Представители гаплогруппы R были праиндоевропейцами. Позже гаплогруппа R распалась на две ветви — R1 и R2.
R2 — это цыгане.
R1 распалась на две ветви — R1a (арии) и R1b (кельты).
R2 (цыгане) взаимодействовали с R1a (ариями) и стали говорить на арийском языке.
R1a + I2 — славяне
R1a + R1b + I1 — германцы
R1b + J + E1b1b1 + I2 — романцы
R1a + R1b + J + E1b1b1 — греки
R1a + R1b + J + E1b1b1 + G — армяне
R1a + R1b + I2 — албанцы
R1a + R1b + G — осетины
У индоевропейцев был общий этноним — русы. Корень «рус» в разных вариациях распространён по всей индоевропейской территории.
Король Артур был русом-сарматом, а сарматы — это кельты (R1b).
Кельты (R1b), поселившись в Иберии, образовали племя кельтиберов, ассимилировали местные племена (представителей гаплогруппы I1). Баски на 10% являются потомками по мужской линии этих племён, по женской линии баски являются носителями гаплогруппы U5, которая коренная в Европе. То есть получается, женщины передали баскам язык иберских племён.
Между R1a и R1b появилась конкуренция, что отразилось в индоевропейской мифологии — война асов и ванов, война асуров и дэвов. Они то воюют, то мирятся.
С развитием генетики, интерес вызывают не только вопросы о происхождении популяций, но также и отдельных людей, особенно если они принадлежали знатному роду. В частности, вопросы о происхождении элиты средневековой Руси IX–XVI вв., где важнейшими действующими лицами были представители правящего рода Рюриковичей, долгое время были предметом дискуссий. Ранее отцовские линии в виде гаплогрупп Y-хромосомы Рюриковичей были реконструированы по биологическим материалам современных людей, чьё происхождение, c разной степенью достоверности восходит к Рюрику. А что касается исследований древних геномов, то их осложняют трудности в идентификации останков Рюриковичей и другой знати. Поскольку до начала XV в. имена погребенных отсутствовали на погребальных сооружениях, надгробиях и саркофагах. Места погребений той эпохи устанавливаются по летописным сообщениям и спискам имён для поминовения XVI–XVII вв. Помимо этого практики повторного использования старых погребальных сооружений, перемещение почитаемых останков и их изъятие в ходе антирелигиозной кампании в советское время, привели к тому, что значительная часть останков XI–XIV вв. из погребений в древнерусских храмах и не только, оказалась утрачена или не может быть достоверно связана с определенными историческими личностями.
1/2
Рака с мощами св. Александра Невского
К примеру, в 1967 году на территории Черниговского Спасо-Преображенского собора во время строительных работ был случайно обнаружен череп мужчины 25-35 лет со следами ран от меча, без археологического документирования. Останки были предположительно отнесены одному из внуков Ярослава Мудрого - Глебу Святославичу, князю тмутараканскому и новгородскому, жившему в 11 веке. Однако точно определить кто был похоронен в каменном саркофаге возле Черниговского Спасо-Преображенского собора не представляется возможным. Особенно в свете недавнего анализ ДНК этого человека, который показал, что он был носителем гаплогруппы Y-хромосомы I2a1a2b1a1a и обладателем митохондриальной линии H5a2a.
Датировки (только археологические): Саркофаг найден на заднем дворе собора. Глубина верхней плиты саркофага составляла 1,9-2,0 м от современной поверхности. Стратиграфия несколько пострадала от более позднего нарушения грунта: трубы водоснабжения проложены на высоте 0,2 м над изголовьем саркофага. Археологи относят его к XI веку нашей эры. гаплогруппа – Y-хромосомы I2a1a2b1a1a гаплогруппа мтДНК – H5a2a
Поэтому особую ценность представляют те немногочисленные захоронения, где костные останки по археологическому контексту, антропологическим данным и комплексу исторических свидетельств надёжно отнесены к представителям рода Рюриковичей.
К таким достоверным захоронениям относится погребение в Спасо-Преображенском соборе Переславля-Залесского князя Дмитрия Александровича, второго сына Александра Ярославича Невского. Прямые сообщения о погребении Дмитрия Александровича в Спасо-Преображенском соборе содержит ряд летописей, в том числе Новгородская четвёртая, Воскресенская, Никоновская летописи, а также Московский великокняжеский свод конца XV века.
А поиск места погребения был облегчен ещё и тем фактом, что после захоронения князя Дмитрия Александровича в 1294 году и его наследника бездетного старшего сына Ивана в 1302 году с последующим переходом Переяславского княжества Москве, этот храм как престижная усыпальница использовался реже чем многие другие в средневековье. Изучив все доступные документы, можно было полагать, что погребение Ивана Дмитриевича находилось в юго-западной части собора, под хорами, а погребение князя Дмитрия Александровича – в юго-восточной части, в южной алтарной апсиде, где до реставрации 1891 г. находились отмечавшие места погребений более поздние кирпичные надгробия XVII в. В результате сам саркофаг с останками, был обнаружен отрядом Института археологии РАН под руководством Владимира Валентиновича Седова в 2014 г.
1/2
Князь Дмитрий Александрович унаследовавший после смерти великого князя Александра Ярославича Невского – Переславское княжество, в разные годы также был новгородским князем и великим князем владимирским. Князь умер в 1294 году на Ламском Волоке, возвращаясь в Переславль из Твери, и был похоронен в Спасо-Преображенском соборе Переславля.
Результаты
Палеогенетический анализ
В результате полногеномного секвенирования фрагментов древней ДНК из наиболее информативного образца - ладьевидной кости стопы Nev2.3, сгенерировано более 15 млн чтений, определены 532154 однонуклеотидных полиморфизма (ОНП). В процессе обработки данных секвенирования установлено, что это был мужчина с митохондриальной гаплогруппой F1b1 и гаплогруппой Y хромосомы N1a1a1a1a1a1a7a~ N-Y59140.
1/2
Анализ Y-хромосомы князя Дмитрия Александровича и других предполагаемых представителей рода Рюриковичей
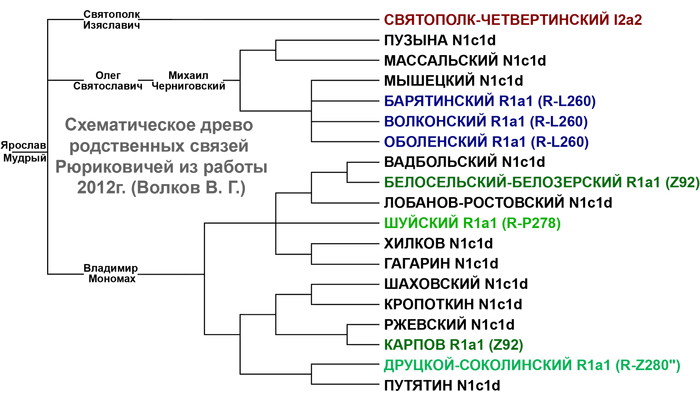
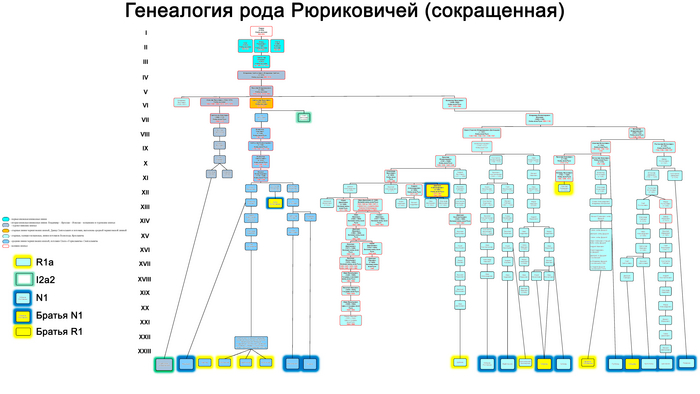
На основании результатов генетических исследований трех древних потомков Рюрика и предполагаемых 43 современных Рюриковичей выдвинуты гипотезы о трех возможных отцовских линиях Рюрика, а именно N1a, R1a и I2a.
В результате филогенетического позиционирования Y хромосома князя Дмитрия Александровича кластеризовалась с Y хромосомами современных вероятных потомков Рюриковичей, принадлежащих к гаплогруппе N1a, которые произошли из различных аристократических родов: Мстиславичей и Юрьевичей, потомков Мономаха от великого князя Мстислава Владимировича и великого князя Киевского Юрия Долгорукого, а также Ольговичей, черниговских Рюриковичей, потомков князя Олега Святославича или «Гориславича».
1/2
При этом важно отметить, что вероятные захоронения князя Глеба Святославича Черниговского, из рода Ольговичей, с линией I2a и Изяслава Ингваревича Луцкого, из рода Мстиславичей, с линией R1a, не подтверждены археологическими данными, что ставит под сомнение и гипотезы, которые вытекают из генетического анализа указанных образцов.
В целом как древние, так и современные предполагаемые Рюриковичи с линиями гаплогрупп R1a и I2a, обладают высокой гетерогенностью нуклеотидной последовательности Y хромосом и не указывают на единое происхождение по прямой мужской линии. В то время как большинство ныне живущих потомков Рюрика по прямой мужской линии согласно их родословным, с гаплогруппой N1a, обладают максимально похожими вариантами Y хромосомы как между собой, так и с Y хромосомой князя Дмитрия Александровича. Что совместно с полногеномными данными средневековых и современных Рюриковичей указывает на то, что, как минимум, со времени великого князя Ярослава Мудрого (XI в), мужчины этого рода были носителями гаплогруппы Y-хромосомы N1a.
Поиск живших в прошлом людей наиболее близких к князю Дмитрию Александровичу по Y-хромосоме
На филогенетическом дереве Y хромосомы видно, что помимо современных Рюриковичей, линия Дмитрия Александровича группируется с древними людьми середины II тысячелетия до н.э. Оленеостровского могильника в Кольском заливе Баренцева моря.
В предыдущих работах на примере этих образцов был показан поток генов из Сибири на север и восток Европы (восточно-евразийская компонента).
1/3
Высокая степень биологического соответствия Y-хромосомы представителя русского княжеского рода и людей эпохи раннего металла привела авторов работы к гипотезе о возможном вкладе восточно-евразийского генофонда в формирование североевропейского населения раннего средневековья, поскольку, согласно летописям, Рюриковичи имели с ними генетическую связь.
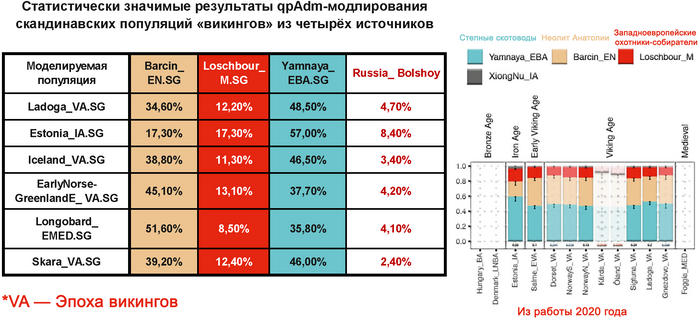
Авторы работы исследовали вклад людей с Большого Оленьего острова в формирование средневекового населения, жившего вдоль побережья Балтийского моря из современных Финляндии, Дании, Швеции и Норвегии, используя геномы периода «викингов» из работы 2020 года, обзор которой тоже есть на канале. В статистических анализах удалось выявить существенный вклад геномов древних представителей Большого Оленьего острова в формирование генофонда популяций периода «викингов», скорее всего, благодаря контактам последних с финно-угорским населением прибалтийского региона. При этом у представителей периода «викингов» с гаплогруппой N1a этот вклад выше чем у носителей линий R1a. Результаты показывают, что в формировании генофонда отдельных жителей из Швеции, Норвегии и Балтийского региона участвовали и люди из Сибири с восточноазиатскими корнями. При этом в наиболее подходящих моделях, помимо европейских охотников-собирателей, неолитических земледельцев и степных скотоводов, скандинавские популяции моделировались с участием обитателей Большого Оленьего острова или представителей хунну железного века.
Здесь стоит адекватно воспринимать компоненты в моделях. В данном случае хунну добавлены для проверки географического происхождения компоненты из Сибири, если говорить упрощённо.
А что касается предков князя Дмитрия Александровича по мужской линии, то с большой вероятностью, среди них были мужчины, оставившие могильник на Большом Оленьем острове в Кольском заливе Баренцева моря около 3600 лет назад.
Анализ мтДНК князя Дмитрия Александровича
Митохондриальную гаплогруппу F1b1, определенную у князя Дмитрия Александровича относят к восточно-евразийскому кластеру, она встречается с разной частотой у большинства исследованных древних и современных популяций Байкальского региона и сопредельных территорий Центральной Азии.
Погребения мужчин с митохондриальными линиями F1b1b и F1b1f, а также с линией Y-хромосомы N1a1a1a1a3a (F4205) характеризовались довольно богатым погребальным инвентарем с золотыми и позолоченными предметами, что указывает на их принадлежность к знати.
● A1811 – Женщина, захороненная с костями овец или коз. мтДНК F1b1f ● A1812 – Мужчина из богато обставленной могилы, с золотыми серьгами, золотыми заколками для волос, ремешками из золотой фольги с переплетающимися узорами из серебра и мечом, украшенным золотыми пластинами. мтДНК – F1b1f, Y-хромосомы – N1a1a1a1a3a (F4205) ● A1821 – Пожилой мужчина, захороненный с исключительно богатыми погребальным инвентарём, включающим позолоченный меч и пояс. мтДНК – F1b1b, N1a1a1a1a3a (F4205)
Однако с уверенностью говорить о происхождении материнской линии князя Дмитрия Александровича довольно сложно, так как практически во всех исторических периодах прослеживается повышенная изменчивость и разнообразие митохондриальных гаплогрупп в одном регионе.
Здесь также важно понимать разницу между генофондом популяций и отдельно взятых людей, особенно представителей знатных родов, у которых была широко распространена практика привлечения самых разных по своему происхождению женщин для союзов, официального и неофициального характера, в качестве выгодных или вынужденных политических шагов. Поэтому материнская линия F1b князя Дмитрия Александровича могла быть связана как с древними миграциями из Сибири, так и с миграциями кочевников раннего средневековья. При этом у этих разных по времени групп могло быть общее происхождение этих линий.
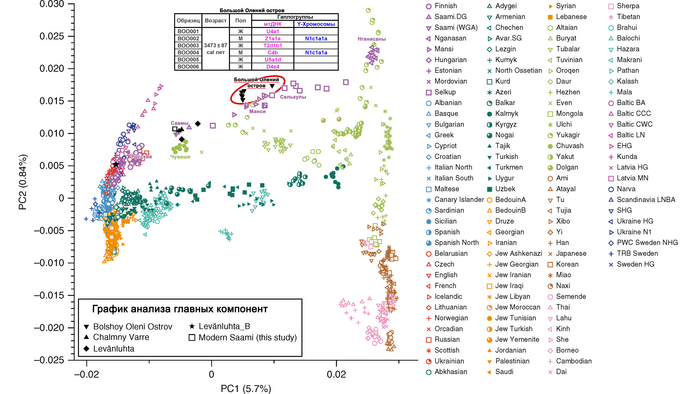
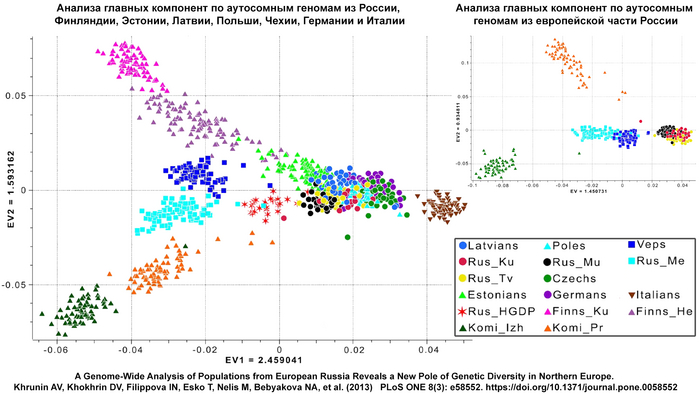
Результаты анализа главных компонент
Генетическую близость генома князя Дмитрия Александровича к другим известным древним и современным популяциям оценили с помощью метода главных компонент (PCA).
Сразу стоит отметить, что надо адекватно понимать положение на этом графике отдельных представителей популяций, особенно из знатного рода. Если положение большинства образцов сочетается с их географией, то положение отдельно взятых людей от смешанных союзов будет где-то между положением его родителей. При этом генетическая схожесть в этом анализе не подразумевает общую историю, а лишь схожие пропорции генетических компонент.
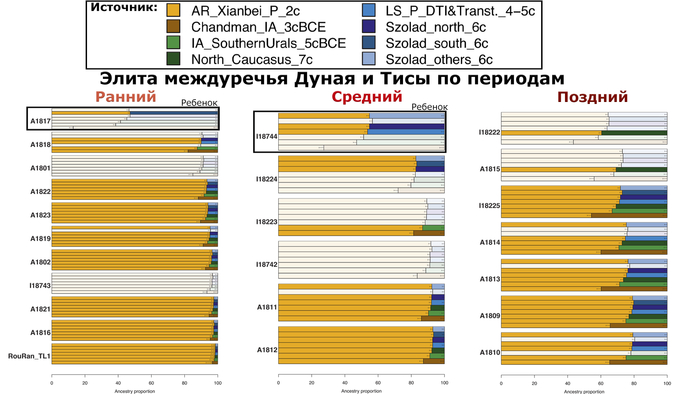
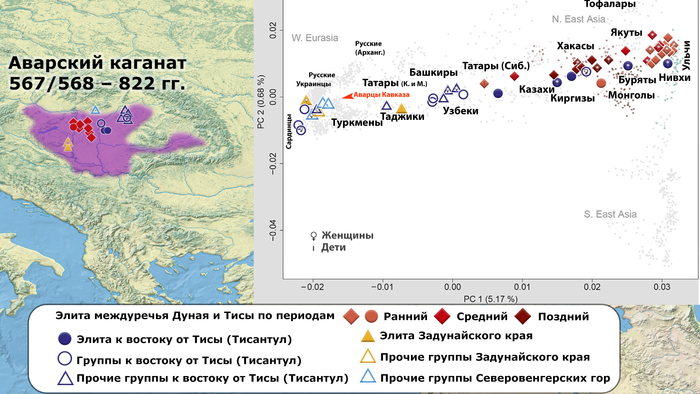
В нашем случае геном князя Дмитрия Александровича занимает промежуточное положение между древними и современными образцами из Европы и Центральной Азии. Наиболее близкими генетически и хронологически к князю Дмитрию Александровичу, расположены некоторые представители авар позднеаварского периода из междуречья Дуная и Тисы, а также из предполагаемого вторичного центра власти Аварского каганата к востоку от Тисы.
Здесь стоит напомнить, что в Аварском каганате (VI–IX вв.), помимо европеоидного большинства ярко выраженный монголоидный тип, имели, преимущественно, представители элиты, который размывался со временем к позднеаварскому периоду.
Как известно из работы 2022 года, популяции аварского периода демонстрируют значительную генетическую изменчивость, а их образцы разбросаны на графике анализа главных компонент от популяций Западной Евразии до популяций Северо-Восточной Азии.
Однако несмотря на эту общую неоднородность, существуют четкие закономерности генетической подструктуры, соответствующие как хронологии и географии Карпатского бассейна, так и социально-археологическим категориям. Почти все представители элиты раннеаварского периода образуют плотный кластер с высоким содержанием компоненты «древних северо-восточных азиатов» (АНА) до 98%. Однако образцы представителей элиты позднеаварского периода уже сильнее смещены в сторону Западной Евразии, а количество родословной «древних северо-восточных азиатов» у них сократилось уже до 70-80%.
При этом среди простых жителей раннего и позднего аварского периодов преобладал более западный генетический профиль, демонстрирующий сильную связь с населением Карпатского бассейна доаварского периода, с нулевым или небольшим вкладом компоненты «древних северо-восточных азиатов». И как раз именно к этой группе образцов относится расположенный рядом с князем Дмитрием Александровичем представитель позднеаварского периода со смешанным генетическим профилем (Hungary_LateAvar), примерно, на 80% состоящим из компоненты максимально выраженной у предшествующих местных жителей Карпатского бассейна и на 20% из компоненты «древних северо-восточных азиатов» из восточной степи.
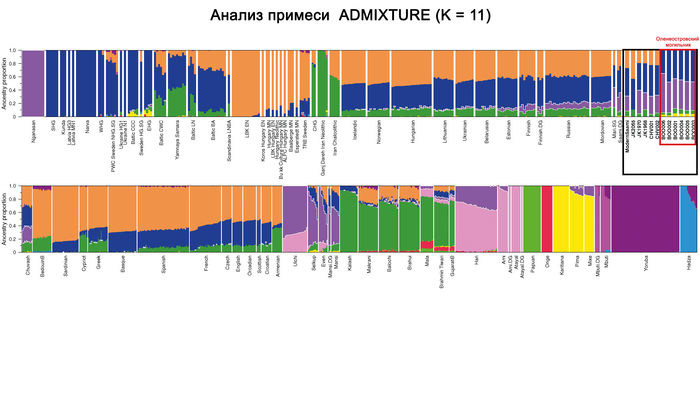
Анализ примесей (Admixture)
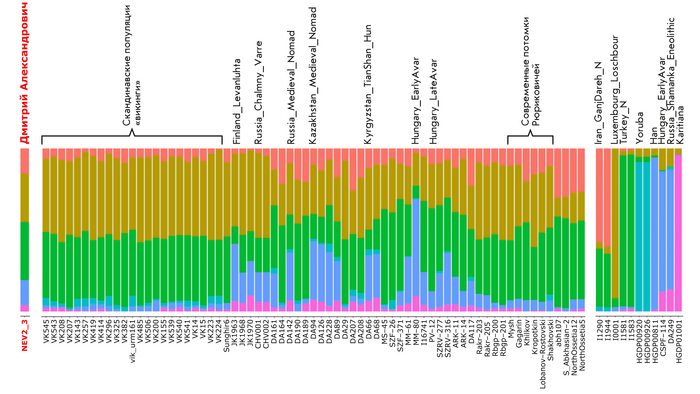
В анализе примесей, по предковым компонентам отмечено генетическое сходство князя Дмитрия Александровича с представителями раннесредневекового населения востока Скандинавии периода викингов.
Что может свидетельствовать в пользу версии о «варяжском» (скандинавском) происхождении Рюрика, родоначальника княжеского рода, призванного на Русь, о чем прямо говорит Летопись. При этом авторы, используя термин «викинги», подчёркивают, что это неоднородная и сложная по своему историческому формированию европейская популяция, объединенная лишь образом жизни и ареалом проживания. Однако есть и отличие генома князя от скандинавских популяций эпохи «викингов», в том числе с территории современной России, это отличие связано с наличием у него дополнительной восточно-евразийской компоненты (обозначенной синим цветом на графике), отсутствующей в таком количестве у подавляющего большинства «викингов». Эта компонента максимально выражена у нганасан, финно-угорского народа Западной Сибири манси, представителей народа хань в Китае, у элиты авар из междуречья Дуная и Тисы (Hungary_DanubeTisza_MLAvar), а также у более древних представителей эпохи раннего неолита Байкала и Монголии (Russia_Shamanka_Eneolithic. SG и Mongolia_North_N). В меньшей степени эта компонента обнаружена у жителей раннего средневековья с территории современной Финляндии (Finland_Levanluhta), у кочевника Прикаспийской степи (Russia_Medieval_Nomad), современника князя Дмитрия Александровича, а также у более древних ираноязычных степных кочевников железного века с территорий современных Казахстана и Кыргызстана (Kazakhstan_TianShan_Saka, Kyrgyzstan_TianShan_Hun). Суммарно, на основании данных анализа главных компонент, анализа примесей и информации о митохондриальной ДНК можно утверждать, что в геноме князя Дмитрия Александровича имеется существенный вклад восточной предковой компоненты, что отличает его от раннесредневекового населения востока Скандинавии, «викингов» и средневекового славянского образца из Владимира (Сунгирь6), но сближает в первую очередь с древним населением Финляндии, Кольского полуострова и раннесредневековым населением Центральной Европы с этой компонентой. Вероятно, этот вклад получен и по мужской, и по женской линиям, что соответствует процессам древней миграции из Сибири по степному евразийскому коридору в 1 тыс. до н.э.–1 тыс. н.э.
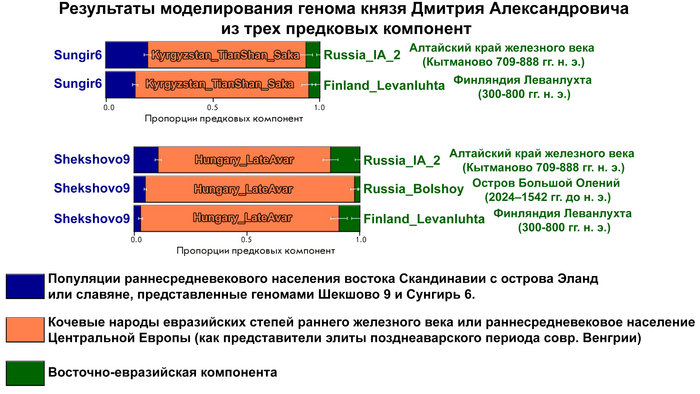
Моделирование генома князя Дмитрия Александровича из геномов предковых популяций
После анализа различных комбинаций предковых компонент наиболее подходящая модель указывает на вклад в происхождение князя Дмитрия Александровича трех компонент, как у следующих популяций:
Раннесредневекового населения востока Скандинавии с острова Эланд. Или славян, представленных геномами Шекшово9 и Сунгирь6.
Представителей степных кочевых народов евразийских степей железного века или раннесредневекового населения Центральной Европы (степных кочевников с территории Венгрии).
Восточно-евразийской компоненты.
Предсказание фенотипических признаков князя Дмитрия Александровича на основании генетических данных
С большой вероятностью глаза князя Дмитрия Александровича были карими (P = 0.962), волосы темными (P = 0.810) или коричневыми (P = 0.555), а оттенок кожи промежуточным (0.635), т.е. не светлым и не темным.
Выводы
Благодаря палеогенетике и применению статистических методов проверки гипотез, подтверждается связь Рюриковичей со Скандинавией по мужской линии, хотя отсутствие генетических данных от населения с территории севера Руси в период, непосредственно предшествовавший становлению княжеского правления, не позволяет предметно судить о присутствии предков князя Дмитрия Александровича и других Рюриковичей на материковом севере и северо-востоке Европы. Анализ генеалогического древа Рюриковичей показал, что современные представители этого рода, с гаплогруппой Y-хромосомы N1a, группирующиеся с образцом князя Дмитрия Александровича, относятся к трем разным ветвям этого рода – Ольговичам, Мстиславичам и Юрьевичам. Которые восходят к князю Ярославу Мудрому, как к ближайшему их общему предку.
В свою очередь митохондриальная гаплогруппа F1b1 у князя Дмитрия Александровича может свидетельствовать о вкладе восточных популяций в его генотип, что подтверждается и аутосомными данными. Согласно результатам анализа главных компонент и анализа примесей, геном князя Дмитрия Александровича имеет сходство с геномами представителей скандинавских популяций эпохи «викингов», но отличается от них заметным вкладом восточной компоненты. Что может быть связано с распространенной практикой браков русских князей с дочерями половецких ханов с конца XI в. Такие браки скрепляли союзнические отношения и усиливали политическое взаимодействие.
Мать Дмитрия, супруга Александра Невского – Александра Брячиславна происходит из рода полоцких Изяславичей. Сведения о женах этих князей скудны, имя и происхождение матери Александры неизвестно. Однако, известно, что представители полоцкой ветви Рюриковичей не избегали брачных союзов с половчанками. Из половецкого рода происходила вторая жена полоцкого князя Святополка Изяславича (1050–1113), Елена, дочь хана Тугоркана.
Однако восточная компонента в геноме Дмитрия Александровича могла быть получена одновременно и по мужской, и по женской линиям, отражая браки Рюриковичей с представительницами династий Центральной и Южной Европы, как сербские Вукановичи или венгерские Арпады, к примеру. Элиты этих территорий могли в большей степени, чем «простое» местное население, сохранить восточные гены, привнесенные миграционными потоками первого тысячелетия нашей эры. Исходя из перечисленных вероятностей, объяснения присутствия «восточной компоненты» в геноме Дмитрия Александровича, пока остаются гипотетическими, как и состав его предков по материнской линии.
Но несмотря на оставшиеся вопросы, впервые на примере представителя древнего рода Рюриковичей показана генетическая составляющая сложного характера межэтнических взаимодействий в формировании знати средневековой Руси.
Источник: Жур К.В., Шарко Ф.С., Седов В.В., Добровольская М.В., Волков В.Г., Максимов Н.Г., Сеславин А.Н., Макаров Н.А., Прохорчук Е.Б. Рюриковичи: первый опыт реконструкции генетического облика правящего рода средневековой Руси по данным палеогеномики // Acta Naturae. - 2023. - Т. 15. - №3. - C. 50-65. doi: https://doi.org/10.32607/actanaturae.23425
Примечание: текста очень много, тема сложная и состоит из двух постов с полным названием "Генофонд народов Восточного Кавказа на основе гаплогрупп Y-хромосомы и аутосомных данных". Выводы к обеим частям во второй части, после аутосомных данных.
Примечание к видео: там, где я говорю: "включая линии" P58, M67, M12, M458 и другие субклады, ошибка! Правильно "без этих линий" Пример: J2(×M67, M12) - надпись в скобках обозначает отсутствие этих мутаций, т.е., не включая этих ветвей. (Коррекцию предложил внести один из соавторов работы – Насиб Искандаров)
Вступление
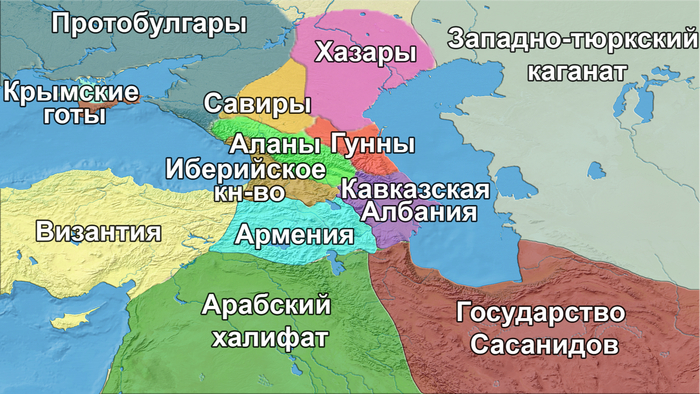
Восточный Кавказ, который охватывает современные Дагестан и Азербайджан, является важным сухопутным мостом, соединяющим Европу с Передней или Западной Азией (далее буду употреблять термин — Западная Азия). Одним из путей через Кавказ является Большой Кавказский проход, прибрежная полоса которая проходит между горами Большого Кавказа и Каспийским морем от Дербента до Сумгаита. Эта территория была населена представителями рода людей, носителями олдувайской культуры, ещё около 2 миллионов лет назад.
*Не предки современных людей
А что касается людей современной анатомии, то древнее поселение Чох в нынешнем Дагестане является ключевым памятником культур мезолита, неолита и бронзового века, которые раньше существовали в горных районах Северо-Восточного Кавказа. Данные недавнего радиоуглеродного датирования свидетельствуют о том, что одомашнивание животных, выращивание сельскохозяйственных культур и изготовление керамики в этом регионе началось не позднее конца 7-го или начала 6-го тысячелетий до нашей эры.
Экономическое развитие в период неолита (земледелие и животноводство, появление керамики, ткачества, новых технологий камнеобработки) привело к быстрому росту населения и колонизации новых территорий. На территории современного Азербайджана развитые неолитические общества возникли на заре 6-го тысячелетия до нашей эры как преемники неолитических обществ Юго-Западной Азии. Открытие металлов в эпоху энеолита (~7 тыс. лет назад) стимулировало развитие технологий. Бронзовый век (6-4 тыс. лет назад) ознаменовался появлением новых методов обработки металла, экономическими и социальными изменениями, включающими социальное неравенство и племенные объединения, а также торговыми отношениями с населением Юго-Восточной Европы и Западной Азии. А уже в среднем бронзовом веке на территорию Восточного Кавказа прибыли кочевники из евразийских степей, привнеся в регион традиции курганных захоронений.
В VII–VI веках до нашей эры на юг региона через Дербентский перевал проникли ираноязычные скифы, которые образовали военно-политический союз, известный как Ишкуза, или Скифское царство, на землях современного Западного Азербайджана и Северо-Западного Ирана. Позже эта территория была присоединена к Мидийскому царству. В конце первого тысячелетия до нашей эры из союза племён в северной части региона возникло государство Кавказская Албания. У него была своя собственная письменность, и его население приняло христианство в IV веке нашей эры. Кавказская Албания в значительной степени состояла из автохтонных кавказских племен, говоривших на лезгинских языках нахско-дагестанской семьи, хотя ее господство распространялось и на некоторые иранские племена.
Территория современного Дагестана была колыбелью таких ранних государств, как Лакз, Табасаран, Кайтагское уцмийство, Сарир, Царство Гумик и др., возникших в первой половине 1-го тысячелетия нашей эры.
В течение первых нескольких столетий нашей эры на Кавказ прибывали другие иранские племена, включая сарматов, маскутов и алан. А Южный Кавказ подвергался набегам гуннов, савиров и хазар, приходившим с севера через Дербентский перевал в раннем средневековье (между концом IV и VIII веками).
В VII веке нашей эры север равнинного Дагестана был захвачен развивающимся Хазарским каганатом, в то время как юг региона был в составе государства Сасанидов с IV века.
В VII–IX веках Восточный Кавказ контролировался Арабским халифатом, который проводил агрессивную миграционную политику, приведшую к исламизации местного населения.
Вторжения тюркоязычных сельджуков, основателей могущественной империи, охватывающей Центральную и Западную Азию, а также Южный Кавказ, начались в XI веке, когда захватчики продолжали экспансию из Средней Азии на Восточный Кавказ, включая Дербент.
Тюркские народы из Половецкой или Кыпчакской степи, мигрировали на Южный Кавказ в XI- XII веках, а их потомки, возможно, внесли свой вклад в возникновение карапапахской группы среди азербайджанцев.
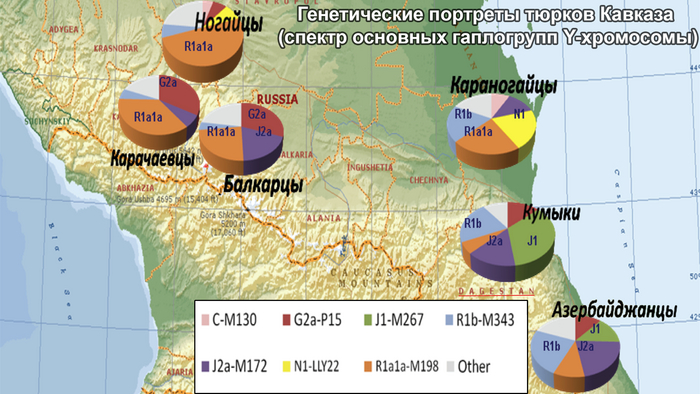
Другими тюркоязычными народами Восточного Кавказа являются кумыки и ногайцы. Единого мнения о происхождении кумыков нет. Преобладающая гипотеза прослеживает их происхождение от местного автохтонного населения, имевшего тесные этнокультурные контакты с чужеземными тюркскими племенами, такими как савиры, хазары или кыпчаки (половцы). В свою очередь, ногайцы были кочевниками поздней миграционной волны; большинство из них осело в Дагестане и представлено караногайцами.
После Монгольских завоеваний, с XIV века, Дагестан снова попал под исламское влияние.
Местные средневековые княжества того времени просуществовали до XIX века, когда они были поглощены Российской империей.
Южные регионы Восточного Кавказа находились под иранским влиянием в течение XIV-XIX веков; они были включены в состав Российской империи после русско–персидских войн, когда в 1813 и 1828 годах были подписаны мирные договоры. Позже эти территории вошли в состав Азербайджана. Это очень сокращённая история демонстрирует, что на генофонды автохтонных популяций Восточного Кавказа оказали влияние как массовые древние миграционные волны ираноязычных народов, так и более поздние миграции евразийских тюркоязычных народов. Богатая история региона отражена также и в языковом разнообразии Восточного Кавказа.
Языки региона включают нахско-дагестанские языки (на которых говорят более 30 этнических и этнографических групп), тюркские языки алтайской языковой семьи (на которых говорят азербайджанцы, кумыки и караногайцы) и иранские языки индоевропейской языковой семьи (на которых говорят таты, горские евреи, езиды, курды и талыши). Такое значительное разнообразие популяций Восточного Кавказа и их необычайная структура является серьезным препятствием в изучении генофонда народов, проживающих в регионе.
Некоторые предыдущие популяционные исследования указывают на сходство между генофондами азербайджанцев и их соседей на Кавказе и в Западной Азии. Азербайджанцы генетически родственны тюрко- и ираноязычному населению Западной Азии, но отличаются от туркмен. А иранские азербайджанцы имеют генетическое сходство со своими ираноязычными соседями, в то время как представители этнографической группы азербайджанцев Дагестана - терекеме демонстрируют генетическое сходство с кумыками.
1/3
В целом опубликованные данные свидетельствуют о том, что 18% генофонда азербайджанцев отражают средневековые миграции из Центральной Азии и 6% связаны с миграциями из Восточной Азии.
А полногеномные данные азербайджанцев Северо-Западного Ирана рассмотренные в работе, посвященной следам тюркской экспансии, указали на 5%-й вклад восточноевразийской предковой компоненты, что можно объяснить распространением тюркских языков на запад согласно модели «доминирования элит» (это когда происходит смена языка без значительного изменения генофонда).
При этом генетических данных из Азербайджана было очень мало. А ведь именно эта территория является значимой для выявления многовекового влияния Персии на регион, в том числе и с точки зрения генетики. При этом интересно: распространялось ли это генетическое влияние только на южные территории или на весь Каспийский регион? Кроме того, данные из восточных регионов Кавказа и Ирана могут также прояснить и генетическую историю ряда малочисленных народов Северного Кавказа.
В более изученных популяциях Дагестана обнаружена значимая корреляция между частотами гаплогрупп, их географией и языком (о чем свидетельствуют лексикостатистические данные).
При этом несмотря на сильное межпопуляционное разнообразие коренных народов Дагестана была подтверждена гипотеза о происхождении всех нахско-дагестаноязычных популяций от одной прапопуляции около 6000-6500 лет назад. Которая в свою очередь восходит к единой послеледниковой предковой популяции, которая претерпела изменения из-за смены ландшафта, генетического дрейфа и эффекта основателя. В отличие от горных народов Дагестана, его равнинные популяции обладают большим разнообразием Y-хромосом, что можно объяснить их контактами с населением Центральной Азии. Дидойцы или цезы, представляющие аваро-андо-цезскую ветвь нахско-дагестанской языковой семьи, демонстрируют более скромное генетическое разнообразие из-за преобладания гаплогруппы J1 и значительного эффекта основателя.
Две работы этого года, опубликованные с интервалом в несколько месяцев, касаются аутосомного генофонда народов Восточного Кавказа и изменчивости Y-хромосомы в тех же популяциях. Такой систематический анализ генофондов популяций, населяющих исторически связанные территории восточной части Кавказа позволяет увидеть наиболее объемную и объективную картину изменчивости генофонда популяций этой физико-географической области. А результаты этих исследований могут быть использованы для моделирования расселения ранних людей по Кавказу, Западной Азии и Евразии, используя обновленные и расширенные данные ДНК.
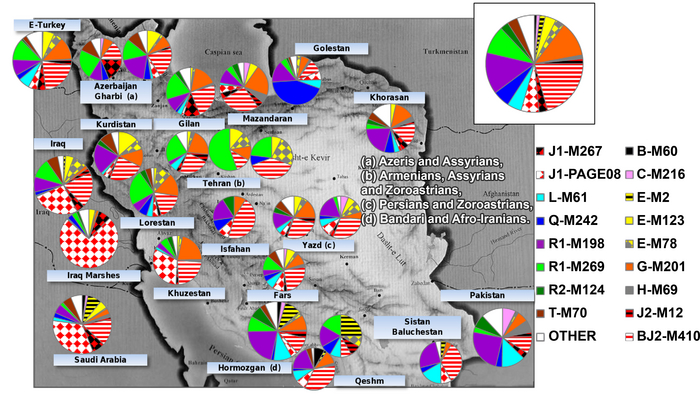
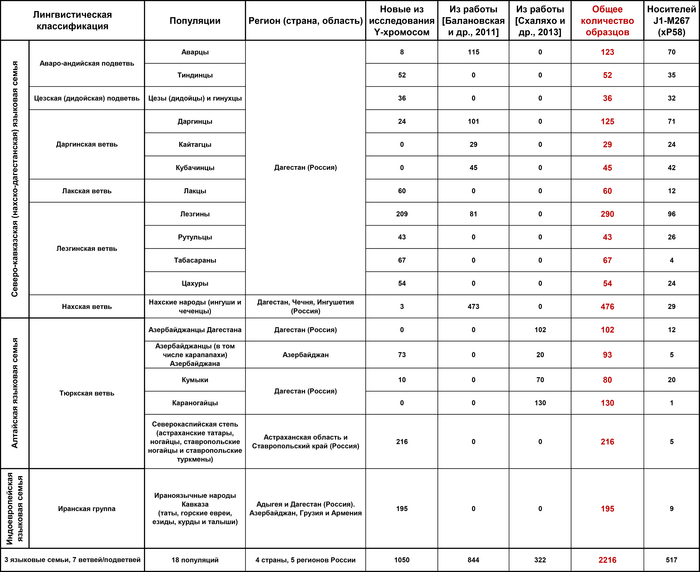
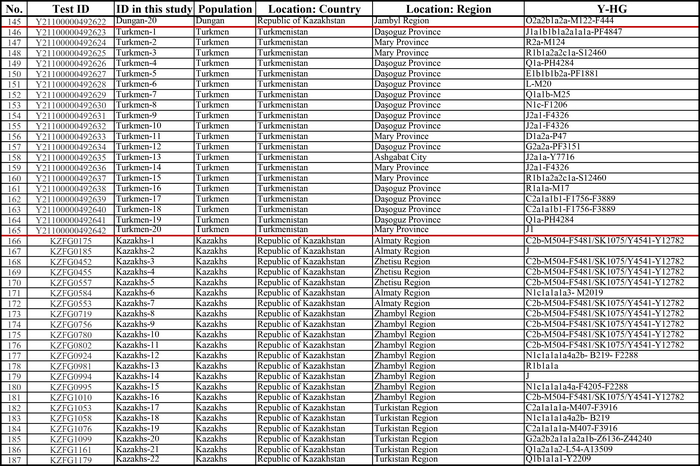
Всего в работах было проанализировано 2216 человек из 22 этнических групп Восточного Кавказа и сопредельных регионов, представляющих 3 языковые семьи: нахско-дагестанскую (аваро-андо-цезская, лакская, лезгинская и нахская ветви), алтайскую (тюркская ветвь) и индоевропейскую (иранская группа).
В исследованиях основное внимание уделялось Восточному Кавказу, генетические данные 1329 человек, представляющих коренные народы Дагестана и Азербайджана, были проанализированы более подробно, чем популяции степей Северного Каспия, носители нахских языков и ираноязычные народы Кавказа, которые вошли в группы сравнения и были представлены объединенными выборками без дальнейшего разделения на более мелкие подгруппы.
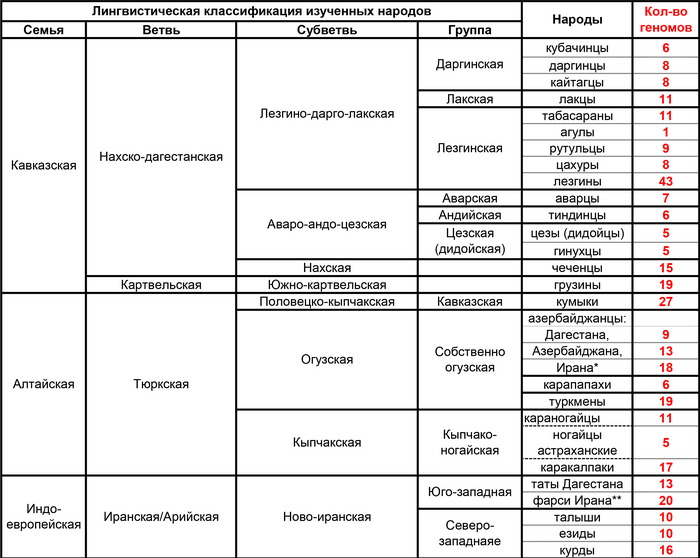
Нахско-дагестанская языковая семья была представлена 14 этническими группами: аварцами и тиндинцами (аваро-андийская языковая подветвь), цезами и гинухцами (цезская подветвь), даргинцами, кайтагцами и кубачинцами (даргинская ветвь), лакцами (лакская ветвь), лезгинами, рутулами, табасаранами и цахурами (лезгинская ветвь нахско-дагестанской семьи), а также чеченцами и ингушами (нахская ветвь). Тюркоязычные народы были представлены населением Восточного Кавказа (караногайцами, кумыками, терекеме Дагестана и азербайджанцами Азербайджана) и западной части Евразийской степи (ставропольскими ногайцами, астраханскими татарами, ногайцами и ставропольскими туркменами или трухменами). Ираноязычное население Кавказа было представлено татами, горскими евреями, езидами, курдами и талышами.
А в исследовании аутосомного генофонда 22 этнические группы Восточного Кавказа представлены данными о 243 геномах по широкогеномным панелям SNP-маркеров, в дополнение к 43 ранее опубликованным, которые были изучены в основном по узким панелям маркеров.
Более половины изученных образцов опубликованы в новых работах впервые.
Гаплогруппы Y-хромосомы в популяциях Восточного Кавказа и прилегающих регионов
Географическая изменчивость проанализированных гаплогрупп Y-хромосомы на Восточном Кавказе неодинакова для представителей разных языковых семей: она более выражена для нахско-дагестанских языковых групп, чем для тюркоязычного и ираноязычного населения.
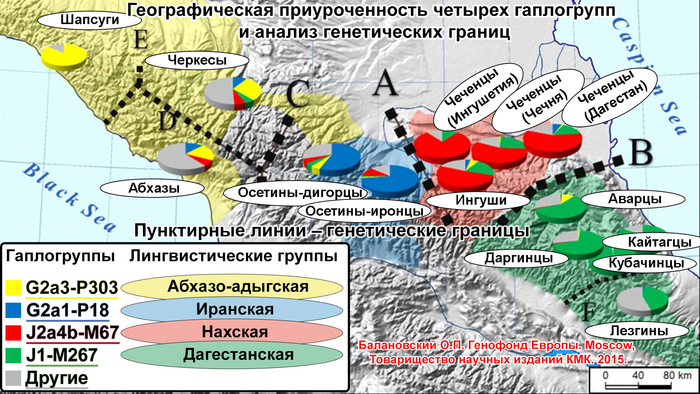
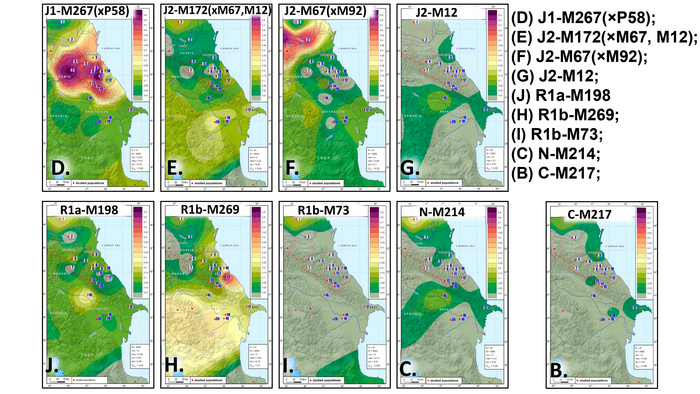
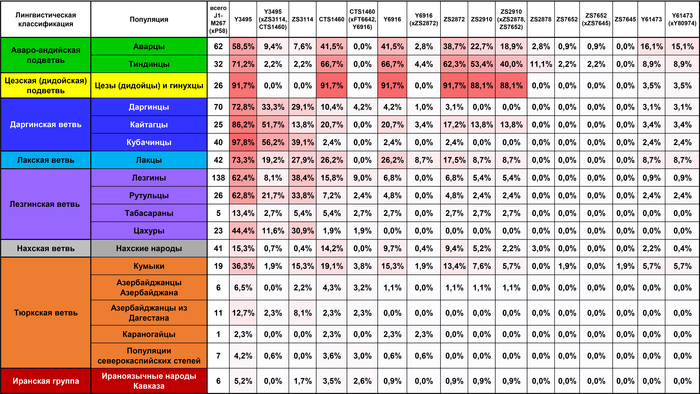
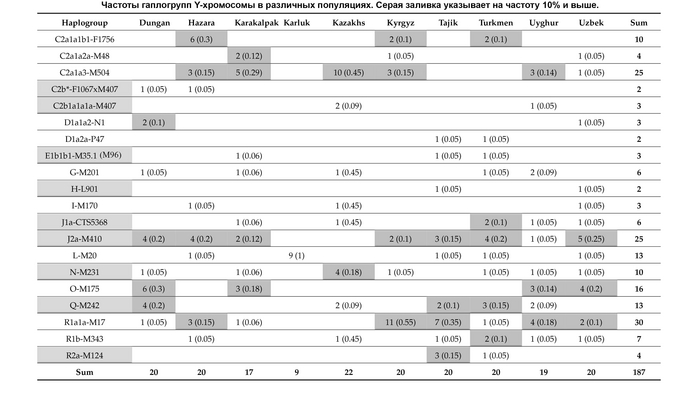
В целом, авторы выделили 5 основных гаплогрупп с различными географическими моделями распределения: J1, J2, R1a, R1b и N. Их разнообразие продемонстрировано на картах частотного распределения.
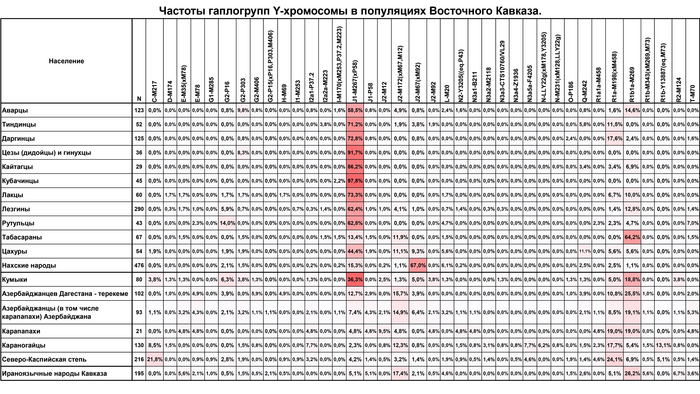
J1
Для гаплогруппы J1 характерна очаговая структура распространения, в Дагестане её частота снижается во всех направлениях от центра к периферии.
Возраст гаплогруппы J1 составляет от 9000 до 24000 лет и, как было показано в предыдущих исследованиях, она возникла в Западной Азии, где ее ветвь J1-P58 является наиболее распространенной и достигает пика частоты у арабов Ближнего Востока. Однако линия J1-P58 крайне редка для популяций Восточного Кавказа. Хотя в целом на долю линий гаплогруппы J1-M267(×P58) приходится более 50% генофонда дагестаноязычных популяций. За исключением цахуров (44%) и табасаран (13%). А максимума частота J1-M267(×P58) достигает у кубачинцев (98%), ареал обитания которых ограничен селом Кубачи. J1-M267(×P58) также преобладает и в других популяциях Дагестана (от 59 до 92%) и часто встречается среди чеченцев (17-25%). Однако её частота резко снижается по мере удаления от носителей нахско-дагестанских языков. При этом высокая частота гаплогруппы J1-M267(×P58), наблюдаемая у кумыков, может указывать на значительный генетический вклад нахско-дагестаноязычных популяций, поскольку эта гаплогруппа обычно не встречается в других тюрко- и ираноязычных популяциях Восточного Кавказа.
Высокая доля гаплогруппы J1-M267(×P58) в популяциях Восточного Кавказа может объясняться не только действием дрейфа генов, но и наличием иных – кроме J1-P58 - ветвей в данном регионе. Поскольку ранние носители гаплогруппы J1 появились на Кавказе еще в палеолите (~13 тыс. лет назад; охотники-собиратели на территории современной Грузии), а возникновение земледельческих сообществ в неолите привело к росту населения и появлению новых гаплогрупп, возможно, несколько местных линий возникли в пределах J1-M267(×P58). К этому вопросу ещё вернёмся.
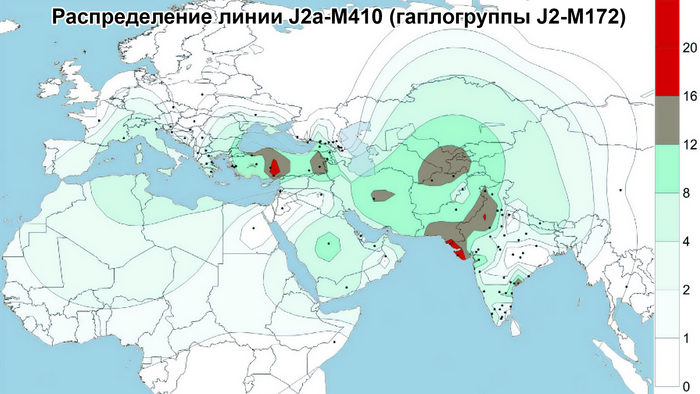
J2
Гаплогруппа J2-M172(×M67, M12) также имеет очаговую структуру, но ее пиковые частоты приходятся на юг региона: совокупный вклад и разнообразие ветвей J2 больше у азербайджанцев и ираноязычного населения Кавказа. Она редко встречается в популяциях Дагестана, но ее ветвь J2-M67(×M92) встречается с частотой 67% у их соседей — чеченцев и ингушей, носителей нахских языков. Происхождение гаплогруппы J2 прослеживается до Западной Азии, где наблюдается наибольшее разнообразие её ветвей (J2a-M92 и J2a-M530). Её частота достигает максимума в ираноязычных популяциях.
В Западной Азии основными носителями гаплогруппы J2 являются турки, азербайджанцы, курды и персы (20-35%). Наиболее распространенной является ветвь J2-M172 (×M67, M12) встречающаяся с частотой 15 и 20% среди тюркоязычных и ираноязычных популяций соответственно. А на Восточном Кавказе её частота составляет 11% у тюркоязычных и 17% у ираноязычных народов. Таким образом, частоты гаплогруппы J2 и ее ветви J2-M172(×M67, M12) среди азербайджанцев Дагестана - терекеме и азербайджанцев Азербайджана близки к частотам, наблюдаемым не только у ираноязычных народов Кавказа, но и у населения Западной Азии.
R1a и R1b
В целом, гаплогруппы R1a и R1b выявляют связь между степными популяциями на севере и группами, расселившимися вдоль Каспийского побережья на юге. Их совместный вклад наиболее значителен для тюркоязычных народов Дагестана (24-40%), азербайджанцев Азербайджана (30%) и ираноязычного населения Кавказа (37%). Их частота плавно снижается в нахско-дагестанских популяциях Дагестана, за исключением табасаран.
R1b
Гаплогруппа R1b представлена 2 субкладами: R1b-M269 и R1b-M73, из которых субклада M269 наиболее распространена. Её максимальная частота наблюдается у табасаран (64%), однако у большинства других носителей нахско-дагестанских языков не превышает 15%. R1b-M269 встречается у ираноязычных популяций Кавказа с частотой в 26%, азербайджанцев (23%) и кумыков (19%). Помимо этого, эта линия встречается с высокой частотой на юге региона среди азербайджанских талышей (48%).
В целом во всей Западной Азии вклад R1b значительно выше для тюркоязычного населения (20% для азербайджанцев и турок), чем для ираноязычного (5%). Схожие частоты R1b-M269 среди ираноязычного населения Кавказа и тюркоязычных народов Западной Азии могут указывать на генетические взаимодействия между их предковыми популяциями. R1b-M73 встречается с заметной частотой среди караногайцев (13%), популяций северокаспийских степей и тюркоязычных популяций Западного Кавказа (карачаевцев и балкарцев).
R1a
Гаплогруппа R1a представлена в основном ветвью R1a-M198(×M458), которая очень распространена среди карапапахов (19%) и караногайцев (18%), но встречается с меньшей частотой в других популяциях Восточного Кавказа, проанализированных в этой работе (2-11%), за исключением цезов, кубачинцев и табасаран. Гаплогруппа R1a широко распространена в Евразии: за исключением ветви R1a-M458, которая типична для популяций Восточной Европы и редко встречается в популяциях Восточного Кавказа. R1a включает в себя удивительное разнообразие линий, охватывающих обширную географию от Индии до Скандинавии.
N
Гаплогруппа N достигает самых высоких частот в степной популяции караногайцев на севере Дагестана (22%) и азербайджанских карапапахов (10%). Однако у них разные линии: у караногайцев субклады N3a5a-F4205 и N3a2-M2118, распространенные в Сибири и Центральной Азии, а также N-LLY22g (×M178, Y3205); а у карапапахов N2-Y3205 и N3a1-B211, которые являются типичными для популяций Уральского региона и Западной Сибири.
Основные закономерности географической изменчивости
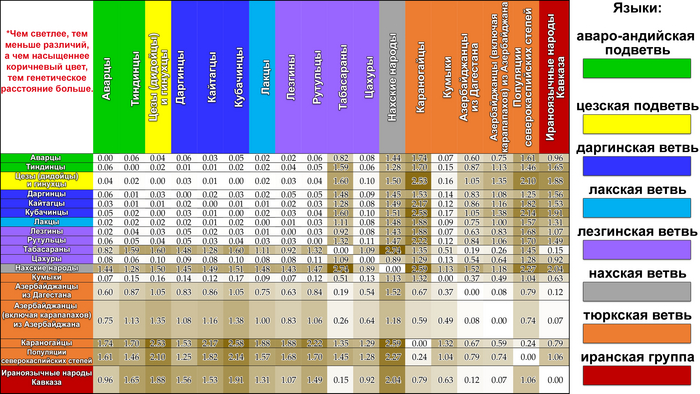
Карты генетических расстояний Нея, которые объединяют данные по всем гаплогруппам, показывают степень сходства и различия между изучаемыми популяциями, а также дают общую характеристику генофонда Y-хромосомы для каждой популяции и выделяют регионы с наибольшим генетическим сходством.
В целом генофонд Восточного Кавказа представлен тремя основными паттернами: дагестанским, иранским и степным.
Дагестанский паттерн
Карты генетических расстояний между говорящими на языках, входящих в нахско-дагестанскую языковую семью, имеют общую закономерность, единственным исключением является табасараны.
Этот дагестанский паттерн характеризуется близкими генетическими расстояниями между популяциями, встречается только в Дагестане и не коррелирует с языком. Похожая, но менее выраженная картина у тюркоязычных кумыков. Дагестанский паттерн показывает глубокую связь между генофондами популяций Дагестана с точки зрения гаплогрупп Y-хромосомы. Эта закономерность в значительной степени определяется присутствием гаплогруппы J1-M267(×P58). В рамках данного исследования наблюдается генетическое сходство между популяциями Дагестана и Северного Ирана, которое определяется вкладом гаплогрупп J1 и J2 и может отражать самые ранние волны миграции на Восточный Кавказ.
В целом дагестанский паттерн указывает на связь с древним автохтонным кавказским населением породившем множество генофондов благодаря мощному генетическому дрейфу, которые до сих пор сохраняют свое единство.
Иранский паттерн
Существует генетическое сходство между всеми азербайджанскими и ираноязычными популяциями Кавказа (татами, талышами, езидами, курдами). Вклад гаплогрупп J2-M172 (×M67, M12) и R1b-M269 в этот паттерн является наибольшим, что можно увидеть, сравнив карты их распределения и генетических расстояний. Эта закономерность проявляется на карте генетических расстояний от табасаран, которые представляют лезгинскую ветвь нахско-дагестанских языков, хотя для табасаран эта закономерность менее выражена.
Иранский паттерн связан с древним иранским населением, пришедшим из Мидии, и более ранними волнами миграции в 3-2 тысячелетиях до нашей эры.
Степной паттерн
Карты генетических расстояний также демонстрируют значительные различия между тюркоязычными популяциями. Так, генофонд кумыков близок к генофонду кавказоязычного населения Дагестана, а генофонд азербайджанцев схож с генофондом ираноязычных народов Кавказа. Единственная закономерность обнаружена у караногайцев и отражает весьма отчетливую связь с генофондами евразийской степи. На значительно более слабом уровне такая закономерность есть и у азербайджанских карапапахов. Степной паттерн отражает одну из последних миграционных волн тюркского населения и является более поздним с точки зрения своего происхождения и вклада в генофонд Восточного Кавказа.
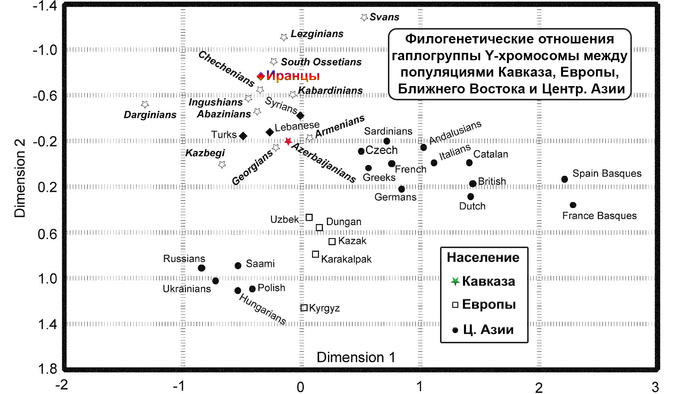
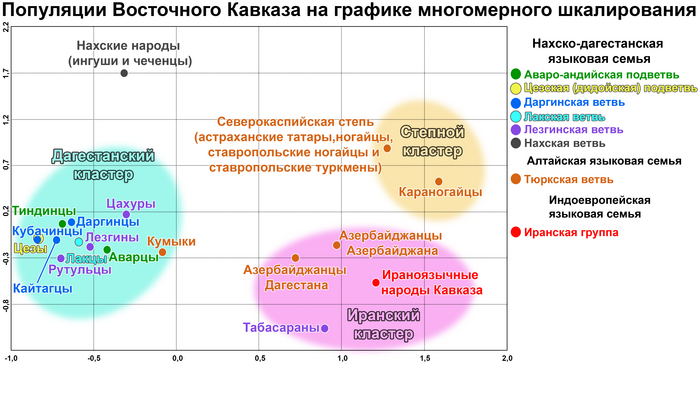
Восточный Кавказ в многомерном генетическом пространстве
Картографический анализ генетических расстояний выявляет на одной карте области генетического сходства одной популяции. В свою очередь, визуальное представление всей попарной матрицы генетических расстояний Нея посредством многомерного шкалирования показывает положение всех изученных популяций. На графике наблюдается три кластера — дагестанский, иранский и степной, которые отражают географические, а не лингвистические закономерности.
1/2
Северо-восточный вектор влияния Евразийской степи проявляется в степном кластере. Юго-восточный вектор отражает влияние Западной Азии и формирует произвольный иранский кластер. Вектор автохтонного кавказского населения проявляется в дагестанском кластере, объединяющем все дагестаноязычные народы. При этом между иранским и степным кластерами генетическое расстояние меньше, чем между иранским и дагестанским. А максимальное расстояние между дагестанским и степным кластерами. Аналогичные закономерности наблюдаются и при анализе главных компонент.
Степной кластер образован караногайцами Дагестана и объединенными популяциями тюркоязычных народов, населяющих север Прикаспийской степи (туркмены, ногайцы и астраханские татары). Кластер характеризуется наибольшей неоднородностью и отражает миграции внутри евразийских степей, в том числе средневековые миграционные волны, повлиявшие на генофонды ногайцев и астраханских татар, новейшую миграцию туркмен из Прикаспийского региона в XVII в. и др. Прошлые генетические взаимодействия между популяциями Восточного Кавказа и евразийских степей проявляются только на северной границе региона в генофонде дагестанских караногайцев и гораздо слабее у кумыков и карапапахов.
Иранский кластер включает представителей трех языковых семей: тюркоязычных азербайджанцев из Дагестана и Азербайджана, ираноязычного населения Кавказа и табасаран, говорящих на языке лезгинской группы нахско-дагестанской семьи. Этот кластер намного плотнее, чем степной, но менее объединенный, чем дагестанский. Присутствие табасаран в этом кластере может отражать наследие Табасаранского майсумства (государственного образования на юге Дагестана), где они жили рядом с другими популяциями кластера, или, альтернативно, может указывать на влияние средневековых миграций из провинции Табаристан на севере Ирана. Иранский кластер отражает взаимодействие между местным населением и населением Западной Азии и, следовательно, может быть назван иранским. Народы Восточного Кавказа и Западной Азии поддерживали свои контакты на протяжении длительных периодов истории; их контакты происходили на обширной территории, простирающейся за пределы Восточного Кавказа, поэтому необходимы дальнейшие исследования для изучения источников и истории этого компонента.
Дагестанский кластер включает в себя все кавказоязычные популяции Дагестана, за исключением табасаран, и тюркоязычных кумыков Дагестана, которые географически близки друг к другу. Распределение популяций внутри кластера отдаленно напоминает их географию: проживающие севернее даргинцы, лакцы и цезы, занимают условно “северо-западную” часть графика. Лезгинские популяции, расселенные дальше к югу, занимают “юго-восточную” часть графика, то есть противоположную часть скопления. Положение кумыков на границе кластера представляет собой основной пласт их генофонда, связывающий их с кавказоязычным населением и предполагающий генетические «заимствования» у других тюркоязычных популяций. Нахские народы (чеченцы и ингуши) расположились выше границ кластера.
Географически плотный дагестанский компонент генофонда Восточного Кавказа, встречающийся в большинстве дагестанских популяций, в значительной степени определяется высокой частотой гаплогруппы J1-M267 (×P58) поэтому рассмотрим её структуру более подробно.
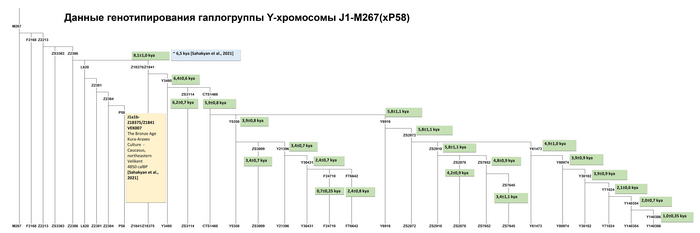
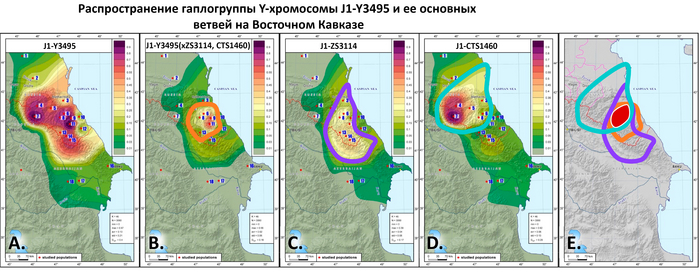
Филогеография гаплогруппы J1-M267(×P58)
Ветвь J1-Y3495, возрастом 6,4 ± 0,6 тыс. лет, является наиболее распространенной в регионе.
До сих пор субклада J1-Y3495 не была подробно описана в литературе, но ее предковая линия J1-Z18375 (или филогенетический эквивалент Z1841) хорошо известна и авторы сосредоточились на ней. Для J1-Z1841 время до ближайшего общего предка по разным расчётам составляет от 8 до 6,5 тыс. лет. Эта линия обнаружена у одного из трех представителей куро-араксской культуры (село Великент Дербентского района Дагестана), относящейся к эпохе бронзы (~5000 лет назад).
Это натолкнуло авторов работы на предположение о географическом происхождении J1-Z1841 на Кавказе или в его окрестностях. Её сублиния J1-Z1842 была обнаружена у человека на востоке Анатолии или Малой Азии, жившего ~5000 лет назад. Тот факт, что линия J1-Z1841/Z18375 была распространена в Малой Азии и Леванте около 3000 лет назад, предполагает контакты между носителями хуррито-урартских и нахско-дагестанских языков.
Единственной линией, представляющей гаплогруппу J1-Z1841 в популяциях Восточного Кавказа, является J1-Y3495. Авторам удалось идентифицировать 17 полиморфных линий этой гаплогруппы.
Карты показывают, что линии J1-ZS3114 и J1-CTS1460, а также J1-Y3495 (×ZS3114, CTS1460), возрастом от 6400 до 5900 лет, встречаются почти повсеместно в регионе: J1-ZS3114 более распространена среди носителей даргинской, лакской и лезгинской ветвей нахско-дагестанской семьи языков; J1-CTS1460 типична для носителей языков аваро-андо-цезской ветви той же семьи; J1-Y3495(×ZS3114, CTS1460) широко распространена среди носителей даргинской ветви и несколько менее часто встречается у носителей лезгинской ветви нахско-дагестанской семьи.
География этих трех линий пересекается с заметной частотой (более 20%) в Дагестане, что позволяет предположить, что линия J1-Y3495, возможно, появилась на его территории.
Небольшая разница, около 500 лет, в средних датах происхождения между гаплогруппой J1-Y3495 и двумя ее ветвями J1-ZS3114 и J1-CTS1460, указывает на рост населения в регионе в раннем бронзовом веке (~6 тыс. лет назад), во время появления новых методов обработки металла, экономических и социальных изменений, включающих социальное неравенство и племенные объединения, а также торговых отношений с населением Юго-Восточной Европы и Западной Азии.
Лингвистические данные свидетельствуют о том, что раскол нахско-дагестанского праязыка произошел в конце 3-го тысячелетия до н.э. (~5 тыс. лет назад), что согласуется с новыми генетическими данными: более древние линии, возрастом ~6000 лет, встречаются почти во всех дагестанских популяциях, проанализированных в этой работе, и у некоторых нахских народов, хотя пики частот для J1-CTS1460 и J1-ZS3114 наблюдаются в ряде различных языковых групп.
Таким образом, основная часть генофонда населения Дагестана по отцовской линии (носителей гаплогруппы J1-M267(×P58)) может быть прослежена до предковой линии J1-Y3495. Авторы предполагают, что в позднем медном веке и раннем бронзовом веке (~6,5 тыс. лет назад) носители этой линии расселились по центральной части горного Дагестана, где география основных субклад J1-Y3495 пересекается.
При этом носители этой линии были частью сообщества, говорившего на пра-нахско-дагестанском языке.
Похожий вывод делали и другие учёные в работах с более скромными техническими и методологическими возможностями. Наличие одной основной гаплогруппы в генофонде Дагестана может свидетельствовать о сокращении древней популяции в более ранние периоды, когда другие линии, сосуществовавшие с J1-Y3495, вымерли из-за эффекта бутылочного горлышка. Т.е. могло иметь место сокращение численности популяции с последующим её восстановлением, но с меньшим генетическим разнообразием.
Эффект бутылочного горлышка с последующим ростом популяции, но с меньшим генетическим разнообразием.
Географические особенности распространения наиболее часто встречающихся вариантов линии J1-CTS1460 (Y6916 (×ZS2872), Y61473, ZS2910 (×ZS2878, ZS7652) и ZS2878) показывает, что ее географический центр находится в районах, занятых людьми, говорящими на языках аваро-андо-цезской ветви нахско-дагестанских языков, и распространяется на некоторые районы южнее. Датировка линий около 4-5 тыс. лет назад предполагает непрерывный рост популяций после их разделения.
В первой части обзора, мы рассмотрели отцовские линии, однако не только мужчины участвуют в формировании всего генофонда популяций, как уже отмечалось ранее, не все линии гаплогрупп Y-хромосомы дошли до наших дней, однако в аутосомном генофонде остаются следы всех его родоначальников в той или иной мере. Поэтому во второй части рассмотрим общегеномные данные в этих же популяциях Восточного Кавказа.
Источник:
Agdzhoyan, A.; Iskandarov, N.; Ponomarev, G.; Pylev, V.; Koshel, S.; Salaev, V.; Pocheshkhova, E.; Kagazezheva, Z.; Balanovska, E. Origins of East Caucasus Gene Pool: Contributions of Autochthonous Bronze Age Populations and Migrations from West Asia Estimated from Y-Chromosome Data. Genes 2023, 14, 1780. https://doi.org/10.3390/genes14091780
Центральная Азия расположена на пути расселения людей современной анатомии с Ближнего Востока в Северную Евразию, что отражено в геномах древних людей, таких как усть-ишимский человек, обитатели Янской стоянки на севере Якутии и стоянки Мальта в Иркутская область. При этом климатические изменения, связанные с последним ледниковым максимумом (LGM) существенно изменили распределение популяций древних людей в Центральной Азии и прилегающих регионах. Древние носители ботайской культуры благодаря предковым линиям древних северных евразийцев генетически связаны с представителями стоянок Мальта в районе озера Байкал и Афонтова гора у современного Красноярска. Последующие миграции земледельцев с Ближнего Востока и скотоводов из степи, дополнили разнообразие в регионе. Распространение на восток в Центральную Азию и восточно-евразийские степи индоевропейских популяций привело к возникновению энеолита и бронзового века. Известны довольно примечательные Бактрийско-Маргианский археологический комплекс и андроновская культурно-историческая общность, предковая, как считается, ираноязычным популяциям.
Карасукская культура возникла в Южной Сибири и распространилась по обширным территориям, дав начало ряду преемников. Позднее подъём скифов вызвал широкомасштабные миграции и примесь через евразийские степи, включая Центральную Азию.
В более поздний исторический период тенденция к расселению людей с запада на восток изменилась, юэчжи, усуни и эфталиты мигрировали с востока на запад и создали ряд государств в Центральной Азии. А хунну/гунны массово мигрировали на запад и значительно изменили этническую структуру в Центральной Азии и Восточной Европе. Позже древние тюркские и монгольские племена распространились на запад и рассеялись по всей Центральной Азии, Ближнему Востоку и Восточной Европе. При этом потомки вышеупомянутых и не упомянутых, в сильно сокращенной предыстории, древних людей в течение длительного исторического периода смешивались и формировали современные популяции в Центральной Азии.
Предыдущие генетические исследования современных людей пролили свет на происхождение и генетическую структуру популяций из Центральной Азии. А анализ древней ДНК останков различных исторических периодов выявил закономерности смешения и генетическую структуру древних людей, а также их вклад в генофонды современных популяций.
В целом отцовский, материнский и аутосомный генофонды большинства популяций этого региона характеризуются высоким разнообразием и примесью с популяциями Ближнего Востока, Европы, а также Южной, Восточной и Северной Азии. Также у популяций этого региона выявлена сильная племенная структура, которая привела к значительным различиям в доминирующих отцовских линиях между племенами. Однако внутри племён разнообразие коротких тандемных повторов на отцовских Y-хромосомах (Y-STR), как правило, невелико.
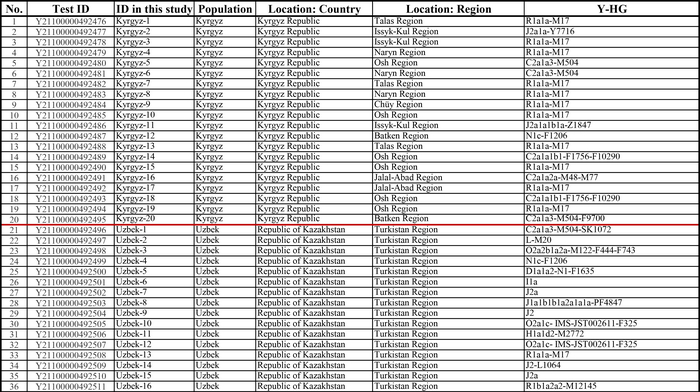
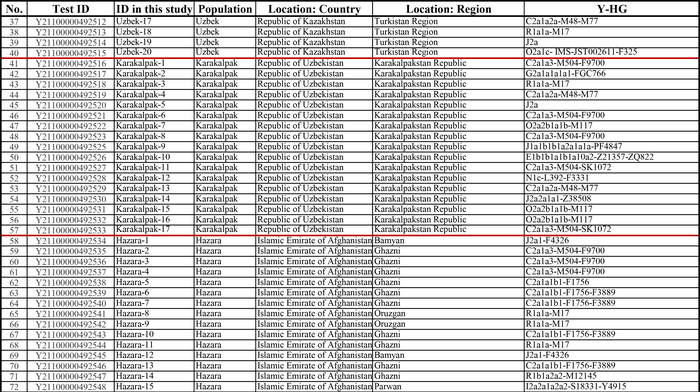
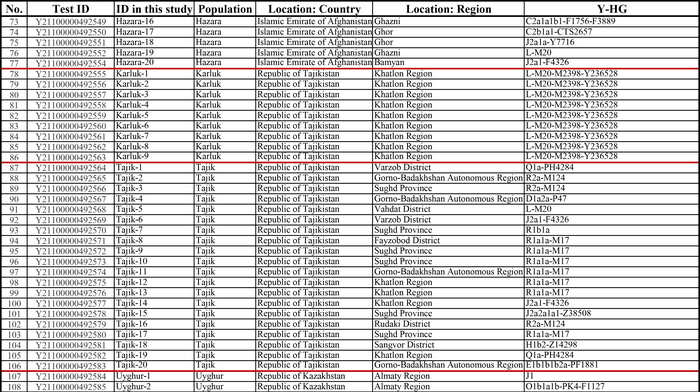
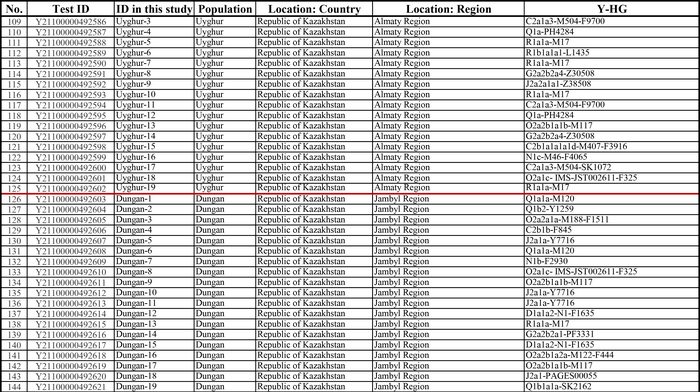
И хотя поток генов является обычным явлением, генетические взаимоотношения между популяциями по-прежнему тесно связаны с языком. Например, таджики генетически родственны другим индоевропейским популяциям, в то время как у кыргызов обнаружено много генетических компонентов из Сибири. Кроме того, были оценены последовательности Y-хромосом конкретных племен или семейных кланов для изучения формирования родственных популяций. Тем не менее, демографическая история населения Центральной Азии сложна, поэтому любые новые данные дополняют и уточняют наши знания об истории региона. Так и в новой работе авторы дополняют данные полноразмерными последовательностями Y-хромосом 187 человек. В выборку вошли казахи, туркмены, узбеки, таджики, кыргызы, каракалпаки, хазарейцы, карлуки, уйгуры и дунгане.
Основной целью авторов работы было выявление на основе полноразмерных последовательностей Y-хромосом общих закономерностей в отцовских генофондах. После чего идентифицировать отцовские линии, которые почти уникальны для какой-то конкретной популяции, и проанализировать их демографическую историю. В частности, исследователи оценили различные периоды генетического обмена и расхождения среди древних популяций в Центральной Азии.
Результаты
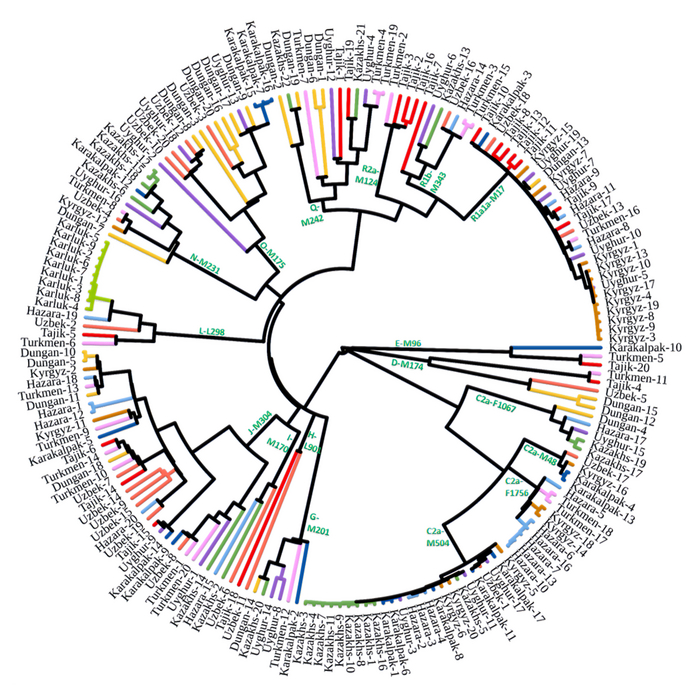
Пересмотренное филогенетическое древо, основанное на Y-хромосомах популяций Центральной Азии
Гаплогруппа C2a1a1b1-F1756 была преобладающей отцовской линией древних племен дунху и сяньби, а появление этой линии у хазарейцев, кыргызов и туркмен указывает на то, что их предки по отцовской линии из Восточной Евразии. Гаплогруппа C2a1a2a-M48-F6379 имела высокую частоту в Младшем жузе казахов и ойратов — западных монголоязычных народов. Появление этой линии у каракалпаков свидетельствовало о близком родстве этого населения с казахами и древними монголами. Считается, что экспансия в Центральной Евразии С2а1а3- M504 напрямую связана с деятельностью древних монгольских племен в прошлом тысячелетии. В исследовании отмечена высокая частота этой линии в популяциях хазарейцев, каракалпаков, казахов, кыргызов и уйгуров, что согласуется с предполагаемым монгольским происхождением этих популяций из предыдущих исследований разных авторов. Линии N-M231 у казахов связаны с древнетюркскими племенами. А линии D-M174 и O-M175, обнаруженные в популяциях Центральной Азии, появились в результате смешения за последние 2000 лет с популяциями из Восточной Азии.
Как уже известно, гаплогруппы E-M96, G-M201, H-L901, I-M170 и L-M20 были широко распространены с низкой частотой в популяциях из Центральной Азии. Эти линии представляют собой смесь из Южной Азии, Ближнего Востока и Европы. Тем не менее, исследователи обнаружили особую сублинию L-M20 у 9 карлуков, а именно L1a2a-M2398-Y236528, которая около 1500 лет назад отделилась от линий, обнаруженных в Индии.
Предыдущие исследования показали, что Южная Сибирь, вероятно, является центром распространения гаплогруппы Q-M242, начавшейся около 30 тысяч лет назад. Кроме того, многие второстепенные сублинии Q-M242 были обнаружены в популяциях из Внутренней Евразии, включая монголо- и тюркоязычные популяции. В новой работе авторы также обнаружили различные сублинии Q-M242.
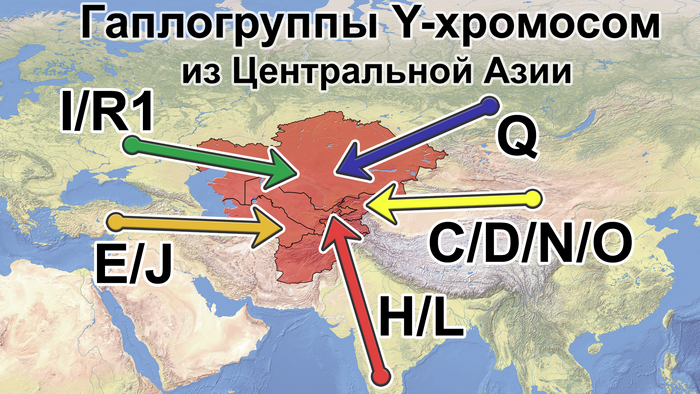
Также известно, что R1a1a-M17 и R1b-M343 являются двумя основными отцовскими линиями индоевропейского населения и большинство современных популяций в Центральной Азии по-прежнему демонстрируют высокую частоту R1a1a-M17. Пересмотренное филогенетическое дерево высокого разрешения с оценкой возраста дает подробный обзор вклада по отцовским линиям в популяциях Центральной Азии. Высокое разнообразие линий макрогаплогрупп предполагает интенсивную примесь из разных периферийных областей Евразийского континента, так линии C/D/N/O из Восточной Евразии, E/J с Ближнего Востока, H/L из Южной Азии, I/R1 из Западной Евразии и Q из Южной Сибири.
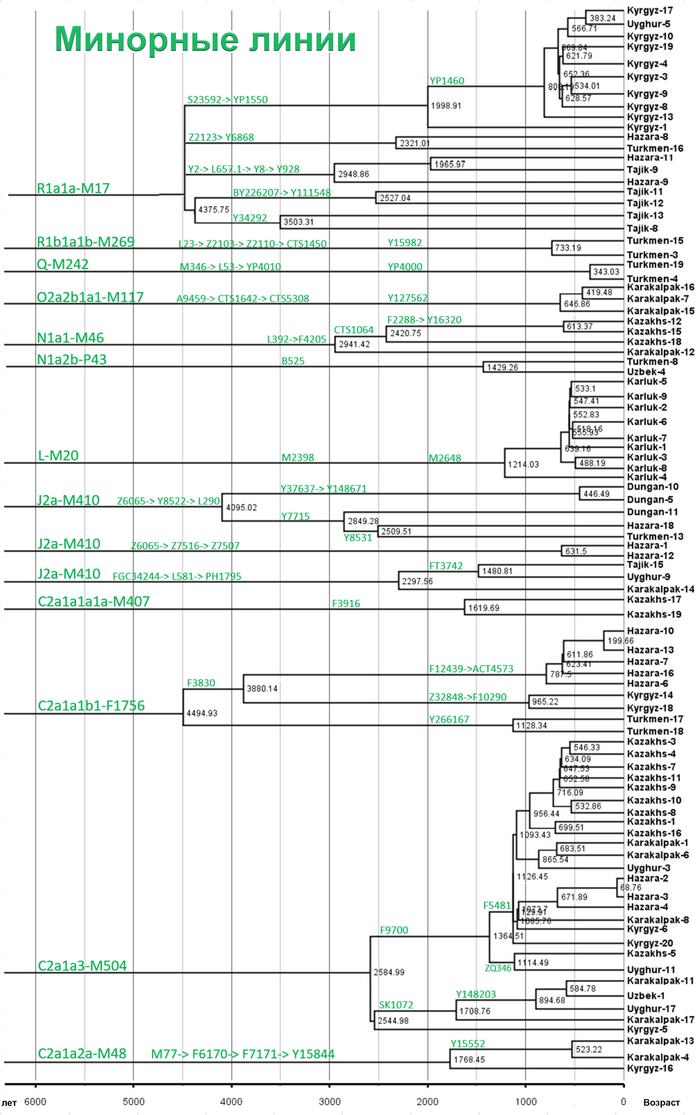
Однако наиболее важным открытием этой работы является идентификация уникальных редких, так называемых, минорных линий, которые в Центральной Азии распространились недавно. В том числе 33 такие линии моложе 2000 лет.
Выводы
Множественные события миграций и различные линии гаплогрупп
Центральная Азия не является регионом зарождения земледелия эпохи неолита или технологических систем эпохи бронзы. Поэтому роста населения, вызванного этими технологическими революциями, в Центральной Азии не было. Однако генетические данные показывают, что источниками популяций для Центральной Азии были различные регионы Евразии, включая Южную Азию, Ближний Восток, Европу, Северную и Восточную Азию. Этот общий сценарий согласуется с географическим положением Центральной Азии как центра Евразии. Причём эти миграции происходили постепенно в разные периоды. Так, неолитическая революция на Ближнем Востоке привела к глобальному распространению гаплогруппы J2-M172 и её дочерних линий, параллельно с появлением первых неолитических поселений в южной части Центральной Азии.
А позже непрерывная миграция с Ближнего Востока привела к появлению дополнительных сублиний J2-M172.
Многие исследования древней ДНК показали, что распространение отцовских линий R1a1a-M17 и R1b-M269 согласуется с экспансией индоевропейских популяций, и это может быть связано с разнообразными сублиниями R1a1a-M17 в Центральной Азии. Кроме того, гаплогруппа G-M201, вероятно, также происходит от индоевропейцев. Однако основываясь на ранее опубликованных генетических данных древних и современных популяций, авторы предполагают, что наличие в Центральной Азии некоторых линий Q-M242, N1a1-M46, R1a1a-M17 и C2a1b1-F1756 может быть связано с миграцией тюркских народов. А монголоязычные популяции привнесли в Центральную Азию линии C2a1a3-M504, отдельные C2a1b1-F1756 и сублинии C2a1a2a-M48. Во время миграции с Востока на Запад, на протяжении сотен лет, многие отцовские гаплогруппы распространились из Восточной Азии в Центральную Азию, включая несколько сублиний D-M174, C2a-F1096 и O-M175. В целом из-за высокой скорости мутаций Y-хромосома легко образует специфические для популяции сублинии или субклады, как кому удобнее.
Однако в комментариях люди часто неверно трактуют предковые гаплогруппы опираясь только на те популяции, у которых встречаются их сублинии. Не говоря про тех, которые им приписывают какие-то невероятные свойства.
Недавние миграции и формирование современного населения
Авторы работы предполагают, что недавнее появление редких минорных предковых линий напрямую способствовало формированию современных популяций Центральной Азии.
Основные отцовские гаплогруппы популяций в разных регионах Евразии включают C-M217, D-M174, E-M96, G-M201, H-L901, I-M170, J-M304, L-L298, T-M184, N-M231, O-M175, Q-M242, и R-M207. Эти макрогаплогруппы распространены и в популяциях Центральной Азии, однако их частоты существенно различаются. На уровне сублиний население Центральной Азии имеет определенные доминирующие линии.
Но как показано на графике, среди населения Центральной Азии было и много мелких минорных предковых линий характерных для некоторых этнических групп, как к примеру, R1a-M17-Y34292 и R1a-M17-Y111548 у таджиков. Наблюдается множество минорных предковых линий, которые расширились за последние 1000 лет, таких как C2a1a3-M504-F5481, L-M20-M2648 и R1a1a-M17-YP1460. Многие такие линии от R1a1a-M17 у таджиков связаны с их индоевропейскими предками. А в целом минорные линии наблюдались у казахов, кыргызов, карлуков, каракалпаков и хазарейцев.
Авторы считают, что расширение этих специфических линий напрямую связано с формированием современных этнических групп в Центральной Азии.
Таким образом, построено филогенетическое дерево линий Y-хромосом с высоким разрешением для популяций из Центральной Азии. Где значительное разнообразие отцовских линий, указывает на примесь из различных регионов Евразии в разные периоды времени. А множество минорных предковых линий указывает на недавние события примеси, которые могут соответствовать формированию современных популяций Центральной Азии за последнюю тысячу лет.
Источники: Maxat Zhabagin, Lan-HaiWei, Zhaxylyk Sabitov, Peng-Cheng Ma , Jin Sun, Zhanargul Dyussenova, Elena Balanovska, Hui Li and Yerlan Ramankulov. Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosome from Central Asia // Genes. 2022, 13, 1776. doi.org/10.3390/genes13101776
В древности был военачальник и легендарный воин, который завоевал Индию. Предания об этом воине сохранились в русских былинах, греческих мифах и индийских сказаниях.
Этот воин родился от женщины и змея, либо одного из богов, принявшего вид змея. При его рождении был сильный гром и бегство животных. Этот воин был оборотнем - мог превращаться в животных и даже насекомых. Он превращал в насекомых свое войско, чтобы пробраться в крепость врага. Этот воин любил выпить.
Остается неизвестным, когда именно он жил.
Имя этого воина на русском языке Волх Всеславьевич, у греков он Вакх, у индийцев он Индра. Приведенные выше его способности и жизнеописание соответствуют каждой этой личности.
Как могли быть связаны русские люди, население Греции и население Индии?
Конечно, после веков изучения искаженной истории, доказательства могут быть косвенными.
Кроме сходства в истории этих народов, можно вспомнить и другие факты.
В "Сказании о Словене и Русе", входящем в «Повесть о стране Вятской» (Вятский летописец): памятник Вятской письменности XVII–XVIII века», изданной в 1905 году, праправнуки Ноя - Скиф и Зардан основали страну Скифию. Их потомки Словен, Рус, Болгар, Коман и Истер из-за вражды разошлись и стали завоевывать новые земли.
Есть версия, что скифы - прямые предки русских людей, а славяне - покоренные скифами народы Восточной Европы, воспринявшие язык ("слово") завоевателей. Считается. что Скифия занимала огромную территорию Украины, Юга России и Казахстан. Империя простиралась до Средиземного моря. Так что скифы вполне могли заселить даже Грецию и оставить там - родственное нам - русское население. На древних картах Северный океан именуется Скифским морем.
Карта Тартарии Абрахама Ортелиуса 1570 г.
Да, скифы - мы! Да, азиаты - мы,
С раскосыми и жадными очами!
А.А. Блок.
Славяне населяли Европу до разных ее окраин. Доримское население Италии называлось этруски. Их самоназвание расены. Догреческое население Греции называлось пеласги и лелеги. На украинском языке журавлей и сейчас называют "лелеки".
Между русским языком и древним языком Индии - санскритом есть тысячи сходств в словах. Наиболее заметно это по географическим названия - они не меняются веками. Индийские предания, говорят, что предки их вождей и брахманов пришли с Севера.
Современные научные открытия профессора Анатолия Клесова в генетике подтверждают это. После исследований гаплотипов выяснилось, что примерно 4,8 тысяч лет назад русские люди с генетическим маркером R1a появились на Русской равнине. Это произошло на 1,2 тысячи лет раньше исхода евреев из Египта и на 200 лет раньше извержения вулкана Санторин (Тера), уничтожившего минойскую цивилизацию на острове Крит. Через тысячу лет наши предки уже жили на Урале, построили город Аркаим, через 200 лет его покинули (подтверждается археологами) и пришли в Индию. Еще этих людей называют арии.
Таким образом, завоевание столицы Индии русским богатырем Волхом Всеславьевичем, именовавшимся среди местных как Индра, произошло 3,6 тысяч лет назад.
История наших народов прекрасна и потрясающая тем более, чем более задумываешься о ней.
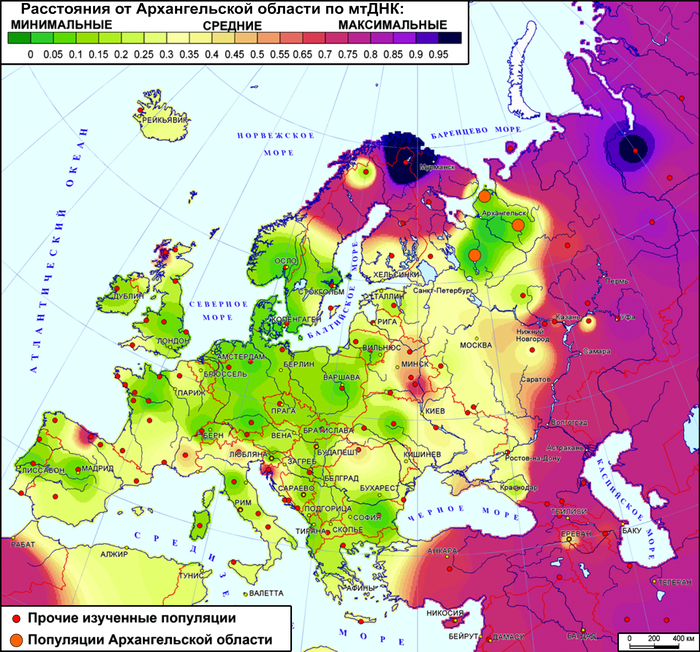
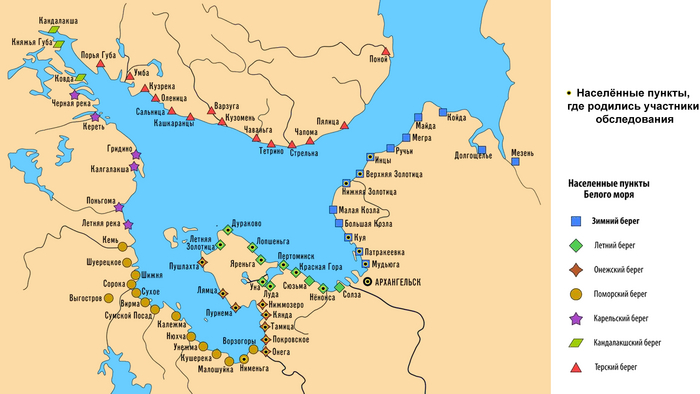
Сегодня речь пойдёт о поморах – представителях самых северных русских популяций Беломорья. Считается, что это потомки новгородцев, расселявшихся с XII в по берегу Белого моря от Онеги до Кеми, а также по берегу Баренцева моря. Поморы адаптировали свой быт и культуру к суровым природным условиям северных морей, занимаясь рыболовством, морской охотой, предпринимательством и мореходством на Белом море, в Северном Ледовитом океане и на его островах, конкурируя с норвежцами.
И хотя поморы отличались по быту от северных русских, они всё же сохранили черты древнерусской культуры, при этом взаимодействуя с населением Северной и Северо-Восточной Европы. Поэтому население этого региона крайне своеобразно, что и заинтересовало учёных наряду с нюансами эволюции популяций на самой периферии этнического ареала.
Заселение Беломорья началось около 8 тысяч лет назад ещё в мезолите. В неолите свои следы оставили представители «каргопольской» и «беломорской» культур.
А в бронзовом веке, около 3-4 тыс. лет назад, в регион прибыли мигранты, которых связывают с финно-угорскими популяциями (прежде всего, с группами, говорившими на языках, близких к саамским). Последней была славянская волна мигрантов. И первые упоминания о постоянных русских поселениях на беломорском побережье в конце XIII — начале XIV века связаны с Новгородской колонизацией северных земель. Где, согласно летописям, переселенцы встретили племена "заволочской чуди", как в летописях называли финноязычное население, часто ассоциирующееся с современными вепсами. Проживание на самом севере ареала русского народа, где в топонимике прочно сохранились следы дославянского населения, совместно с уникальной культурой и бытом, предполагает также и разнообразие генофонда поморов.
В предыдущих работах — по крайне узким панелям маркеров Y-хромосомы и мтДНК с небольшими выборками — поморы рассматривались в составе популяции «северных русских», а особенности их генофонда объяснены возможной ассимиляцией урало- или балтоязычного населения славянским. Данные об аутосомном геноме поморов Мезенского района Архангельской области также указывают на сходство с финноязычным населением, причем больше с финно-пермским, чем с финно-волжским.
На рисунке: данные об аутосомном геноме поморов Мезенского района Архангельской области
Однако в исследованиях не поморов, а обширного круга популяций Русского Севера, выявлена их близость по мтДНК и аутосомным ДНК-маркерам к населению Северной Европы.
А разнообразие линий Y-хромосомы обнаружило сходство генофонда жителей Русского Севера с населением от Прибалтики до Печоры и объяснялось наследием древнейших палеоевропейских популяций.
Интригу к вопросу о происхождении поморов добавляет и тот факт, что у жителей самого севера Архангельской области не обнаружена «новгородская» предковая компонента по широкогеномной панели аутосомного генома.
В новой работе детально изучены наиболее сохранившиеся и труднодоступные популяции поморов Белого моря. В ходе экспедиции 2021 г. обследованы практически все населенные пункты Онежского полуострова (Онежский и Летний берега), а также западного (Прионежского) фрагмента Зимнего берега. И только для краткости все три популяции названы «онежскими поморами».
Авторы надеялись, что столь трудно добытые новые данные о генофонде поморов, изученном по современным панелям маркеров, смогут дать ключ к пониманию значительного генетического разнообразия и своеобразия популяций Русского Севера. Принимая во внимание традицию патрилокальности у поморов, авторы работы изучили маркеры Y-хромосомы для того, чтобы создать «генетические портреты» трёх впервые исследованных популяций поморов и попробовать найти в их генофонде генетическую память о новгородской колонизации.
Результаты
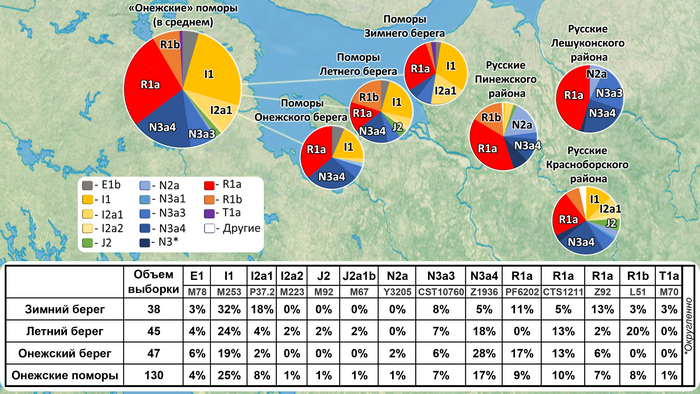
Разнообразие гаплогрупп Y-хромосомы у поморов
У онежских поморов самыми частыми оказались три гаплогруппы: 26% R1a (три ветви); 25% I1 (одна ветвь), 24% N3a (две ветви). А всего у исследованных мужчин было выявлено 14 линий Y-хромосомы: I1-M253, N3a4-Z1936, R1a-CTS1211, R1a-PF6202, R1b-L51, I2-P37.2, R1a-Z92, N3a3-CST10760, E-M78 и T1a-M70, встречающиеся с частотами от 25 до 1% (перечислены по убыванию) и линии I2-M223, J2-M92, J2-M67, N2a-Y3205 с частотами менее 1%.
При этом, несмотря на небольшое расстояние между тремя популяциями поморов (от 80 до 170 км,) заметно отличаются частоты их гаплогрупп:
– у поморов Зимнего берега снижена частота гаплогруппы N3a4 и повышена частота I2;
– у жителей Онежского берега наоборот, снижена частота гаплогруппы I2 и повышена частота N3a4.
– а у находящихся между ними представителей Летнего берега не обнаружена гаплогруппа R1a-PF6202, характерная для двух других популяций, но повышена частота гаплогруппы R1b.
Примечательно, что у поморов Летнего берега встречается линия R1b-L51, более характерная для народов Северо-Западной Европы, в отличие от другой линий R1b, обнаруженной у русских из Пинежского района Архангельской области.
В зависимости от географического положения популяций поморов наблюдается снижение частот гаплогрупп N3a4 и E1 с юго-запада на северо-восток, а частоты гаплогрупп I1 и I2a наоборот возрастают в этом направлении. Однако этой тенденции нет у ветвей гаплогруппы R1a, частота которой высока в генофондах Зимнего (29%) и Онежского берегов (36%), но в два раза ниже на расположенном между ними Летнем берегу (причем только ветви R1a-CTS1211 (13%) и R1a-Z92 (2%)). Зато на Летнем берегу наблюдается аномально высокая частота R1b-L51 (20%), что может быть результатом миграций и дрейфа генов.
Сходство онежских поморов с другими русскими популяциями Архангельской области наблюдается по частотам гаплогрупп R1a и N3a4 (кроме Зимнего берега, где N3a4 редка). А различия наиболее выражены по частоте гаплогруппы I1: у онежских поморов она составляет в среднем четверть генофонда (25%), но для других северных русских не характерна (в Красноборском районе Архангельской области – 12%, в Пинежском – 1%, а в Лешуконском не обнаружена вовсе).
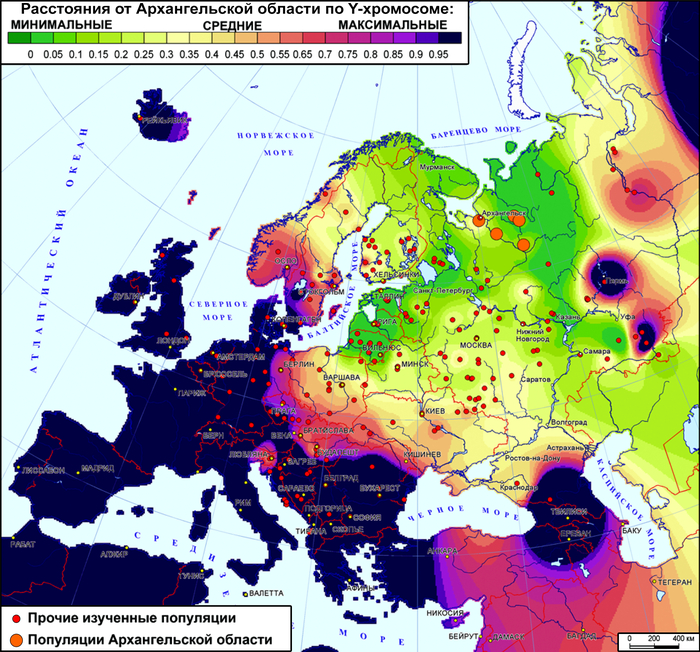
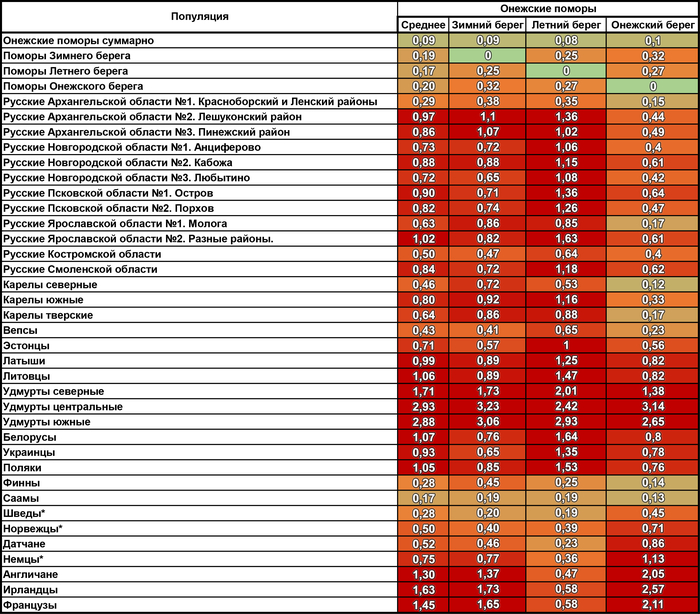
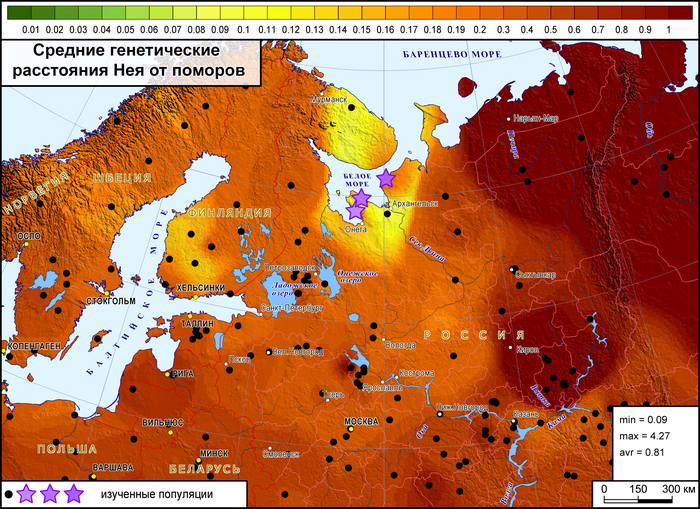
Генетические расстояния между поморами и другими популяциями Северной Европы
При оценке сходства разных генофондов суммарно по всем гаплогруппам наиболее примечательны значительные генетические расстояния между тремя популяциями онежских поморов, вопреки их географической близости.
Но ещё более значительны расстояния от поморов до других русских популяций. Есть лишь два исключения. Во-первых, сходство поморов Онежского берега с географически далекими русскими Красноборского района Архангельской области. Во-вторых, их сходство с русскими из района Мологи Ярославской области. Причём ранее было показано, что среди ярославских популяций именно представители Мологи сохранили явный генетический след дославянского населения (предположительно мери). Однако поморы Онежского берега ещё ближе к северным карелам, а также финнам и саамам, чем к этим русским популяциям. Поморы Летнего берега также близки к саамам, но ещё они близки и к шведам с датчанами, причём до них генетические расстояния меньше, чем до коллег по быту. Похожая картина у поморов Зимнего берега, только они более близки к саамам и шведам по генетическим расстояниям.
В целом онежские поморы демонстрируют наибольшее генетическое сходство с народами Северной Европы.
Причём расстояние до генофондов финнов и саамов уменьшается по мере движения на юго-запад - от Зимнего берега к Онежскому в 4 раза, а генетическое расстояние до скандинавов уменьшается к Летнему берегу от Зимнего и Онежского берегов. При этом поморы Летнего берега помимо близости к датчанам, норвежцам и шведам, также близки и к немцам с англичанами, хоть и не настолько, как к скандинавам. Также представители Летнего берега генетически ближе к ирландцам и французам, чем к соседним русским популяциям.
Среди других народов России к онежским поморам наиболее генетически близки вепсы и северные карелы, причем в основном за счет поморов Онежского берега, которые сами ближе к финноязычным соседям, чем к другим популяциям поморов.
А наиболее генетически удалёнными от поморов оказались удмурты – представители восточного крыла финноязычных народов, что входит в противоречие с выводами предыдущих публикаций о сходстве геномов финно-пермских народов с поморами.
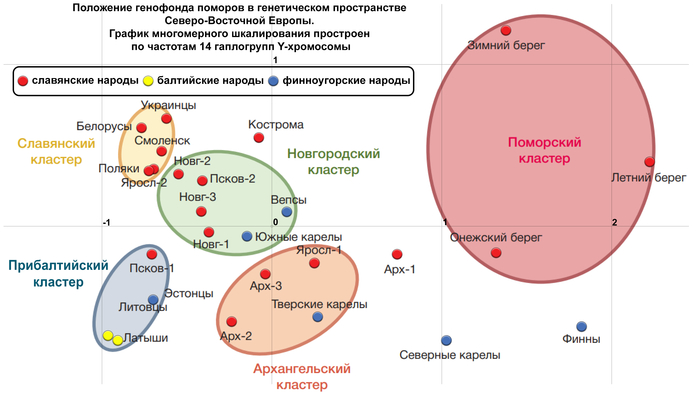
Генетическое положение кластеров популяций
На графике многомерного шкалирования (построенном по 14 «поморским» гаплогруппам Y-хромосомы) выделяются 5 кластеров, с условными названиями:
1. «Славянский» кластер, в него вошли белорусы, украинцы, поляки, а также русские Смоленской и Ярославской областей.
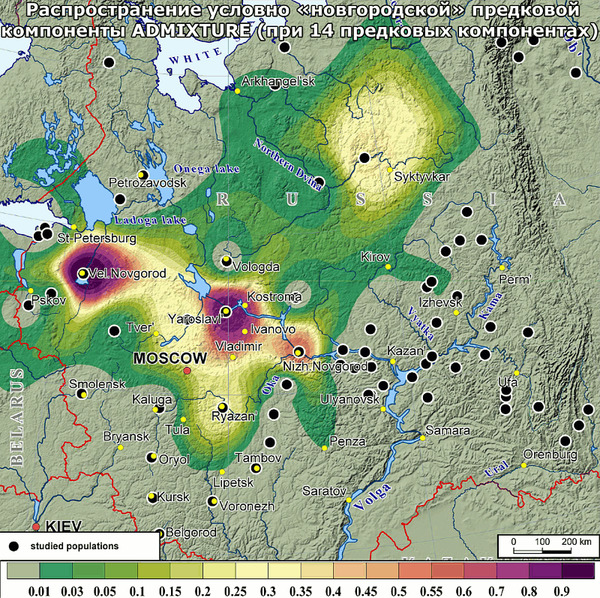
2. «Новгородский» кластер, близкий к «Славянскому», объединил все три популяции Новгородской области и Порхов Псковской области, который входил в Новгородские земли. К ним присоединились также финноязычные популяции – вепсы и южные карелы.
3. «Прибалтийский» кластер, помимо латышей, литовцев и эстонцев, включил население Островского района Псковской области.
4. «Архангельский» кластер объединил популяции Пинежского и Лешуконского районов Архангельской области с Мологой Ярославской области и тверскими карелами.
5. «Поморский» кластер, из-за значительных расстояний между популяциями поморов, оказался лишь немногим меньше суммы расстояний предыдущих четырех кластеров, включивших финноязычные, балтоязычные и славянские популяции.
При этом авторы подчёркивают, что вопреки значительным различиям между поморскими популяциями, все вместе они занимают своё собственное «поморское» место среди популяций Северо-Восточной Европы.
Карты генетических расстояний
Более точно определить регионы, с которыми сходны генофонды поморов, и значительно расширить круг популяций сравнения помогают карты генетических расстояний по методу Масатоши Нея, рассчитанные по 26 гаплогруппам Y-хромосомы, характерным для всего региона.
Суммарный генофонд поморов помимо некоторых соседних популяций генетически более близок к южной части Финляндии, а не к северной, представленной саамами.
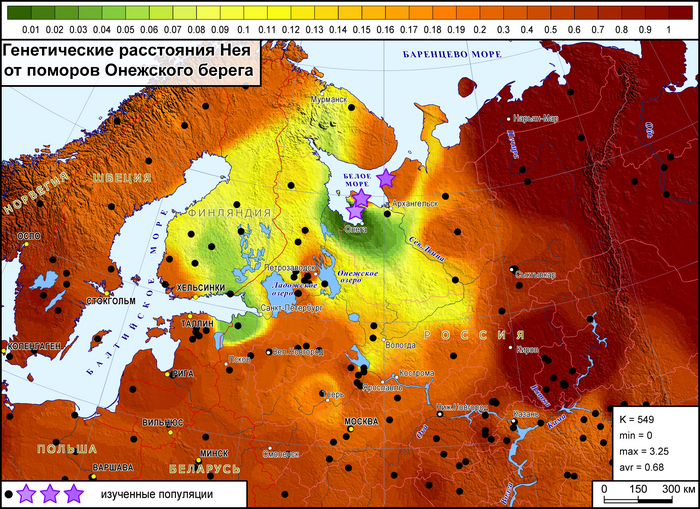
Генофонд поморов Онежского берега на карте генетических расстояний демонстрирует близость к популяциям на обширных географических участках, охватывая почти всю Финляндию на западе, кроме крайних северных и южных участков, а также от рек Сухона и Северная Двина на востоке России до Ярославской и Ленинградской областей на юге и юго-западе.
Эта область сходств охватывает финноязычные народы и те русские популяции, в генофонде которых прослеживается существенный вклад дославянского населения.
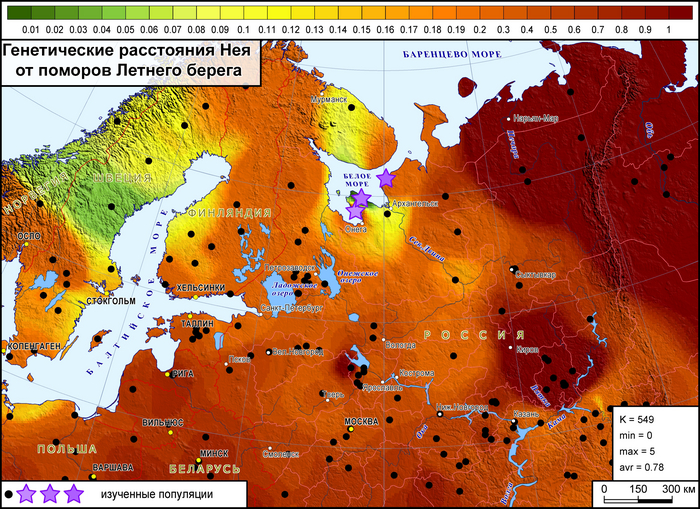
Генофонд поморов Летнего берега демонстрирует наибольшее сходство со шведами и норвежцами, а также с саамами, но слабее.
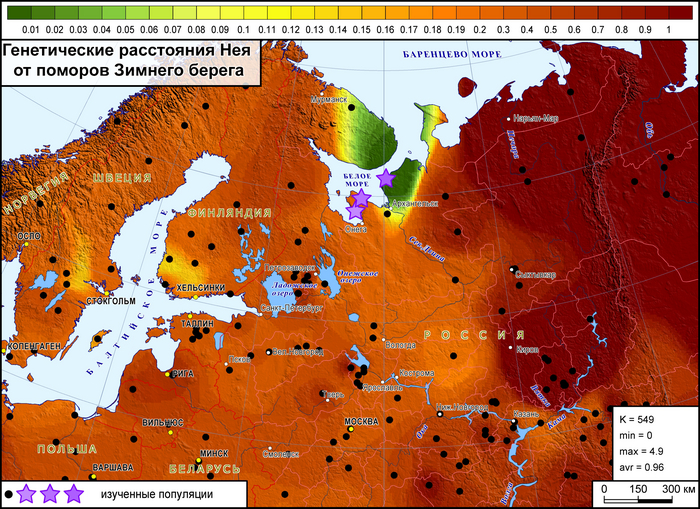
Генофонд поморов Зимнего берега, представленный «прионежской» его частью, показывает лишь некоторое сходство с отдельными популяциями финнов и шведов.
Это единственная популяция поморов, для которой можно допустить, что ее генетический портрет в значительной степени сформирован дрейфом генов, однако исследование популяций остальной части Зимнего берега ещё проводится.
Обсуждение и выводы
Учитывая схожий быт и культуру, а также географическую близость трёх исследованных популяций поморов разумно было бы предположить, что и их генофонды будут довольно схожи. Однако эти ожидания не подтвердились, и каждая из трех исследованных поморских популяций продемонстрировала свои уникальные отличия.
Поморы Онежского берега оказались близки к финноязычным народам России и Финляндии, а также к русским популяциям Архангельской области. Поморы Летнего берега продемонстрировали генетическое сходство только с населением Скандинавии. А у поморов Зимнего берега практически нет сходных генофондов за исключением некоторой близости к финнам и шведам.
Различия между популяциями поморов почти столь же велики, как между рассмотренными популяциями, говорящими на разных языках.
Связать такое уникальное своеобразие генофондов поморов с действием лишь дрейфа генов невозможно, хотя не оспаривается, что дрейф генов был важным фактором в генетической истории всех популяций поморов, численность которых в последнее время сокращалась.
Полногеномные исследования в будущем позволят пролить свет на происхождение поморов и на миграционные волны, оставившие след в их генофонде.
Кстати, обращаю Ваше внимание на необходимость дальнейших исследований, чтобы удержаться от поспешных выводов только по однородительским маркерам.
Но и результаты анализа отцовских линий, учитывая патрилокальность поморов прямо указывают, что причина генетического своеобразия популяций онежских поморов заключается в различном генетическом происхождении их отличающихся субстратов, с последующим дрейфом генов, который размывал генетическую память.
Ещё один важный вопрос заключался в поиске сходства генофондов поморов и новгородцев. Но среднее генетическое расстояние между этими популяциями оказалось довольно значительным, почти как между поморами и другими рассмотренными русскими популяциями. Примечательно, что в предыдущей работе авторы сообщали о различии аутосомных геномов новгородцев и русских севера Архангельской области. И в новой работе показаны различия между генофондами новгородцев и поморов по маркерам Y-хромосомы. Оба результата противоречат мнению о том, что генофонд Русского Севера был сформирован за счёт Новгородской колонизацией северных земель.
Здесь важно отметить, что генетические данные не перечёркивают связи поморов с новгородцами - просто культура и генетика порой отражают разные плоскости взаимодействий между разными группами населения. И генетическая память отражает лишь те события в истории народов, которые сопровождались демографическими изменениями.
Источник:
Окованцев, В. С., Пономарев, Г. Ю., Агджоян, А. Т., Агджоян, А. Т., Пылёв, В. Ю., Балановская, Е. В. Своеобразие поморов Онежского полуострова и Зимнего берега в генетическом контексте севера Европы // Вестник РГМУ. 2022. №5. С. –. DOI: 10.24075/vrgmu.2022.046
Дополнительные источники:
1. Генофонд Русского Севера: Славяне? Финны? Палеоевропейцы? Балановская, Д.В. Пежемский, А.Г. Романов, Е.Е. Баранова, М.В. Ромашкина, А.Т Агджоян, А.Г. Балаганский, И.В. Евсеева, Р. Виллемс, О.П. Балановский 2. Two Sources of the Russian Patrilineal Heritage in Their Eurasian Context Oleg Balanovsky, * Siiri Rootsi, Andrey Pshenichnov, Toomas Kivisild, Michail Churnosov, Irina Evseeva, Elvira Pocheshkhova, Margarita Boldyreva, Nikolay Yankovsky, Elena Balanovska, and Richard Villems DOI 10.1016/j.ajhg.2007.09.019. 2008 by The American Society of Human Genetics. 3. Своеобразие Новгородского генофонда в контексте народонаселения европейской части России. Е.В. Балановская, Д.К. Черневский, О.П. Балановский doi.org/10.34680/2076-8052.2021.3(124).51-57 4. The Eurasian Heartland: A continental perspective on Y-chromosome diversity R. Spencer Wells, Nadira Yuldasheva, Ruslan Ruzibakiev, Walter F. Bodmer et. all doi.org/10.1073/pnas.171305098 5. Khrunin AV, Khokhrin DV, Filippova IN, Esko T, Nelis M, Bebyakova NA, et al. (2013) A Genome-Wide Analysis of Populations from European Russia Reveals a New Pole of Genetic Diversity in Northern Europe. PLoS ONE 8(3): e58552. doi.org/10.1371/journal.pone.0058552
Мое увлечение генеалогией началось около 10-ти лет назад.
Спасибо маме, которая расспрашивала родственников, все записывала и накидала примерную схему древа. Она дала мне хорошую базу, которую я переписал, оформил все в графическом редакторе и забросил)))
Второе дыхание открылось когда я сдал генетический тест.
В первую очередь мне хотелось узнать свой этнический состав. Результат на картинке.
Как видите, результат довольно "скучный", никакого разнообразия.
Оказалось, что при помощи ДНК-теста можно найти родственников, а также построить генеалогическое древо.
Это толкнуло меня на новые поиски и вот уже некоторые веточки протянулись до 1791 года.
Но некоторые так и не могут продлиться далее прабабушки, о чем я расскажу далее.
Их есть у нас! Красивая карта, целых три уровня и много жителей, которых надо осчастливить быстрым интернетом. Для этого придется немножко подумать, но оно того стоит: ведь тем, кто дойдет до конца, выдадим красивую награду в профиль!