О корреляции формы мозга летающих архозавров и занимаемой экологической нише
Среди Диапсид, птичий мозг по объему и соотношению с размером тела не уступает мозгу млекопитающих. Такое увеличение коэффициента энцефализации и соответствующая анатомия мозга объясняется конгитивными функциями, которые требуются для полета: обработка сенсорной информации, контролирование сложного летательного аппарата. Изменения форм мозга, уменьшение размера тела – такие эволюционные изменения могли появиться в результате педоморфоза (способ передвижения), который повлиял на появление остальных характерных анатомических птичьих признаков, в том числе и развитого мозга.
Ранее неинвазивный анализ отпечатков внутренней поверхности черепной коробки ископаемых остатков динозавров был невозможен или затруднен (в случае, если ископаемое по плотности примерно такое же, как и материнская порода, в которой оно сохранилось). Однако сейчас нам доступен детальный анализ с помощью методик КТ и компьютерного моделирования размеров и форм мозга динозавров на заре возникновения полета. Было обнаружено, что мозг раннего летающего авиала Archaeopteryx, представляющего прекрасный пример перехода между нелетающими и летающими тероподами, имеет анатомию, не слишком отличающуюся от таковой у нелетающих Maniraptoriformes1.
У археоптерикса присутствует единственная структура мозга, ассоциированная с полетом - the wulst (бугорки на лобных долях), которая также есть у современных птиц (Neornithes), но полностью отсутствует у нелетающих Maniraptoriformes и мезозойских не-птичьих Ornithurae13,14, Эндокранам архозавров обычно характерны две различные формы при латеральном обзоре. Первому типу характерна удлиненная форма и нечеткие границы между функциональными зонами (Fig. 1). Такая форма наблюдается у современных крокодилов (Fig. 2), большинства не-манирапторовых динозавров и современных рептилий (Fig. 1). Второй тип структуры описан у птерозавров – это выпуклый мозг и четкое разделение на функциональные зоны (Fig. 1). Хотя не-птичьим манирапторам характерны разнообразные формы эндокранов и разграничение функциональных зон (Fig. 1), их эндокраны достаточно выпуклые и компартментализация мозга также высокая. Современные птицы (Neornithes) имеют очень выпуклый мозг с четко разграниченными функциональными зонами (Figs 1 and 2). Также, у птиц присутствует уникальная структура на конечном мозге (передних долях), которой нет у архозавров – the wulst1
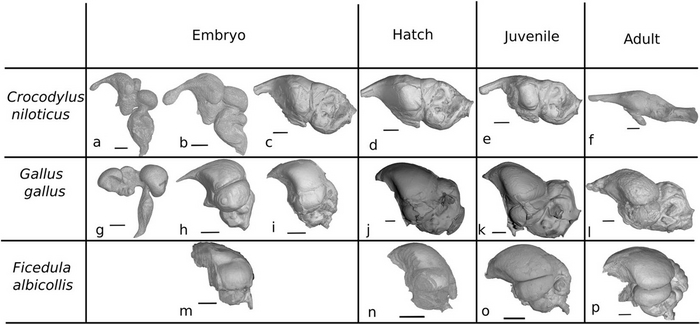
В течение эмбрионального развития внутри яйца Crocodylus niloticus происходит стремительное удлинение мозга и стирание границ между его функциональными зонами (Fig. 2). Вытягивание и распрямление мозга продолжается и после вылупления, но более постепенно, чем при эмбриональном развитии. В итоге, формы мозга крокодила существенно отличаются у эмбриона и при онтогенезе после вылупления.
Для эмбрионального развития курицы Gallus gallus характерно лишь частичное выпрямление мозга, при этом четкая компартментализация мозга сохраняется. Выпрямление мозга G. gallus останавливается после вылупления и не продолжается в течение остальных стадий онтогенеза (Fig. 2). Изменения форм мозга в процессе онтогенеза у страуса Struthio camelus (выборка более ограничена n = 7) и фазана Phasianus colchicus (n = 3) происходят таким же образом, как и у курицы (Fig. 4). Более специализированные птицы, таких как воробьинообразная мухоловка Ficedula albicollis, также сохраняют эмбриональную компартментализацию мозга в течение жизни. Однако, мозг взрослой птицы более куполообразный в сравнении с более сжатым мозгом 6-дневных эмбрионов(Fig. 4).
Другой признак, описывающий изменение формы мозга (РС1) – это отношение длины черепного шва от самого переднего края переднего мозга до самого дорзального края возле черепного отверстия (C) к самому короткому прямому расстоянию между этими двумя точками (D).
И признак Type I, и PC1 описывают изменение формы мозга от почти прямой до выраженной куполообразной.
Куполообразность, выпуклость мозга и коэффициент энцефализации коррелируют у проанализированных видов архозавров. Другой признак, описывающий изменение формы мозга (РС1) – это отношение длины черепного шва от самого переднего края переднего мозга до самого дорзального края возле черепного отверстия (C) к самому короткому прямому расстоянию между этими двумя точками (D).
И признак Type I, и PC1 описывают изменение формы мозга от почти прямой до выраженной куполообразной.
Для не-птичьих манирапторовых, таких как Halszkaraptor или мелкий дромеозаврид Bambiraptor feinbergi (Fig. 3), характерен высокий уровень выпуклости мозга (коэффициент выпуклости C/D = 1.15). Хотя строение тела Halszkaraptor указывает на неприспособленность к полету, размер его тела и выпуклость мозга напоминает таковые у современных птиц. В то же время, Archaeopteryx’у также характерна общая для манирапторов выпуклость мозга. Форма мозга Archaeopteryx’а имеет общие черты с формой мозгов мелких нелетающих манирапторов, в частности, Halszkaraptor. Это может говорить о том, что такая форма могла возникнуть в разное время у разных групп и является результатом параллельной эволюции корневых Парависов, и это, вероятно, не является свидетельством зарождающейся способности к полету. Редукция обонятельных луковиц у современных птиц не была достигнута у Archaeopteryx – это может говорить о более остром обоняни у археоптерикса по сравнению с современными Килевыми птицами. Это также свидетельствует о том, что особенности анатомии мозга, характерные летающим птицам, возникли непосредственно вследствие способности к полету. То есть, строение мозга, типичное для летающих птиц, появилась не до способности к полету, но как следствие адаптации к экологической нише (необходимостью к контролю летательного аппарата, координации и ориентации в пространстве).
Среди птиц, курообразным, гусеобразным и летающим бескилевым (тинамуобразным) по степени выпуклости характерна схожая форма черепа (коэффициент C/D vs log D, показывающий соотношение выпуклости черепа к его длине) (Fig. 6). Примерно похожие показатели коэффициента C/D vs log D наблюдаются и у мелких нелетающих манирапторовых и Archaeopteryx. Все эти виды преимущественно передвигаются по земле. Вполне вероятно, благодаря этой экологической нише (прибрежно-околовоной) указанные виды пережили Меловое вымирание. Те признаки, которые помогли им выжить (в частности, коэффициент C/D vs log D и занимаемая экологическая ниша, околоводный ареал), сохранились в начале палеогена и по сегодняшний день.
Это наблюдение подтверждается также значениями коэффициента C/D vs log D у ископаемых птиц Lithornis and Presbyornis (C/D=1.20 для Lithornis и 1.23 для Presbyornis) – эти значения близки к показателю C/D vs log D cсовременных летающих бескилевых тинаму и гусеобразных; филогенетически эти группы близки к Archaeopteryx.
Распространение видов килевых птиц в новые экологические ниши привело к специализации их летательного аппарата и отразилось на форме их черепа. Хотя способность к полету Archaeopteryx является общепризнанным фактом, его мозг не приспособлен к контролю полноценного полета в сравнении с современными птицами. В то же время, Энанциорнисы были отличными летунами, а мозг базального авиала Cerebavis (circa 122 Ma) имел структуры, необходимые для полета. Однако, строение тела ископаемых базальных килевых птиц говорит о том, что их способности к полету уступали современным птицам. Ни летающих бескилевые, ни гусе- ил курообразные не способны к ловкому полету, и это коррелирует со слабой выпуклостью их мозга. Однако, для видов птиц, которые приспособились добывать пищу благодаря ловкому и высокоточному полету, как, например, соколы, или к убеганию от преследования, маневрируя в трехмерном пространстве леса (с случае сов и воробьинообразных), или же координируют полет перед точным ударом (как это делают олуши), характерен высокий уровень выпуклости мозга.
Таким образом, на основе нашего анализа можно предположить, что занимаемые экологических ниши привели к совершенствованию полета, что далее повлекло за собой ковергентное усложнение мозга, повышение его объема и выпуклости.
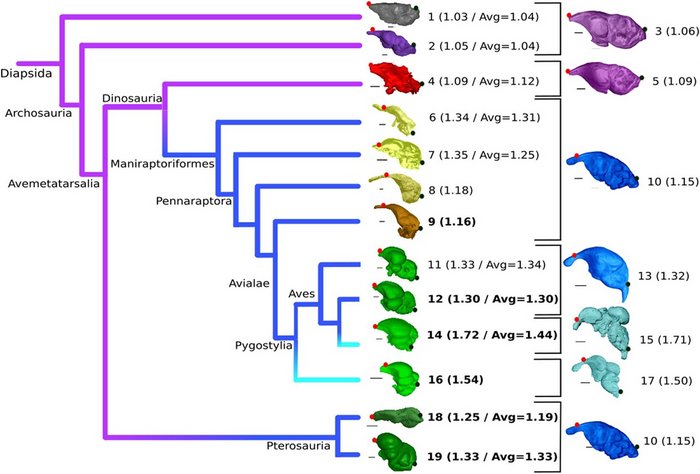
Figure 1. Гетерохронная эволюция формы эндокранов архозавров птичьей стем-группы. Фиолетовые линии указывают на группы с мало выраженной куполообразностью мозга, синие – промежуточную куполообразность, голубые – высокая куполообразность мозга.
Летающие виды выделены жирным, в скобках – группы с малой выборкой. Изображения мозга крокодила на соответствующих стадиях онтогенеза крокодила расположены справа. Реальные размеры мозга не учтены, масштаб для каждого изображения указан в названии образца. Точки указывают передний (красная) и задний (черная) конец мозга. На схеме представлены такие виды:
Podarcis muralis (1; масштаб 0.65 mm; Рептилии);
Caiman crocodilus (2; 5 mm; Crocodylia);
Crocodylus niloticus в возрасте 1 год (3; 2.5 mm);
Arcovenator escotae (4; 20 mm; не-манирапторовый динозавр);
C. niloticus сразу после вылупления (5; 2.5 mm);
Struthiomimus altus (6; 10 mm36; Ornithomimosauria);
Incisivosaurus gauthieri (7; 10 mm96; не-птичий манирапторовый динозавр);
Halszkaraptor escuillei (8; 3 mm);
Archaeopteryx lithographica (9; 2.5 mm);
68-дневный эмбрион C. niloticus (10; 2.5 mm);
Dromaius novaehollandiae (11; 6 mm; Palaeognathae);
Phasianus colchicus (12; 3 mm; Galloanserae);
63-дневный эмбрион C. niloticus embryo (13; 3 mm);
Ficedula albicollis (14; 1.5 mm; Neognathae);
24- дневный эмбрион C. niloticus (15; 1,5 mm);
Cerebavis cenomanica (16; 5 mm);
41- дневный эмбрион C. niloticus (17; 2.5 mm);
Rhamphorhynchus muensteri (18; 5 mm97; Rhamphorhynchidae);
Araripesaurus santanae (19; 4.5 mm; Azhdarchidae).
Морфологический признак Type I отражает изменение формы эндокрана от практически прямого дорзального черепного шва до выраженно-куполообразного (Fig. 5b,d),
Figure 2. Изменение формы эндокрана в онтогенезе крокодила и разных видов птиц.
(a) 29- дневный эмбрион Crocodylus niloticus (масштаб: 1.5 mm);
(b) 41- дневный эмбрион C. niloticus (масштаб: 2.5 mm);
(c) 93- дневный эмбрион C. niloticus (scale: 3 mm);
(d) свежевылупленная особь C. niloticus (scale: 3 mm);
(e) ювенильная особь C. niloticus возрастом 1 год (scale: 4 mm);
(f) взрослая особь C. niloticus (scale: 15 mm);
(g) 5.5- дневный эмбрион Gallus gallus (scale:1.5 mm);
(h) 12- дневный эмбрион G. gallus (scale: 3 mm);
(i) 19- дневный эмбрион G. gallus (scale: 3.5 mm);
(j) трехнедельная ювенильная особь G. gallus (scale: 2 mm);
(k) ювенильная особь G. gallus возрастом 6 недель (scale: 3 mm);
(l) взрослая особь G. gallus (scale: 3.5 mm);
(m) 6- дневный эмбрион Ficedula albicollis (scale: 1.5 mm);
(n) свежевылупленная особь F. albicollis (scale: 2.5 mm);
(o) ювенильная особь F. albicollis (scale: 2.5 mm);
(p) взрослая особь F. albicollis (scale: 2 mm).
Этапы a и g – первая треть эмбрионального развития, этапы b, h и m – половина эмбрионального развития, этапы c и i – финальные стадии развития мозга эмбриона перед вылуплением.
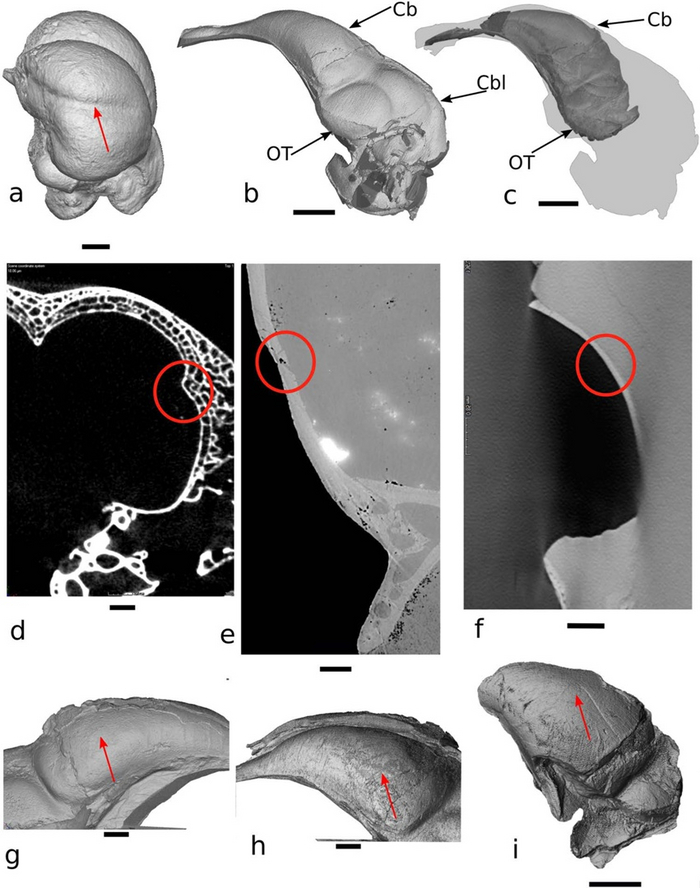
Figure 3. Отпечатки wulst на эндокранах современных птиц и Archaeopteryx.
(a) Трехмерная визуализация эндокрана полярной совы Bubo scandiacus;
(b) Леволатеральный обзор полного эндокрана лондонского образца Archaeopteryx;
(c) Реконструированный эндокран образца Archaeopteryx из Мюниха, спроецированный на лондонский образец Archaeopteryx;
(d)Срез черепной коробки B. scandiacus в том месте, где находится отпечаток wulst (указан красным кружком);
(e) Срез черепной коробки лондонского Archaeopteryx в том месте, где потенциально расположен “wulst1” рядом с местом разлома (красный кружок);
(f) Срез черепа мюнихского Archaeopteryx в зоне, где мог быть расположен “wulst1”;
(g) Детализированный праволатеральный обзор эндокрана лондонского образца
Archaeopteryx;
(h) Детализированный леволатеральный обзор эндокрана лондонского образца
Archaeopteryx;
(i) Леволатеральный обзор области зоны переднего (конечного) мозга мюнихского эндокрана Archaeopteryx.
Красные стрелки указывают положение wulst на поверхности эндокрана B. scandiacus и предполагаемые зоны расположения wulst на эндокранах лондонского и мюнихского образцов Archaeopteryx.
Аббревиатуры: Cb-Cerebrum; Cbl-Cerebellum; OT-Optic tectum. Scale bar: a-4 mm; b-c-
3.5 mm; d-0.25 mm; e-0.3 mm; f-0.85 mm; g-h-i-1.5 mm.
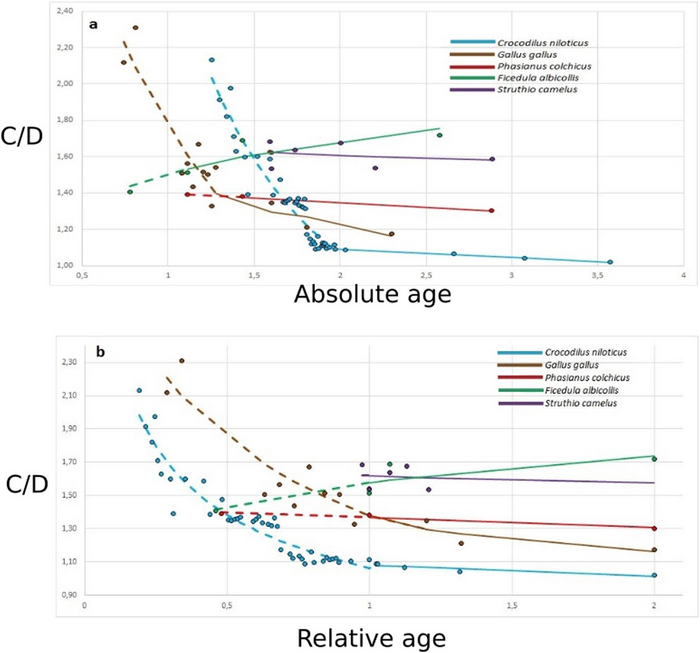
Figure 4. Зависимость возраста образцов архозавров от коэффициента куполообразности мозга (C/D). На графике представлен абсолютный возраст представлен в днях (a) и относительный возраст, соотнесенный с этапом онтогенеза до или после вылупления (1; b). Стадии развития до вылупления показаны прерывающейся линией , после вылупления – непрерывной линией. Отметки справа указывают на возраст половой зрелости.
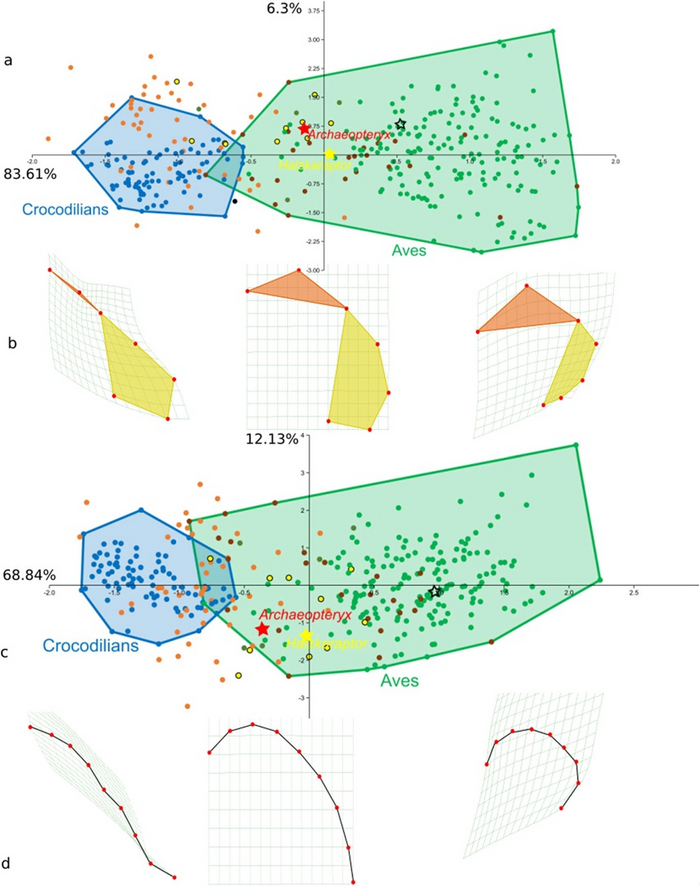
Figure 5. Распределение признака Type I и РС1 (длина и форма дорзального черепного шва, указывающая на степень выпуклости и изменение формы мозга).
(a) Распределение выраженности признака Type I;
(b) Усредненное схематическое изображение типичной формы мозга по признакам PC1 (куполообразность) и Type I (длина дорзального шва);
(c) График Principal Component Analysis (сухая статистика – прим. переводчика);
(d) Линия, показывающая общее изменение формы мозга (контур дорзального черепного шва).
Образцы крокодилов указаны голубыми точками, область распределения признаков крокодила ограничена голубой чертой. Не-манирапторовые динозавры указаны оранжевыми точками, Манирапторовые - желтыми точками, Paleognathae (Бескилевые птицы) коричневыми точками, Neognathae (Килевые) – зелеными точками. Область распределения признаков птиц (Paleognathae + Neognathae) ограничена зеленой линией.
Вариации форм эндокранов (b,d) указаны красными точками. Окрашенные области (рис. b) обозначают функциональные зоны мозга – конечный (передний) мозг (оранжевый) более нижние отделы (средний+промежуточный) (желтый).
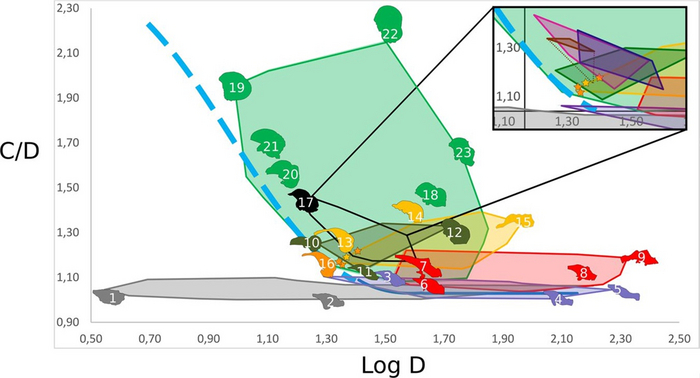
Figure 6. Кривая зависимости выпуклости мозга от длины черепа. Голубым уветом указаны мозги крокодилят до вылупления. Траектории изменения форм в онтогенезе указаны прерывчатой голубой линией до вылупления, непрерывной – после. Разноцветные области распределения признаков соответствуют таким группам животных: Рептилии (серый), не-манирапторовые динозавры (красный), не-птичьи Манирапторы (желтый), Птерозавры (темно-зеленый), летающие не-птичьи авиалы (черный), нелетающие Бескилевые и Килевые птицы (светло-зеленый). Вставка показывает разнообразие эндокранов вымерших летающих не-птичьих авиалов: летающие Paleognathae (коричневый), Anseriformes (синий), Galliformes (темно-розовый). Прерывчатая коричневая линия указывает на принадлежность вымершей летающей птицы Lithornis plebius к Бескилевым. На схеме указаны эндокраны таких видов:
1-Podarcis muralis;
2-Varanus exanthematicus;
3-Caiman crocodylus;
4-Crocodylus niloticus;
5-Alligator mississipiensis;
6-Heterodontosaurus tucki;
7-Psittacosaurus lujiatunensis;
8-Arcovenator escotae;
9-Tyrannosaurus rex;
10-Rhamphorhynchus muensteri;
11-Paraspicephalus purdoni;
12-Araripesaurus santanae;
13-Halszkaraptor escuillei (желтая звездочка);
14-Incisivosaurus gauthieri;
15-Struthiomimus altus;
16-Archaeopteryx lithographica (оранжевая звездочка);
17-Phasianus colchicus;
18-Leptoptilos crumeniferus;
19-Thalurania furcata;
20-Cerebavis cenomanica;
21-Ficedula albicollis;
22-Strix nebulosa;
23-Struthio camelus.
Источник: https://vk.com/dromeo