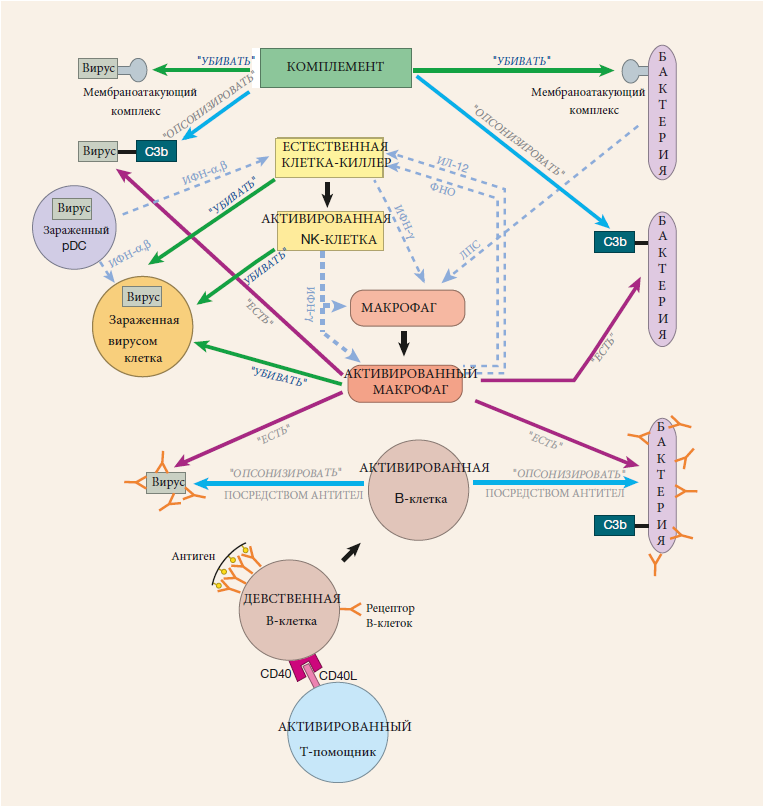
Рецепторы В-клетки работают как “глаза” и фактически состоят из двух частей: части распознавания (состоящей из белков тяжелой и легкой цепей) и сигнальной части (состоящей из двух других белков, Iga и Igβ). Гены, кодирующие часть распознавания, создаются путем смешивания и сопоставления сегментов генов. В результате получается набор В-клеток с настолько разнообразными рецепторами, что они, вероятно, могут распознать любую органическую молекулу во вселенной. Для того чтобы эти рецепторы сигнализировали о том, что они видели, требуется, чтобы несколько рецепторов В-клеток были сгруппированы (сшиты). И когда достаточное количество молекул Iga и Igβ группируется таким образом, сигнал “задействованный рецептор” отправляется в ядро В-клетки.
У В-клеток также есть на своей поверхности молекулы ко-рецепторов, которые могут распознавать опсонизированный антиген. Рецепторы и ко-рецепторы В-клеток задействованы антигеном, количество рецепторов, которые должны быть сшиты для активации сигнала, резко уменьшается. Следовательно, эти ко-рецепторы фокусируют внимание В-клеток на антигенах, которые уже были распознаны врожденной системой как опасные и которые были опсонизированы.
Для активации девственной В-клетки требуется два “ключа”. Сшивание рецепторов В-клеток служит первым ключом, но также требуется второй, “ко-стимулирующий” ключ. Этот ключ обычно обеспечивается Т–клеткой-помощником и включает контакт между клетками, во время которого молекулы CD40L на поверхности Т-клетки-помощника связываются с белками CD40 на поверхности В-клетки. В-клетки также могут быть активированы без помощи Т-клеток. Первое требование для этой независимой от Т-клеток активации состоит в том, что большое количество рецепторов В-клеток должно быть сшито. Обычно это происходит, когда поверхность вредителя состоит из множества копий антигена, с которым связываются рецепторы В-клетки (ее родственный антиген). Хотя сшивание многих рецепторов В-клеток служит необходимым условием для независимой от Т-клеток активации наивной В-клетки, этого недостаточно. Также необходим второй, стимулирующий сигнал. Эта совместная стимуляция осуществляется в форме “сигнала опасности”, который подтверждает, что существует подлинная угроза. Требуя, чтобы для активации B-клетки были предоставлены два ключа, создается отказоустойчивая система, которая защищает от неправильной активации B-клетки.
Антитела IgM — это первые антитела, вырабатываемые В-клетками в ответ на патоген, с которым ранее не сталкивались. Однако по мере созревания В-клетки она может выбирать для выработки антител другого класса: IgG, IgA или IgE. Это переключение классов не изменяет антиген-связывающую область (Fab) антитела. Следовательно, антитело распознает один и тот же антиген до и после переключения его класса.
Что меняется во время переключения классов, так это константная область (Fc) тяжелой цепи. Это та часть молекулы, которая определяет, как функционирует антитело, причем некоторые функции лучше подходят для определенных вредителей, чем для других. Важно отметить, что выбор класса антител определяется цитокинами, присутствующими в локальной среде В-клетки, когда происходит переключение классов. Таким образом, организуя производство соответствующих цитокинов в соответствующих местах, можно создать правильный класс антител для защиты от конкретного вредителя.
Другое изменение, которое может произойти по мере созревания В-клетки, — это соматическая гипермутация. В отличие от переключения классов, при котором антитело получает другую константную область, соматическая гипермутация изменяет антиген-связывающую область антитела. Поскольку вероятность того, что В-клетка будет размножаться, зависит от аффинности ее рецептора к антигену, В-клетки, которые размножаются больше всего, будут теми, для которых соматическая гипермутация увеличила аффинность их рецепторов. В результате получается набор В-клеток, у рецепторов которых в среднем более тесная связь с вредителем, чем исходные, немутированные рецепторы. Эти модернизированные В-клетки особенно полезны в качестве клеток памяти. После созревания аффинности рецептора В-клетки, рецепторы более чувствительны к небольшим количествам антигена, поэтому эти В-клетки могут быть реактивированы в начале второй инфекции, пока число вредителей все еще относительно невелико.
В-клетки могут быть активированы с помощью Т-клеток или без них, но результаты в этих двух случаях обычно очень разные. Независимая от Т-клеток активация обычно приводит к выработке антител IgM. Напротив, активация, зависящая от Т-клеток, может привести к образованию антител IgG, IgA или IgE, аффинность которых уже созрела. Одна из причин этого различия заключается в том, что как переключение классов, так и соматическая гипермутация требуют лигирования белков CD40 на В-клетках. Этот сигнал обычно обеспечивается CD40L, белком, обнаруженным на поверхности активированных Т-клеток-помощников.
По мере созревания В-клеток они должны решить, становиться ли короткоживущими плазматическими клетками, которые производят огромное количество антител, или оставаться в качестве более долгоживущих В-клеток памяти. Эти В-клетки памяти отвечают за выработку антител, которые могут защитить нас от последующего нападения того же патогена.
ИТОГОВАЯ СХЕМА
Вопросы для размышления
1.В-клетки производятся в соответствии с принципом клонального отбора. Что именно это значит?
2.Опишите, что происходит во время Т-клеточно-зависимой активации В-клеток.
3.Как можно активировать В-клетки без помощи Т-клеток и почему независимая от Т-клеток активация В-клеток важна для защиты нас от определенных патогенов?
4.Опишите отказоустойчивые системы, которые участвуют в активации В-клеток.
5.Каковы основные характеристики антител IgM, IgG, IgA и IgE?
6.Почему переключение классов и соматическая гипермутация производят В-клетки, которые лучше способны защищаться от вредителей?
Перевод книги LAUREN SOMPAYRAC "HOW THE IMMUNE SYSTEM WORKS", продолжение следует.