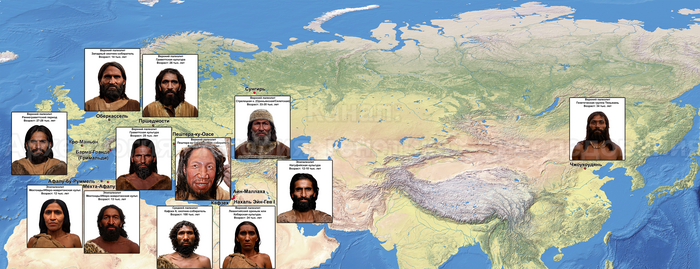
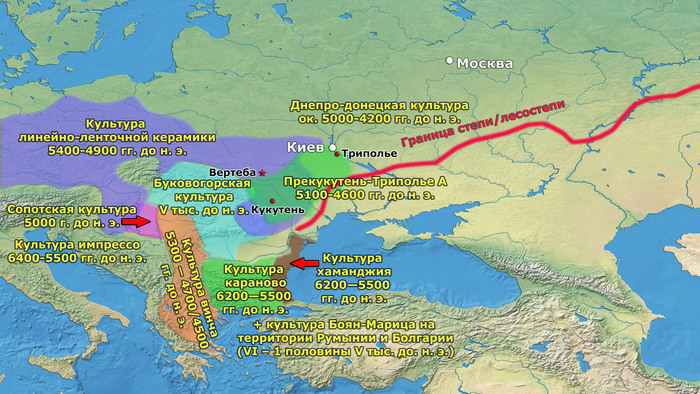
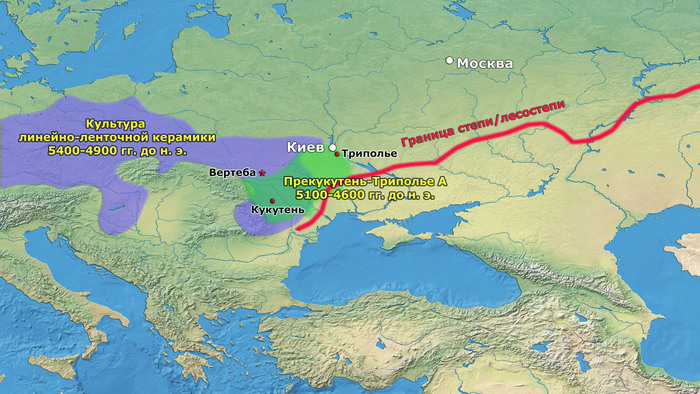
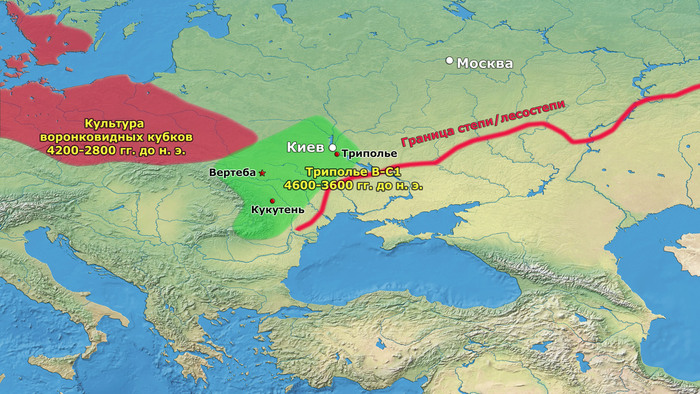
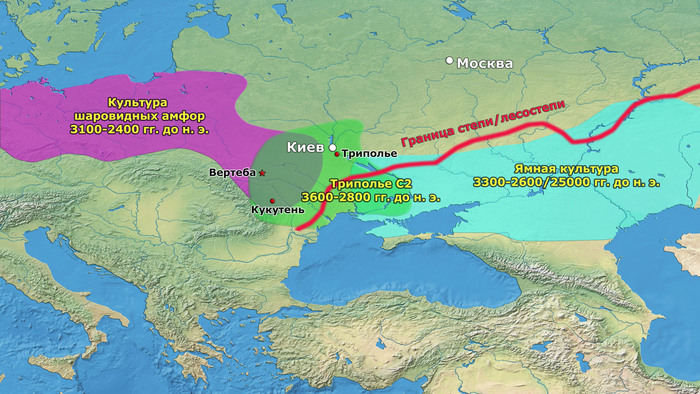
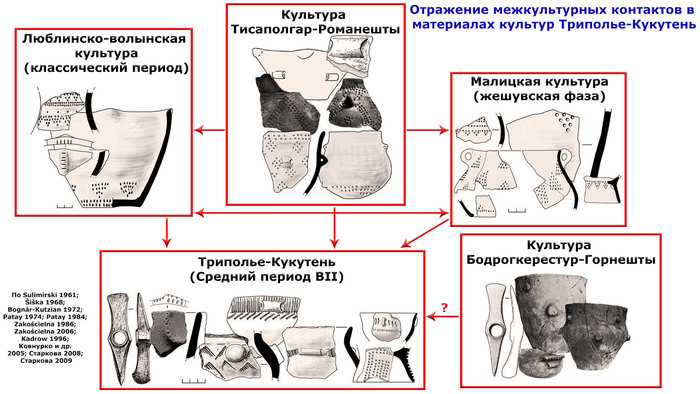
В статье речь пойдёт о генетических данных представительницы древних тюрков – принцессы, а после брака с императором китайско-сяньбийской династии Северной Чжоу - У-ди, императрицы – Ашины из одноимённого рода, дочери Мукан-кагана, третьего кагана древних тюрков – Ашина Сыцзинь, при котором Тюркский каганат расширил свои границы от Жёлтого до Чёрного моря, став крупнейшей империей того времени.
Кстати, о происхождении и внешности супруга Ашины - третьего императора китайско-сяньбийской династии Северной Чжоу - У-ди (Юйвэнь Юна) будет в следующей статье.
А в данном исследовании авторы, проанализировав генетический профиль императрицы Ашины, пытаются ответить на три ключевых вопроса:
1. Какое было происхождение у древних тюрков?
2. Есть ли генетическая связь между Ашиной и другими тюркскими народами, а также различными популяциями Центральной и Восточной степи после железного века?
3. Есть ли генетическая связь между древними тюрками и современными тюркоязычными народами?
Вступление
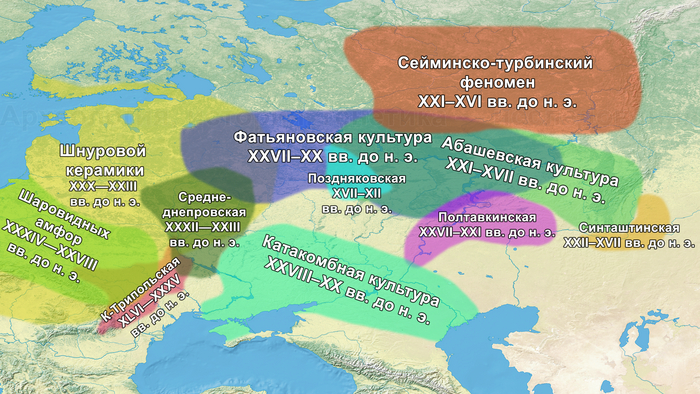
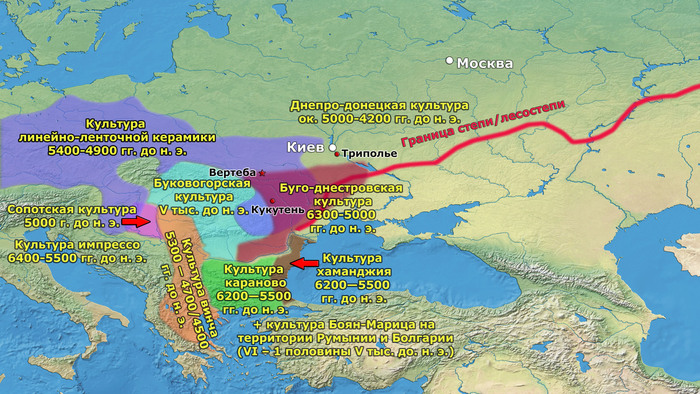
Первый расцвет тюркских народов пришелся на период с VI по VIII век, когда возникла могущественная кочевая империя в виде Тюркского каганата, основанного правителями из рода Ашина, простиравшегося от Монгольского плато до Каспийского моря. Появившись вслед за хунну, сяньби и другими кочевыми конфедерациями, влияние тюрков ощущалось по всей Евразии, что повлияло на языковой ландшафт значительной её части в течение следующих столетий. Тюркский язык, состоящий из более чем 40 языков, является самой многочисленной группой в гипотетической алтайской языковой семье, и около 200 миллионов человек говорят на тюркских языках более чем в 10 странах.
К сожалению, исторические сведения о Тюркском каганате носят разрозненный характер и часто противоречивы. Происхождение древних тюрков является предметом споров, но если не брать мифические истории с тотемными предками по типу появления от хуннского царевича и волчицы, их происхождение может быть сведено к трем конкурирующим гипотезам:
1. Представители племени Ашина выходцы из одноимённого племени хунну в Северо-Восточной Азии.
2. Их происхождение связано с Причерноморско-Каспийской степью или Центральной Азией с последующей миграцией на восток.
3. Сложносоставное происхождение в окрестностях Пинляна или Гаочана в Турфанской впадине, охватывающее как восточные, так и западные евразийские этнические группы.
Поэтому происхождение представителей правящего рода из племени Ашина вызывает большой интерес и является ключом к разгадке происхождения древних тюрков. Тем более с учётом того, что среди тюркской знати была широко распространена кремация.
Результаты
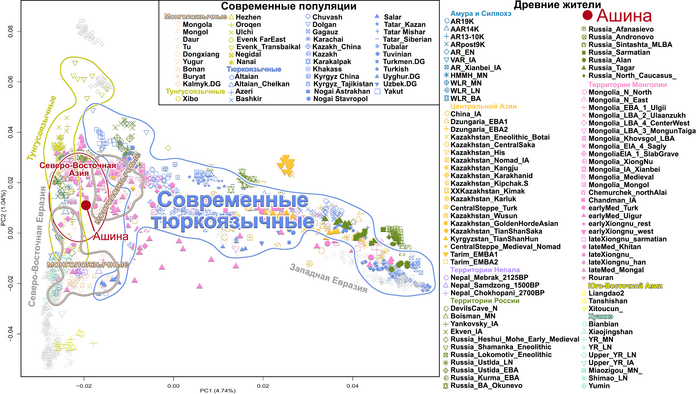
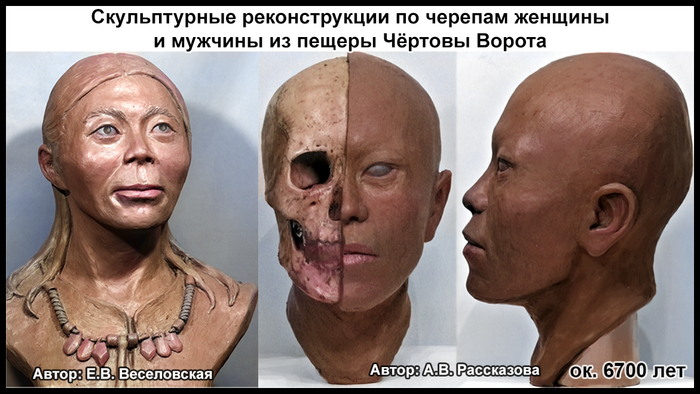
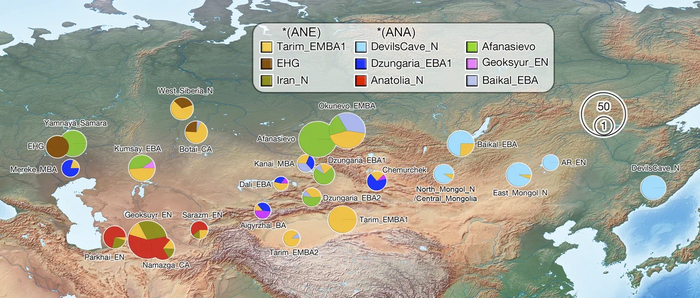
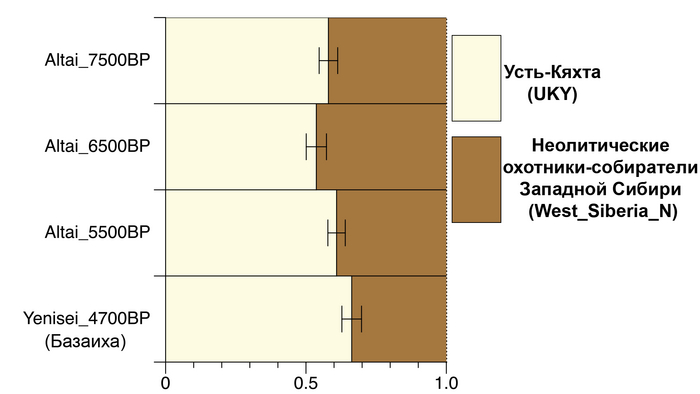
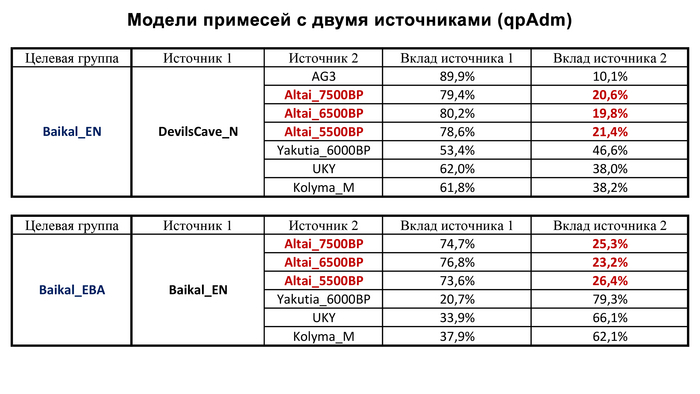
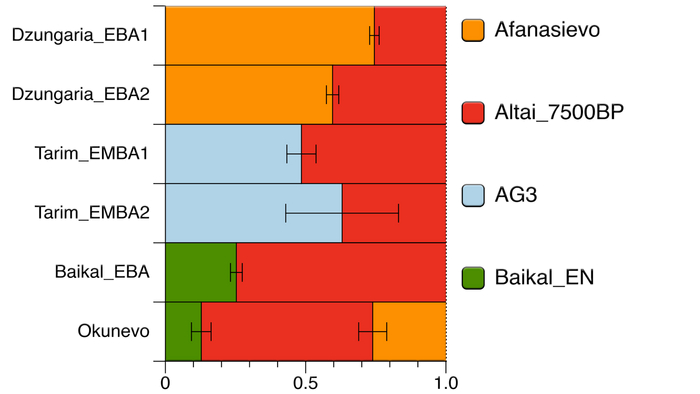
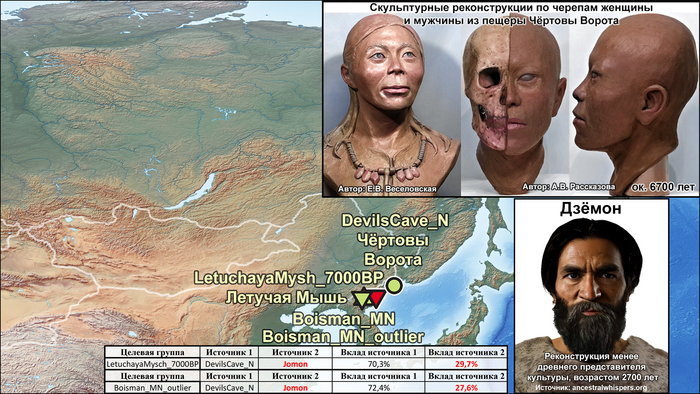
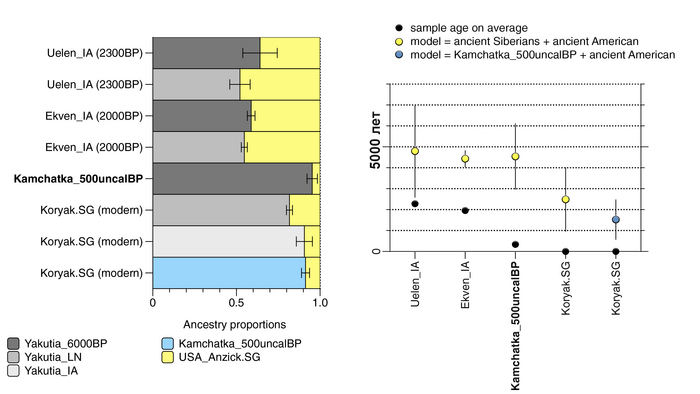
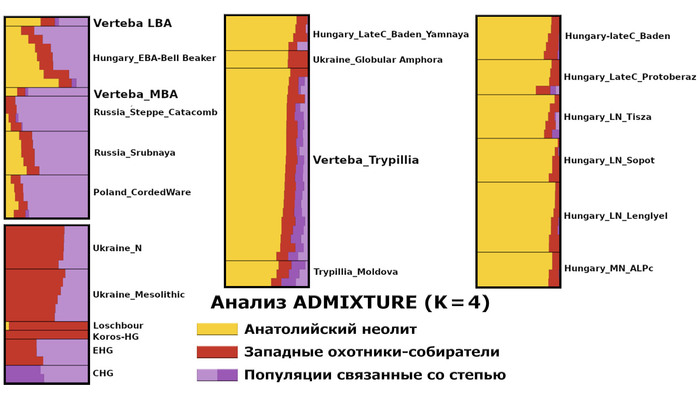
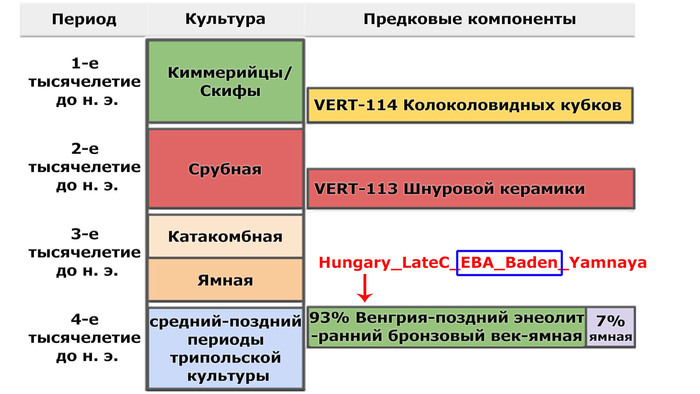
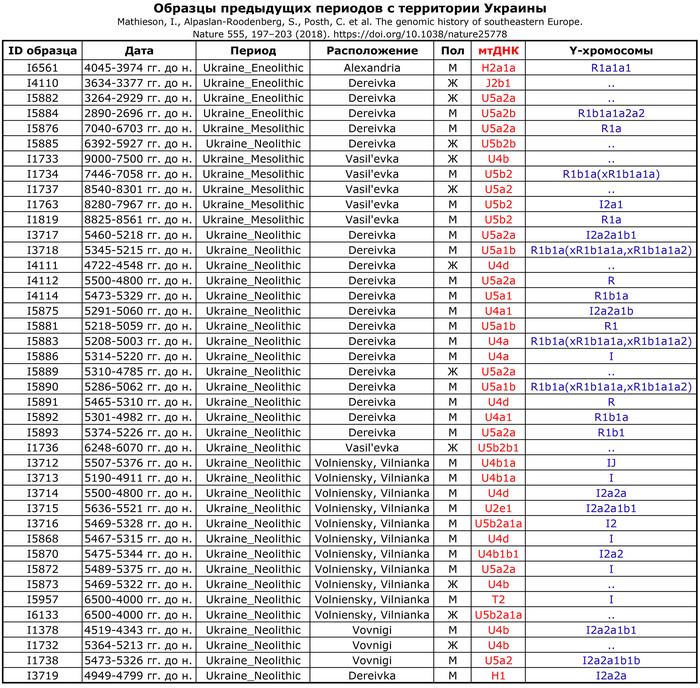
По данным мтДНК императрица Ашина принадлежала к гаплогруппе F1d, распространенной в Северо-Восточной Азии. А по результатам полногеномных анализов она группировалась с древними популяциями Северо-Восточной Азии и востока Монгольского плато, но особенно, с древними северо-восточными азиатами, как представители неолита из пещеры Чёртовы Ворота, возрастом около 6700 лет и носители бойсманской культуры среднего неолита из археологического памятника Бойсмана-2, Приморья, представителями неолита Амура и Северной Монголии, а также с кочевниками Восточной степи после железного века, включая сяньбийцев, жужаней, киданей и части монгольского населения. Из современных популяций она была генетически близка с современными тунгусо-маньчжурскими и монголоязычными народами.
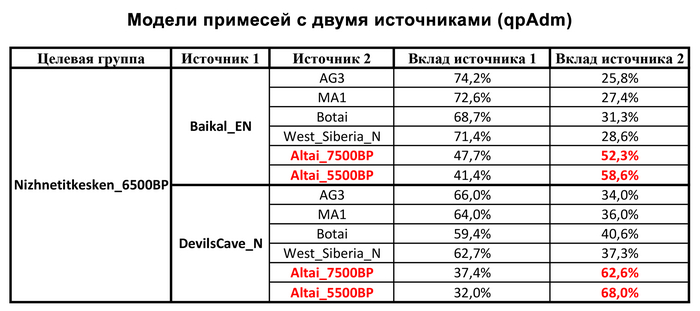
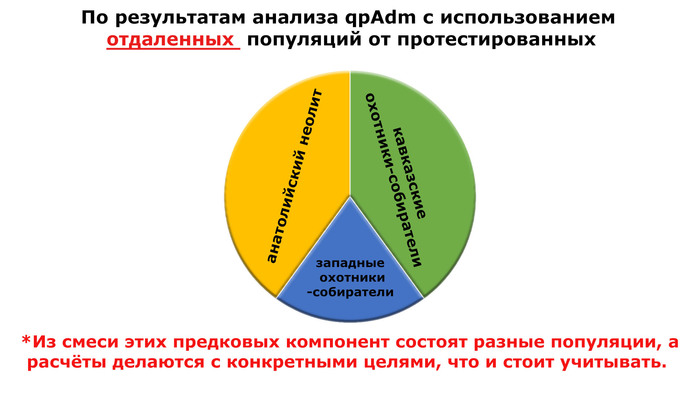
При этом вклада от древних китайцев, связанных с жителями реки Хуанхэ у Ашины не было обнаружено. Однако отмечен небольшой вклад от жителей Западной Евразии – 2,3-3,9%, включающий степную родословную с примесью анатолийских земледельцев, который произошел у предков Ашины за 1566 ± 396 лет до её жизни. В целом генетические данные опровергают гипотезы о западно-евразийском и множественном происхождении представителей знатного рода тюркских каганатов – Ашина в пользу восточноазиатского происхождения.
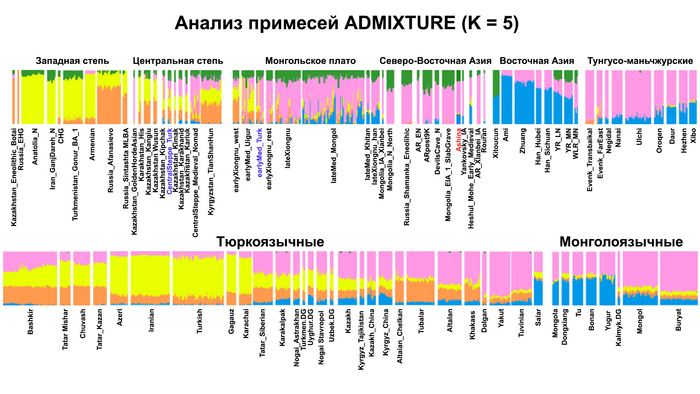
Однако стоит отметить, что у других образцов в тюркский период наблюдалось смешение компонент древних северо-восточных азиатов, которых в среднем было более половины, с компонентами западноевразийских популяций Бактрийско-Маргианского археологического комплекса и кочевников степи железного века, что также соответствует заметному увеличению гаплогрупп Y-хромосомы R и J, в течение тюркского периода в Монголии.
Есть ли генетическая связь между Ашиной и другими тюркскими народами, а также популяциями Центральной и Восточной степи после железного века?
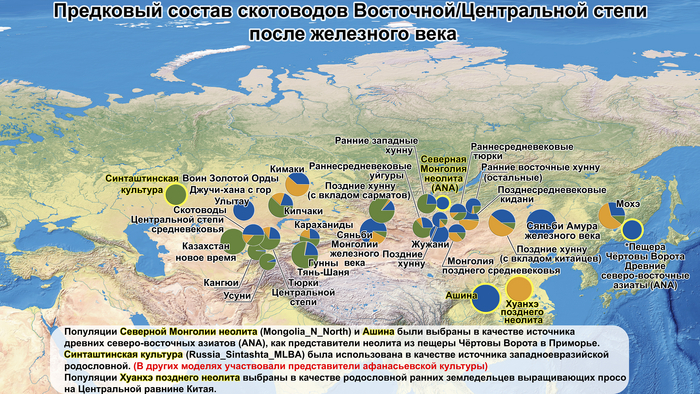
Начиная с железного века в Восточной Степи возникали и рушились, выходящие далеко за её пределы, различные государственные образования кочевников: хунну, сяньби, жужани, тюрки, уйгуры, кидани, кыргызы и – наиболее агрессивные и успешные из всех – монголы. Соседняя средняя часть Евразийской степи или Центральная степь, также была свидетелем перемен в судьбах кочевых образований: гунны, усуни, кангюи, карлуки, кимаки, караханиды и кыпчаки.
Однако представительница элиты Тюркского каганата продемонстрировала генетическую связь только с древнемонгольскими племенами сяньбийцев, киданей и жужаней, а также с представителями тунгусо-маньчжурских племён мохэ. А вот монголы, уйгуры, карлуки, кимаки, кипчаки, а также караханиды были отдалены от кластера Ашины. Хунну вообще был союз разных по происхождению племён, они сильно отличались между собой по географии и по времени. Некоторые западные их представители вообще на 68,4% имели компоненты как у людей из афанасьевской культуры.
Кстати в моделях надо адекватно воспринимать компоненты, они подразумевают схожие генофонды, а не непосредственных представителей культур.
Среди скотоводов Центральной степи усуни, кангюи и гунны Тянь-Шаня большую часть своего происхождения (62,4–73%) ведут от западных степных групп афанасьевской культуры.
А тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, имеют 35–50,6% компонент как у афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но доля восточно-евразийского происхождения увеличилась у средневековых скотоводов Центральной степи с 67,3% до 82,5%, за некоторым исключением. Однако стоит отметить, что ещё в новое время на территории Казахстана встречались люди с генетическим профилем на 75,5% происходящем от западных степных скотоводов. При этом Ашина не продемонстрировала близкого генетического родства с тюрками Центральной степи или раннего средневековья. Но в отличие от Ашины, тюрки Центральной Степи и раннесредневековые тюрки продемонстрировали высокую, но вариативную степень западно-евразийского происхождения, что указывает на существование генетической подструктуры ещё у древних тюркоязычных народов.
Есть ли генетическая связь между древними тюрками и современными тюркоязычными народами?
В скотоводческих образованиях Восточной Степи утверждалось, что распространение монгольских языков было связано с сяньбийскими, жужаньскими, киданьскими и монгольскими группами, в то время как хунну, тюрки и уйгуры считались связанными с распространением тюркских языков. В последнем случае были выдвинуты гипотезы о двух волнах распространения: булгаро-тюркской диффузии, начавшейся в гуннский период, спровоцированной более ранней экспансией хунну, и последующей демографической экспансии, связанной с Тюркским каганатом. При этом во втором и третьем веках нашей эры Центральная степь была населена ираноязычными группами, которые постепенно переходили на тюркские языки.
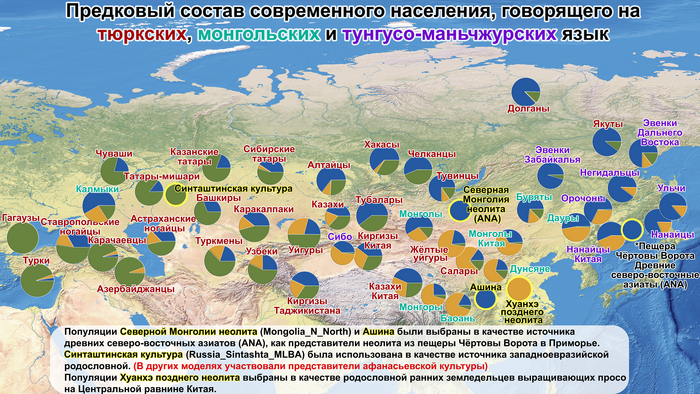
Поэтому генетические данные не подтверждают связи между древними и современными тюркоязычными группами и указывают на значительные различия между тюркоязычными популяциями и популяциями, связанными с распространением тюркских языков.
А пропорции предков древних северо-восточных азиатов, которые составляли основу древних тюрков, варьировались у тюркоязычных групп и даже полностью отсутствовали у некоторых самых западных их представителей. Это указывает на то, что распространение тюркских языков было в основном обусловлено культурными факторами, а не демографической диффузией и интеграцией населения. Поэтому натягивать современные тюркоязычные популяции на тюркизированные древние в качестве доказательства тюркоязычности этих древних популяций абсолютно не имеет смысла и противоречит всем данным.
Пояснение
А подвести итог хочу простым языком, отклонившись от строгой научной линии повествования. Ещё раз: это не научная часть!
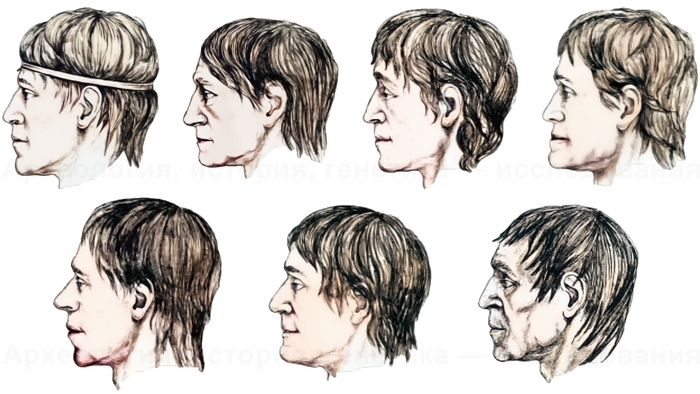
Древние тюрки, и тем более из рода Ашина, по происхождению были близки к древним северо-восточным азиатам, с незначительным вкладом от западных степных скотоводов. По реконструкциям понятно даже как они выглядели. Это характерные отличия.
Древние и современные тюрки имеют разное происхождение. И многие современные тюркоязычные народы объединяет лишь язык.
Отдельные представители тюркских народов в поисках своих предков почему-то обращаются к древним тюркам, однако схожих с ними компонент у них не так много, а у некоторых вообще почти нет. Их генофонд состоит из смеси различных древних популяций, говоривших на разных языках, при этом у разных народов и разный генетический коктейль и своя интересная история. Но если древних тюрков начинать искать от современных, то можно выйти на разные популяции, которые внесли свой вклад в современные и намеренно или искренне заблуждаясь, объявить их тюрками.
Характерным примером таких манипуляций выглядит якобы доказательство тюркоязычности скифов, основанное только на словах из исследования 2017 года, что современные потомки западно-скифских групп встречаются среди различных современных групп на Кавказе и в Центральной Азии, в то время как сходство с восточно-скифскими группами встречается более широко, но почти исключительно среди современных тюркоязычных групп, особенно из кыпчакской ветви тюркского языка, которые в прошлом были кочевниками. Что не удивительно с учётом огромного генетического разнообразия этих групп, значительно большего чем у всех народов Европы вместе взятых. Трудно промахнуться в стадо слонов из дробовика. К тому же ещё и с учётом того, что сами тюркоязычные карлуки, кыпчаки и караханиды, в разных моделях, помимо древних северо-восточных азиатов, как уже было сказано выше, имеют 35–50,6% предков от афанасьевцев, 10,5–21,7% от жителей Бактрийско-Маргианского археологического комплекса и 38,9–49,4% от выходцев из бассейна реки Хуанхэ железного века. Но они же по определению тюрки, и мы не называем кыпчаков индоевропейцами или монголами с китайцами! С такой логикой если в Анадыре с вами поздоровался и разговаривает чукча на чисто русском языке, то его предки по мнению таких вот специалистов в кавычках, должны быть исключительно русскими.
Работа о скифах 2017 года: Unterländer, M., Palstra, F., Lazaridis, I. et al. Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat Commun 8, 14615 (2017). https://doi.org/10.1038/ncomms14615
И это исследование 2017 года в комментариях, не цитировал разве что только ленивый защитник тюркского происхождения скифов и не только. И продолжат это делать, вот увидите. При этом само исследование они не читали и вряд ли понимают…
Толкование скифских слов из тюркского несостоятельно хотя бы потому, что фонетика засвидетельствованных скифских слов противоречит фонетическим характеристикам пратюркского (распад которого по глоттохронологическим данным относится к I в. до н. э., т. е. много позже появления на исторической арене скифов – соответственно, сравнивать скифские слова имело бы смысл только с пратюркскими, а не с их рефлексами в отдельных древних и современных тюркских языках). Так, пратюркское слово не могло начинаться на сонанты *m
и *l, тогда как нам известны скифские имена Мадий и Липоксай; невозможными в пратюркском были и скопления согласных в анлауте (начале слова), засвидетельствованные в самоназвании скифов и таких именах, как, например, Спаргапиф. Одним словом, попытки объявить скифов тюрками представляют интерес разве что с точки зрения современной этнопсихологии. Сергей Кулланда http://генофонд.рф/?page_id=3267
1/3
Интервью одного из авторов работы о скифах 2017 г., руководителя межинститутского сектора молекулярной палеогенетики Института цитологии и генетики СО РАН, Института археологии и этнографии СО РАН Александра Пилипенко на научно-популярном сайте elementy.ru
К примеру, от союза русского и китаянки положение их детей может быть на том же графике где-то около ногайцев или ближе к казахам.
*Очень грубый пример
И по пропорциям главных компонент они будут на них похожи, но ведь нельзя же будет их ребенка назвать казахом или ногайцем. Часто вижу непонимание этого элементарного биологического аспекта в комментариях. Но когда нужно что-то доказать о себе любимом и, тем более, великом, тут логика ломается.
Или упор часто делают на то, что если кто-то до наших дней сохранил традиции, связанные с кочевым образом жизни, питанием и тому подобно… То только его предки в прошлом могли этим заниматься. Мне вот интересно, чем занимались по такой логике предки тех, кто сейчас компьютерные технологии развивает или в космос ракеты запускает? А так и бомжам можно сказать, что они продолжатели славных традиций охотников и собирателей… Уж пусть не обижаются на меня современные кочевники и скотоводы, а также их потомки. В сохранении традиций нет ничего плохого, если не сочинять себе историю…
Примечание: текста много, тема сложная и состоит из двух постов. Разделы – "Социокультурные изменения" и "Итоги" (Появление и сокращение генетического разрыва), во второй части.
Вступление
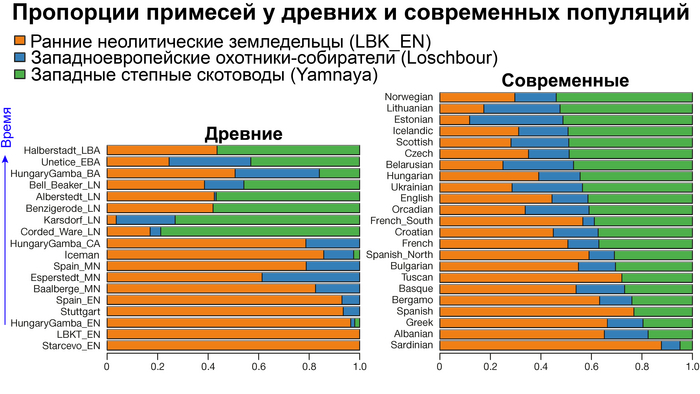
Генетическое разнообразие человеческих популяций Западной Евразии в значительной степени сформировалось в результате трех основных доисторических миграций:
1. Миграции охотников-собирателей, а именно анатомически современных людей, находящихся на ранней стадии социально-экономического развития, которые занимали территорию, примерно, с 45 тыс. лет назад. При этом вклад в последующие популяции внесли выжившие их линии после последнего ледникового максимума около 25-19 тыс. лет назад.
2. Далее, гораздо ещё более существенный вклад, внесли ранние неолитические земледельцы, пришедшие с Ближнего Востока примерно с 11 тыс. лет назад.
3. Степные скотоводы, распространившиеся из Причерноморско-Каспийской степи около 5000 лет назад, привнесли третью основную генетическую компоненту в популяции Западной Евразии.
Аутосомный генофонд практически всех европейских популяций по большей части состоит из различных пропорций этих предковых компонент, при этом независимо какие у них гаплогруппы.
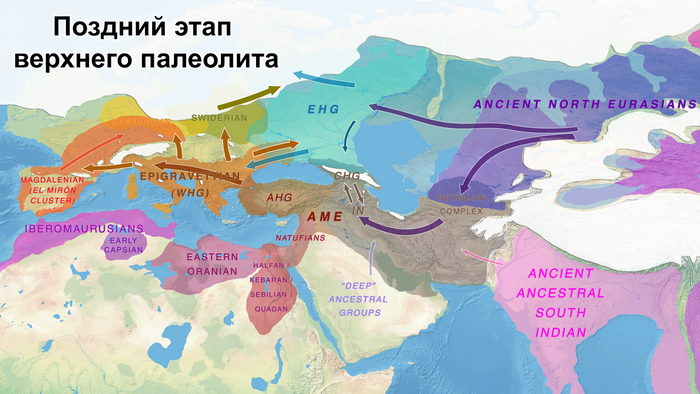
Европейский континент заселялся людьми современной анатомии в несколько волн, при этом не все линии этих людей были предками современных. Первые «сапиенсы» Европы (более 40 тыс. лет назад) пересекались и скрещивались с неандертальцами, пока те совсем не вымерли. Об этой примеси свидетельствуют геномы людей из пещеры Бачо Киро в Болгарии, связанных с археологической культурой начального верхнего палеолита, и со стоянки Пештера-ку-Оасе в Румынии. Другие люди того периода, такие как женщина из пещеры Златый кунь Чехии и человек из Усть-Ишима России, не несут в своих геномах значительного количества неандертальского вклада, чем другие неафриканские группы. При этом люди, жившие в Европе более 40 тыс. лет назад, не оставили существенных следов в генетическом составе современных евразийских популяций, что говорит о том, что они в значительной степени вымерли или были ассимилированы последующими группами людей современной анатомии.
В те времена, на демографию древних людей влияли различные факторы, в том числе и климатические. Пик похолодания во время последнего ледникового максимума (LGM) был около 21 тыс. лет назад, когда Северная Европа была в значительной степени покрыта льдом, а южная граница ледниковых щитов проходила через Германию и Польшу. Многие участки Европы стали холодными пустынями с тундростепями и лесостепями на побережье Средиземного моря. А люди и животные переместились в южные широты, о чем свидетельствует, например, солютрейская культура на Пиренейском полуострове и на юге Франции. Другими предложенными рефугиумами или климатическими убежищами в тот период были Апеннинский полуостров, Балканы и юго-восточная часть Европейской равнины.
А в результате послеледниковой колонизации, палеогенетический анализ выявил разделение популяций охотников-собирателей на западных и восточных. Источники родословной западных охотников-собирателей (WHG) связаны с носителями эпиграветтской, азилийской и других эпипалеолитических культур генетического кластера Виллабруна, появление которого обусловлено миграциями в конце ледникового периода и появлением новой генетической компоненты, связанной с Ближним Востоком того времени, начиная с бёллинг-аллерёдского потепления около 14 тыс. лет назад, что археологически коррелирует с культурными переходами.
А родословная восточных охотников-собирателей (EHG) демонстрирует дальнейшую примесь с верхнепалеолитическими популяциями Сибири - древними северными евразийцами (ANE).
Предковый состав западных охотников-собирателей (WHG) был регионально изменчивым в мезолитических популяциях. Если охотники и собиратели из Британии и северо-западной континентальной Европы продемонстрировали однородный состав от палеолита и мезолита, а также вплоть до неолитической экспансии около 6000 лет назад, то их коллеги из Испании, представляли собой смесь людей из двух рефугиумов, когда к генетическому профилю кластера Виллабруна, добавляется примесь от более ранних местных пиренейских охотников-собирателей, связанных с солютрейской и мадленской культурами верхнего палеолита.
Этот генетический профиль сохранился во время последнего ледникового максимума в Юго-Западной Европе около 25-19 тыс. лет назад.
А время появления примеси сформировавшей восточных охотников-собирателей (EHG), оценивается в 15–13 тыс. лет назад, однако её пропорции в популяциях, по-видимому, соответствуют клине, которая в целом коррелирует с географией, так охотники-собиратели Прибалтики и Украины демонстрируют большее сходство с предками кластера Виллабруна, чем охотники и собиратели западной части современной России, у которых было больше вклада от древних северных евразийцев (ANE).
В свою очередь скандинавские охотники и собиратели продемонстрировали различные пропорции смеси западных и восточных охотников и собирателей (WHG и EHG) в позднем мезолите.
Стоит отметить, что охотники и собиратели мезолита и тем более палеолита, были очень разнообразны и неоднородны, но из-за ограниченного количества данных, наши знания о мезолитической структуре населения и процессах демографического смешения долгое время были скудны и содержали существенные хронологические и географические пробелы в информации. Отчасти это объясняется малым количеством хорошо сохранившихся мезолитических человеческих останков возрастом более 8000 лет, а отчасти и потому, что большинство исследований древней ДНК, посвященных периодам мезолита и неолита, были ограничены преимущественно находками из Европы.
1/3
Хотя давно напрашивается более детальная классификация охотников и собирателей по их характерным отличиям. Так, археологические данные указывают на условную границу от восточной Балтики до Черного моря, к востоку от которой сообщества охотников и собирателей существовали гораздо дольше, чем в Западной Европе, несмотря на аналогичное расстояние до центра распространения раннего земледелия на Ближнем Востоке.
Компоненты восточного и западного происхождения у охотников-собирателей в этом пограничном регионе сильно различаются, но более точные представления о демографических процессах, связанных с разделением на восток и запад и о динамике численности населения к востоку от Европы, включая Северную и Центральную Азию за тот же период времени, весьма ограничены. В этих регионах термин «неолит» характеризуется культурными, экономическими и социальными изменениями, а также использованием керамики. К примеру, неолитические культуры степей Центральной Азии и таежного пояса России уже изготавливали керамику, однако продолжали вести прежний образ жизни с присваивающей экономикой, как у предшествующих мезолитических культур охотников-собирателей, с различными инновациями в зависимости от условий среды обитания. Однако пока недостаточно данных для понимания того, как неолитизация отличалась по своим срокам, механизмам и последствиям в разных уголках Евразии.
В Европе, кроме восточной её части, переход от охоты и собирательства к земледелию сопровождался прибытием мигрантов ближневосточного происхождения, практикующим производящее хозяйство на основе одомашненных растений и животных. С последующим значительным ростом населения. В Юго-Восточной, Центральной и Западной Европе анатолийские земледельцы впоследствии в некоторой степени смешались с местными охотниками и собирателями, а на территории Британии, прибывшие с континентальной Европы ранние европейские земледельцы полностью заменили местных охотников и собирателей.
А вот в восточно-балтийском регионе наблюдается заметно иная модель неолитизации, с появлением одомашненных животных только при появлении культуры шнуровой керамики (CWC) около 4800 лет назад. Аналогичным образом в Северном Причерноморье люди с генетическим профилем мезолитических охотников и собирателей тысячелетиями сосуществовали с земледельческими группами, расположенными дальше на запад.
И третий крупнейший генетический пласт в истории европейских популяций связан со степными скотоводами раннего бронзового века, как представители ямной культуры около 5 тыс. лет назад. Этот генетический профиль, быстро распространился по Европе посредством экспансии как носителей культуры шнуровой керамики (CWC), так и родственных ей культур.
Хотя предыдущие исследования выявили эти крупномасштабные миграции в Европу и Центральную Азию, основные аспекты, касающиеся демографических процессов, не решены. Ямная или степная родословная в общих чертах охарактеризована как смесь восточноевропейских (EHG) и кавказских охотников-собирателей (CHG), образовавшаяся в результате гипотетической смеси «северного» степного и «южного» кавказского источников. Однако точное происхождение этих предковых источников не было установлено. Более того, за некоторым исключением, известные гаплогруппы Y-хромосомы представителей ямной культуры не соответствуют гаплогруппам, обнаруженным у европейцев после 5000 тыс. лет назад и происхождение этой отцовской линии также не установлено. К тому же "степная" родословная в Европе до сих пор была выявлена только в смешанной форме, а детали и механизмы, с помощью которых эта родословная впоследствии распространилась вместе культурой шнуровой керамики (CWC), оставались неясными.
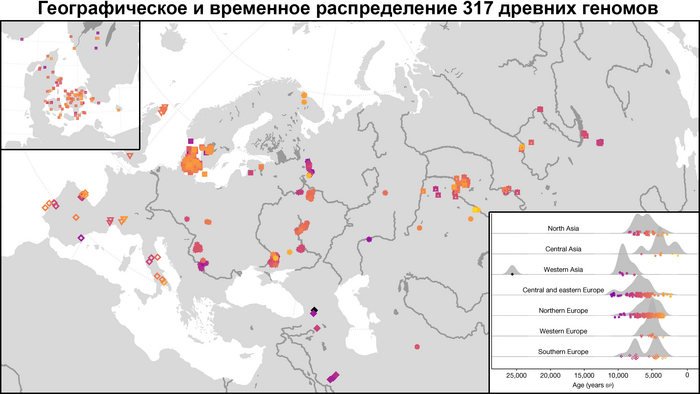
В новой работе учёные под руководством Эске Виллерслева из образцов, датированных радиоуглеродным методом при помощи ускорительной масс-спектрометрии, секвенировали геномы 317 человек, 309 из них датируются периодом от 11 до 3 тысяч лет назад.
А совместно с ранее опубликованными данными получилась выборка из 1600 древних геномов, секвенированных с помощью метода дробовика.
Географически 317 отобранных образцов охватывают обширную территорию Евразии, по долготе от Атлантического океана до озера Байкал, а по широте от Скандинавии до Ближнего Востока, и происходят из контекстов, которые включают курганы, пещеры, болота и дно водоёмов.
Географически образцы можно разделить на три региона:
1. Центральная, Западная и Северная Европа.
2. Восточная Европа, включая западную часть России, Беларусь и Украину.
3. Урал и Западная Сибирь.
Образцы включают представителей многих ключевых культур Западной Евразии мезолита, неолита и последующих периодов, такие как маглемозе, эртебёлле, воронковидных кубков (TRB) и шнуровой керамики или одиночных погребений в Скандинавии; импрессо или кардиумной керамики в Средиземноморье; кёрёш и линейно-ленточной керамики (LBK) в Юго-Восточной и Центральной Европе; и многие археологические культуры Украины, западной части России и Зауралья (как веретьё, льяловская, волосовская, китойская и др.). Выборка была особенно плотной в Дании, где сотня геномов, охватывала период от раннего мезолита до бронзового века. Обширная выборка была также получена из Украины, Западной России и Зауралья, охватывая период от раннего мезолита до неолита, примерно до 5000 лет назад.
Результаты
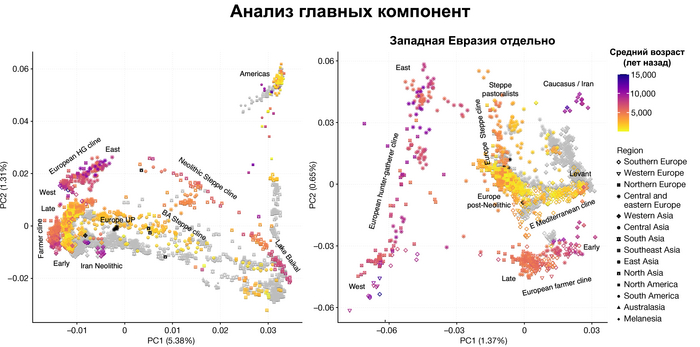
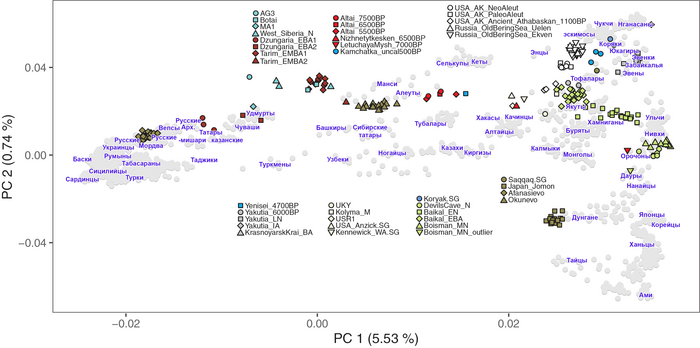
Полученный набор генетических данных позволил при анализе главных компонент использовать за основу древние геномы, и уже на них спроецировать положение современных популяций, а не наоборот, как это было во многих предыдущих исследованиях.
И хотя результаты согласуются с тем, что древние люди сильнее друг от друга генетически отличались, чем современные, такой подход позволил выявить ещё более значительные генетические различия между древними людьми, которые ранее не наблюдались, что отражает меньший эффективный размер их популяции и большую генетическую изоляцию.
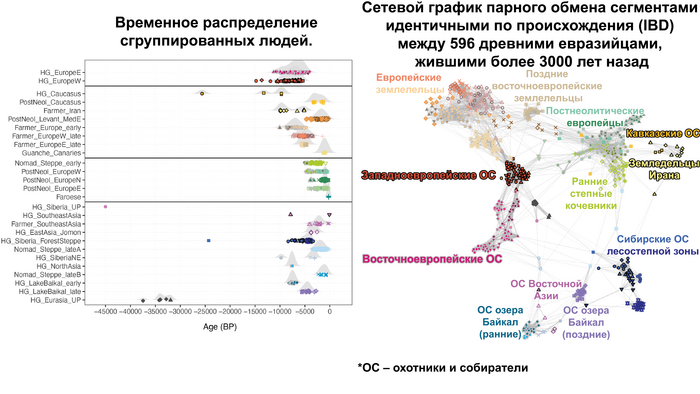
Чтобы получить более точную генетическую характеристику в пространстве и времени, авторы использовали попарную идентификацию сегментов ДНК идентичных по происхождению, чтобы сгруппировать анализируемых людей в связанные кластеры сходного генетического происхождения, а не по культурным или территориальным признакам.
В более крупном масштабе они отражали ранее описанные группы западных и восточных охотников-собирателей, а вот в более мелком масштабе они сгруппировались не только в пределах ограниченных пространственно-временных диапазонов и/или археологических контекстов, но также продемонстрировали ранее неизвестные связи в более широких географических диапазонах.
Полученные кластеры далее использовались для моделирования состава предков последующих популяций.
Популяционная структура охотников и собирателей после максимума последнего оледенения (LGM)
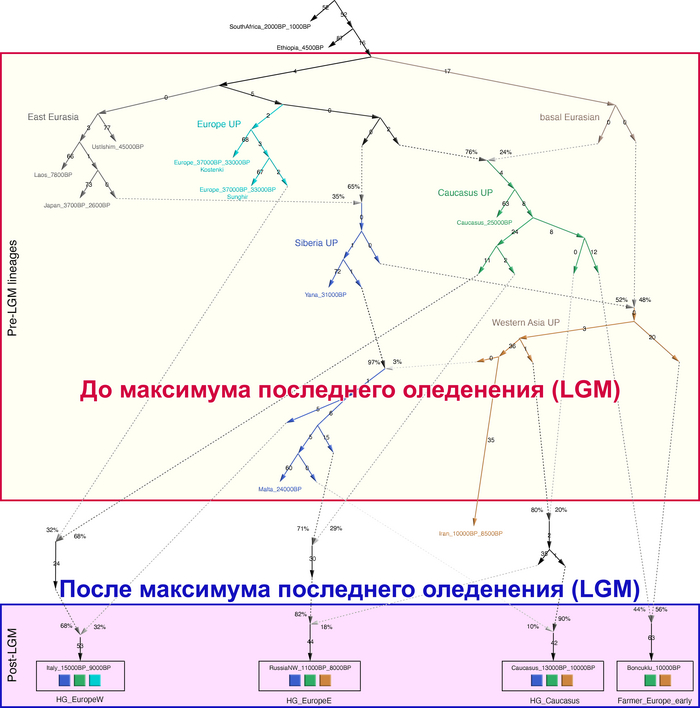
До последнего ледникового максимума, популяции Евразии довольно сильно отличались и в разной степени смешивались с неандертальцами. Некоторые популяции пока остаются гипотетическими, как к примеру, так называемые, «базальные евразийцы» с незначительной или нулевой примесью от неандертальцев, их прямых образцов, подтвержденных генетикой, на данный момент нет, но генетические следы этой популяции были обнаружены у охотников-собирателей Кавказа и Западной Азии верхнего палеолита, а также у более поздних западноевропейских охотников и собирателей.
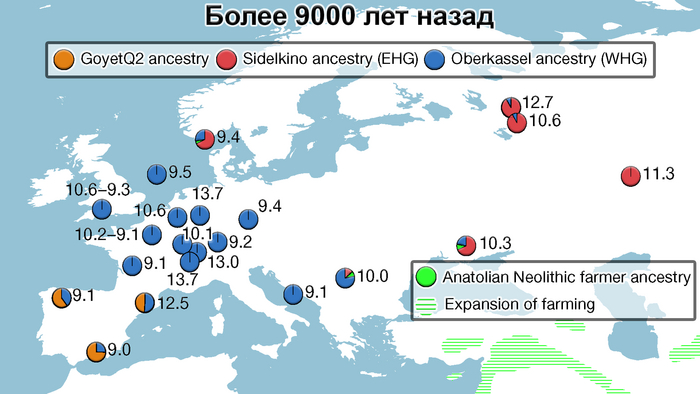
В данной работе авторы сообщают о генетических данных полученных из зуба женщины верхнего палеолита, жившей около 26-25 тыс. лет назад в окрестностях карстового грота Котиас Клде в современной Грузии, не путать с более молодыми образцами кавказских охотников-собирателей из этого археологического комплекса, у которых было менее 20% этой кавказской родословной верхнего палеолита. А сам геном этой представительницы Кавказа верхнего палеолита представляет собой смесь 76% западноевразийских охотников-собирателей верхнего палеолита (т.е. до максимума последнего оледенения) и 24% «базальных евразийцев». Кстати популяции Ближнего Востока верхнего палеолита, почти наполовину произошли от «базальных евразийцев», которые, практически не имели примеси неандертальцев и отделились от других неафриканских линий ещё до их разделения между собой. При этом западноевропейские охотники-собиратели, жившие после последнего ледникового максимума, моделируются как итоговая, за несколько событий, смесь 46% популяций, связанных с Кавказом верхнего палеолита, 22% европейцев, живших около 33-37 тыс. лет назад, связанных со стоянками Сунгирь и Костёнки и 32% древних популяций, связанных с Сибирью верхнего палеолита, как представитель стоянки Мальта, возрастом 24 тыс. лет, т.е. древних северных евразийцев.
В свою очередь восточноевропейские охотники-собиратели моделируются как смесь 58% популяций, связанных с обитателями стоянки Мальта в Прибайкалье, 28% популяций, связанных с Кавказом верхнего палеолита, как женщина из Котиас Клде (но в разные периоды, сначала 24%, а потом 4%) и 14% представителей Западной Азии верхнего палеолита. Эта смесь также проходила в несколько этапов. Тем самым новые данные подтверждают генетическое разделение охотников и собирателей в Восточной и Западной Европе после максимума последнего оледенения, когда люди снова расселялись по Евразии, при этом эти различия сохранялись на протяжении всего мезолита.
В анализе примесей, эти сложносоставные генофонды представлены отдельными компонентами. А различия между ними, а именно между западноевропейскими и восточноевропейскими охотниками-собирателями мезолита максимально выражены у представителей Италии, возрастом от 15 до 9 тыс. лет и Северо-Запада современной России возрастом от 11 до 8 тыс. лет.
Обратите внимание, что генетический состав одинаков даже у людей с разными гаплогруппами и наоборот, по всему геному люди с одинаковыми линиями гаплогрупп отличаются.
Кстати, в представлении сложных процессов, не стоит забывать о временных промежутках, о которых идёт речь.
Что касается североевропейских охотников и собирателей, то они отличались по пропорциям смеси компонент как у западноевропейских и восточноевропейских охотников-собирателей, за исключением Дании, где популяции продемонстрировали южноевропейское происхождение и были генетически схожи с охотниками-собирателями из Италии, возрастом от 15 до 9 тыс. лет. При чём эта родословная сохранялась в регионе на протяжении 5 тыс. лет.
В целом результаты указывают на то, что существовало по меньшей мере три отчетливые волны миграций охотников и собирателей в Скандинавию:
1. Первая волна связана с миграцией людей южноевропейского происхождения на территорию Дании и прибрежной части Юго-Западной Швеции.
2. Вторая волна в Прибалтику и Юго-восточную Швецию связана с популяциями из Юго-Восточной Европы.
И 3. Третья волна миграций на крайний север Скандинавии, а затем на юг вдоль атлантического побережья Норвегии связана с охотниками и собирателями с северо-запада современной России.
Эти миграции, вероятно, представляют собой послеледниковую экспансию из рефугиумов или районов-убежищ, общих как для людей, так и для многих видов растений и животных.
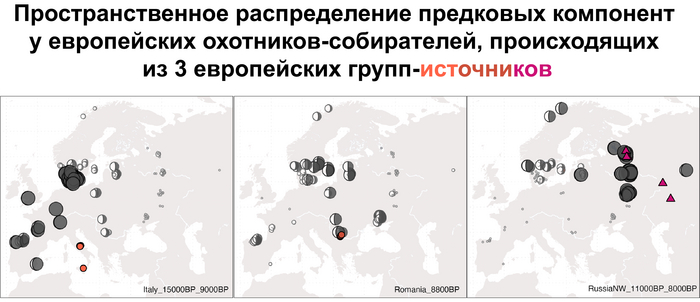
Стоит отметить, что генофонд европейских охотников-собирателей верхнего палеолита, живших до максимума последнего оледенения был в значительной степени заменён генофондом, происходящим из рефугиума в Южной Италии. На Пиренейском полуострове самые древние образцы из этого исследования, в том числе охотника-собирателя, возрастом около 9500 лет (NEO694) из бухты Санта-Майра на востоке Испании, показали преимущественно южноевропейское происхождение с незначительным вкладом охотников и собирателей верхнего палеолита, который, вероятно, отражает генофонд, связанный с мадленским периодом до максимума последнего оледенения.
А более поздние жители севера Пиренейского полуострова, были ближе к охотникам и собирателям Юго-Восточной Европы, демонстрируя около 40–30% вклада балканских охотников-собирателей.
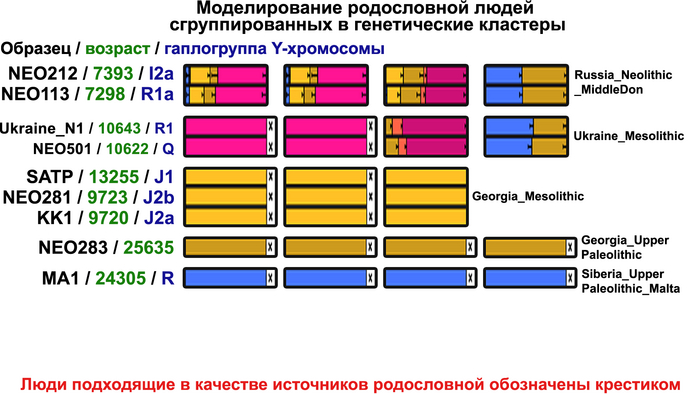
Поток генов, связанных с охотниками-собирателям Юго-Восточной Европы был направлен и в Восточную Европу после мезолита, на территорию современной Украины и далее. А у двух человек из могильника мариупольского типа раннего энеолита (NEO212, NEO113 ) близ хутора Голубая Криница на юге Воронежской области, возрастом около 7300 лет, генетический профиль состоял уже из смеси более ранних охотников-собирателей с территории Украины с вкладом от 18 до 24% из источника, связанного с охотниками-собирателями Кавказа.
1/2
Поэтому на графике анализа главных компонент они отклонены в сторону Ирана и Кавказа. По расчётам эта примесь произошла примерно 8300 лет назад. Результаты указывают на более ранний генетический контакт между популяциями Кавказа и степи ещё до появления культур кочевников в Причерноморско-Каспийской степи. Помимо этого, этот контакт задокументирован западнее, чем предполагалось ранее.
Основные генетические изменения в Европе
Предыдущие исследования древней геномики задокументировали несколько эпизодов крупномасштабной смены населения в Европе за последние 10 тыс. лет, но 317 новых геномов из данной работы, заполняют важные пробелы в наших знаниях.
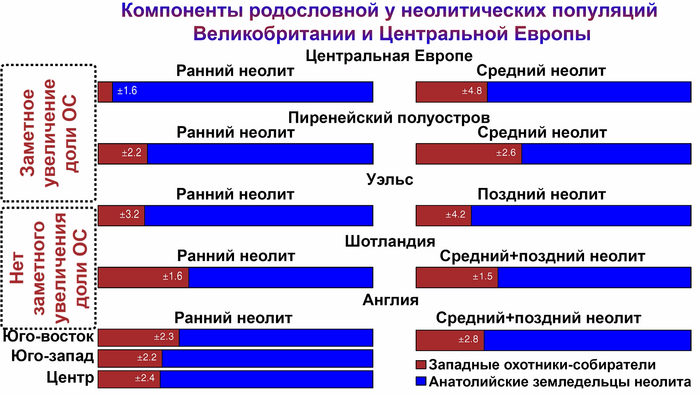
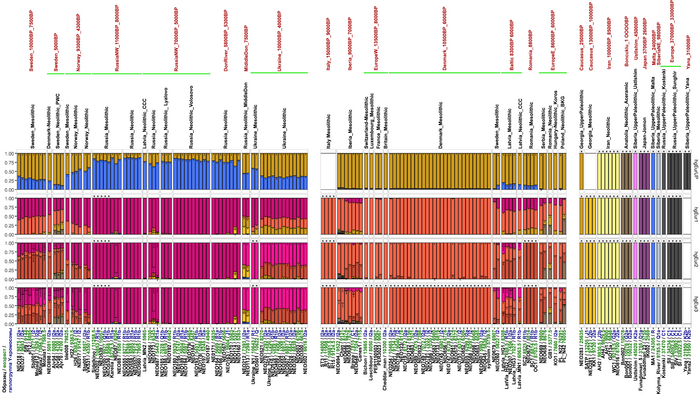
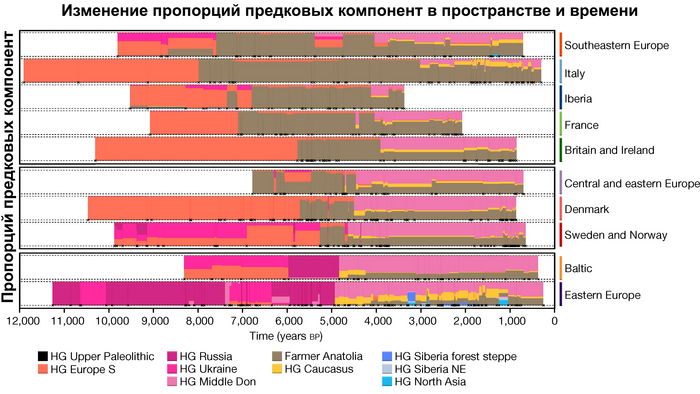
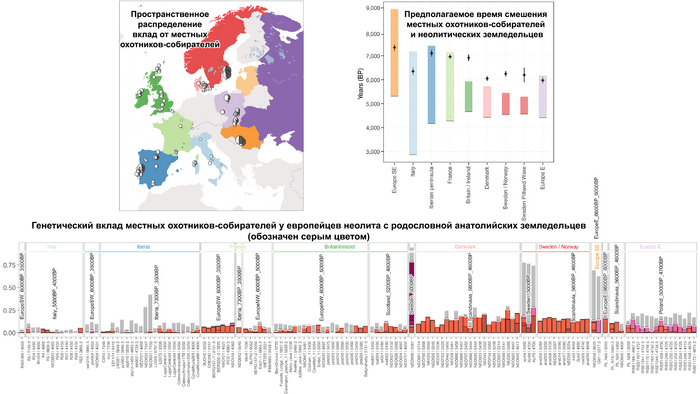
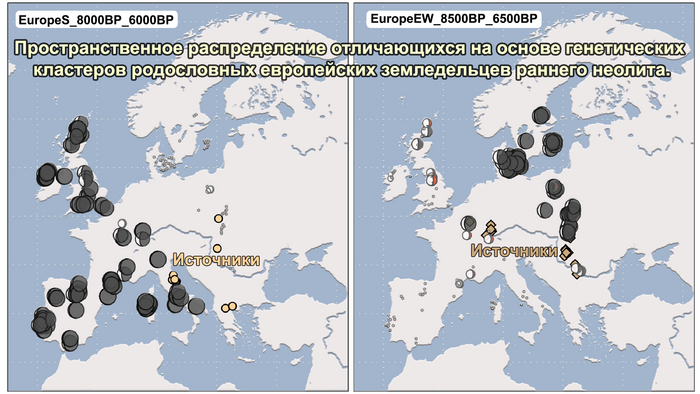
Анализ выявил глубокие различия в пространственно-временной динамике неолитизации между восточной и западной частями Европы вдоль пограничной зоны, простирающейся от Черного моря до Балтийского. К западу от этой пограничной зоны, переход к неолиту сопровождался крупномасштабными сдвигами в генетической родословной, от местных охотников-собирателей к земледельцам анатолийского происхождения (Boncuklu_10000BP). Этот переход в зависимости от региона охватывал обширный период времени, более 3 тыс. лет, от самых ранних свидетельств на Балканах около 8700 лет назад до 5900 лет назад в Дании.
В последующие периоды смешение между ранними европейскими земледельцами и местными охотниками-собирателями привело к частичному увеличению родословной охотников и собирателей во многих регионах Европы, но этот вклад редко превышал 10%, помимо заметных исключений, наблюдаемых у людей из района Железных ворот на границе современных Сербии и Румынии, а также у носителей культур ямочной керамики из Швеции и кардиумной керамики раннего неолита из Португалии, которые, по оценкам, содержат от 27 до 43% вклада от охотников и собирателей, как представители Пиренейского полуострова, возрастом от 9 до 7 тыс. лет (Iberia_9000BP_7000BP).
Обитатели эпохи неолита с территории современной Дании продемонстрировали до 25% генофонда охотников-собирателей (EuropeW_13500BP_8000BP), но преимущественно не местных.
Также обнаружены региональные различия в происхождении последующих неолитических групп. Так последующие неолитические популяции Западной Европы происходят от ранних земледельцев Южной Европы. А в последующие неолитические группы Восточной Европы и Скандинавии внесли свой вклад преимущественно ранние земледельцы Центральной Европы.
А вот с восточной стороны упомянутой пограничной зоны, на территории Прибалтики, Украины и западной части России, не наблюдается существенных генетических изменений, а предковые линии местных охотников и собирателей, без заметного вклада от анатолийских земледельцев,сохраняются примерно до 5 тыс. лет назад. Эта генетическая преемственность согласуется с археологическими данными, которые свидетельствуют о сохранении в этом обширном регионе групп охотников, рыболовов и собирателей, которые использовали керамику, но не практиковали земледелие и животноводство ещё на несколько тысяч лет.
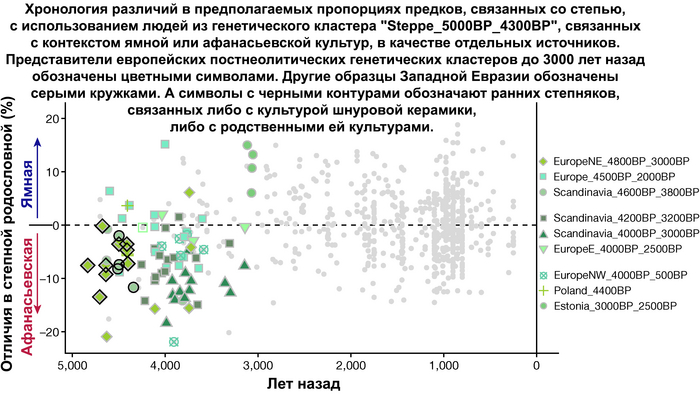
Но несмотря на то, что влияние степной родословной на популяции Евразии хорошо известно, вопросы о её происхождении оставались не решенными. В данной работе авторы показывают, что состав степной родословной (Steppe_5000BP_4300BP) может быть смоделирован как смесь примерно 65% компонент, связанных с охотниками-собирателями, использующими керамику, из района Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет (MiddleDon_7500BP) и 35% компонент, связанных с охотниками и собирателями Кавказа, возрастом от 13 до 10 тыс. лет (Caucasus_13000BP_10000BP).
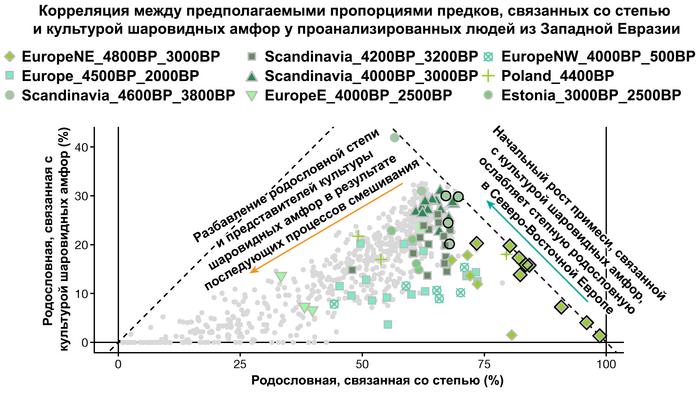
Таким образом, охотники-собиратели Среднего Дона, которые уже имели родословную, связанную с кавказскими охотниками-собирателями, являются неизвестным до сих пор ближайшим так называемым северным источником большей части родословной у людей, связанных с ямной культурой. При этом генетический состав людей из могильника Голубая Криница, отличается от такового у других древних людей из могильников мариупольского типа Приднепровья. Однако распространение степной родословной в Европе связно с культурой шнуровой керамики, носители которой представляют собой смесь предковых линий, связанных со степью и неолитическими земледельцами, происхождение которых может быть связано исключительно с культурой шаровидных амфор.
Это наблюдение имеет большое значение для понимания возникновения культуры шнуровой керамики. Хотя уже давно высказывались предположения о стилистической связи между керамикой культур шаровидных амфор и шнуровой керамики, включая использование сосудов в форме амфор и узоры, выполненные шнуром. При этом незадолго до появления самых ранних групп культуры шнуровой керамики восточные группы культуры шаровидных амфор и западные группы ямной культуры обменялись культурными элементами в лесостепной переходной зоне к северо-западу от Черного моря, где керамические амфоры и кремневые топоры культуры шаровидных амфор были включены в погребения ямной, а типичное для ямной использование охры встречалось в погребениях культуры шаровидных амфор, что указывает на тесное взаимодействие между этими группами. Предыдущие генетические анализы нескольких людей из зоны контакта предполагали, что эти контакты были ограничены культурным влиянием, а не примесью. Однако, в свете новых генетических данных, похоже, что эта зона — и, возможно, другие подобные зоны контакта между культурой шаровидных амфор и группами из степи (как из ямной культуры) — были ключевыми в формировании культуры шнуровой керамики, благодаря которой генетические компоненты, связанные со степью, и культурой шаровидных амфор, распространились далеко на запад и север, что привело к регионально разнообразным ситуациям взаимодействия и смешения.
При этом распространение культуры шнуровой керамики происходило преимущественно благодаря связям, которые были выстроены культурой шаровидных амфор в предшествующий период. Но несмотря на сходство по аутосомным данным, различные линии гаплогрупп Y-хромосомы у представителей культур шнуровой керамики и ямной предполагают, что известные на данный момент образцы представителей ямной культуры не подходят в качестве непосредственного источника степной родословной у носителей культуры шнуровой керамики.
Это предположение было подтверждено моделированием предковых компонент у европейцев со степным происхождением возрастом от 3 тыс. лет назад с участием представителей родственных ямной и афанасьевской культур, потому как модели показывают, что представители афанасьевской культуры, хоть и незначительно, но оказались более лучшим источником степной родословной у европейцев, несмотря на разное географическое распространение.
Стабильность популяций охотников-собирателей к востоку от Урала
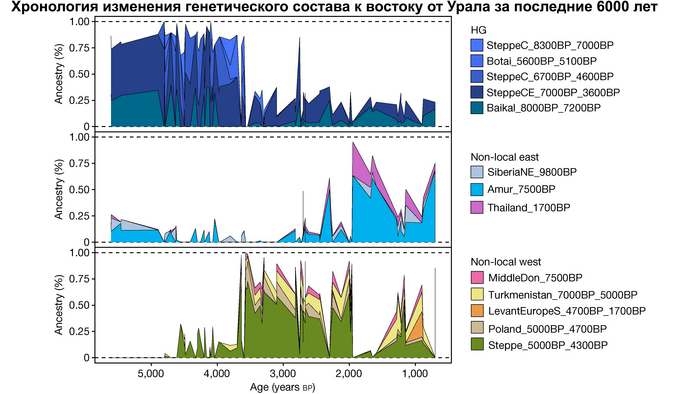
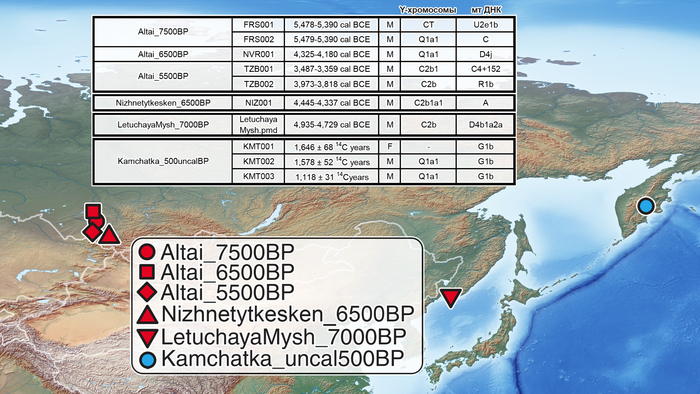
В отличие от значительного числа геномов древних охотников-собирателей из Западной Евразии, которые были изучены до настоящего времени, геномные данные охотников-собирателей к востоку от Урала были скудны. Эти регионы характеризуются ранним появлением керамики из районов, расположенных дальше к востоку, они были населены сложными обществами охотников-собирателей с постоянными, а иногда и укрепленными поселениями. В данной работе сообщается о 38 новых образцах, 28 из которых от охотников-собирателей, использующих керамику 8300-5000 лет назад. Большинство этих геномов попадает на неолитическую степную клину, которая охватывает сибирские лесостепные зоны бассейнов рек Иртыш, Ишим, Обь, Енисей, а также озера Байкал.
Эти охотники и собиратели могут быть смоделированы как смесь компонент из трёх генетических кластеров:
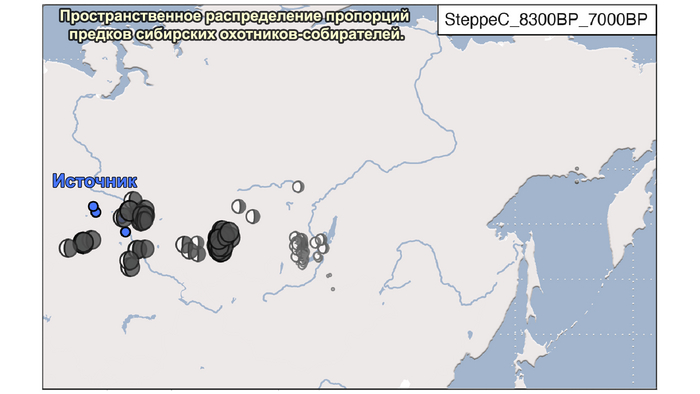
1. Компонента западносибирских охотников-собирателей возрастом от 8300 до 7000 лет (SteppeC_8300BP_7000BP), доминировала в западной части лесостепной зоны.
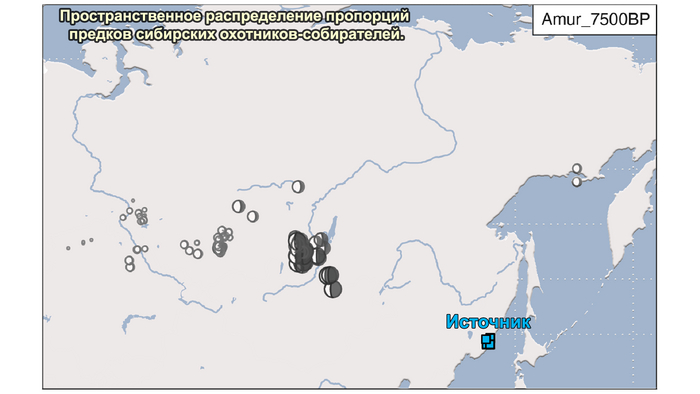
2. Компонента охотников-собирателей Северо-Восточной Азии возрастом около 7500 лет (Amur_7500BP) или древних северо-восточных азиатов из пещеры Чёртовы ворота в Приморье, была максимально выражена у популяций Байкала.
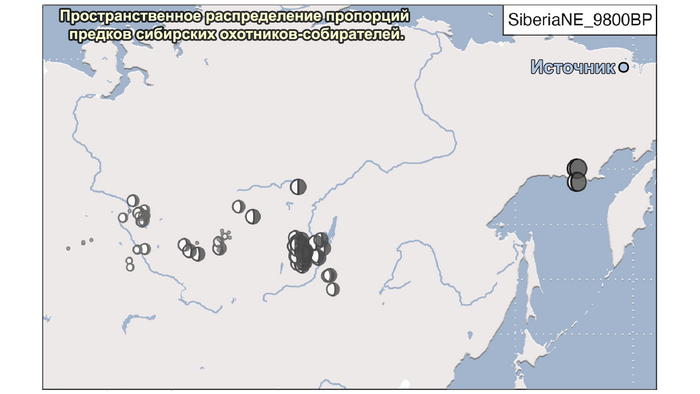
И 3. Компонента представлена палеосибирской родословной возрастом 9800 лет (Siberian_9800BP) как у обитателя Дуванного Яра в низовьях реки Колымы, она снижались по мере удаления от озера Байкал на запад.
1/5
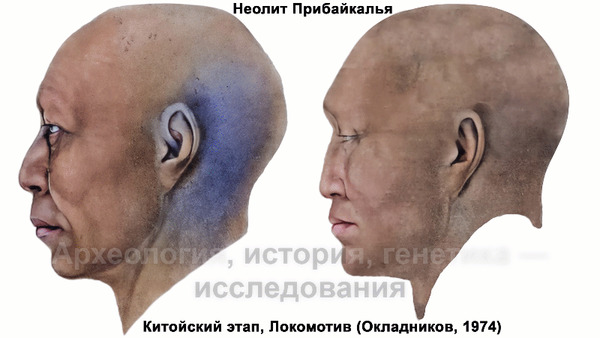
Авторы использовали кластеры также и для исследования пространственно-временной динамики генетического состава в степи и районе озера Байкал после неолита. Результаты подтверждают увеличение компоненты охотников-собирателей лесостепи возрастом от 7000 до 3600 лет (SteppeCE_7000BP_3600BP) у жителей Байкала позднего неолит-ранней бронзы (кластеры Baikal_5600BP_5400BP и Baikal_4800BP_4200BP). Однако родословная, относящаяся к этому кластеру, уже наблюдалась у вновь описанных неолитических охотников-собирателей как у озера Байкал (NEO199 и NEO200), так и вдоль Ангары на севере (NEO843) около 7000 лет назад. У двух мужчин Байкала была определена гаплогруппа Y-хромосомы Q1b1, характерная для последующих групп позднего неолита-ранней бронзы из того же региона. Что совместно с расчётной датой примеси около 7300 лет назад, позволяет предположить, что поток генов между охотниками-собирателями Байкала и лесостепей Южной Сибири уже был во времена раннего неолита, что согласуется и с археологическими данными.
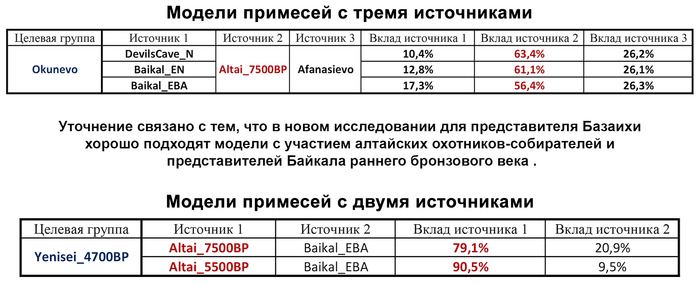
Новые образцы также проливают свет на генетическое происхождение носителей окуневской культуры раннего бронзового века в Минусинской котловине на юге Сибири. Её представителей можно смоделировать как смесь сибирских охотников-собирателей лесостепной зоны из кластеров возрастом 6700-4600 и 7000-3600 лет (Steppe_6700BP_4600BP и SteppeCE_7000BP_3600BP) со степными скотоводами возрастом от 5000 до 4300 лет (Steppe_5000BP_4300BP), связанными исключительно с афанасьевской культурой, время этой примеси датировано около 4600 лет назад.
1/2
А примерно 3700 лет назад жители в степном регионе и у озера Байкал демонстрируют уже заметно отличающиеся генетические профили. Ранние стадии этого перехода характеризуются притоком предков, связанных со степью, количество которых снижается со временем, от 70% на начальном этапе.
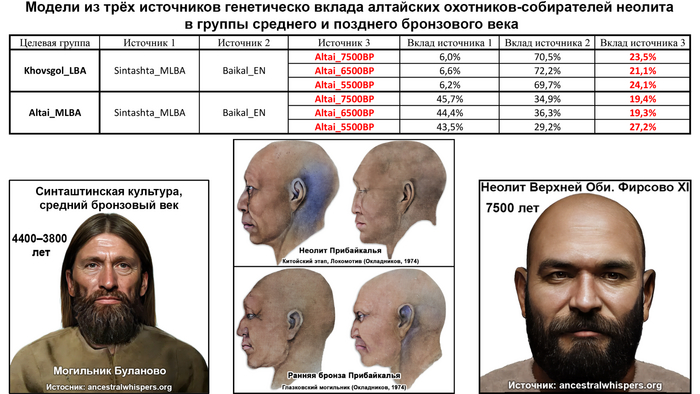
Аналогично динамике в Западной Евразии, происхождение, связанное со степью, здесь соотносится с таковым как у представителей культуры шаровидных амфор с территории современной Польши, возрастом 5000-4700 лет, подтверждая ранее задокументированный поток генов от групп из культуры шаровидных амфор в соседние группы степи и лесостепи, а также поток этой уже смешанной с европейскими земледельцами родословной дальше на восток уже с синташтинской и андроновской культурами в бронзовом веке. Однако у людей из окуневской культуры отсутствует вклад от людей культуры шаровидных амфор, а люди со степным происхождением начиная от 3700 лет назад демонстрируют чуть большую близость к ямной, чем к афанасьевской культуре, что указывает на две волны миграций западных степных скотоводов на восток. Первая волна в раннем бронзовом веке была связана с ямной культурой, а вторая, позже, была связана с синташтинской и андроновской культурами. Более поздние этапы переходного периода характеризуются увеличением родословной, связанной с иранскими земледельцами Центральной Азией (Turkmenistan_7000BP_5000BP) и древними северо-восточными азиатами Приморья (Amur_7500BP).
Суммарно, эти результаты показывают, что на востоке Евразийской степи родословная охотников и собирателей со сложной генетической структурой сохранялась и преобладала значительно дольше, чем в западной Евразии. Пока менее чем 4 тыс. лет назад по степи не прокатились волны миграций, которые привнесли в регион одомашненные породы лошадей с последующим появлением нового снаряжения для верховой езды и колесницы со спицами. Также эти миграции способствовали распространению проса в качестве сельскохозяйственной культуры.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
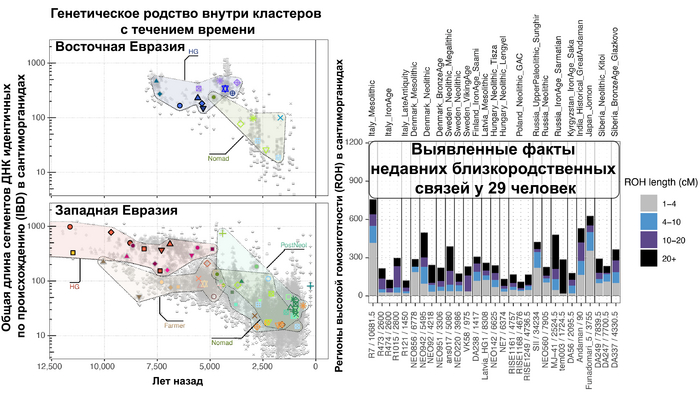
Закономерности распределения сегментов ДНК идентичных по происхождению между людьми, продемонстрировали снижение родства внутри кластеров с течением времени как в западной, так и в восточной частях Евразии. Что согласуется с увеличением эффективного размера популяции в течение этого времени. Однако наблюдаются заметные демографические различия между западной и восточной Евразией. Если на западе во время перехода к неолиту около 9000-6000 лет назад, люди с родословной анатолийских земледельцев демонстрируют снижение доли сегментов ДНК идентичных по происхождению, по сравнению с охотниками-собирателями, то на востоке генетическое родство оставалось высоким примерно до 4000 лет назад, что согласуется с гораздо более длительным существованием небольших локализованных групп охотников-собирателей.
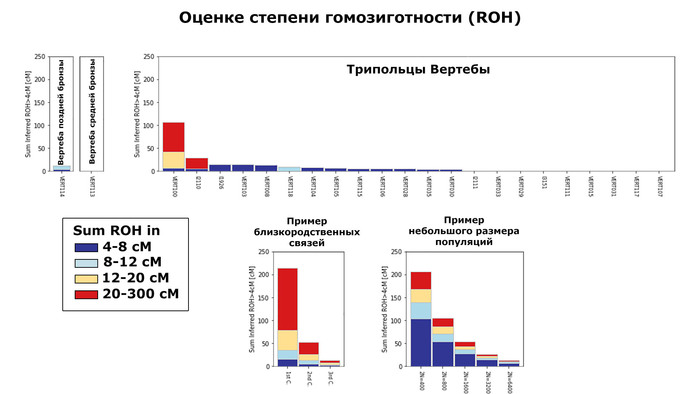
Помимо этого, анализ сегментов гомозиготности (ROH) не выявил значительного числа близкородственных связей, только 29 из 1396. При этом каких-то пространственно-временных или культурных закономерностей в распространенности близкородственных связей обнаружено не было. Зато авторы задокументировали первый случай однородительской дисомии по древнему геному (когда человек получает две копии хромосомы или части хромосомы от одного родителя и ни одной копии от другого родителя). Эта особенность обнаружена у представителя из Темясовских курганов позднесарматской культуры возрастом около 1700 лет.
Также были выявлены люди с различной степенью родства.
Итоги
Появление и сокращение генетического разрыва
После максимума последнего оледенения около 25-19 тыс. лет назад во время расселения древних людей по Евразии, установилось чёткое генетическое разделение между востоком и западом по линии от Черного моря до Балтийского, что отражено и в археологических данных. Это разделение сохранялось на протяжении всего мезолита и неолита. В Восточной Европе распространение земледелия происходило с опозданием примерно на 3000 лет, и эта задержка может быть связана с особенностями окружающей среды, поскольку регионы к востоку от условной границы имеют более континентальный климат и более суровые зимы, возможно, менее подходящие для ближневосточных методов ведения земледелия. На этих территориях сохранились высокоразвитые сообщества охотников-собирателей со стабильными, сложными, а иногда и укрепленными поселениями, с обменом на большие расстояния и крупными кладбищами. Рацион, включающий пресноводную рыбу, зафиксирован как по изотопным данным из этого исследования, так и по анализам остатков липидов в керамике.
В северных лесных районах этой пограничной зоны сообщества охотников-собирателей сохранялись вплоть до появления культуры шнуровой керамики около 5 тыс. лет назад, тогда как в южных и восточных степных регионах охота и собирательство со временем были дополнены животноводством с крупным рогатым скотом и овцами и, возможно, коневодством в Центральной Азии, но этот факт ещё под вопросом. Некоторые из этих групп, такие как представители хвалынской культуры Северного Кавказа и Среднего Поволжья, стали свидетелями появления мужских сообществ, вовлеченных в широкомасштабные обменные связи на основе медных изделий с Восточной и Центральной Европой, а также Кавказом. Поселения в то время были приурочены в основном к равнинным поймам и долинам рек, тогда как степной пояс оставался в значительной степени неиспользованным.
Окончательное исчезновение этой генетической, экономической и социальной границы было вызвано событиями, развернувшимися в степном регионе. Здесь археологически можно наблюдать две временные фазы технологических инноваций: широкое распространение колесных повозок, запряженных волами, около 5500 лет назад и более позднее развитие колесниц со спицами и верховой езды.
Начиная ещё с первой фазы инноваций, в сочетании с возможными изменениями условий окружающей среды, степь стала выгодной экономической зоной, которая позволила группам по типу ямной культуры эксплуатировать степь в качестве кочевых степных скотоводов около 5000 лет назад. А поселения эпохи энеолита вдоль речных долин были заменены этой новой формой мобильной экономики на основе скотоводства, которая окончательно разрушила генетическую границу, существовавшую в предыдущие тысячелетия.
Далее, после второй фазы инноваций около 4 тыс. лет назад, изобретение боевых колесниц и использование проса в качестве продовольственной культуры позволили андроновцам и родственным им группам окончательно распространиться на восток в Среднюю Азию и за ее пределы, оставив глобальное наследие в виде распространения индоиранских языков.
Это исследование внесло свою лепту в наши знания о прошлом, особенно касательно степных миграций.
Во-первых, авторы выявили до сих пор неизвестный источник родословной, который способствовал появлению степных скотоводов, на его роль хорошо подходили охотники-собиратели Среднего Дона, как люди из могильника мариупольского типа Голубая Криница раннего энеолита на юге Воронежской области, возрастом около 7300 лет.
А во-вторых, было определено, что степная родословная, которая распространилась по Европе, благодаря представителям культуры шнуровой керамики, сформировалась в результате культурного и генетического слияния степных групп, родственных ямной культуре и культуре шаровидных амфор. При этом благодаря своему смешанному культурному и генетическому происхождению, носители культуры шнуровой керамики практиковали смешанную экономику, используя различные стратегии выживания в различных условиях. Такая гибкость в значительной степени способствовала их успеху в расселении и адаптации к очень разным экологическим и климатическим условиям за очень короткий период времени.
Источник: Allentoft, M.E., Sikora, M., Refoyo-Martínez, A. et al. Population genomics of post-glacial western Eurasia. Nature 625, 301–311 (2024). https://doi.org/10.1038/s41586-023-06865-0
Взять с собой побольше вкусняшек, запасное колесо и знак аварийной остановки. А что сделать еще — посмотрите в нашем чек-листе. Бонусом — маршруты для отдыха, которые можно проехать даже в плохую погоду.
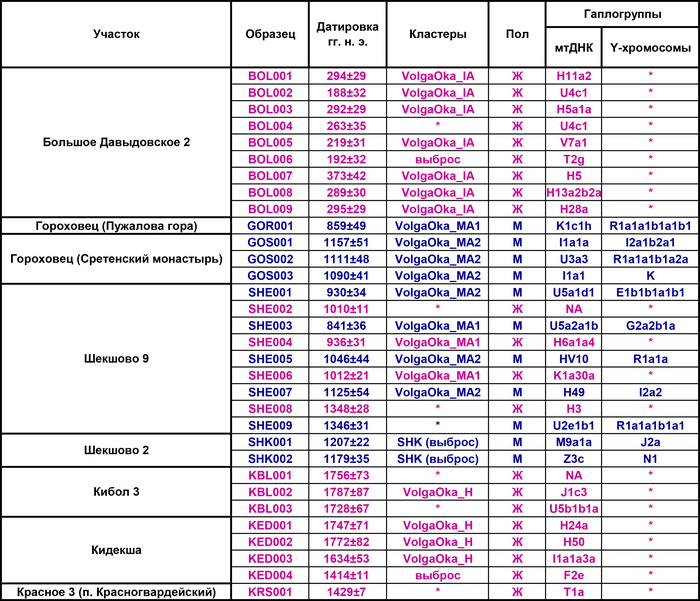
С развитием генетики, интерес вызывают не только вопросы о происхождении популяций, но также и отдельных людей, особенно если они принадлежали знатному роду. В частности, вопросы о происхождении элиты средневековой Руси IX–XVI вв., где важнейшими действующими лицами были представители правящего рода Рюриковичей, долгое время были предметом дискуссий. Ранее отцовские линии в виде гаплогрупп Y-хромосомы Рюриковичей были реконструированы по биологическим материалам современных людей, чьё происхождение, c разной степенью достоверности восходит к Рюрику. А что касается исследований древних геномов, то их осложняют трудности в идентификации останков Рюриковичей и другой знати. Поскольку до начала XV в. имена погребенных отсутствовали на погребальных сооружениях, надгробиях и саркофагах. Места погребений той эпохи устанавливаются по летописным сообщениям и спискам имён для поминовения XVI–XVII вв. Помимо этого практики повторного использования старых погребальных сооружений, перемещение почитаемых останков и их изъятие в ходе антирелигиозной кампании в советское время, привели к тому, что значительная часть останков XI–XIV вв. из погребений в древнерусских храмах и не только, оказалась утрачена или не может быть достоверно связана с определенными историческими личностями.
1/2
Рака с мощами св. Александра Невского
К примеру, в 1967 году на территории Черниговского Спасо-Преображенского собора во время строительных работ был случайно обнаружен череп мужчины 25-35 лет со следами ран от меча, без археологического документирования. Останки были предположительно отнесены одному из внуков Ярослава Мудрого - Глебу Святославичу, князю тмутараканскому и новгородскому, жившему в 11 веке. Однако точно определить кто был похоронен в каменном саркофаге возле Черниговского Спасо-Преображенского собора не представляется возможным. Особенно в свете недавнего анализ ДНК этого человека, который показал, что он был носителем гаплогруппы Y-хромосомы I2a1a2b1a1a и обладателем митохондриальной линии H5a2a.
Датировки (только археологические): Саркофаг найден на заднем дворе собора. Глубина верхней плиты саркофага составляла 1,9-2,0 м от современной поверхности. Стратиграфия несколько пострадала от более позднего нарушения грунта: трубы водоснабжения проложены на высоте 0,2 м над изголовьем саркофага. Археологи относят его к XI веку нашей эры. гаплогруппа – Y-хромосомы I2a1a2b1a1a гаплогруппа мтДНК – H5a2a
Поэтому особую ценность представляют те немногочисленные захоронения, где костные останки по археологическому контексту, антропологическим данным и комплексу исторических свидетельств надёжно отнесены к представителям рода Рюриковичей.
К таким достоверным захоронениям относится погребение в Спасо-Преображенском соборе Переславля-Залесского князя Дмитрия Александровича, второго сына Александра Ярославича Невского. Прямые сообщения о погребении Дмитрия Александровича в Спасо-Преображенском соборе содержит ряд летописей, в том числе Новгородская четвёртая, Воскресенская, Никоновская летописи, а также Московский великокняжеский свод конца XV века.
А поиск места погребения был облегчен ещё и тем фактом, что после захоронения князя Дмитрия Александровича в 1294 году и его наследника бездетного старшего сына Ивана в 1302 году с последующим переходом Переяславского княжества Москве, этот храм как престижная усыпальница использовался реже чем многие другие в средневековье. Изучив все доступные документы, можно было полагать, что погребение Ивана Дмитриевича находилось в юго-западной части собора, под хорами, а погребение князя Дмитрия Александровича – в юго-восточной части, в южной алтарной апсиде, где до реставрации 1891 г. находились отмечавшие места погребений более поздние кирпичные надгробия XVII в. В результате сам саркофаг с останками, был обнаружен отрядом Института археологии РАН под руководством Владимира Валентиновича Седова в 2014 г.
1/2
Князь Дмитрий Александрович унаследовавший после смерти великого князя Александра Ярославича Невского – Переславское княжество, в разные годы также был новгородским князем и великим князем владимирским. Князь умер в 1294 году на Ламском Волоке, возвращаясь в Переславль из Твери, и был похоронен в Спасо-Преображенском соборе Переславля.
Результаты
Палеогенетический анализ
В результате полногеномного секвенирования фрагментов древней ДНК из наиболее информативного образца - ладьевидной кости стопы Nev2.3, сгенерировано более 15 млн чтений, определены 532154 однонуклеотидных полиморфизма (ОНП). В процессе обработки данных секвенирования установлено, что это был мужчина с митохондриальной гаплогруппой F1b1 и гаплогруппой Y хромосомы N1a1a1a1a1a1a7a~ N-Y59140.
1/2
Анализ Y-хромосомы князя Дмитрия Александровича и других предполагаемых представителей рода Рюриковичей
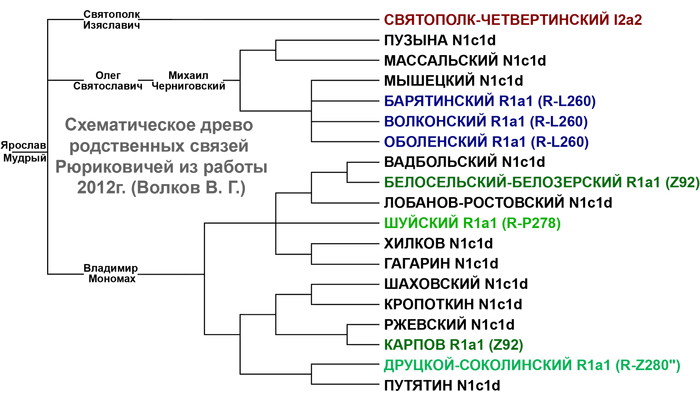
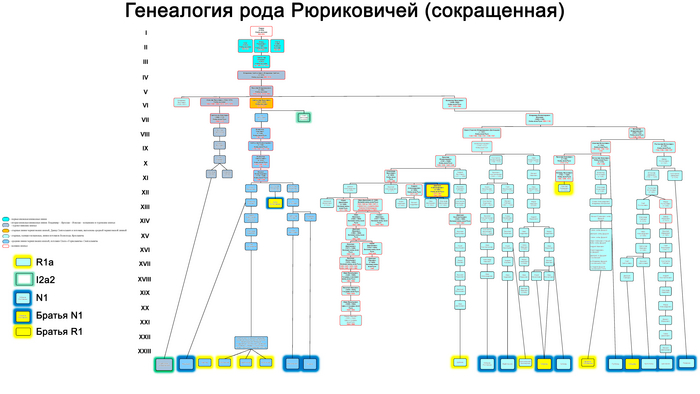
На основании результатов генетических исследований трех древних потомков Рюрика и предполагаемых 43 современных Рюриковичей выдвинуты гипотезы о трех возможных отцовских линиях Рюрика, а именно N1a, R1a и I2a.
В результате филогенетического позиционирования Y хромосома князя Дмитрия Александровича кластеризовалась с Y хромосомами современных вероятных потомков Рюриковичей, принадлежащих к гаплогруппе N1a, которые произошли из различных аристократических родов: Мстиславичей и Юрьевичей, потомков Мономаха от великого князя Мстислава Владимировича и великого князя Киевского Юрия Долгорукого, а также Ольговичей, черниговских Рюриковичей, потомков князя Олега Святославича или «Гориславича».
1/2
При этом важно отметить, что вероятные захоронения князя Глеба Святославича Черниговского, из рода Ольговичей, с линией I2a и Изяслава Ингваревича Луцкого, из рода Мстиславичей, с линией R1a, не подтверждены археологическими данными, что ставит под сомнение и гипотезы, которые вытекают из генетического анализа указанных образцов.
В целом как древние, так и современные предполагаемые Рюриковичи с линиями гаплогрупп R1a и I2a, обладают высокой гетерогенностью нуклеотидной последовательности Y хромосом и не указывают на единое происхождение по прямой мужской линии. В то время как большинство ныне живущих потомков Рюрика по прямой мужской линии согласно их родословным, с гаплогруппой N1a, обладают максимально похожими вариантами Y хромосомы как между собой, так и с Y хромосомой князя Дмитрия Александровича. Что совместно с полногеномными данными средневековых и современных Рюриковичей указывает на то, что, как минимум, со времени великого князя Ярослава Мудрого (XI в), мужчины этого рода были носителями гаплогруппы Y-хромосомы N1a.
Поиск живших в прошлом людей наиболее близких к князю Дмитрию Александровичу по Y-хромосоме
На филогенетическом дереве Y хромосомы видно, что помимо современных Рюриковичей, линия Дмитрия Александровича группируется с древними людьми середины II тысячелетия до н.э. Оленеостровского могильника в Кольском заливе Баренцева моря.
В предыдущих работах на примере этих образцов был показан поток генов из Сибири на север и восток Европы (восточно-евразийская компонента).
1/3
Высокая степень биологического соответствия Y-хромосомы представителя русского княжеского рода и людей эпохи раннего металла привела авторов работы к гипотезе о возможном вкладе восточно-евразийского генофонда в формирование североевропейского населения раннего средневековья, поскольку, согласно летописям, Рюриковичи имели с ними генетическую связь.
Авторы работы исследовали вклад людей с Большого Оленьего острова в формирование средневекового населения, жившего вдоль побережья Балтийского моря из современных Финляндии, Дании, Швеции и Норвегии, используя геномы периода «викингов» из работы 2020 года, обзор которой тоже есть на канале. В статистических анализах удалось выявить существенный вклад геномов древних представителей Большого Оленьего острова в формирование генофонда популяций периода «викингов», скорее всего, благодаря контактам последних с финно-угорским населением прибалтийского региона. При этом у представителей периода «викингов» с гаплогруппой N1a этот вклад выше чем у носителей линий R1a. Результаты показывают, что в формировании генофонда отдельных жителей из Швеции, Норвегии и Балтийского региона участвовали и люди из Сибири с восточноазиатскими корнями. При этом в наиболее подходящих моделях, помимо европейских охотников-собирателей, неолитических земледельцев и степных скотоводов, скандинавские популяции моделировались с участием обитателей Большого Оленьего острова или представителей хунну железного века.
Здесь стоит адекватно воспринимать компоненты в моделях. В данном случае хунну добавлены для проверки географического происхождения компоненты из Сибири, если говорить упрощённо.
А что касается предков князя Дмитрия Александровича по мужской линии, то с большой вероятностью, среди них были мужчины, оставившие могильник на Большом Оленьем острове в Кольском заливе Баренцева моря около 3600 лет назад.
Анализ мтДНК князя Дмитрия Александровича
Митохондриальную гаплогруппу F1b1, определенную у князя Дмитрия Александровича относят к восточно-евразийскому кластеру, она встречается с разной частотой у большинства исследованных древних и современных популяций Байкальского региона и сопредельных территорий Центральной Азии.
Погребения мужчин с митохондриальными линиями F1b1b и F1b1f, а также с линией Y-хромосомы N1a1a1a1a3a (F4205) характеризовались довольно богатым погребальным инвентарем с золотыми и позолоченными предметами, что указывает на их принадлежность к знати.
● A1811 – Женщина, захороненная с костями овец или коз. мтДНК F1b1f ● A1812 – Мужчина из богато обставленной могилы, с золотыми серьгами, золотыми заколками для волос, ремешками из золотой фольги с переплетающимися узорами из серебра и мечом, украшенным золотыми пластинами. мтДНК – F1b1f, Y-хромосомы – N1a1a1a1a3a (F4205) ● A1821 – Пожилой мужчина, захороненный с исключительно богатыми погребальным инвентарём, включающим позолоченный меч и пояс. мтДНК – F1b1b, N1a1a1a1a3a (F4205)
Однако с уверенностью говорить о происхождении материнской линии князя Дмитрия Александровича довольно сложно, так как практически во всех исторических периодах прослеживается повышенная изменчивость и разнообразие митохондриальных гаплогрупп в одном регионе.
Здесь также важно понимать разницу между генофондом популяций и отдельно взятых людей, особенно представителей знатных родов, у которых была широко распространена практика привлечения самых разных по своему происхождению женщин для союзов, официального и неофициального характера, в качестве выгодных или вынужденных политических шагов. Поэтому материнская линия F1b князя Дмитрия Александровича могла быть связана как с древними миграциями из Сибири, так и с миграциями кочевников раннего средневековья. При этом у этих разных по времени групп могло быть общее происхождение этих линий.
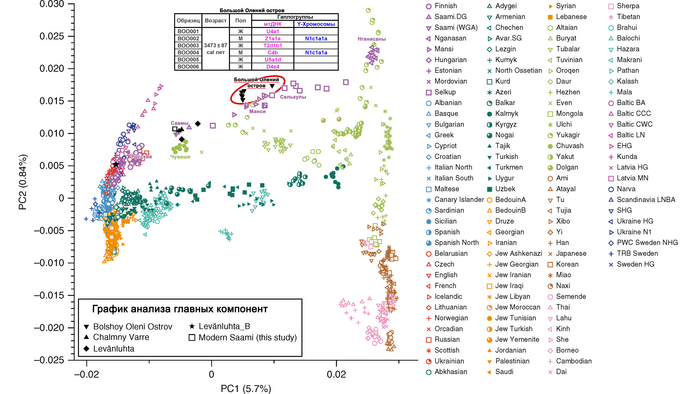
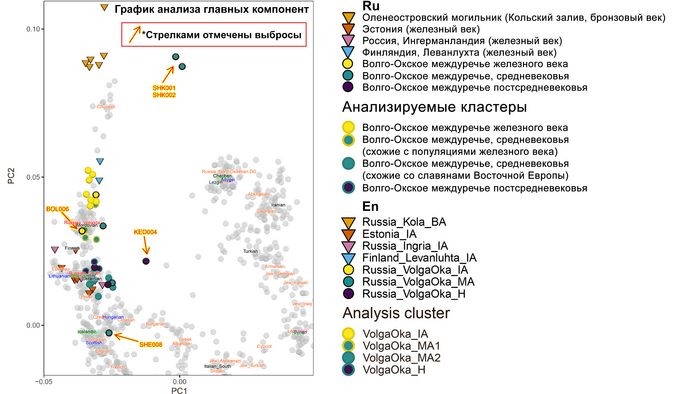
Результаты анализа главных компонент
Генетическую близость генома князя Дмитрия Александровича к другим известным древним и современным популяциям оценили с помощью метода главных компонент (PCA).
Сразу стоит отметить, что надо адекватно понимать положение на этом графике отдельных представителей популяций, особенно из знатного рода. Если положение большинства образцов сочетается с их географией, то положение отдельно взятых людей от смешанных союзов будет где-то между положением его родителей. При этом генетическая схожесть в этом анализе не подразумевает общую историю, а лишь схожие пропорции генетических компонент.
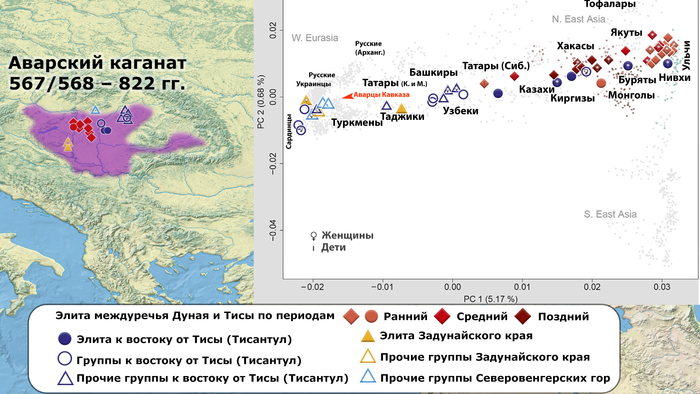
В нашем случае геном князя Дмитрия Александровича занимает промежуточное положение между древними и современными образцами из Европы и Центральной Азии. Наиболее близкими генетически и хронологически к князю Дмитрию Александровичу, расположены некоторые представители авар позднеаварского периода из междуречья Дуная и Тисы, а также из предполагаемого вторичного центра власти Аварского каганата к востоку от Тисы.
Здесь стоит напомнить, что в Аварском каганате (VI–IX вв.), помимо европеоидного большинства ярко выраженный монголоидный тип, имели, преимущественно, представители элиты, который размывался со временем к позднеаварскому периоду.
Как известно из работы 2022 года, популяции аварского периода демонстрируют значительную генетическую изменчивость, а их образцы разбросаны на графике анализа главных компонент от популяций Западной Евразии до популяций Северо-Восточной Азии.
Однако несмотря на эту общую неоднородность, существуют четкие закономерности генетической подструктуры, соответствующие как хронологии и географии Карпатского бассейна, так и социально-археологическим категориям. Почти все представители элиты раннеаварского периода образуют плотный кластер с высоким содержанием компоненты «древних северо-восточных азиатов» (АНА) до 98%. Однако образцы представителей элиты позднеаварского периода уже сильнее смещены в сторону Западной Евразии, а количество родословной «древних северо-восточных азиатов» у них сократилось уже до 70-80%.
При этом среди простых жителей раннего и позднего аварского периодов преобладал более западный генетический профиль, демонстрирующий сильную связь с населением Карпатского бассейна доаварского периода, с нулевым или небольшим вкладом компоненты «древних северо-восточных азиатов». И как раз именно к этой группе образцов относится расположенный рядом с князем Дмитрием Александровичем представитель позднеаварского периода со смешанным генетическим профилем (Hungary_LateAvar), примерно, на 80% состоящим из компоненты максимально выраженной у предшествующих местных жителей Карпатского бассейна и на 20% из компоненты «древних северо-восточных азиатов» из восточной степи.
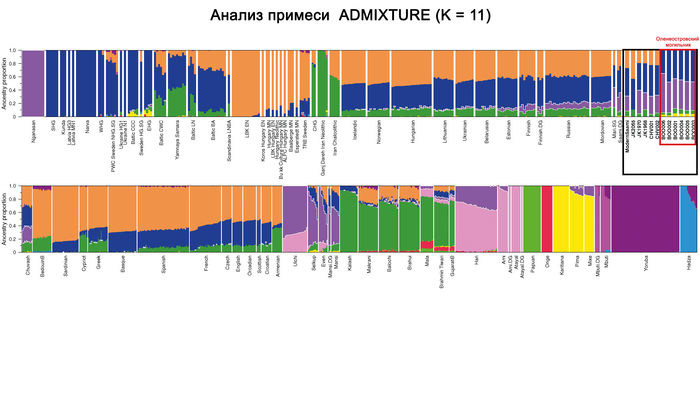
Анализ примесей (Admixture)
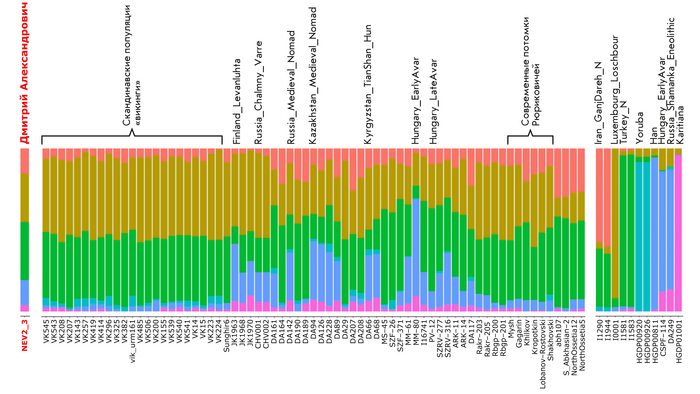
В анализе примесей, по предковым компонентам отмечено генетическое сходство князя Дмитрия Александровича с представителями раннесредневекового населения востока Скандинавии периода викингов.
Что может свидетельствовать в пользу версии о «варяжском» (скандинавском) происхождении Рюрика, родоначальника княжеского рода, призванного на Русь, о чем прямо говорит Летопись. При этом авторы, используя термин «викинги», подчёркивают, что это неоднородная и сложная по своему историческому формированию европейская популяция, объединенная лишь образом жизни и ареалом проживания. Однако есть и отличие генома князя от скандинавских популяций эпохи «викингов», в том числе с территории современной России, это отличие связано с наличием у него дополнительной восточно-евразийской компоненты (обозначенной синим цветом на графике), отсутствующей в таком количестве у подавляющего большинства «викингов». Эта компонента максимально выражена у нганасан, финно-угорского народа Западной Сибири манси, представителей народа хань в Китае, у элиты авар из междуречья Дуная и Тисы (Hungary_DanubeTisza_MLAvar), а также у более древних представителей эпохи раннего неолита Байкала и Монголии (Russia_Shamanka_Eneolithic. SG и Mongolia_North_N). В меньшей степени эта компонента обнаружена у жителей раннего средневековья с территории современной Финляндии (Finland_Levanluhta), у кочевника Прикаспийской степи (Russia_Medieval_Nomad), современника князя Дмитрия Александровича, а также у более древних ираноязычных степных кочевников железного века с территорий современных Казахстана и Кыргызстана (Kazakhstan_TianShan_Saka, Kyrgyzstan_TianShan_Hun). Суммарно, на основании данных анализа главных компонент, анализа примесей и информации о митохондриальной ДНК можно утверждать, что в геноме князя Дмитрия Александровича имеется существенный вклад восточной предковой компоненты, что отличает его от раннесредневекового населения востока Скандинавии, «викингов» и средневекового славянского образца из Владимира (Сунгирь6), но сближает в первую очередь с древним населением Финляндии, Кольского полуострова и раннесредневековым населением Центральной Европы с этой компонентой. Вероятно, этот вклад получен и по мужской, и по женской линиям, что соответствует процессам древней миграции из Сибири по степному евразийскому коридору в 1 тыс. до н.э.–1 тыс. н.э.
Моделирование генома князя Дмитрия Александровича из геномов предковых популяций
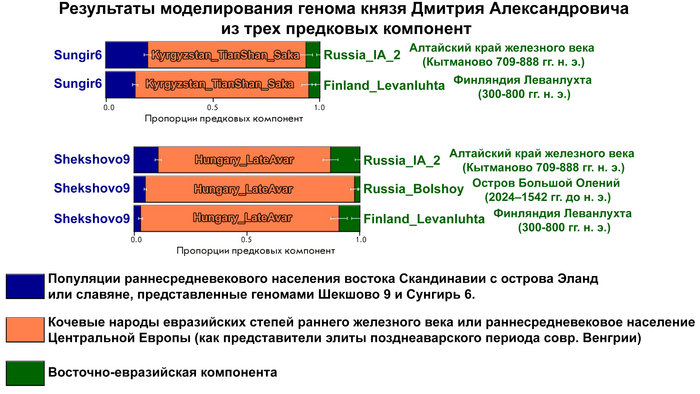
После анализа различных комбинаций предковых компонент наиболее подходящая модель указывает на вклад в происхождение князя Дмитрия Александровича трех компонент, как у следующих популяций:
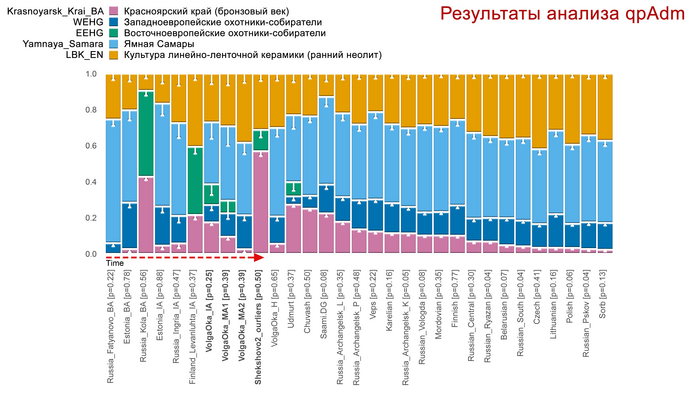
Раннесредневекового населения востока Скандинавии с острова Эланд. Или славян, представленных геномами Шекшово9 и Сунгирь6.
Представителей степных кочевых народов евразийских степей железного века или раннесредневекового населения Центральной Европы (степных кочевников с территории Венгрии).
Восточно-евразийской компоненты.
Предсказание фенотипических признаков князя Дмитрия Александровича на основании генетических данных
С большой вероятностью глаза князя Дмитрия Александровича были карими (P = 0.962), волосы темными (P = 0.810) или коричневыми (P = 0.555), а оттенок кожи промежуточным (0.635), т.е. не светлым и не темным.
Выводы
Благодаря палеогенетике и применению статистических методов проверки гипотез, подтверждается связь Рюриковичей со Скандинавией по мужской линии, хотя отсутствие генетических данных от населения с территории севера Руси в период, непосредственно предшествовавший становлению княжеского правления, не позволяет предметно судить о присутствии предков князя Дмитрия Александровича и других Рюриковичей на материковом севере и северо-востоке Европы. Анализ генеалогического древа Рюриковичей показал, что современные представители этого рода, с гаплогруппой Y-хромосомы N1a, группирующиеся с образцом князя Дмитрия Александровича, относятся к трем разным ветвям этого рода – Ольговичам, Мстиславичам и Юрьевичам. Которые восходят к князю Ярославу Мудрому, как к ближайшему их общему предку.
В свою очередь митохондриальная гаплогруппа F1b1 у князя Дмитрия Александровича может свидетельствовать о вкладе восточных популяций в его генотип, что подтверждается и аутосомными данными. Согласно результатам анализа главных компонент и анализа примесей, геном князя Дмитрия Александровича имеет сходство с геномами представителей скандинавских популяций эпохи «викингов», но отличается от них заметным вкладом восточной компоненты. Что может быть связано с распространенной практикой браков русских князей с дочерями половецких ханов с конца XI в. Такие браки скрепляли союзнические отношения и усиливали политическое взаимодействие.
Мать Дмитрия, супруга Александра Невского – Александра Брячиславна происходит из рода полоцких Изяславичей. Сведения о женах этих князей скудны, имя и происхождение матери Александры неизвестно. Однако, известно, что представители полоцкой ветви Рюриковичей не избегали брачных союзов с половчанками. Из половецкого рода происходила вторая жена полоцкого князя Святополка Изяславича (1050–1113), Елена, дочь хана Тугоркана.
Однако восточная компонента в геноме Дмитрия Александровича могла быть получена одновременно и по мужской, и по женской линиям, отражая браки Рюриковичей с представительницами династий Центральной и Южной Европы, как сербские Вукановичи или венгерские Арпады, к примеру. Элиты этих территорий могли в большей степени, чем «простое» местное население, сохранить восточные гены, привнесенные миграционными потоками первого тысячелетия нашей эры. Исходя из перечисленных вероятностей, объяснения присутствия «восточной компоненты» в геноме Дмитрия Александровича, пока остаются гипотетическими, как и состав его предков по материнской линии.
Но несмотря на оставшиеся вопросы, впервые на примере представителя древнего рода Рюриковичей показана генетическая составляющая сложного характера межэтнических взаимодействий в формировании знати средневековой Руси.
Источник: Жур К.В., Шарко Ф.С., Седов В.В., Добровольская М.В., Волков В.Г., Максимов Н.Г., Сеславин А.Н., Макаров Н.А., Прохорчук Е.Б. Рюриковичи: первый опыт реконструкции генетического облика правящего рода средневековой Руси по данным палеогеномики // Acta Naturae. - 2023. - Т. 15. - №3. - C. 50-65. doi: https://doi.org/10.32607/actanaturae.23425
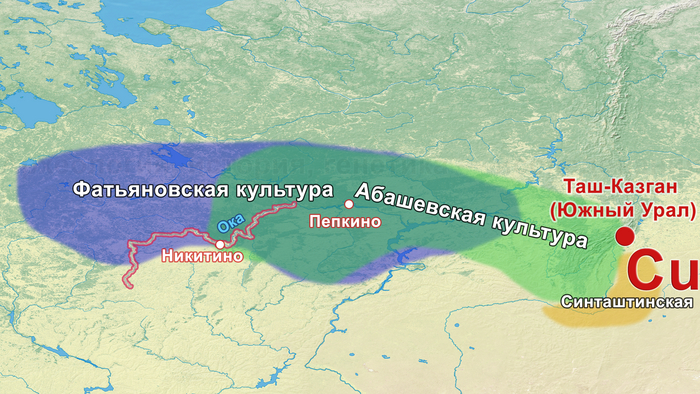
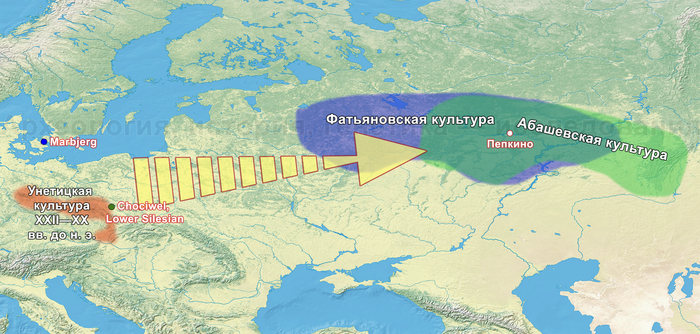
Приветствую! Сегодня речь пойдёт о научной работе, авторы которой рассматривают фундаментальный вопрос о происхождении и особенностях миграций носителей фатьяновской и средневолжской абашевской культур. Цель работы заключалась в том, чтобы на основании новых лабораторных данных о древней ДНК рассмотреть гипотезы о пришлом происхождении палеопопуляций фатьяновской и средневолжской абашевской археологических культур на Русской равнине.
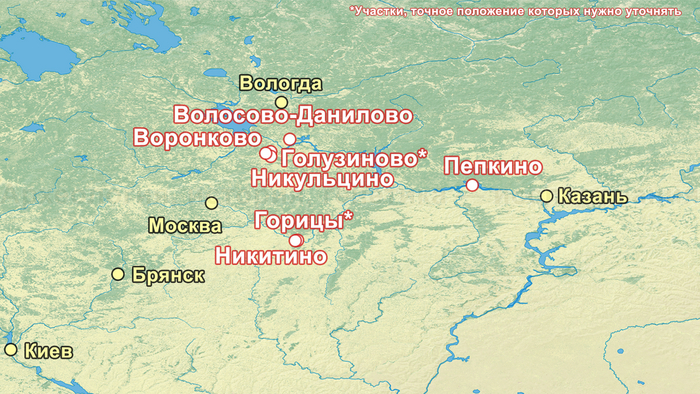
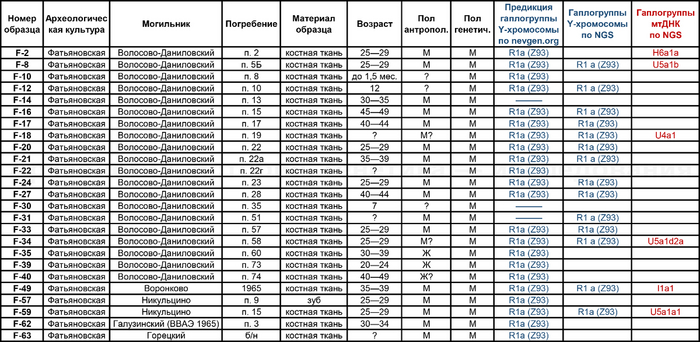
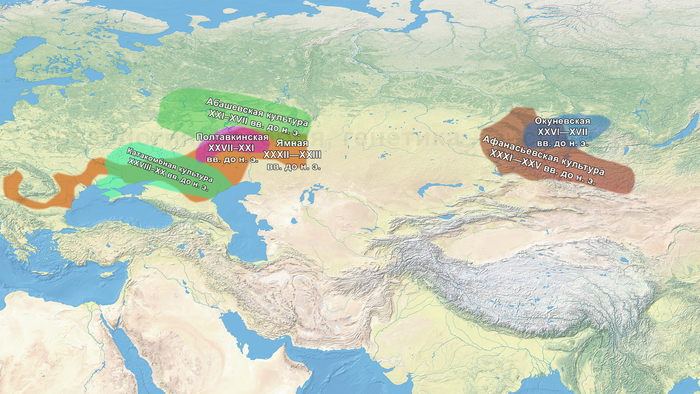
В исследовании были проанализированы образцы 25 человек, относящихся к фатьяновской культуре (20 образцов было получено от захороненных в пределах Волосово-Даниловского могильника, два из раскопок в Никульцино и по одному из Воронково, а также Голузинского и Горицкого могильников). Стоит отметить, что Волосово-Даниловский могильник — наиболее репрезентативный погребальный комплекс фатьяновской археологической культуры.
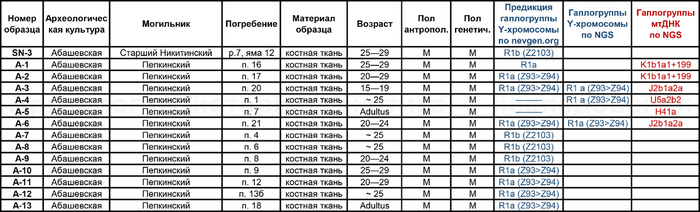
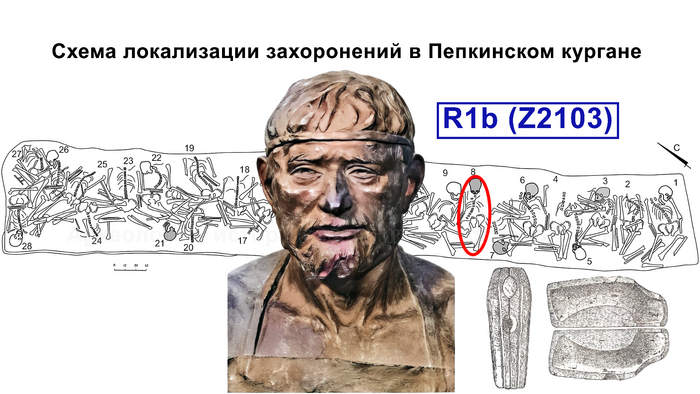
Помимо этого, впервые был проведен палеогенетический анализ 14 человек, относящихся к абашевской культуре (тринадцати человек из Пепкинского кургана и одного из Старшего Никитинского могильника). Пепкинский курган, открытый у деревни Пепкино, представляет собой коллективное захоронение группы молодых мужчин, не менее 27, средневолжской абашевской археологической культуры, погибших в бою.
Кстати, для исследования авторы работы намеренно отбирали представителей мужского пола разного возраста, так как для них возможен анализ однородительских маркеров как по мужской, так и по женской линиям.
Здесь сразу хотелось бы выразить благодарность авторам работы за их вклад в наше понимание прошлого.
Введение
Обнаружение в геномах европейских групп компонента восточных охотников и собирателей, соотносимого с генетическим наследием населения ямной археологической культуры, в своё время распространенной на обширных территориях евразийских степей, стало одним из наиболее спорных вопросов в российской археологии.
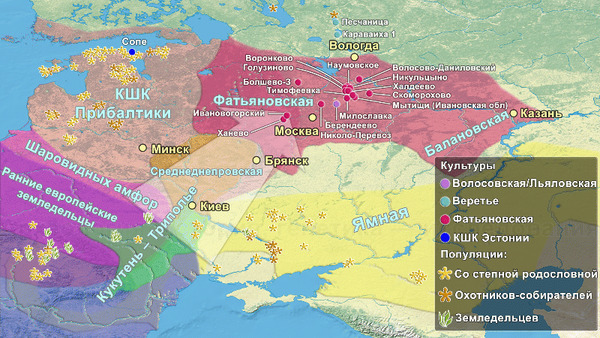
И хотя между распространением культуры и генов нельзя ставить знак равенства, генетический вклад от носителей ямной культуры зафиксирован во многих регионах Европы. Наличие этого степного генетического сигнала отличает представителей культур шнуровой керамики и колоколовидных кубков, а также более поздних жителей Европы эпохи бронзы от ранних неолитических земледельцев этого региона. При этом на территории европейской части современной России подобные генетические исследования до недавнего времени отсутствовали, пока в 2021 году не была опубликована работа, обзор которой также есть на канале, под названием: Генетическая история населения запада России при переходе от каменного века к бронзовому. В этом, более раннем, исследовании авторы пришли к выводу о зарождении носителей фатьяновской культуры благодаря миграции смешанного населения, потомков носителей ямной культуры и европейских ранних земледельцев.
Тогда авторы сообщали, что в моделях только две популяции, а именно из культуры шаровидных амфор Украины и Польши, а также трипольской культуры Украины, оказались потенциальными источниками для смешения со степными скотоводами с генетическими профилями как у представителей ямной культуры Самары или Калмыкии, для образования генетической структуры популяции фатьяновцев. Также была отмечена близость этих людей с представителями культуры шнуровой керамики.
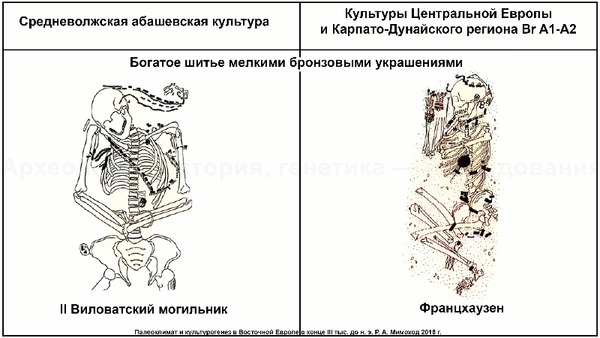
На основании данных археологии в последние годы уже высказывались предположения, что на территории Восточно-Европейской равнины в конце III тыс. до н. э. происходили масштабные миграции, с первоначальным импульсом из центральноевропейского региона. В пользу этого довода говорит схожесть керамического комплекса средневолжской абашевской культуры, следы которой выявлены непосредственно в Поволжье на территориях Брянской, Московской и Ярославской областей, с керамикой поздней фазы культуры колоколовидных кубков в Южной Германии.
Результаты
Анализ Y-хромосомы
По результатам микросателлитных исследований и анализа Y-хромосомы по методу секвенирования нового поколения (NGS) для образцов, относящихся к фатьяновской культуре, получена единственная гаплогруппа R1a (Z93).
Вероятно, такая однородность гаплогрупп Y-хромосомы связана с эффектами основателя и дрейфа генов в изолированной древней группе неместного происхождения.
Эффект основателя — вариант генетического дрейфа, при котором снижение и смещение генетического разнообразия происходит из-за заселения малым количеством представителей вида новой географической территории.
Дрейф генов — Изменение частот аллелей и генотипов в популяции, имеющее случайный характер. Такие случайные отклонения в частотах аллелей происходят из поколения в поколение, особенно при небольшом эффективном размере популяции.
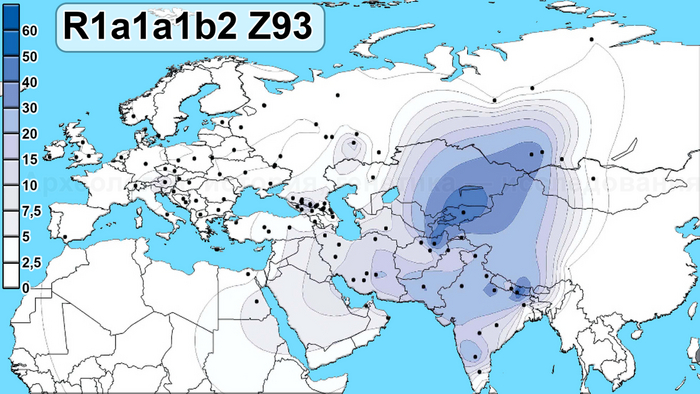
На данный момент линии гаплогруппы R1a1a1b2 Z93 с заметной частотой встречаются в Южной Сибири среди алтайцев и хакасов (>30 %), также сравнительно небольшие частоты встречаются среди татар Поволжья (5 %), в Кыргызстане (6 %) и во всех популяциях Ирана (1—8 %).
Среди археологических культур указанная гаплогруппа обнаружена, в частности, в сагсайской культуре XV—XII вв. до н. э. Субветвь этой гаплогруппы R1a1a1b2a Z94 представлена рядом образцов, относящихся к археологическим культурам средней бронзы центральной и западной части степного коридора Евразии — андроновской, полтавкинской, синташтинской и также сагсайской (XX—XV вв. до н. э.).
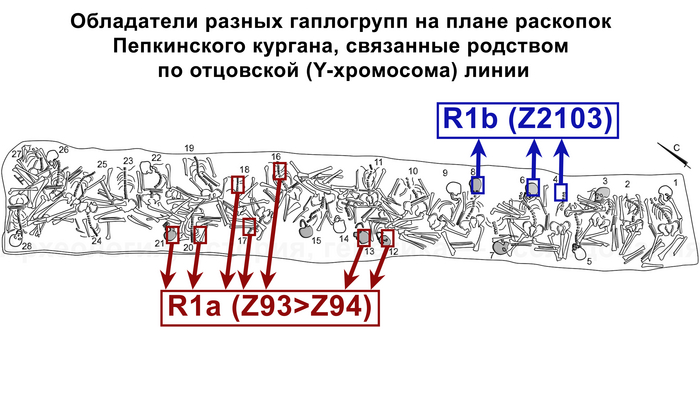
А вот для образцов абашевской культуры полученные результаты менее однородны. Для большинства образцов из Пепкинского кургана определена гаплогруппа R1a (Z93>Z94), однако у троих мужчин с этого участка была гаплогруппа R1b (Z2103).
Примечательно, что для абашевца захороненного на значительном удалении от Пепкинского кургана в Старшем Никитинском могильнике, был получен достаточно полный гаплотип, также предсказывающий принадлежность к той же гаплогруппе R1b (Z2103).
Указанная гаплогруппа была выявлена у мужчин, относящихся к ямной, афанасьевской, полтавкинской, катакомбной и окуневской культурам III—II тыс. до н. э.
Микросателлитные исследования в палеогенетике являются важным инструментом, особенно при исследовании групповых захоронений, поскольку позволяют выявить близость с точки зрения происхождения по мужской линии, а иногда и прямого родства, когда гаплотипы по всем считанным STR локусам совпадают, при этом результат тем точнее, чем выше число считанных локусов. Такие совпадения были найдены как для фатьяновцев погребенных в Волосово-Даниловском могильнике, так и для абашевцев из Пепкинского кургана, а также Старшего Никитинского могильника.
Что указывает на возможное близкое родство по отцовской линии людей с совпадающими STR локусами. А у людей, которые продемонстрировали минимальные отличия в значениях аллелей в одном или двух локусах, общий предок по отцовской линии мог жить несколькими поколениями ранее.
Анализ митохондриальной ДНК
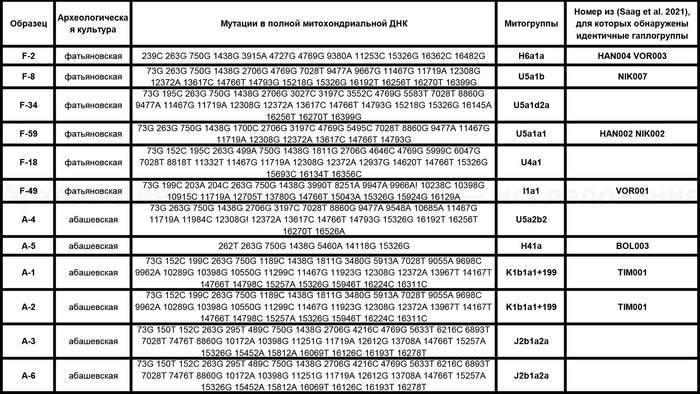
Материнские линии фатьяновцев были более разнообразны, чем отцовские. При этом основной гаплогруппой мтДНК была U с сублиниями — U5a1b, U5a1d2a и U5a1a1. А среди редких, так называемых минорных линий, определены H6a1a и I1a1
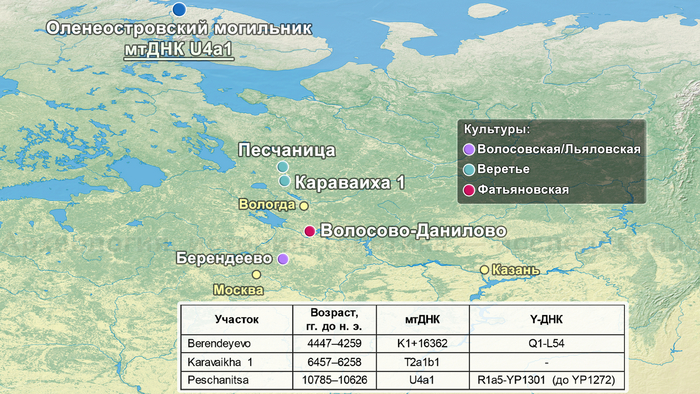
В большинстве линии гаплогрупп совпадают с задокументированными и в работе 2021 года, но с некоторыми дополнениями, как линии U5a1d2a и U4a1 у фатьяновцев из Волосово-Даниловского могильника. Однако в предыдущем исследовании линия U4a1 определена у одного из охотников-собирателей культуры Веретье эпохи мезолита, захороненного в могильнике Песчаница 1 на юго-западе Архангельской области. А ещё ранее эта линия была обнаружена у восточных и скандинавских охотников-собирателей. В частности, у охотников и собирателей эпохи неолита Большого Оленьего острова в Кольском заливе.
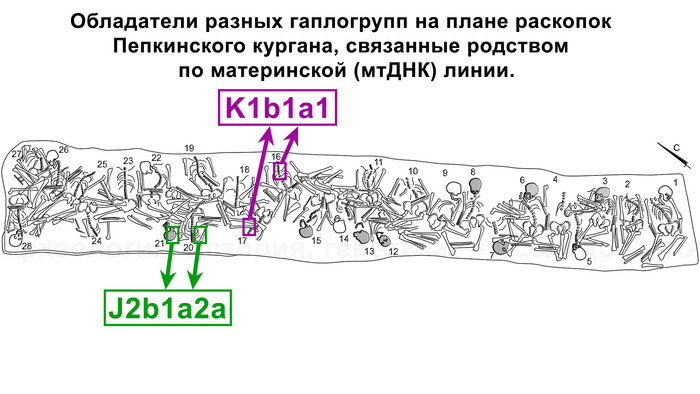
А что касается хронологически более поздних представителей абашевской культуры, то часть их линий, таких как U5a2b2, H41, ранее были обнаружены и у фатьяновцев. Однако неожиданностью для авторов стало наличие гаплогрупп J2b1a2a и K1b1a1+199 у абашевцев из Пепкинского кургана, ведь распространение этих линий связывают с неолитическими земледельцами. При этом пары с этими линиями имеют полностью совпадающий набор мутаций, что указывает на их родство по материнской линии.
Анализ аутосомных маркеров
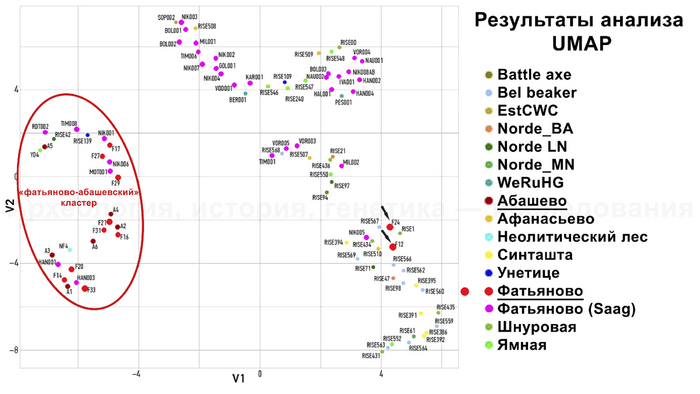
Сравнительный анализ аутосомных генетических вариантов с использованием метода понижения размерности проводился для 19 образцов в сочетании с ранее опубликованными данными 75 человек. По результатам анализа большая часть исследованных в новой работе образцов фатьяновской и абашевской культур объединились между собой в одном кластере.
Туда же вошли некоторые фатьяновцы из предыдущей работы. Неожиданный результат был получен для двух фатьяновцев из Волосово-Даниловского могильника, которые оказались ближе к скоплению образцов из контекста различных археологических культур эпохи бронзы, а именно из культур шнуровой керамики, колоколовидных кубков, афанасьевской и синташтинской. Что указывает на генетическую близость жителей эпохи бронзы, несмотря на их разную хронологию и географию.
Обсуждение и выводы
В новой работе у представителей фатьяновской культуры не обнаружено других отцовских линий, кроме R1a (Z93), что, вероятно, связано с эффектом основателя (когда небольшая группа отделяется от первоначальной популяции и формирует новую) и с дрейфом генов (когда одна линия сохранилась из-за случайных событий).
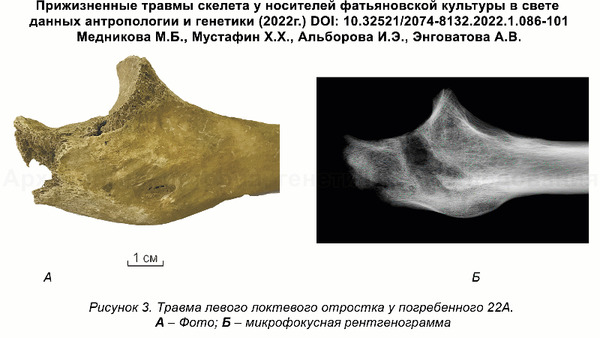
Это подтверждает миграцию группы представителей культуры шнуровой керамики из Центральной Европы и отсутствие связей с представителями местной волосовской культуры, что также упоминалась и в предыдущем исследовании, хотя его авторы использовали другие образцы и методы. Примечательно, что ранее, по результатам анализов травм левой руки в области локтевого сустава и кисти у мужчин из Волосово-Даниловского могильника, авторы исследования, опубликованного в 2022 году, пришли к выводу о схожей тактике боя у фатьяновцев Верхней Волги и шнуровиков Германии и Польши. Что служит независимым индикатором их принадлежности к единой культурной общности.
Помимо этого, совпадение митохондриальных гаплогрупп в обоих упомянутых генетических исследованиях также подтверждает гипотезу о неместном происхождении носителей фатьяновской культуры. При этом однообразный погребальный обряд фатьяновцев, различный для мужчин, женщин и детей, а также их низкое генетическое разнообразие, добавляют аргументов в пользу существования строгих правил внутри общества, которые можно объяснить его сплочённостью в связи с миграцией в иную среду. А учитывая некоторую близость по материнской линии захороненных в Волосово-Даниловском могильнике, с преобладанием ветвей гаплогруппы U, можно сделать вывод, что фатьяновцы переселялись семьями.
Датировка образцов из Волосово-Даниловского могильника указывает на его использование в течение более трёхсот лет, минимум с середины третьего тысячелетия до н. э. (интервал калиброванных дат: 2563-2037 гг. до н. э.).
При этом Волосово-Даниловский могильник всеми исследователями рассматривался как наиболее поздний.
А спустя несколько сотен лет после первого появления фатьяновцев возникла абашевская археологическая культура, в лесостепных районах среднего Поволжья и верхнего Дона, предположительно, основанная другими мигрантами. При этом Пепкинский курган, на котором были захоронены мужчины, погибшие в одном сражении, на данный момент является главным археологическим памятником средневолжской абашевской культуры. На первый взгляд это могли быть воины одного племени, потому как они захоронены в одной могиле и по схожему обряду, между 2130-1950 гг. до н. э.
Однако анализ древней ДНК выявил как минимум две группы разного происхождения. Экспертиза бронзовых изделий из Пепкинского кургана и Старшего Никитинского могильников показала, что по составу металла оба памятника относятся к единому этапу средневолжской абашевской культуры. А мышьяк и серебро в составе их металла, могут указывать на связь с рудными источниками Южного Урала. При этом для образца из Старшего Никитинского могильника была получена несколько более поздняя дата чем для Пепкинского кургана, отличающаяся на время жизни одного поколения абашевцев (3557 ± 31 BP).
Примечательно, что в Пепкинском кургане, один из абашевцев, погребенный с набором предметов кузнеца-бронзолитейщика с гаплогруппой Y-хромосомы R1b (Z2103), по данным изотопного анализа провел свои юные годы в геохимических условиях, которые соответствовали степным районам Южного Урала.
К тому же, известно о происхождении металла некоторых изделий, найденных в Пепкинском кургане, из Таш-Казганского месторождения меди.
При этом по соотношениям изотопов стронция в зубной и костной ткани другой группы мужчин из Пепкинского кургана, все они оказались «неместными» для территории современной республики Марий Эл, где они были похоронены. Примерно до 10 лет они жили в регионе соответствующему западному ареалу проживания абашевцев в бассейне Оки. Похожие значения характерны и для захороненных в пределах Старшего Никитинского могильника. Интересно, что абашевцы с гаплогруппой R1b(Z2103) были более мобильны, но при этом связаны достаточно тесным родством. Можно предположить, что именно эта группа в составе абашевского населения была тесно связана с металлургическим производством. Однако другая, более многочисленная часть мужчин из Пепкинского кургана с гаплогруппой R1a (Z93>Z94), очевидно, имела иное происхождение. Отцовские линии и аутосомные маркеры сближают их с фатьяновцами, прежде всего, из Волосово-Даниловского могильника. Однако неясно, связано ли происхождение абашевцев с фатьяновцами, что в целом не подтверждается археологически, или же с новой миграцией из Европы спустя несколько веков после миграции предков фатьяновцев. И хотя митохондриальные линии поддерживают обе гипотезы, отдельные линии обнаружение у некоторых людей из Пепкинского кургана, такие как J2b1a2a и K1b1a1+199, скорее, свидетельствуют о недавней миграции из Европы, где генетический след неолитических земледельцев был очень заметен.
Ранее была предложена гипотеза общей энеолитической подосновы населения абашевской культуры и культур шнуровой керамики.
А по черепам из Пепкинского кургана антропологами была отмечена высокая изменчивость у абашевских мужчин.
Графические реконструкции мужчин погребенных в Пепкинском кургане (Реконструкции Г. В. Лебединской).
Но, в целом, метод главных компонент выявил сближение выборки мужчин, погребенных в Пепкинском кургане, с южноевропеоидными формами, ранее отнесенными к грацильным представителям средиземноморской расы. Что больше свидетельствует в пользу недавней миграции из Европы.
На графике с использованием метода понижения размерности (UMAP) люди из Волосово-Даниловского могильника сближаются со второй, более многочисленной группой из Пепкинского кургана, что может указывать на преемственность. Однако абашевец из погребения 7 Пепкинского кургана близок и к мужчине унетицкой культуры из Польши, а также к представителю позднего неолита из Марбьерга в Дании, которые жили примерно в то же время, что и он. А это уже аргумент в пользу недавней миграции.
У носителей фатьяновской культуры, на протяжении всего периода существования Волосово-Даниловского могильника, наблюдается достаточно стандартный погребальный обряд, который роднит их с культурами шнуровой керамики Европы, чего нельзя сказать для набора инвентаря поселений фатьяновцев. Но обряд свидетельствует о формировании могильника представителями пришлого населения, несущего традиции европейских групп шнуровой керамики.
А вот вопрос о происхождении представителей средневолжской абашевской культуры решается не столь однозначно. В ее составе присутствуют две составляющие — с гаплогруппами R1a (Z93>Z94) и R1b (Z2103).
Для носителей линии R1a (Z93>Z94) типично захоронение в могиле рядом близких родственников по материнской линии (причем здесь выделяются пары с европейскими «раннеземледельческими» митохондриальными гаплогруппами: как J2b1a2a из погребений 20, 21 и K1b1a1+199 из погребений 16,17).
В то время как захороненных рядом мужчин с гаплогруппой Y-хромосомы R1b (Z2103) сближает родство по отцовской линии. Это указывает на то, что система родства в группах разного происхождения отличалась.
Примечательно, что абашевцы из Пепкинского кургана с гаплогруппой R1a (Z93>Z94), в том числе, связанные тесным родством, всегда имели с собой костяные поделки и украшения, в отличие от носителей линии R1b (Z2103), у отдельных носителей которой прослеживаются связи со степными районами Южного Урала.
Однако решение вопроса о происхождении двух составляющих абашевского населения лежит в плоскости будущих исследований.
Источник: А. В. Энговатова, И. Э. Альборова, Х. Х. Мустафин, В. Ю. Луньков, Ю. В. Лунькова (Москва, Россия), А. А. Канапин, А. А. Самсонова (Санкт-Петербург, Россия), М. Б. Медникова (Москва, Россия) Древняя ДНК носителей фатьяновской и абашевской культур (к вопросу о миграциях населения эпохи бронзы в лесной полосе на Русской равнине) doi.org/10.55086/sp232207228
Сибирь занимает значительную часть Евразии, от Урала до Тихого океана. А самая восточная часть региона, благодаря Берингии, связывала Евразию с Северной Америкой. Начиная с верхнего палеолита, на этой обширной территории жили и смешивались люди с различным генетическим происхождением.
На сегодняшний день Саяно-Алтай представлен геномами людей со стоянок Афонтова гора верхнего палеолита, у современного Красноярска, возрастом около 17 тыс. лет. С предковой компонентой типичной для древних северных евразийцев (ANE) по аутосомному геному, которая в прошлом была широко распространена в тундростепи. Но после периода жизни представителей Афонтовой горы, не было информации о генетическом профиле людей, проживавших в пределах Алтайско-Саянской горной страны на протяжении почти 120 веков. Хотя в это время группы охотников-собирателей соседних регионов продемонстрировали различный предковый состав, что вызывает вопросы относительно их перемещений и связей. В частности, популяции Западной Сибири демонстрируют значительную долю аутосомной компоненты древних северных евразийцев (ANE), тогда как их современники из Восточной Сибири несли значительную долю компоненты древних северо-восточных азиатов (ANA), как у представителей неолита из пещеры Чёртовы Ворота в Приморье, возрастом около 6700 лет. Ранее, самое западное присутствие этой предковой компоненты было зарегистрировано в районе озера Байкал 7500 лет назад и в Центральной Монголии 5600 лет назад.
И третья, палеосибирская аутосомная предковая компонента, была определена у древнего жителя Усть-Кяхты (UKY) Забайкалья, возрастом около 14 тыс. лет и у обитателя Дуванного Яра в низовьях реки Колыма (Kolyma_M), возрастом 9800 лет. Эта компонента связывает древних жителей Сибири с коренными американцами. Однако подробности того, как между собой взаимодействовали люди с этими отличающимися предковыми линиями и как они связаны с последующими популяциями Северной Азии до конца не ясны, поэтому любые новый данные вызывают большой интерес.
В данной работе авторы получили полногеномные данные шести охотников-собирателей из четырех археологических памятников Саяно-Алтая, охватывающих период от 7,5 до 5,5 тыс. лет назад, а также охотника-собирателя из пещеры Летучая Мышь на Дальнем Востоке России, к северо-западу от Находки, возрастом 7 тыс. лет и трех менее древних обитателей Камчатки с некалиброванными датами, которые помещают их в интервал между XII и XVII веками нашей эры, точнее определить не позволяет эффект морского резервуара.
График анализа главных компонент с участием современных популяций Евразии и коренных американцев демонстрирует клинальный характер генетической изменчивости с запада на восток, где восточная часть клины, справа, представлена древними северо-восточными азиатами (ANA), как неолитические обитатели пещеры Чёртовы Ворота в Приморье, а западная часть клины, слева, представлена популяциями с высокой долей восточноевропейских охотников-собирателей (EHG), которые жили в тот же период, что и неолитические охотники-собиратели Саяно-Алтая.
Другой генофонд, который был в значительной степени распространен в северной Евразии со времен верхнего палеолита, связан с древними северными евразийцами (ANE), которые отклоняются вверх от вышеупомянутой клины на графике анализа главных компонент.
Пятеро древних алтайских охотников-собирателей из данной работы, представленных людьми из могильников Фирсово-XI (напротив Барнаула на правом берегу реки Обь, между селами Санниково и Фирсово), Новоалтайск-Развилка (в районе остановочного пункта Развилка города Новоалтайска) и Тузовские Бугры-I (в 30 км к югу от Барнаула), возрастом около 7500, 6500 и 5500 лет соответственно, в масштабах Евразии, расположились почти посередине между древними северо-восточными азиатами и древними северными евразийцами, а точнее между популяциями с высоким вкладом упомянутых компонент. Но в региональном масштабе алтайские охотники-собиратели попадают между представителями окуневской культуры и образцами Байкала, близко к человеку, обнаруженному в районе реки Базаиха на правом берегу Енисея, возрастом около 4700 лет. В работе 2020 года, над которой работали и некоторые авторы этой работы, этот человек хорошо подходил на роль представителя популяции, которая в последствии смешалась с группами, связанными с афанасьевской культурой и образовала генетический профиль носителей окуневской культуры.
При этом самые ранние свидетельства генетической родословной, связанной с кочевниками-скотоводами из западно-евразийских степей, появляются на Алтае начиная с 5000 лет назад с распространением афанасьевской культуры.
Однако давайте рассмотрим участки отбора образцов более детально:
Фирсово-XI
На этом участке обнаружено восемь погребений, которые первоначально были отнесены к большемысской культуре энеолита. Они включали пять одиночных могил, два парных и одно коллективное захоронение, глубиной от 0,4 до 1,7 м. Люди в них были похоронены в два ряда, непараллельно, лёжа на спине, а головами на северо-восток, за исключением одного мужчины 35–45 лет, который был расположен в сидячем положении. Сопроводительный инвентарь представлен в основном заготовками костяных орудий, наконечниками стрел треугольной формы, скребками, топорами и ножами. Украшения представлены ожерельями, сделанными из костяных подвесок и зубов животных с отверстиями. В работе этот участок представляют 2 человека из могил 9 и 14, похороненных около 7500 лет назад. Важно отметить, что могила № 14 представляет собой самое богатое захоронение данного некрополя. И хотя по антропологическим показателям скелет человека возрастом 18–20 лет был похож на женский, набор погребального инвентаря и генетический анализ свидетельствуют в пользу его принадлежности к мужскому полу. На скелете обнаружено девять ожерелий из 21 резца волка, 50 резцов лося и 19 моляров и премоляров медведя.
Новоалтайск-Развилка
Это место было обнаружено и раскопано в Новоалтайске на ул. Репнина в обрыве надпойменной террасы правого берега Оби. В процессе раскопок расчищен неполный скелет юноши 15–16 лет, погребенного около 6300 лет назад в вытянутом положении на спине, головой на север и руками вдоль туловища. Сопроводительный инвентарь включает каменные и костяные предметы: подвеску из клыка барсука с просверленным в корне отверстием, кость птицы, ребристый скол, отщеп, прямоугольные украшения из раковин крупных речных моллюсков, каменное шлифованное долото, позвонок животного, резцы бобра, а также орнаментированный игольник, изготовленный из лучевой кости птицы (размером с крупную утку). Трасологический анализ последнего показал, что орнамент нанесен металлическим ножом, который использовали в качестве резчика. При зачистке на уровне могильного пятна к востоку от скелета найден фрагмент большемысской керамики, орнаментированный отпечатками гладкой качалки, кстати, орнамент характерный для «афанасьевцев».
Тузовские Бугры-I
После обнаружения в 1995 году трёх могил, в 2000 году на памятнике Тузовские Бугры-I уже было исследовано 37 погребений. Из них 19 относятся к эпохе энеолита – ранней бронзы, остальные к раннему железному веку. Часть погребений раннего комплекса была нарушена более поздним. Энеолитические могилы располагались четырьмя рядами, глубиной 0,5–1,15 м, ориентированными по линии СЗ–ЮВ.
Большая часть захоронений одиночные, но также было три парных и два коллективных от 3 до 5 человек. Большинство захороненных лежали на спине в вытянутом положении, с руками вдоль туловища, а головой на северо-восток. Сопроводительный инвентарь представлен в основном украшениями одежды и головных уборов из раковин моллюсков и зубов животных, а также изделиями из камня и кости.
В одной из могил обнаружены металлические височные кольца и два сосуда баночной формы. Зубы животных встречены в девяти погребениях. В исследовании этот участок представляют 2 человека из погребений 8 и 33, возрастом около 5500 лет.
Антропология
Чтобы лучше представить людей, о которых идёт речь, дополним картину описанием антропологических типов неолитического населения из Фирсово XI, которое по данным исследования 2017 года Константина Солодовникова и Светланы Тур, было морфологически промежуточным между европеоидным и монголоидным, с условной долей монголоидного элемента суммарно у мужчин и женщин 57,3%. А по сочетанию признаков люди из Фирсово XI близки к неолитическим жителям предгорного Алтая.
Реконструкции внешности удобнее смотреть в видео, в контексте.
Кстати черепа женской части группы Фирсово XI, были схожи с черепом мужчины из карстовой пещеры Нижнетыткескенская I с умеренно выраженными монголоидными особенностями в сочетании с брахикранной формой мозговой коробки и широким, но невысоким (по монголоидному масштабу) лицом.
Генетические данные человека из пещеры Нижнетыткескенская-1, также представлены в этом исследовании. Подобный комплекс признаков отмечается также на неолитических черепах из могильников Базаиха и Долгое Озеро в Красноярско-Канской лесостепи.
Пещера Нижнетыткескенская-1
Пещера расположена в горном массиве на высоте 130 метров над уровнем реки Катунь у устья её левого притока реки Тыткескень. В пещере зафиксированы культурные слои от конца мезолита до средневековья, а также уникальное для Сибири захоронение большемысской культуры, возрастом около 6500 лет, с большим набором как хозяйственно-бытовых, так и ритуальных предметов. Ориентация скелета не может быть установлена в силу специфичности положения его частей. При этом все кости принадлежат одному человеку в возрасте 40-45 лет. Расчлененный скелет может быть связан с шаманской практикой, поскольку большинство предметов, которые описаны в дополнительных материалах, вероятно, составляют шаманский костюм сибирских племен 4 тысячелетия до н.э. Значение костюма может быть истолковано как символ превращения человека в животное, подобное птице.
На графике анализа главных компонент, этот человек смещён от вышеописанных охотников-собирателей Алтая в сторону древних северо-восточных азиатов близко к более древним жителям Америки.
В данном случае, по результатам статистики внешней группы f3, алтайские охотники-собиратели демонстрируют наибольшее генетическое сходство друг с другом и с представителем Базаихи (Yenisei_4700BP). Анализ также указывает на высокий уровень общих аллелей с людьми и популяциями, со значительным вкладом от древних северных евразийцев (ANE), такими как Мальта 1, Афонтова гора 3, неолитические охотники-собиратели Западной Сибири, представители ботайской культуры и жители Таримского бассейна бронзового века из могильников Сяохэ и Гумугоу, последние более известны как таримские мумии. Следовательно, авторы использовали в моделях эти популяции в качестве источника родословной от древних северных евразийцев (ANE). При этом ботайцы и неолитические охотники-собиратели Западной Сибири (West_Siberia_N), содержащие родословную восточноевропейских охотников-собирателей, лучше подходят для моделирования компоненты древних северных евразийцев (ANE) у алтайских охотников и собирателей вместе взятых.
А для источника древних северо-восточных азиатов (ANA) у алтайских охотников-собирателей, учитывая в расчётах некоторую близость к ним современных коренных американцев михе, авторы сосредоточились на древних геномах, которые связаны с палеосибирской родословной, присутствующей и в популяциях коренных американцев, как у представителя верхнего палеолита из Усть-Кяхты Забайкалья (UKY), возрастом около 14000 лет, обитателя стоянки Дуванный Яр — Колыма1, возрастом 9800 лет, у популяций Якутии из Центральной Сибири, возрастом около 6000 лет, а также у древних жителей Северной Америки, как Анзик-1 из современного штата Монтана возрастом 12700 лет и Кенневикский человек из штата Вашингтон, возрастом около 8-9 тыс. лет. В расчётах представители Дуванного Яра, Усть-Кяхты и древние популяции Якутии, лучше подходят для моделирования родословной древних северо-восточных азиатов (ANA) у групп алтайских охотников-собирателей, чем жители неолита из пещеры Чёртовы Ворота в Приморье.
Таким образом, авторы смоделировали представителей участков Фирсово-XI, Новоалтайск-Развилка, Тузовские Бугры-I и Базаихи, как смесь популяции с генетическим профилем как у древнего жителя Усть-Кяхты (UKY) Забайкалья от 53,7% до 66,3%, чтобы представить компоненты палеосибирского происхождения и неолитических охотников-собирателей Западной Сибири (West_Siberia_N) от 33,7% до 46,3%, как источника родословной древних северных евразийцев (ANE).
По оценкам это смешение произошло в интервале от 11000 до 7400 лет назад. Что, возможно, свидетельствует о множественных и/или непрерывных событиях смешения в процессе формирования генофонда алтайских охотников-собирателей.
При этом индивидуальная близость охотников и собирателей Алтая к древним американцам, лучше всего объясняется различными пропорциями палеосибирского происхождения, которое связано и с древними американцами.
В целом, анализ показывает, что древние алтайские охотники-собиратели имели сибирский генетический профиль, ранее известный только благодаря человеку, обнаруженному в районе реки Базаиха, возрастом около 4700 лет (Yenisei_4700BP), но который начал формироваться ещё в раннем голоцене в результате характерной смеси предковых линий палеосибиряков и древних северных евразийцев.
Генетический вклад алтайских охотников-собирателей в соседние и более поздние популяции
К востоку от Алтайского края байкальские охотники-собиратели раннего неолита (EN) и раннего бронзового века (EBA), помимо родословной древних северо-восточных азиатов (ANA) имели и часть родословной древних северных евразийцев (ANE). Следовательно, авторы исследовали, могла ли родословная описанных алтайских охотников-собирателей способствовать генетическому профилю байкальских охотников-собирателей, о которых сообщалось ранее.
В результате было обнаружено, что все три алтайские группы подходят для моделирования прибайкальских охотников-собирателей раннего неолита (EN) и раннего бронзового века (EBA) в моделях примесей с двумя источниками (qpAdm).
Для моделирования прибайкальских охотников-собирателей раннего неолита (EN) в дополнение к родословной древних северо-восточных азиатов (ANA), как у представителей неолита из пещеры Чёртовы Ворота в Приморье, требуется около 20% родословной охотников-собирателей Алтая. А для моделирования охотников-собирателей раннего бронзового века (EBA), в дополнение к родословной охотников-собирателей раннего неолита дополнительно требуется около 25% родословной алтайских охотников и собирателей. При этом даты примеси родословной охотников-собирателей Алтая для байкальского неолита и бронзового века близки и составляют 8296 ± 624 лет назад и 8202 ± 995 лет назад соответственно.
В целом алтайские охотники-собиратели представляют собой более подходящий источник родословной древних северных евразийцев, которая участвовала в формировании генетического профиля байкальских охотников-собирателей и обнаруживается в районе озера Байкал около 8 тыс. лет назад.
Помимо этого, новые данные проливают свет на генетическое происхождение более поздних популяций, как носители окуневской культуры бронзового века, возрастом около 4600–3900 лет, которая представляет собой археологический феномен, возникший в Минусинской котловине вслед за афанасьевской культурой.
В предыдущих исследованиях сообщалось о гаплогруппе Q Y-хромосомы у людей, связанных с окуневской культурой, что наблюдается и у алтайских охотников-собирателей. А по полногеномным данным, в работе 2020 года окуневцы были смоделированы как трёхсторонняя смесь компонентов ботайцев, Байкала бронзового века и степных скотоводов. Или как более простая смесь компонент как у представителя Базаихи и носителей афанасьевской культуры. Авторы уточняют генетический состав групп окуневской культуры как трёхсторонняя смесь, в основе которой алтайские охотники-собиратели, географически расположенные к югу от Минусинской котловины, а также представители афанасьевской культуры и в меньшей степени охотники-собиратели Байкала раннего неолита.
У представителей окуневской культуры примесь алтайских и байкальских охотников-собирателей произошла примерно 6000 ± 700 лет назад, а вклад представителей афанасьевской культуры произошел позже, за 16-18 поколений до её возникновения, что перекрывается с датировками присутствия в регионе носителей афанасьевской культуры, примерно около 5300–4500 лет назад.
Что подтверждает археологические данные, согласно которым начало окуневской культуры последовало сразу же за концом афанасьевской.
Также авторы проанализировали и другие популяции бронзового века, так представителей Алтая среднего/позднего бронзового века можно смоделировать как смесь охотников-собирателей Байкала и Алтая с представителями синташтинской культуры среднего/позднего бронзового века.
Похожая модель подходит и для представителей Хубсугул позднего бронзового века из современной Монголии, которые ранее описывались как смесь компонент западноевразийских степей и Байкала раннего бронзового века.
Алтайский край также представляет собой географический коридор, соединяющий Минусинскую котловину с Джунгарской и Таримской равнинами в Синьцзяне. В недавнем исследовании сообщалось об изолированном и условно автохтонном генофонде у обитателей Таримского бассейна раннего-среднего бронзового века, связанному с древними северными евразийцами 4100–3700 лет назад. А их соседи из Джунгарии раннего бронзового века как смесь этого местного генетического профиля с предковыми линиями охотников-собирателей Байкала раннего бронзового века и афанасьевцев. Однако в свете новых данных, представителей Джунгарии можно смоделировать как смесь местного генофонда Таримского бассейна раннего-среднего бронзового века участков Сяохэ и Гумугоу с описанными в данной работе алтайскими охотниками-собирателями.
Но эти модели требуют дополнительных анализов. И хотя в целом генофонд алтайских охотников-собирателей является подходящим источником предков для групп Внутренней Азии бронзового века, и подобная родословная сохранялась в значительных количествах во времени и пространстве, генетические данные указывают на небольшой эффективный размер популяции алтайских охотников-собирателей, но не до такой степени как у древних жителей Таримского бассейна.
Родословная древних северо-восточных азиатов от Дальнего Востока до Алтая
В отличие от представителей кластера алтайских охотников-собирателей, человек, обнаруженный в пещере Нижнетыткескенская-1, возрастом 6500 лет, смещён в сторону древних северо-восточных азиатов (ANA) на графике анализа главных компонент. Используя алтайских охотников-собирателей из Фирсово-XI, в качестве местного источника предков у представителя Нижнетыткескенской пещеры, можно смоделировать его как смесь алтайских охотников-собирателей с 47,7% ± 5,2% родословной представителей Байкала раннего неолита или с 37,4% ± 4,3% неолитических жителей Приморья из пещеры Чёртовы Ворота.
Событие примеси датируется за 52 ± 11 поколений до жизни захороненного в пещере Нижнетыткескенская-1 или примерно 8000 ± 300 лет назад. Теперь, благодаря новым данным, самое западное зарегистрированное присутствие предковой компоненты древних северо-восточных азиатов (ANA) сместилось более чем на 1000 км западнее Байкала, до Алтайского края по крайней мере 6500 лет назад.
В работе также авторы сообщают о генетических данных охотника-собирателя из пещеры Летучая Мышь на Дальнем Востоке России, возрастом около 7000 лет, который демонстрирует наибольшее генетическое сходство с ранее опубликованными популяциями древних северо-восточных азиатов (ANA) из пещеры Чёртовы Ворота в Приморье, возрастом около 6700 лет и носителями бойсманской культуры среднего неолита из археологического памятника Бойсмана-2, который располагается на восточном побережье залива Петра Великого в Приморье, возрастом 6300 лет.
Примечательно, что охотник-собиратель из пещеры Летучая Мышь демонстрирует дополнительное сходство с охотниками-собирателями, связанными с культурой дзёмон из Японии, возрастом 9000 лет, чей вклад в генофонд человека из пещеры Летучая Мышь достигает в моделях (qpAdm) 29,7% ± 9,8%. Похожая пропорция предков наблюдается у одного образца из бухты Бойсмана, который считается аутлаером по отношению к другим из археологического памятника Бойсмана-2.
Эти данные предполагают существование ранее недооцененных генетических связей между группами охотников-собирателей Японских островов и Дальнего Востока современной России уже 7000 лет назад.
Непрерывный или повторяющийся поток генов через Берингово море
Палеосибирская родословная, связывающая древних жителей Сибири с коренными американцами была широко распространена, от Алтая и Забайкалья через Якутию до Северного Ледовитого океана. Предыдущие исследования показали, то представитель стоянки Дуванный Яр — Колыма_М, возрастом 9800 лет и популяций Якутии из Центральной Сибири, возрастом около 6000 лет,имеют тесные генетические связи с современными группами Камчатки, говорящими на чукотско-камчатских языках и коренными американцами. Новые генетические данные трёх жителей камчатки, с некалиброванным возрастом в 500 лет указывают на их генетическое родство с современными популяциями Камчатки, такими как коряки и ительмены, а также с древним жителем Дуванного Яра и представителем культуры саккак из Гренландии, возрастом около 4 тыс. лет, представляющем палеоэскимосов. В связи с чем авторы проанализировали новые образцы Камчатки вместе с древними популяциями из Якутии, побережья Берингова моря и Гренландии чтобы исследовать популяционную динамику в Северо-Восточной Сибири и Берингии.
Генетические данные указывают на значительные демографические изменения у жителей Северной Сибири в интервале между 6000 и 2000 лет назад, что можно наблюдать на графике анализа главных компонент в виде клины между представителями Якутии разных периодов, благодаря которой генетический профиль древних жителей региона смещается в сторону современных северо-восточных сибирских популяций, таких как эвенки и эвены.
Повторный анализ ранее опубликованных геномов показывает, что генетические изменения в Якутии между 6000 и 4000 лет назад связаны с увеличением родословной древних северо-восточных азиатов (ANA) почти на 50%. В то время как изменения от 4000 до 2500 лет назад, возможно, связаны с примесью из Северо-Восточной Сибири, что хорошо заметно у современных нганасан. При этом демографическая история групп Северо-Восточной Сибири также включает в себя обратный поток генов, связанных с коренными американцами через Берингово море.
Используя группу Якутии позднего неолита в качестве исходной линии происхождения, поток генов, связанных с коренными американцами, составляет почти 50% в группы железного века из района самых восточных поселений России, как Эквен и Уэлен датируемые около 2000-2300 лет соответственно.
Таким образом, генетический профиль жителей Камчатки, возрастом около 500 лет, можно смоделировать как смесь предковых линий древних популяций Якутии и коренных американцев.
А в результате исследования генетического формирования современных популяций Камчатки в свете древних образцов, было обнаружено, что коряки несут дополнительную родословную, связанную с коренными американцами, в отличие от более древних жителей полуострова. Другие современные популяции Камчатки и Чукотки, такие как ительмены и чукчи, также имеют дополнительное количество родословной коренных американцев от 5% до 20%.
Причём этот обратный поток генов, по оценкам, произошел 5500-4400 лет назад у древних популяций и около 1500 лет назад у предков современных коряков. Что с учётом увеличенного вклада от коренных американцев в генофонде коряков по сравнению с более древними обитателями Камчатки, указывает на непрерывный или повторяющийся поток генов из Северной Америки или Берингии в популяции Камчатки в течение последних нескольких тысячелетий.
Итоги
Благодаря полученным данным, в частности по алтайским охотникам-собирателям, авторы работы улучшили модели происхождения некоторых популяций и выявили генетические связи между Алтайским краем, районом озера Байкал, а также с Джунгарским и Таримским бассейнами Внутренней Азии, начиная со среднего голоцена.
Сам же генетический профиль алтайских охотников-собирателей в значительной степени сохранился локально в генофонде скотоводов, связанных с окуневской культурой, несмотря на изменение стратегии ведения хозяйства. А на Алтае, родословная эпохи неолита была существенно сокращена к среднему и позднему бронзовому веку, возможно, в результате миграций в регион популяций как с востока, так и с запада. Данные также указывают на то, что формирование генетического кластера алтайских охотников-собирателей не связано с расширением родословной древних северо-восточных азиатов (ANA). Однако близкий по времени к ним человек, захороненный в пещереНижнетыткескенская-1, возрастом 6500 лет, демонстрирует значительное количество родословной древних северо-восточных азиатов (ANA), что, примерно, на 1500 км западнее, чем было зарегистрировано ранее. Такой отчетливый генетический профиль указывает на то, что предковые линии древних северо-восточных азиатов (ANA) прибыли в Алтайский регион до распространения афанасьевской культуры, что, возможно, связано с различными культурными особенностями, наблюдаемыми у человека из пещеры Нижнетыткескенская-1.
Присутствие этого «генетического аутлаера» может указывать на ранее недооцененные связи с Востоком на большие расстояния. Кстати похожая недооценка, по-видимому, была и в случае с древними жителями Дальнего Востока России. В частности, человек из пещеры Летучая Мышь и аутлаер из бухты Бойсмана, возрастом 7000 и 6300 лет соответственно, демонстрируют дополнительное сходство с охотниками-собирателями, связанными с культурой дзёмон из Японии, обнаруживая неожиданные генетические связи между группами охотников-собирателей Японских островов и Дальнего Востока современной России.
А в целом, новые генетические данные в сочетании с ранее опубликованными, охватывающими территорию от Центральной Азии до Северной Америки, выявили сложную динамику перемещения групп охотников-собирателей по Северной Азии, начиная с раннего голоцена.
Помимо этого, авторы обнаружили, что генофонд современных популяций Камчатки был сформирован в результате длительного периода потока генов, связанных с коренными американцами, на протяжении нескольких тысячелетий.
Ke Wang et al. Middle Holocene Siberian genomes reveal highly connected gene pools throughout North Asia DOI: 10.1016/j.cub.2022.11.062
В истории человечества распространение языков часто сопровождалось миграциями, однако люди могли переходить на другие языки практически без миграций и, соответственно, без генетических изменений. И наоборот, смешиваться без языковых изменений.
Таким образом, выводы о прошлых контактах населения, основанные либо на текущем распространении языков, либо только на компонентах генетического происхождения, могут ввести в заблуждение. Тем не менее исследования древней ДНК позволяют нам напрямую наблюдать за изменениями в генофондах популяций во времени и пространстве, выявляя связи между генетическими, археологическими и лингвистическими данными. Волго-Окское междуречье имеет интригующую историю притока населения и смены языков. В настоящее время большинство жителей региона говорят по-русски, но до средневековья северо-запад России населяли уралоязычные народы. А постепенный переход к славянским языкам начался во второй половине первого тысячелетия н. э., с экспансией славянских племен, в результате которой было основано государство Киевская Русь в конце IX века н. э. Средневековая Русь была многокультурной и многоязычной. Но после крещения Руси и распространения кириллической письменности в X-XI веках, влияние и привлекательность славянского языка усилилось, что привело к языковому сдвигу с уральского на славянский. При этом несмотря на замену языка, современные русские северо-западных регионов, демонстрируют явную близость к своим уралоязычным соседям, что предполагает генетический вклад от этих предшествующих популяций.
Суздальское Ополье является историческим ядром Северо-Восточной Руси, которое сыграло важную роль в политической истории Волго-Окского междуречья XI – XIII вв. Для него характерно непрерывное существование сельских поселений, возникших в X – XII вв., часто на территориях, заселенных уже в I тысячелетии н. э. До IX– X вв. этот регион был населен уралоязычными группами, а славянская колонизация началась в X в. Ранние летописи связывают мерян, вымершую уралоязычную группу, с первыми жителями региона. Средневековые города – Суздаль, Владимир, Юрьев-Польский и сельские поселения Суздальского Ополья тщательно изучены археологами. Их материальная культура представляет собой классическую древнерусскую традицию XI – XIII вв. А предшествующий период характеризуется сложной и многокомпонентной материальной культурой, сочетающей в себе элементы как славянских групп, так и финно-угров Поволжья. Таким образом, этнокультурную историю Суздальской земли в средние века можно рассматривать как один из этапов формирования и закрепления региональной группы средневековой Руси (именуемой в летописях как «суздальцы») на территории, ранее занимаемой финно-уграми Поволжья. Тем не менее подробности взаимодействия и слияния славянского и финно-угорского населения в значительной степени неизвестны. Некрополи X–XII вв. в пределах Суздальского Ополья известны в основном по раскопкам, проведенным в середине XIX в., и их документация не дает четкого представления о погребальных обрядах и датировке. А поселения второй половины первого тысячелетия, т. е. периода, предшествующего расселению славян, и вовсе практически не изучены, да и могильники того времени также не известны. В новой работе авторы при помощи анализа древней ДНК и методов радиоуглеродного датирования исследовали останки 32 древних жителей Волго-Окского междуречья из шести археологических памятников Суздальского Ополья на территории Владимирской и Ивановской областей и двух из Гороховца на восточной окраине Владимирской области.
В целом образцы охватывают период от 188 до 1787 года н. э. (плюс/минус) в том числе и времена демографических изменений, связанных с распространением славянских языков.
Кроме того, чтобы реконструировать изменения в рационе и образе жизни людей того времени, были получены данные о соотношении стабильных изотопов углерода и азота на участках Большое Давыдовское 2 и Шекшово 2 и 9. При этом авторы, пытаясь найти связь между лингвистическими и генетическими изменениями, подчёркивают, что по генетическим данным сложно судить о языке или этнической принадлежности. Как собственно и по материальной культуре. Поэтому к терминологии, используемой в исследовании стоит относиться адекватно и с пониманием.
Результаты
Генетический сдвиг в Волго-Окском междуречье при переходе от железного века к средневековью
На графике анализа главных компонент между популяциями Западной и Восточной части Евразии, по оси абсцисс, наблюдаются три генетические клины, которые примерно соответствуют экорегионам Центральной её части.
Самая северная клина проходит по лесотундре и большинство говорящих на уральском языке попадают на неё. В соответствии с их географией представители железного века из могильника Большое Давыдовское 2 попадают на эту клину, между русскими с побережья Белого моря в Архангельской области и современными популяциями Поволжья. Анализ примесей также показал, что люди из Большого Давыдовского имели долю древнего сибирского происхождения, как и большинство современных уралоязычных групп.
Эта предковая компонента максимально выражена у современных нганасан Таймыра, но, возможно, в прошлом, она была ещё более широко распространена. А вот образцы последующих за железным веком периодов из Шекшово, Гороховца, Кибола, Кидекши и Красногвардейского, расположены ближе к западноевропейским популяциям на графике анализа главных компонент, что указывает на генетические изменения в регионе после железного века. Однако около половины средневековых жителей оставались по-прежнему генетически близки к людям железного века из могильника Большое Давыдовское 2. А другая их часть продемонстрировала близость к современным славянам Восточной Европы. Эта закономерность предполагает, что как минимум к началу XI века, в Волго-Окском междуречье всё ещё продолжалось смешение или же две различающиеся популяции сосуществовали в одном и том же регионе. Однако уже после средневековья большинство жителей Ополья, которое ещё называют Владимирским или Юрьевским, уже было ближе к современным центральным и южным русским. И эти люди не демонстрируют таких же различий в происхождении, как их предшественники.
Изотопный анализ
Данные изотопного анализа костных фрагментов с участков Большое Давыдовское 2 и Шекшово 2 и 9 также указывают на перемены между железным веком и средневековьем. У людей железного века из могильника Большое Давыдовское 2 рацион был с высоким содержанием белка и растениями с C4-фотосинтезом (вероятно, просом), тогда как у людей из средневекового курганного могильника Шекшово 9, несмотря на генетические различия между захороненными, рацион был основан на растениях с C3-фотосинтезом, как пшеница и ячмень, к примеру.
Примечание (Кластеры):
Для последующих анализов образцы были распределены по четырем аналитическим кластерам на основе археологического контекста, радиоуглеродных дат и результатов анализа главных компонент и примесей.
7 человек вошли в кластер (VolgaOka_IA) – Волго-Окского междуречья железного века;
4 человека отнесены к кластеру (VolgaOka_MA1) – Волго-Окского междуречья, средневековья как схожие с популяциями железного века из того же региона;
6 человек вошли в кластер (VolgaOka_MA2) Волго-Окского междуречья, средневековья, но в отличие от предыдущего, как схожие со славянскими популяциями Восточной Европы;
4 человека представляют кластер (VolgaOka_H) Волго-Окского междуречья постсредневековья.
Некоторые образцы не были включены в кластеры из-за их близкого генетического родства с другими людьми, загрязнения, и из-за того, что они попали под критерии хронологических и генетических статистических выбросов О них в отдельном разделе).
Популяции Поволжья железного века и древняя сибирская родословная
Результаты статистики внешней группы f3 показали высокий уровень общих аллелей с литовцами для всех анализируемых кластеров. А в тестах кладальности f4 для каждого из вышеописанных кластеров, представители средневековья Волго-Окского междуречья, схожие со славянскими популяциями Восточной Европы были кладальны с восточными славянами (как белорусы с украинцами, к примеру) и современными русскими из Рязанской области, тогда как группа людей из кластера Волго-Окского междуречья постсредневековья была кладальной только с рязанскими русскими. А представители Волго-Окского междуречья железного века и схожие с ними жители средневековья не были кладальны ни с одной из тестируемых популяций, однако они продемонстрировали наименьшие отличия от современных русских из Архангельской области, а это позволяет предположить, что северные русские являются их ближайшими современными родственниками. Тесты на кладальность выявили общую генетическую границу между народами Восточной части Евразии и Волго-Окского междуречья железного века и схожими с ними жителями средневековья, примерно в районе Уральских гор.
При этом упомянутые группы Волго-Окского междуречья имеют значительно больше общих аллелей с популяциями к востоку от Урала, чем современные литовцы. Дальнейшие анализы продемонстрировали, что это родство с восточными популяциями связано древней сибирской родословной, хорошо выраженной у нганасан. Похожая, но более размытая граница наблюдается между народами Восточной части Евразии и Волго-Окского междуречья постсредневековья. Однако у средневековых групп из Волго-Окского междуречья, которые были схожи с современными славянскими популяциями Восточной Европы, не наблюдается большего количества общих аллелей с популяциями Восточной части Евразии чем у современных литовцев. Т.е. они не получали дополнительного количества сибирской родословной. Дальнейшие анализы показали, что при использовании в качестве аналога сибирской родословной, представителя бронзового века из Красноярского края, общие аллели с ним, представителями Волго-Окского междуречья железного века и схожими с ними жителями средневековья, также разделяют, в схожих диапазонах, северные русские из Архангельска и Вологды, а также вепсы, карелы, финны и мордва.
Волго-Окское междуречье близко к ареалу обитания восточноевропейских охотников-собирателей мезолита (EEHG), которые внесли свой вклад в происхождение многих более поздних популяций.
Результаты указали на увеличенный вклад восточноевропейских охотников-собирателей в обитателей Волго-Окского междуречья железного века, а также в представителей бронзового века из Эстонии и Оленеостровского могильника Кольского залива залива, хотя этот вклад был статистически значимым только для образцов из Оленеостровского могильника.
Популяции Центральной России сформировались как продукт местной примеси
Для моделирования состава предков (qpAdm), описанных выше кластеров, авторы построили согласованную модель с пятью источниками, которые включают представителя Красноярского края бронзового века, западноевропейских и восточноевропейских охотников-собирателей, носителей ямной культуры Самары и представителей культуры линейно-ленточной керамики раннего неолита.
При этом восточноевропейские охотники-собиратели мезолита были наиболее актуальны для моделирования обитателей Волго-Окского междуречья железного века, схожих с ними жителей средневековья и современных удмуртов. А для большинства других популяций лучше подходила модель, включающая 4 источника без восточноевропейских охотников-собирателей. Чтобы точно определить временные или географические популяции, которые могли внести свой вклад в население Волго-Окского междуречья железного века, авторы протестировали несколько потенциальных моделей, из которых наиболее подходящие показали, что эти люди примерно наполовину имеют общее происхождение с населением, относящимся к представителям Прибалтики железного века (800 г. до н.э. – 50 г. н. э.), ещё 25% их предков были связаны с людьми бронзового века из Оленеостровского могильника Кольского залива и оставшиеся 25% их родословной связаны со степью железного века, в качестве носителей которой использованы скифы Восточной Европы.
Таким образом, Волго-Окское междуречье, по-видимому, находилось на пересечении потоков генов с нескольких направлений. Примечательно, что представители фатьяновской культуры не подошли в качестве достоверного источника происхождения для представителей железного века Волго-Окского междуречья ни в одной из протестированных моделей, что указывает на отсутствие генетического вклада этих групп, населявших этот же регион в бронзовом веке. А сами представители Волго-Окского междуречья железного века подходят для моделей смешения между ранними славянами и уралоязычными группами, где в качестве славян авторы использовали представителей кластера Волго-Окского междуречья, средневековья, схожих со славянскими популяциями Восточной Европы как белорусы, к примеру. При этом вклад от людей железного века в индивидуальных моделях людей из кластера средневековья схожих с популяциями железного века варьируется от 35 до 75%. А для моделирования жителей Волго-Окского междуречья постсредневековья, помимо вклада от средневековых групп схожих со славянскими популяциями, дополнительно потребовалась родословная от более ранних групп региона, как представители железного века, хотя в качестве этого источника могут выступать и смешанные жители средневековья схожие с популяциями железного века. Датировка появления сибирской родословной у проанализированных людей показала, что эта примесь у предков представителей железного века была в эпоху бронзы, а у предков схожих с ними жителей средневековья, в железном веке.
А вот у предков представителей региона средневековья, схожих со славянскими популяциями Восточной Европы и последующих жителей постсредневековья, появление сибирской родословной датировано VIII-IX веками, что хорошо соответствует предполагаемому началу расселения славян в Волго-Окском междуречье.
Уникальная родословная отдельных представителей средневекового Суздальского Ополья
В наборе данных, помимо сгруппированных в кластеры, было отмечено несколько образцов, отличающихся от других, которые в статистике называют – выбросы. Они хорошо заметны на графике анализа главных компонент. Одна женщина 30–39 лет из могильника Большое Давыдовское 2 (BOL006), генетически была более близка к последующим популяциям средневековья, чем к своим современникам из железного века. А вот женщина из Шекшово 9 (SHE008) была ближе к жителям Центральной и Западной Европы, чем другие представители Шекшово 9. Однако максимальные отличия продемонстрировали двое мужчин 25-35 лет из могильника Шекшово 2 (SHK001 и SHK002), Ивановской области на рубеже XII-XIII веков, с линиями гаплогрупп J2a и N1. Которые были захоронены в неглубоких ямах овальной и прямоугольной формы, лежа на спине головой на Запад/юго-запад, с похожим положением рук: левые на животе, а правые около талии. Без погребального инвентаря, только у одного в районе ключицы обнаружены две бронзовые пуговицы. На графике анализа главных компонент они были ближе к популяциям Восточной Азии и попадают на «лесостепную клину». Их ближайшие соседи на графике - казахи, каракалпаки, сибирские татары и другие тюркоязычные группы из Центральной Азии и Сибири. Оба человека также несли митохондриальные гаплогруппы M9a1a и Z3c, которые более распространены в Азии, чем в Европе. В тестах кладальности f4 они продемонстрировали сходство с каракалпаками. Анализ изотопного состава стронция из исследования 2020 года указал на то, что росли они в другом месте, а в Поволжье прибыли будучи уже взрослыми.
Как считают авторы работы, эти двое мужчин могли представлять тюркоязычные группы, представители которых находились на военной службе в Киевской Руси, в основном охраняя ее южные рубежи. Однако в вышеупомянутом исследовании 2020 года, на которое ссылаются авторы данной работы, соотношения изотопов стронция у этих мужчин были близки к значениям, полученным для девушки 15-19 лет конца X – начала XI века из погребения №14, могильника Шекшово 9, указывая на то, что военная служба - это не единственная интерпретация из возможных, хотя и не отрицая её. Также соотношение изотопов стронция этих мужчин и девушки были близки к таковым у мужчины XII века, умершего в возрасте 30–39 лет, который провел свое детство в Суздальском Ополье, но прожил последние годы своей жизни в других землях, по изотопному составу схожих с таковыми, где росли девушка и упомянутые мужчины, и был похоронен на своей родине в Шекшово. При этом схожий изотопный состав не обязательно может быть только в одном месте. В любом случае, данные указывают на то, что в те времена были регионы, имевшие устойчивые связи с Суздальским Опольем. Помимо этого, интересный предковый состав продемонстрировала женщина из Кидекши (KED004), датированная 1414 г. н. э. и с митохондриальной гаплогруппой F2e, которая преимущественно встречается у современных жителей Восточной и материковой части Юго-Восточной Азии. На графике анализа главных компонент она попадает на клину между Европой и Ираном с Кавказом. А в анализе примесей она продемонстрировала генетическое родство с древними и современными иранцами. При этом наилучшая модель включала средневековых аланов Северного Кавказа в дополнение к родословной популяций славянского и дославянского периодов. Эти аномальные образцы подчеркивают наличие взаимных контактов у населения Волго-Окского междуречья с другими регионами и в целом значимость Суздальского Ополья в эпоху средневековья.
Параллельное распространение генов и языков в Волго-Окском междуречье
Последние полторы тысячи лет истории Волго-Окского междуречья характеризуются постепенным переходом с уральских языков на славянские. Этот процесс также оставил свой отпечаток и в генофонде. Наличие сибирской родословной у жителей Волго-Окского междуречья железного века, объединяет их с большинством современных уралоязычных популяций. В рамках моделирования местная группа железного века была подходящим источником сибирского происхождения для большинства представителей средневековья. Однако учитывая, что культура жителей железного века в пределах участка Большое Давыдовское исчезла уже к VII в, маловероятно, что именно они были непосредственными предками средневековых групп. Близкородственной, но отдельной уралоязычной группой Волго-Окского междуречья, во времена славянских миграций, могли быть представители летописного племени меря, с реконструированным ареалом включающем и Суздальское Ополье. Славянские миграции во второй половине первого тысячелетия нашей эры сформировали языковой ландшафт Северо-Западной Руси. А в северо-восточных регионах Киевской Руси, включающих также Суздальское Ополье, в X-XII веках славянские и уралоязычные народы часто формировали многоязычные сообщества, что может быть отражено и в данных из этого исследования, которые фиксируют появление славянской родословной и сосуществование в средневековье, предположительно, славянских и уральских групп. В Шекшово 9 при равном соотношении людей из обеих генетических групп, не наблюдается видимых различий между их погребениями. Более того, некоторые люди с уральским происхождением были похоронены с элементами “славянского” погребального инвентаря или смесью славянских и “уральских” предметов, что указывает на культурную интеграцию групп. Хотя моделирование и предполагает, что славянская группа внесла значительную долю родословной в генофонд более позднего населения, около 70%. Также несмотря на то, что исторические источники указывают на значительное скандинавское влияние на раннюю Русь, в пределах исследованных участков не обнаружено какого-либо генетического подтверждения этому. Большинство людей, помимо описанных исключений, имели компоненты как славянских, так и уралоязычных популяций в разных пропорциях. Однако люди скандинавского происхождения могли быть захоронены на других участках.
Выводы
Временной охват образцов древней ДНК из этого исследования демонстрирует, что Волго-Окское междуречье располагалось на перекрестке популяционных взаимодействий в течение последних двух тысячелетий. Местный генофонд железного века состоял из трёх основных компонент предков, напоминающих население Прибалтики и степи железного века, а также обитателей побережья Баренцева моря бронзового века. Интересно, что эти источники также связывали жителей Волго-Окского междуречья железного века, с близкими им по ареалу обитания, восточноевропейскими охотниками-собирателями мезолита. Тем не менее, их генофонд, по-видимому, в значительной степени отличался от носителей фатьяновской культуры бронзового века.
А раннее средневековье, в свою очередь, ознаменовалось изменением рациона питания и появлением генетического профиля схожего со славянским, что отражено в письменных источниках и соответствует выводам исторической лингвистики. При этом генетическое разнообразие населения средневекового Суздальского Ополья дополняли люди демонстрирующие генетическую близость к популяциям Центральной Азии, Кавказа и Ирана, подчёркивая взаимные контакты на большие расстояния И хотя результаты работы могут отражать локальную динамику, они показывают, что древняя ДНК может также предоставить дополнительные доказательства к истории языков, когда лингвистические данные скудны.
Источник: Genetic admixture and language shift in the medieval Volga-Oka interfluve Sanni Peltola, Kerttu Majander, Nikolaj Makarov et al. https://doi.org/10.1016/j.cub.2022.11.036
Процесс неолитизации в Европе привел к серьёзным технологическим и культурным сдвигам, которые включали в себя новые методы ведения хозяйства. Существуют две основные группы моделей, которые объясняют процесс неолитизации. Модели демической диффузии описывают неолитизацию как процесс колонизации земледельцами, который был движим экспоненциальным ростом населения, характерным для неолита. Вторая группа моделей – аккультурации – описывает процесс, в котором, по крайней мере, переход к земледелию мог практиковаться местными охотниками-собирателями после периода (той или иной продолжительности) взаимодействия с соседними, пришлыми земледельцами.
Анатолийские земледельцы достигли Балкан и других регионов Юго-Восточной Европы в седьмом тысячелетии до нашей эры и впоследствии распространились дальше через Средиземное море, а затем через Дунайский коридор, существенно вытеснив коренное мезолитическое европейское население.
В отличие от Центральной Европы, области Восточной Европы, включая Украину, Молдову, Западную Россию и Румынию, не занимались сельским хозяйством до позднего неолита (~ 4500 г. до н. э.), хотя различные оседлые и полуоседлые группы охотников-собирателей этих регионов начали использовать глиняную посуду еще в 8500 г. до н. э.
Археологическая общность Триполье-Кукутень представляет собой группу нескольких взаимосвязанных археологических культур среднего неолита/энеолита на территории ряда областей современных Украины, Молдовы и Румынии. Этот комплекс простирается от Южных Карпат до Днепра и назван в честь типовых памятников Кукутень в Яссы (Румыния) и Триполье (Обуховский район Киевской области).
Культуры Кукутень и Триполье имеют общие корни в культуре Прекукутень; самые ранние стоянки археологической общности Триполье-Кукутень обнаружены в предгорьях Карпат, а самые ранние радиоуглеродные даты (от периода Прекукутень II) относятся примерно к 4800 г. до н.э. Археологическая общность Триполье-Кукутень возникла в результате взаимодействия нескольких дунайских неолитических групп, о чем свидетельствует сходство в строительстве домов, керамических стилях и производстве каменных артефактов.
Rassamakin, Y., & Menotti, F. (2011). Chronological Development of the Tripolye Culture Giant-Settlement of Talianki (Ukraine): 14C Dating vs. Pottery Typology. Radiocarbon, 53(4), 645-657. doi:10.1017/S0033822200039102
После возникновения этого культурного комплекса в Карпатском предгорье археологическая общность Триполье-Кукутень расширяла свой ареал и в конечном итоге заняла территорию, охватывающую большую часть современных территорий Украины, Молдовы и Румынии. Первые раннетрипольские (Триполье А) стоянки отошли от культуры Прекукутень ~ 4500 г. до н. э. в долине реки Днестр.
Более поздние перемещения населения, происходящие со среднего периода (Триполье BI) и далее, привели к тому, что трипольская культура распространилась на Волынь на западе и Днепр на востоке.
Считается, что эта территориальная экспансия была вызвана в первую очередь демографическим ростом, связанным с успешной стратегией земледелия и животноводства, с поиском новых пахотных земель для возделывания. Однако некоторый рост населения мог быть результатом включения в состав трипольцев местных групп охотников-собирателей, например, представителей буго-днестровской культуры.
Другим каналом увеличения населения могла быть аккультурация переселенцев после краха неолита в Румынии, Венгрии и Болгарии. В средние и поздние периоды трипольской культуры (Триполье от BII до CI; 4100–3400 гг. до н. э.) некоторые группы трипольской культуры основали чрезвычайно крупные поселения в Центральной Украине, которые достигли размеров 100–320 га.
Быстрый демографический рост трипольцев на рубеже 4 тысячелетия до н. э. потребовал освоения новых территорий, ускорив миграцию в ранее периферийные районы.
Гипотезы возникновения мегапоселений разнообразны. Они могли быть защитным ответом на угрозы, исходящие от степных скотоводов или же от конкурирующих популяций внутри самой археологической общности Триполье-Кукутень. Они могли и просто быть временным эпизодом компактного скопления поселений из-за крупномасштабной миграции из Приднестровья.
Однако, несмотря на высокую плотность поселений в Западной и Центральной Украине, здесь обнаружено крайне мало захоронений. Лишь несколько кладбищ позднетрипольского периода были раскопаны в 1960-х и 1970-х годах, например, Чапаевка на территории Украины и Выхватинцы в Приднестровье. Хотя эти памятники дают некоторое представление о погребальных обрядах трипольцев, они ограничены по временному охвату и не подвергались современным лабораторным исследованиям.
Пример по могильнику в Выхватинцах: Положение скелетов в могилах различно — наряду с преобладающими погребениями в скорченном положении на левом или на правом боку отмечено также несколько случаев погребений на спине, с согнутыми и поднятыми в коленях ногами. Ориентировка довольно устойчива — наибольшее число погребенных положено головой на северо-восток, с небольшим отклонением к востоку. В погребальном инвентаре численно преобладает керамика, представленная двумя типами: сосудами из розовой глиняной массы, украшенными черной и красной росписью; и нерасписными сосудами слабого обжига, орнаментированными веревочными оттисками. Формы и стиль росписи керамики — типичные для позднего Триполья. Число сосудов в погребениях различно — от 1 до 8, чаще — 2-5. Помимо керамики, в мужских погребениях обнаружены роговые мотыги, кремневые вкладыши для серпов, костяные шила, а также каменный топор, кремневый нож, медное шило, глиняная и костяная антропоморфные статуэтки. Женские погребения сопровождались в основном сосудами, лишь в одном были найдены два пряслица и раковина. Выделяется своеобразием инвентарь детских погребений. Кроме сосудов, здесь были найдены антропоморфные глиняные погремушки; антропоморфные женские статуэтки были положены также главным образом в детские могилы. Помимо этого, обнаружены пряслица, бусы из раковин, кремневые отщепы, пластины, костяное шило. М. С. Великанова «Палеоантропология Прутскоднестровского междуречья» (Издательство «Наука», 1975 г)
Чтобы лучше понять происхождение, связи и разнообразие населения археологической общности Триполье-Кукутень, авторы исследовали палеоантропологические образцы из трех камер на участке пещеры Вертеба в Тернопольской области Украины - одном из немногих мест, где они обнаружены.
Результаты исследования методами масс-спектрометрии углерода-14 из останков людей и фауны указывают на присутствие трипольцев в пещере Вертеба в интервале между 3950 и 3520 гг. до н. э.
Lillie, M., Budd, C. E., Potekhina, I., Price, T. D., Sokhatsky, M., & Nikitin, A. G. (2017). First isotope analysis and new radiocarbon dating of Trypillia (Tripolye) farmers from Verteba Cave, Biche Zolote, Ukraine. Documenta Praehistorica, 44, 306–324. doi.org/10.4312p.44.18
По данным керамических комплексов, присутствующих в пещере, и результатам жидкостно-сцинтилляционного метода анализа углерода-14 с более низким разрешением можно предположить, что заселение продолжалось в течение некоторого времени в период позднего Триполья (CII) и перехода к раннему бронзовому веку.
Помимо этого, масс-спектрометрия также выявила отложения в разных местах пещеры, относящиеся к мезолиту (7950–7490 гг. до н. э.), бронзовому веку, железному веку и средневековью.
Останки людей были извлечены из трех отдельных камер пещеры: участок 7, участок 17 и участок 20. Каждая из этих камер содержит материальную культуру, соответствующую археологической общности Триполье-Кукутень.
Однако захоронения в пещере носят вторичный характер, а нарушения, вызванные деятельностью человека в древности и процессом перемешивания останков животными или растениями, усложняют реконструкцию использования пещеры и хронологии. Большинство изученных образцов в этом исследовании происходят из участка 7, который был подробно задокументирован с помощью анализа керамики и радиоуглеродного датирования, причем пик использования этого участка древним населением датируется периодами CI и CII трипольской периодизации (~ 3900-3350 гг. до н. э.).
Интерпретации использования пещеры разнообразны: в качестве временного убежища, места проведения ритуалов или же погребения. Имеются дополнительные сведения, подтверждающие гипотезу о жертвах конфликтов или же человеческих жертвоприношений.
Палеогенетика трипольской популяции до сих пор включила анализ однородительских маркеров мтДНК и полногеномный анализ 8 представителей трипольской культуры.
Митохондриальные гаплогруппы, типичные для древних евразийских земледельческих групп (H, HV, T, K, J), выявлены у образцов из всех трех камер пещеры. В одной из камер обнаружена гаплогруппа W, характерная для степных популяций, связанных с культурами шнуровой керамики и унетицкой культуры Среднего Поволжья.
Полногеномный анализ представителей археологической общности Триполье-Кукутень показал, что предковые генетические компоненты преимущественно (60–80%) относятся к ранненеолитическим земледельческим группам, подтверждая, что ранние земледельцы, заселившие Западную и Центральную Украину, в значительной степени происходили из той же исходной популяции, что и земледельцы Анатолии и Западной Европы. Оставшиеся 40-20% предковых компонент определены с меньшей достоверностью.
Immel, A., Țerna, S., Simalcsik, A. et al. Gene-flow from steppe individuals into Cucuteni-Trypillia associated populations indicates long-standing contacts and gradual admixture. Sci Rep 10, 4253 (2020). doi.org/10.1038/s41598-020-61190-0
Авторы работы 2018 г. [Mathieson et al.] показали, что этот наследственный генетический компонент присутствовал у западных и восточных охотников-собирателей, населявших этот регион в эпоху неолита.
Mathieson, I., Alpaslan-Roodenberg, S., Posth, C. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018). doi.org/10.1038/nature25778
Полногеномные данные четырех человек, происходящих из двух разных памятников на севере Молдовы (поздний трипольский период, 3500–3100 гг. до н. э., то есть на пять столетий моложе пещеры Вертеба), выявили у них значительный вклад степного населения (хотя и в разных пропорциях). Это может быть объяснено постепенной ассимиляцией местных мезолитических и неолитических групп охотников и собирателей трипольским населением, жившем на территории современной Молдовы и Приднестровья.
Ареал трипольцев расположен таким образом, что они контактировали как с центральноевропейскими, так и со степными популяциями. Археологические свидетельства взаимодействия с населением степей обнаруживаются в обожженной глиняной посуде. Некоторые из них выглядят почти идентично керамике, найденной в степях, в то время как другая керамика сочетает обжиг с декоративными мотивами трипольцев. Обнаружены и символические предметы, находящиеся под влиянием или непосредственно импортированные из степных сообществ. Например, каменные булавы обнаружены на некоторых стоянках среднего и позднего Триполья, а обмен глиняной посудой очевиден уже в Триполье BII.
Несомненно, существовала определенная степень взаимодействия между трипольским населением и днепро-донецкой культурой, в то время как взаимодействие трипольской культуры и последующей ямной, вероятно, было очень кратким.
Тем не менее, некоторые трипольские популяции, вероятно, находились в постоянном контакте со степными популяциями. После 3400 г. до н. э. трипольские мегаполисы были в значительной степени заброшены. Причина этого широко обсуждалась. Одна из гипотез - усиление конфликтов из-за расширения степных культур на запад. Такая гипотеза может найти подтверждение в частых свидетельствах насильственной смерти в пещере Вертеба.
Примеры травм у захороненных в пещере Вертеба (Violence at Verteba Cave, Ukraine: New insights into the Late Neolithic intergroup conflict G.D. Madden, J.K. Karsten, S.H. Ledogar, R. Schmidt, M.P. Sokhatsky doi.org/10.1002/oa.2633)
В данном исследовании авторы реконструировали последовательности генома 20 человек (8 женщин и 12 мужчин), захороненных в пещере Вертеба. Восемь образцов датированы масс-спектрометрией по углероду-14 в интервале от 3790 до 825 гг. до н. э, что соответствует трипольской культуре и позднее. Шесть человек (образцы ВЕРТ-035, ВЕРТ-106, ВЕРТ-031, ВЕРТ-100, ВЕРТ-104 и ВЕРТ-015) датируются поздним энеолитом (3790–3535 гг. до н.э.), один (VERT-113) из Зоны 7 датируется средним бронзовым веком (1960–1770 гг. до н.э.) и один из Зоны 17 (VERT-114) датируется поздним бронзовым веком (980–825 гг. до н.э.).
Используя полученные данные, авторы задались целью ответить на ряд вопросов:
1. Есть ли доказательства метисации с местными охотниками и собирателями, как было предположено в предыдущих работах?
2. Используя расширенный набор данных, можно ли уточнить неолитическую предковую составляющую трипольцев: сходны они с ранними земледельцами из Анатолии, представителями культуры линейно-ленточной керамики или иными?
3. Поскольку трипольцы жили в непосредственной близости от степных популяций, есть ли доказательства генетической близости к ямной или более ранним степным популяциям?
4. Имеет ли население более позднего бронзового века, заселившее регион, генетическую преемственность с представителями археологической общности Триполье-Кукутень из пещеры Вертеба?
Полученные результаты
Однородительские маркеры (мтДНК и Y-хромосомы)
Проанализированные образцы эпохи энеолита относятся к гаплогруппам митохондриальной ДНК T2b, H, HV, K1, N1, J1, U5 и T2c.
У представительницы среднего бронзового века (VERT-113) определена гаплогруппа HV, типичная для таких неолитических культур, как культура линейно-ленточной керамики к востоку от Дуная – Альфёльда, а также для жителей европейского бронзового века. Представительница позднего бронзового века (VERT-114) демонстрирует гаплогруппу T2, также связанную с многими культурами бронзового века. В целом эти гаплогруппы обычно встречаются в европейских популяциях эпохи неолита и бронзового века.
У мужчин выявлены гаплогруппы Y-хромосомы G2a, C1a и I2 (более подробно в таблице), которые также были выявлены в популяциях неолита и бронзового века Европы. При этом как гаплогруппы мтДНК, так и гаплогруппы Y-хромосомы всех образцов из Вертебы полностью согласуются с ранее опубликованными данными.
Полногеномные данные
На график анализа главных компонент вместе с 20 геномами из пещеры Вертеба авторы поместили 729 представителей современных популяций из Европы, а также 478 ранее опубликованных древних генома.
18 из 20 человек из Вертебы тесно связаны с европейскими популяциями эпохи неолита и энеолита: культура линейно-ленточной керамики, средний и поздний неолит Центральной Европы, а также трипольцы из Молдовы.
Анализ главных компонент также показал значительное сходство между 18 новыми образцами из Вертебы и четырьмя другими трипольцами из более ранней работы, также обнаруженных в пещере Вертеба. Поэтому их объединили в 22 образца, пометили на графике как Verteba_Trypillia и в дальнейшем проанализировали совместно.
А вот два генома бронзового века являются явными исключениями. Образец VERT-114 (позднего бронзового века) подпадает под разнообразие представителей культуры колоколовидных кубков и, по-видимому, занимает положение, близкое к чешской, венгерской и польской группам этой культуры. А образец, обозначенный как VERT-113 (среднего бронзового века), близок к европейским популяциям культур шнуровой керамики и срубной, демонстрируя значительное сходство со степным населением. Сразу стоит отметить, что статистический анализ указал на отсутствие доказательств связи со срубной культурой.
При этом все 22 трипольца из пещеры Вертеба содержат наследственный компонент, доминирующий у представителей анатолийского неолита (по методу ADMIXTURE).
Это указывает на явную связь с европейскими неолитическими популяциями, как собственно и было показано в предыдущих исследованиях. Однако эти же образцы также демонстрируют присутствие компонентов восточных, кавказских и западных охотников-собирателей. Лишь один образец (I3151), по-видимому, не имеет предков восточных/кавказских охотников-собирателей.
В геноме образца эпохи поздней бронзы (VERT-114, укладывающимся в разнообразие представителей культуры колоколовидных кубков) преобладает компонент анатолийского неолита, но выявлен значительный компонент восточных охотников и собирателей. А геном среднего бронзового века (VERT-113) демонстрирует высокую степень сходства с геномами культуры шнуровой керамики и ямной культуры. При этом хотя моделирование геномов трипольцев свидетельствует об их связи со степными популяциями, статистически (f4) они не связаны с популяциями ямной культуры из ареалов современных России и Украины.
Моделируя величину вклада различных основных предков, которые могли бы внести свой вклад в генетический портрет трипольцев, показано, что 40% вклада связано с анатолийским неолитом, 20% - с западными охотниками-собирателями и 40% - с кавказскими охотниками-собирателями.
При этом охотники-собиратели юго-восточной Венгрии – Кёрёш (Koros_HG), являются источником западных охотников-собирателей у трипольцев из Вертебы. Также неолит Украины и западные охотники-собиратели являются двумя вероятными источниками генетического пласта, связанного с охотниками и собирателями, для трипольцев из Вертебы, в дополнение к смоделированному компоненту Венгрия-поздний энеолит-ранний бронзовый век-ямная (Hungary_LateC_EBA_Baden_Yamnaya).
Это также указывает на очень незначительное присутствие среди предков трипольцев местных охотников-собирателей.
Кстати обратите внимание как происходит замена популяций на одной и той же территории и как сильно современные популяции отличаются от древних, вопреки распространяемым мифам.
Также стоит отметить, что большинство трипольцев из пещеры Вертеба не имеют статистически значимого сходства с молдавской популяцией археологической общности Триполье-Кукутень, но этих различий недостаточно, чтобы считать их разными популяциями.
Образец среднего бронзового века (VERT-113) является единственным, который демонстрирует четкий сигнал степного происхождения, причём наблюдается большая близость к ямной культуре с территории России, а не Украины
Кроме того, это единственный образец, который демонстрирует большую близость к представителям неолита на территории Украины, а не к западным охотникам-собирателям. В его геноме фиксируется до 33% вклад неолита Украины и 66% кавказских охотников-собирателей, что подтверждает большую долю степного происхождения.
Образец позднего бронзового века (VERT-114) генетически близок к геномам колоколовидных кубков на графике главных компонент и при моделировании «примесей» (ADMIXTURE). Однако, хотя модель с популяцией колоколовидных кубков в качестве единственного источника работает, есть и модели двух источников включающие 60% представителей культуры шаровидных амфор с территории Украины и 40% степных популяций.
Авторы также исследовали возможную связь генома поздней бронзы их Вертебы и киммерийцами, которые обосновались на территории современной Украины около 1000 г. до н. э. Однако геном из Вертебы оказался значительно ближе к носителям культуры колоколовидных кубков, чем к киммерийцам.
Анализ близкого родства в изученных геномах
Размеры гомозиготных участков ДНК указывают, что обитатели Вертебы жили в довольно многочисленных популяциях.
Исключением стал образец VERT-100, предки которого, вероятно, были родственниками.
Фенотипические и биохимические особенности
Ни один из проанализированных людей пещеры Вертеба не обладал толерантностью к лактозе.
Лишь два человека, видимо, были темноглазыми, но остальные обладали ДНК-маркером rs12913832, связанным с голубым цветом глаз.
Обсуждение
Генетический анализ 20 человек из пещеры Вертеба, которые датируются четвертым, вторым и первым тысячелетиями до нашей эры, выявил важные генетические изменения как в раннем бронзовом веке, так и в позднем. Эти датировки подтверждены радиоуглеродным датированием образцов 8 человек, поскольку пещера Вертеба многократно использовалась человеком, начиная с мезолита.
Предыдущие анализы митохондриальной ДНК представителей археологической общности Триполье-Кукутень показали их близкое родство по материнской линии с популяциями раннего неолита: гаплогруппы H, HV, T, V, J и K. И в новом анализе, за исключением двух человек с гаплогруппой U5a, все остальные 18 человек, имеют гаплогруппы, сходные с неолитическим населением Центральной Европы. Это разнообразие мтДНК резко контрастирует с населением более ранних неземледельческих неолитических памятников на территории Украины, которые имеют только гаплогруппу U, указывающую на преемственность с мезолитическими охотниками-собирателями.
Mathieson, I., Alpaslan-Roodenberg, S., Posth, C. et al. The genomic history of southeastern Europe. Nature 555, 197–203 (2018). doi.org/10.1038/nature25778
Разнообразие гаплогрупп мтДНК предполагает, что местное население в значительной степени было заменено населением, связанным с трипольской культурой. Большинство представителей Вертебы имеют Y-гаплогруппу G2a2, которая широко распространена среди европейцев эпохи неолита, связанных с Анатолией.
Другие выявленные гаплогруппы - C1 и I2 - также обнаружены среди европейских неолитических популяций.
Это указывает на миграцию и мужчин, и женщин, что контрастирует со степными миграциями во время бронзового века.
Данные популяционной генетики показывают, что люди, похороненные в пещере Вертеба в эпоху позднего энеолита (3790-3535 гг. до н. э.), генетически похожи на другие, ранее проанализированные, геномы из Молдовы. Эти наблюдения в целом предполагают, что люди энеолита археологической общности Триполье-Кукутень произошли от того же или близкородственного населения, которое распространило неолит по большей части Европы (в случае с трипольцами конкретно указывая на баденцев из Венгрии), и практически без признаков метисации с более ранним мезолитическим или неолитическим населением с территории современной Украины, связанным с охотниками-собирателями.
Фактически, большинство трипольцев могут моделироваться энеолитическими популяциями из Европы, имеющими степное происхождение, однако четверо из 20 человек могут быть смоделированы как трипольцы из Молдовы.
Результаты моделирования (qpAdm) предполагают, что существовали различия в составе предков трипольцев, которые могут быть связаны с долей охотников и собирателей, хотя, как отмечалось выше, эта изменчивость недостаточно существенна, чтобы разделить людей с разных участков на разные популяции.
При этом источником генетического пласта, унаследованного от охотников-собирателей у трипольцев, являются в основном западные охотники-собиратели. Значительная часть предков западных охотников-собирателей, обнаруженная у трипольцев (до 18%), может быть связана с увеличением доли этого компонента и в других популяциях среднего неолита Центральной Европы, вероятно, из-за смешения с популяциями на западе, которые уже имели более высокий компонент западных охотников-собирателей, полученный от неолитических групп, связанных с Анатолией до возникновения археологической общности Триполье-Кукутень.
Это также указывает на то, что неолитические популяции охотников-собирателей на территории Украины не внесли большого вклада в происхождение трипольцев. Кроме того, как и в Молдове, выявлен генетический вклад степного происхождения, но в Вертебе он ниже. Это может быть связано с их разными датировками, если предполагать непрерывный импульс движения с востока на запад, постепенно увеличивающий связь с ямной культурой в течение четвертого тысячелетия до н. э.
А вот геном из среднего бронзового века (VERT-113, 1960-1770 гг. до н. э.) значительно отличается от более ранних трипольцев: для него характерен больший вклад кавказских охотников-собирателей/ямной и восточных охотников-собирателей. Результаты (qpAdm) предполагают связь между геномом VERT-113 и представителями культуры шнуровой керамики из Польши. Кроме того, это единственный геном, который ближе к неолиту Украины, чем к западным охотникам и собирателям. Это позволяет предположить, что популяция, возникшая во втором тысячелетии до н. э., возможно, имела общие черты с популяциями неолита на территории Украины.
Интересно, что у генома поздней бронзы (VERT-114) нет значимой связи с геномом средней бронзы (VERT-113). Геном VERT-114 предполагает связь с культурой колоколовидных кубков, несмотря на то, что этот человек почти на 2000 лет моложе конца этого надкультурного феномена, и с датировкой ближе к киммерийцам или скифам. Это подтверждает данные, приведенные в более ранних работах, о притоке западного населения в степь в конце бронзового века.
Однако для понимания более полной картины необходимо получить ещё больше образцов разных периодов из этого региона.
Результаты новой работы имеют важное значение для понимания процесса неолитизации в Восточной Европе. Поскольку население археологической общности Триполье-Кукутень распространилось с современных территорий Румынии и Молдовы в лесостепные районы Западной и Центральной Украины, оно должно было вступить в контакт с населением, связанным с местной буго-днестровской культурой, чья система жизнеобеспечения была сосредоточена главным образом на охоте, рыбалке и собирательстве. Эта группа, вероятно, представляла собой потомков мезолитических охотников и собирателей. Анализ геномов населения пещеры Вертеба предполагает, что местные мезолитические охотники-собиратели не внесли существенного вклада в генофонд более поздних трипольцев. Иными словами, процесс неолитизации в Западной Украине был в большей степени результатом миграции (демической диффузии), а не принятия местными жителями практики земледелия (аккультурации).
Результаты также подтверждают гипотезу длительно существовавшей границы между оседлыми земледельцами лесостепной зоны и соседними кочевыми скотоводами из Причерноморской степи. Эта граница характеризуется резкими контрастами в материальной культуре и способах жизнеобеспечения и, вероятно, сохранялась благодаря этим факторам и языковым различиям. Отсутствие метисации на этой культурной границе является ключом к пониманию контекста, в рамках которого произошла миграция представителей ямной культуры.
В целом результаты показывают, что пещера Вертеба представляет собой важный погребальный комплекс, соединяющий Восток и Запад. Генетическая структура представителей археологической общности Триполье-Кукутень включает предковые компоненты, связанные как с более ранними западными охотниками-собирателями, так и с земледельцами Ближнего Востока, а также те, которые генетически отличаются от предков молдавских представителей этой общности.
Отсутствие местных предков, связанных с охотниками-собирателями эпохи неолита на территории современной Украины, предполагает, что эти земледельцы в основном заменили местных охотников-собирателей и не смешивались с соседними степными популяциями. Кроме того, в течение бронзового века пещера Вертеба использовалась последовательными волнами кочевников-скотоводов с востока, которые в конечном итоге принесли значительные генетические, а также культурные изменения в Европу и в конечном итоге смешались с местными потомками населения трипольской культуры.
Дополнительные генетические данные из этих и более поздних временных периодов позволят дополнить картину, а также ответить на вопросы хронологии и, возможно, указать, причины краха трипольской культуры.
Источник: Gelabert, P., Schmidt, R.W., Fernandes, D.M. et al. Genomes from Verteba cave suggest diversity within the Trypillians in Ukraine. Sci Rep 12, 7242 (2022). doi.org/10.1038/s41598-022-11117-8